Dieback of Euonymus alatus (Celastraceae) Caused by Cytospora haidianensis sp. nov. in China


The Key Laboratory for Silviculture and Conservation of Ministry of Education, Beijing Forestry University, Beijing 100083, China
*
Author to whom correspondence should be addressed.
Forests 2020, 11(5), 524; https://0-doi-org.brum.beds.ac.uk/10.3390/f11050524
Submission received: 26 March 2020
/
Revised: 1 May 2020
/
Accepted: 3 May 2020
/
Published: 7 May 2020
(This article belongs to the Special Issue Emerging Pathogens in Forest Ecosystems)
Abstract
:Euonymus alatus (Celastraceae) is widely cultivated in China for its economic value and landscape benefits. Euonymus alatus dieback occurs due to members of Cytospora and has become one of the most severe diseases affecting its cultivation in China. In this study, we examined the causal agent of bough dieback on campuses of University Road, Beijing, China. Among the strains, three were morphologically consistent with Cytospora, showing hyaline and allantoid conidia. Based on phylogenetic analyses of the concatenated actin (ACT), internal transcribed spacer (ITS), RNA polymerase II second largest subunit (RPB2), translation elongation factor 1-alpha (TEF1-α) and beta-tubulin (TUB2) gene sequences, along with morphological and physiological features, we propose C. haidianensis as a novel species. It was confirmed as a causal agent of dieback of E. alatus by pathogenicity tests. Mycelial growth of Cytospora haidianensis occurred at pH values ranging from 3.0 to 11.0, with optimum growth at 8.3, and at temperatures from 5 to 35 °C, with optimum growth at 19.8 °C. We also tested the growth of C. haidianensis in the presence of six carbon sources. Sucrose, maltose and glucose were highly efficient and xylose was the least. The ability of C. haidianensis to grow at 19.8 °C may help to explain its occurrence causing dieback of E. alatus in Beijing during the autumn season.
1. Introduction
Euonymus alatus (Celastraceae) has been widely cultivated for ornamental landscaping in China because of its tolerance to many environmental conditions [1]. At present, the related research on the fungal diseases of Euonymus is mainly on anthracnose caused by Colletotrichum gloeosporioides, powdery mildew by Oidium euonymi-japonici and dieback by Cytospora euonymicola and C. euonymina [2,3].
The genus Cytospora has wide distribution and has often been regarded as comprising phytopathogens, endophytes or saprobes occurring on a broad range of hosts [3,4]. Several species have been reported as pathogens causing severe branch or trunk dieback disease on monocotyledonous, dicotyledonous and gymnosperm hosts (e.g., Anacardiaceae, Elaeagnaceae, Fabaceae, Juglandaceae, Myrtaceae, Rosaceae, Salicaceae and Ulmaceae) [5,6]. The symptoms of Cytospora canker are elongate, slightly sunken and discoloured areas in the bark at first, then the forming of several prominent black fruit bodies [5]. Conidia emerge from the fructifications in the form of yellow to orange or red gelatinous tendrils under moist conditions [3]. Cytospora species have single or multiple locules (and/or diaporthalean-like perithecia), filamentous conidiophores (and/or clavate to elongate obovoid asci) and allantoid hyaline conidia (and/or ascospores) [5]. As plant pathogens, Cytospora species have also been reported to be associated with other diseases, such as root rot of Chinese jujube and collar rot of pomegranate [7,8].
In the past, it was difficult to name Cytospora species because of their morphological overlap, causing confusion in species delimitation. Previously, identification of Cytospora species was mainly based on host affiliations, often with unclear morphological descriptions. Since the advent of molecular analysis, morphology and phylogeny using internal transcribed spacer (ITS) sequence data were combined to describe 28 species of Cytospora from Eucalyptus, of which 11 species were new to science [5]. Later, similar methods were used to describe 14 species from South Africa [6]. However, only ITS sequences are available for most known Cytospora species, ex-type sequence data are available for only a few species and many taxa need epitypification. Thus, recent studies have subsequently emphasized only part of Cytospora species using a polyphasic approach to solve the confusion in species recognition [3,4,7,8].
Stem and branch dieback have occurred on Euonymus alatus growing on the streets of campuses of University Road in Beijing, China. Typical symptoms of the disease are stem blight and dieback, with lesions extending along the entire branch. Infected stems have light brown to brown pigmentation. According to our observation, the disease seriously affects the colour of Euonymus plants and growth status, along with significant damage to the landscape. The aim of this study was to identify the causal agent causing E. alatus dieback disease based on molecular, morphological and physiological data.
2. Materials and Methods
2.1. Collection and Isolation
Three diseased branches of E. alatus were selected from 20 infected plants observed during collecting trips on the campuses of University Road in Beijing, China. Part of the hymenium containing 3 to 4 fruiting bodies of fresh material was cut horizontally with a sterile blade and crushed in a drop of sterile water on a glass slide. The contents were agitated with the blade until a spore suspension was obtained. Half of the spore suspension was then spread over the surface of 1.8% potato dextrose agar (PDA) in a petri dish and incubated at 25 °C for up to 24 h, and a single germinating conidium was transferred to a fresh PDA plate. Specimens were deposited at the working collection of X.L. Fan (CF) housed at Beijing Forestry University (BJFC) and living cultures were deposited at the China Forestry Culture Collection Centre (CFCC).
2.2. Morphological Observation
Specimens were observed on infected plant tissues, and the structure and size of fruiting bodies, the presence or absence of a conceptacle, and the size and shape of the spores were recorded. Macro-morphological photographs were captured using a Leica stereomicroscope (M205), including size of conidiomata; the presence or absence of special structures such as conceptacle and central column; number and diameter of ostioles per ectostromatic disc; colour, shape and size of discs; and number of locules. Micro-morphological observations such as size and shape of conidiophores and conidia were determined under a Nikon Eclipse 80i microscope equipped with a Nikon digital sight DS-Ri2 high-definition colour camera with differential interference contrast (DIC). Over 30 conidiomata were sectioned and 50 conidia were selected randomly to measure their lengths and widths. Colony diameters were measured, and the colony colours were described after 3 and 14 days according to the colour charts of Rayner [9]. Adobe Bridge CS6 and Adobe Photoshop CS6 were used for manual editing.
2.3. DNA Extraction, PCR Amplification and Sequencing
Mycelium for DNA extraction was grown on PDA with cellophane for 3 days and obtained from the surface of the cellophane by scraping. Genomic DNA was extracted using the modified CTAB method [10]. DNA concentrations were estimated visually by electrophoresis in 1% agarose gel by comparing band intensity with a DNA marker 1 kbp (Takara Bio USA, Inc., Mountain View, CA, USA). PCR amplifications were performed in a DNA Engine (PTC-200) Peltier Thermal Cycler (Bio-Rad Laboratories, Hercules, CA, USA). DNA was amplified from actin (ACT), internal transcribed spacer (ITS), RNA polymerase II second largest subunit (RPB2), translation elongation factor 1-alpha (TEF1-α) and beta-tubulin (TUB2) following Fan et al. [3]. The ACT region was amplified using primers ACT-512F and ACT-1567R [11]. The ITS rDNA region was amplified and sequenced with primers ITS-1 and ITS-4 [12]. The RPB2 was amplified with primers RPB2-5F and fRPB2-7cR [13]. The TEF1-α was amplified with primers EF1-688F and EF1-986R [11,14]. The TUB2 was amplified with primers Bt-2a and Bt-2b [15]. The PCR amplification products were electrophoresed and visualized in gels. The DNA sequencing was performed using an ABI PRISM® 3730XL DNA Analyzer with BigDye® Terminator Kit v.3.1 (Invitrogen) at the Shanghai Invitrogen Biological Technology Company Limited (Beijing, China). DNA sequences generated by each primer combination were used to obtain consensus sequences using Seqman v.7.1 and the DNASTAR Lasergene Core Suite software package (DNASTAR Inc., Madison, WI, USA).
2.4. Phylogenetic Analysis
The current isolates were initially identified as Cytospora sp. based on morphological observations and BLAST results. To clarify their further phylogenetic position, an analysis based on the 5 combined genes (ACT, ITS, RPB2, TEF1-α and TUB2) was constructed to compare Cytospora species from the current study with other strains in the GenBank database. Diaporthe vaccinii CBS 160.32 was selected as the outgroup in all analyses. Subsequent alignments for each gene were generated using MAFFT v.7 [16] and manually adjusted using MEGA v.6 [17]. Ambiguously aligned sequences were excluded from the analysis. Reference sequences were selected based on ex-type or ex-epitype sequences available from recently published literature [5,7,18,19,20,21,22,23,24] (Table 1).
Phylogenetic analyses were formed by PAUP v.4.0b10 for the maximum parsimony (MP) method [25], MrBayes v.3.1.2 for the Bayesian inference (BI) method [26] and RAxML v.7.2.8 for the maximum likelihood (ML) method [27]. Tree length (TL), consistency index (CI), retention index (RI) and rescaled consistency (RC) were calculated [25]. ML analysis was generated using a GTR+G+I model of site substitution following recent study [4], including estimation of gamma distributed rate heterogeneity and proportion of invariant sites [27]. Branch support was evaluated with a bootstrapping method of 1000 replicates [28]. BI analysis was performed using a Markov chain Monte Carlo (MCMC) algorithm with Bayesian posterior probabilities [29]. A nucleotide substitution model was estimated by MrModeltest v.2.3 [30] and a weighted Bayesian analysis was considered. Two MCMC chains were run from random trees for 10,000,000 generations and trees were sampled each 100th generation. The first 40% of trees were discarded as the burn-in phase of each analysis and the Bayesian posterior probability (BPP) was calculated to assess the remaining trees [29]. The branch support from MP and ML analysis was evaluated with a bootstrapping (BS) method of 1000 replicates [28]. Phylograms were constructed using Figtree v.1.3.1 [31]. Sequence data were deposited in GenBank. The aligned matrices used for phylogenetic analysis were submitted through TreeBASE (www.treebase.org; study ID S26000).
2.5. Pathogenicity Test
Three Cytospora strains (CFCC 54184, CFCC 54056 and CFCC 54057) obtained in this study were used to conduct the pathogenicity test. The pathogenicity test was performed on 1-year-old E. alatus plants obtained from seeds kept in a greenhouse at constant 28 °C and 99% relative humidity. On healthy plants, twigs to be used for inoculation were surface disinfected with 75% ethanol for 1 min. The bark surface of each disinfected twig was scalded with a sterilized inoculating loop within a region 5 mm in length to a depth of 2 mm. For mycelial inoculation, a 5 mm diameter PDA plug with mycelium was taken from a 3-day-old colony and inoculated onto the wounded twigs. Three replicates were conducted for each isolate. Non-colonized PDA plugs and sterile water were used as negative controls. Pathogenicity was determined by the length of the necrotic lesion caused by the tested isolates, which was measured 3 weeks after inoculation. Fungal isolates were re-isolated from the infected tissue, and morphological characterization and DNA sequence comparisons were conducted to follow Koch’s postulates.
2.6. Temperature and pH Tests
The 3 Cytospora isolates showed similar growth characteristics, so we used the type strain of the new species (CFCC 54057) to evaluate the effects of temperature and pH on colony growth using PDA plates. Tested temperatures ranged from 0 to 40 °C at intervals of 5 °C (i.e., 0, 5, 10, 15, 20, 25, 30, 35 and 40 °C). In order to clarify the effect of pH on radial mycelial growth, PDA medium was adjusted with 0.1 M NaOH and 0.1 M HCl to obtain pH values from 2.0 to 12.0 at intervals of 1.0 (i.e., 2.0, 3.0, 4.0, 5.0, 6.0, 7.0, 8.0, 9.0, 10.0, 11.0 and 12.0). A 5 mm diameter mycelial plug was placed in the centre of a 90 mm petri dish with PDA medium and incubated at 28 °C in the dark, with 3 replicates for each treatment. The effects of pH and temperature on mycelial growth were determined by measuring the colony diameter after 24, 48, 72 and 96 h of incubation and the data were converted to radial growth in millimetres [32]. Data were analysed in IBM SPSS Statistics v.22.0 (IBM Inc., Armonk, NY, USA) to select the model that best fit the individual data points, and SPSS was used to confirm the selected model. The optimal temperature and pH value of the regression curves were calculated based on the regression equations generated by IBM SPSS Statistics, and output figures with Origin v.8.0.
2.7. Carbon Colony Growth Test
To investigate the utilization of carbon sources, the type strain of the new species (CFCC 54057) was incubated in the dark at 28 °C on PDA medium for 4 days. PDA medium was used as the base medium (potato 20 g, sucrose 20 g, agar 17 g, distilled water to complete 1000 mL). The 20 g of sucrose was replaced by 20 g of fructose, galactose, glucose, maltose, sucrose or xylose to test these compounds as carbon sources. A 5 mm diameter PDA plug of mycelium was transferred to the centre of each sole carbon source medium. Colony growth was determined by measuring the colony diameters after incubation for 24, 48, 72 and 96 h at 28 °C in the dark, and the results were subsequently converted to radial growth [32]. Mean comparisons were conducted using Tukey’s honestly significant difference (HSD) test (α = 0.05) in SigmaPlot v.14.0.
3. Results
3.1. Phylogenetic Analyses
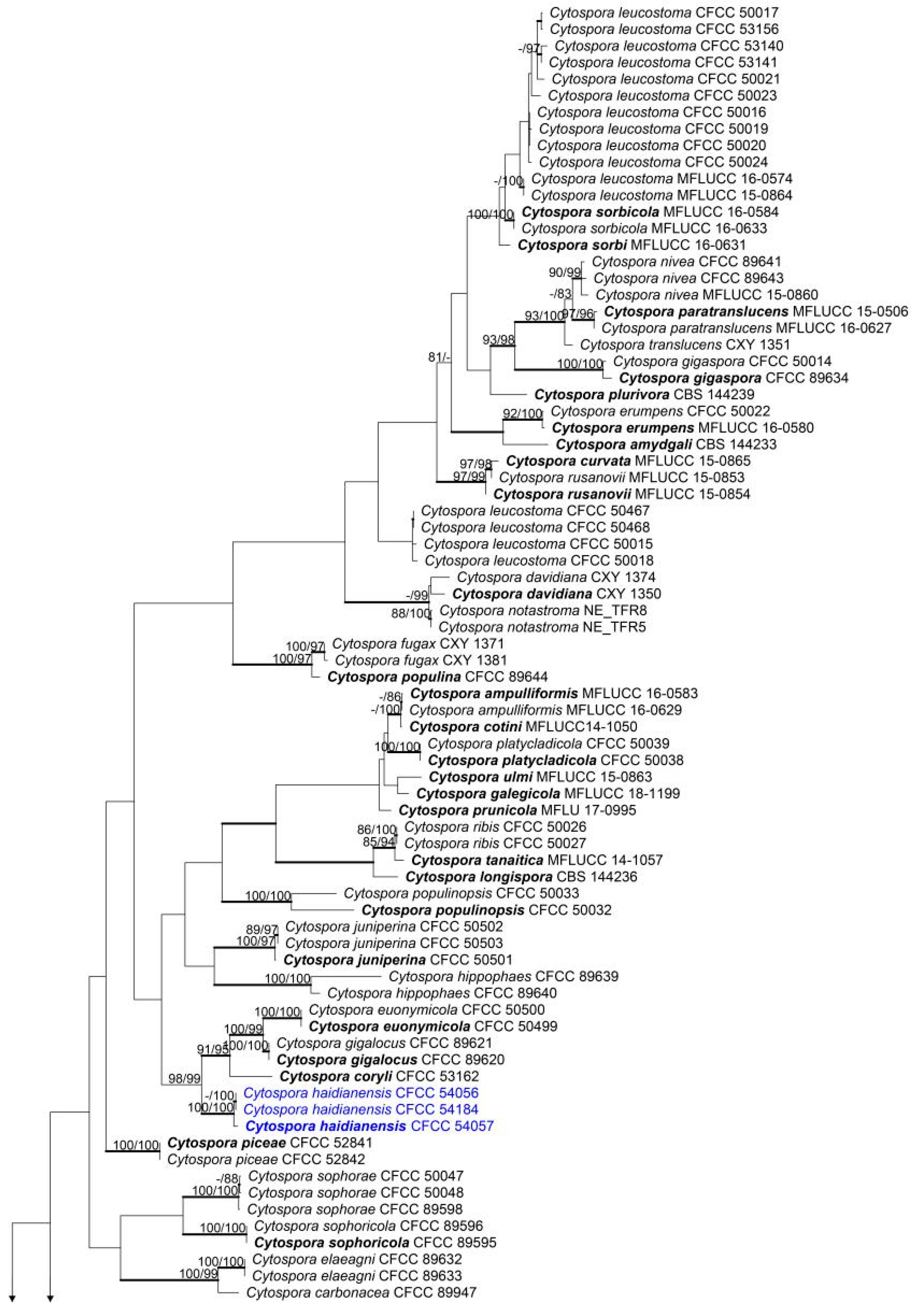
A combined matrix of five gene sequences of Cytospora species was constructed. The combined alignment matrices (ACT, ITS, RPB2, TEF1-α and TUB2) included 192 accessions (3 from this study and 189 retrieved from GenBank) and counted 3056 characters including gaps (350 characters for ACT, 631 for ITS, 726 for RPB2, 725 for TEF1-α and 624 for TUB2), of whih 1594 characters were constant, 130 variable characters were parsimony-uninformative and 1349 (44.14%) characters were variable and parsimony-informative. The MP analysis generated 200 parsimonious trees, the first of which is presented in Figure 1 (TL = 8,573, CI = 0.312, RI = 0.788, RC = 0.246). The tree topologies of ML and BI analyses were similar to that of the MP tree.
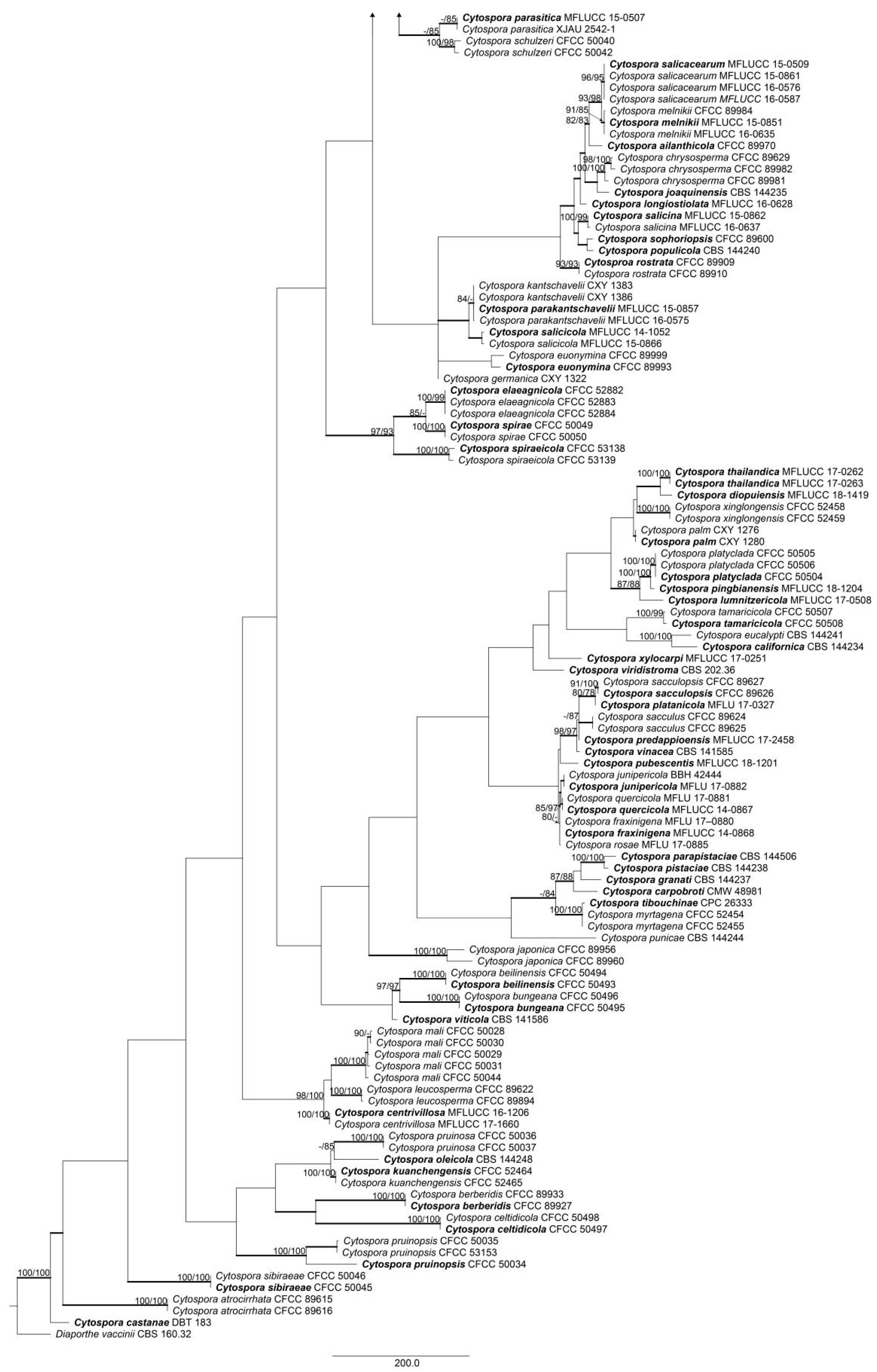
Based on the initial analysis, a second, more inclusive combined matrix was constructed using 27 accessions from the first dataset. The second combined alignment matrix counted 2531 characters including gaps (274 characters for ACT, 529 for ITS, 726 for RPB2, 553 for TEF1-α and 449 for TUB2). In total, 1,819 characters were constant, 182 variable characters were parsimony-uninformative and 547 (21.61%) characters were variable and parsimony-informative. The MP analysis generated one parsimonious tree and the best tree (TL = 1,225, CI = 0.768, RI = 0.853, RC = 0.656) is presented in Figure 2. The tree topologies of ML and BI analyses were similar to that of the MP tree.
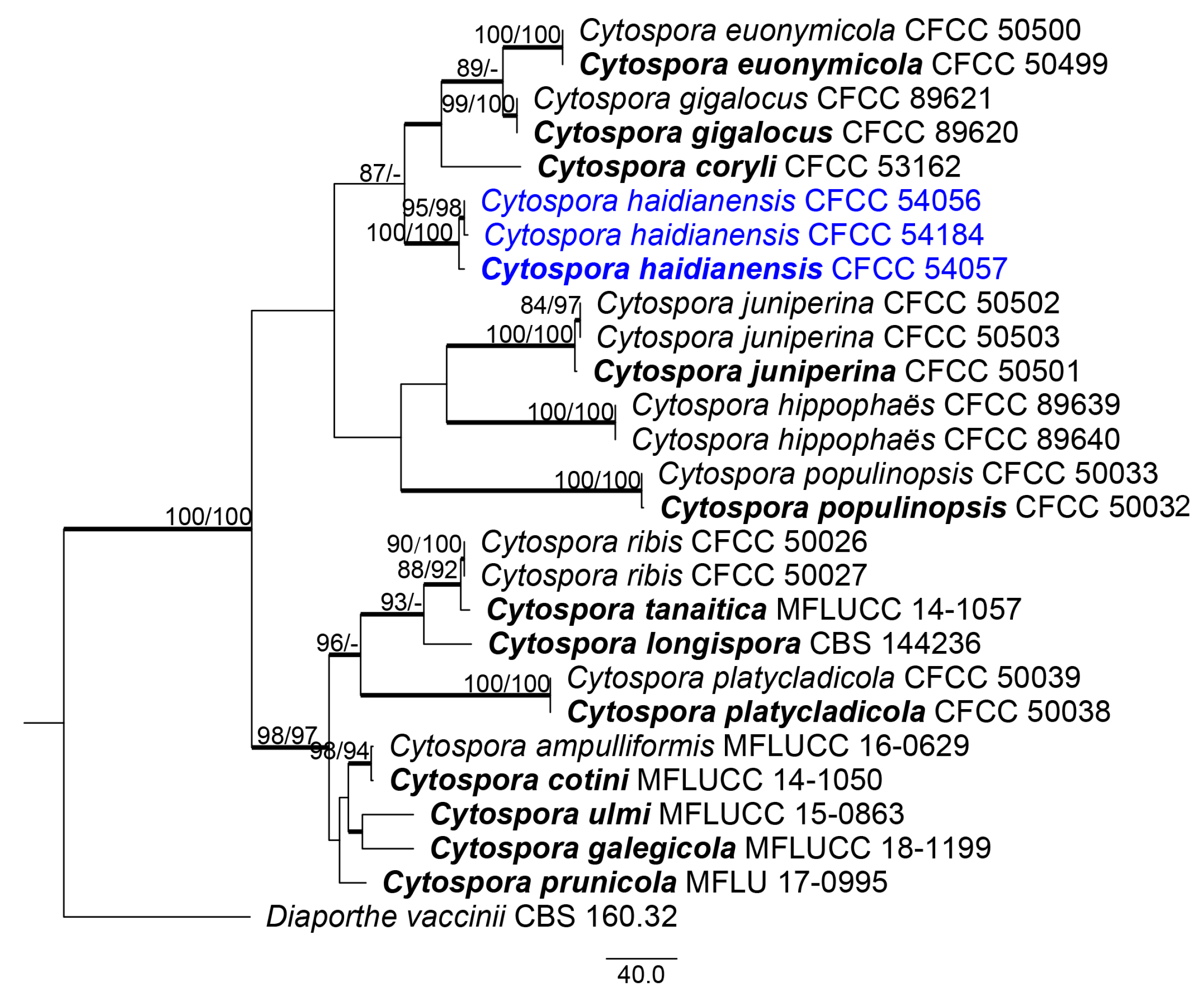
Based on the multilocus phylogeny and morphology, all three strains were assigned to one new species, named Cytospora haidianensis, representing a monophyletic clade with high support value (MP/ML/BI = 100/100/1).
3.2. Taxonomy
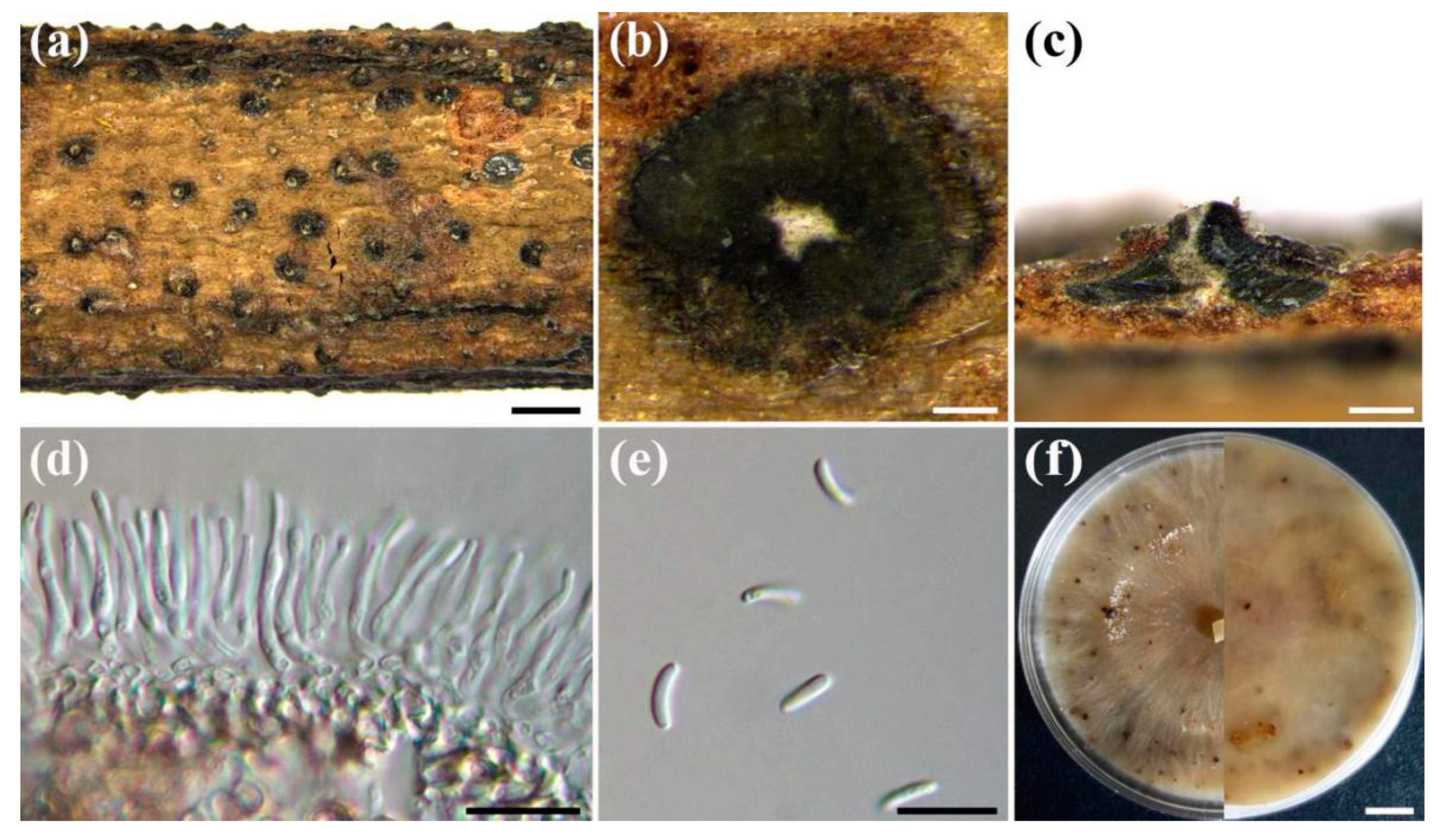
Cytospora haidianensis X. Zhou & X.L. Fan, sp. nov. (Figure 3)
MycoBank MB 835121
Holotype: CF 20198643
Etymology: named after the place where it was first collected, Haidian
Host/Distribution: on cankered Euonymus alatus branches in China
Description: Sexual morph not observed. Pycnidial stromata ostiolate, immersed in bark, scattered, erumpent through the surface, with multiple locules. Conceptacle absent. Ectostromatic disc isabelline to dark brick, conspicuous, circular to ovoid, (330–)380–500(–520) µm (= 460 ± 30 µm, n = 35) diam, with one ostiole per disc. Ostiole in the centre of the disc, black, conspicuous, (170–)179–195(–200) μm (= 188 ± 3 µm, n = 10) diam. Numerous locules, subdivided frequently by invaginations with common walls, (650–)700–800(–1000) µm (= 760 ± 30 µm, n = 30) diam. Conidiophores hyaline, branched at the base or unbranched, thin-walled, (9–)12–15(–16.5) × 1.0–1.5 μm (= 13.5 ± 1.5 × 1.4 ± 0.1 µm, n = 50), embedded in a gelatinous layer. Conidiogenous cells enteroblastic, phialidic, subcylindrical to cylindrical, (8.5–)9–12.5(–13.5) × 1–1.5 μm (= 11 ± 1.5 μm, n = 30), tapering towards the apices. Conidia hyaline, allantoid, smooth, aseptate, thin-walled, (6–)6.5–7.5 × 1–1.5 μm (= 6.8 ± 0.2 × 1.2 ± 0.1 µm, n = 50).
Cultural characteristics: Colonies on PDA are initially white after 3 days, becoming light brown after 14 days. The colonies are thin with a uniform texture, lack aerial mycelium and grow up to 90 mm after 4 days. Pycnidia were randomly observed on the surface of the colony.
Material examined: CHINA, Beijing, Haidian, University Road, 116°20′19.11″ E, 40°00′16.21″ N, 51 m asl, on stems and branches of Euonymus alatus, Xinlei Fan, 12 November 2019 (CF 20198643, holotype; ex-type culture, CFCC 54057). Beijing, Haidian, University Road, 116°35′49.37″ E, 40°00′37.85″ N, 50 m asl, on stems and branches of Euonymus alatus, Xinlei Fan, 12 November 2019 (CF 20198644; living culture, CFCC 54056). Beijing, Haidian, University Road, 116°20′19.11″ E, 40°00′16.21″ N, 51 m asl, on stems and branches of Euonymus alatus, Xinlei Fan, 12 November 2019 (CF 20198646; living culture, CFCC 54184).
Notes: Cytospora haidianensis differs from the phylogenetically related species C. euonymicola and C. gigalocus based on the sizes of the ectostromatic disc (240–350 µm diam in C. euonymicola and 330–620 µm diam in C. gigalocus), ostiole (60–120 µm diam in C. euonymicola and 130–190 µm diam in C. gigalocus), locules (1150–1400 µm diam in C. euonymicola and 1630–2180 µm diam in C. gigalocus), conidiophores (13–21.5 × 1.5–2 µm in C. euonymicola and 16.1–23.6 µm in C. gigalocus) and conidia (4.5–5 × 1 µm in C. euonymicola and 4.6–5.6 × 0.8–1.3 µm in C. gigalocus) [3,21]. Fan et al. [21] typified C. gigalocus based on material collected on the stems of Juglans regia, C. euonymicola and C. euonymina first found on twigs and branches of Euonymus kiautschovicus in China [3]. Similar to the other species, C. haidianensis also differs from the recently described species, C. coryli, based on macro- and micro-morphological characteristics [4]. At the molecular level, C. haidianensis differs from C. euonymicola by ACT (45/350), ITS (35/631), RPB2 (24/726), TEF1-α (47/725) and TUB2 (24/624), and differs from C. gigalocus by ACT (62/350), ITS (32/631), RPB2 (17/726), TEF1-α (41/725) and TUB2 (22/624).
Based on a BLAST search of the NCBI GenBank nucleotide database, the closest hits using the ACT sequence had distant hits with Cytospora gigalocus (strain CFCC 89620; GenBank KU710997; identities = 236/249 (94.78%), 3 gaps (1%)); Cytospora carbonacea (strain CFCC 50055; GenBank KP310838; identities = 237/252 (94.44%), 7 gaps (1%)). The closest hits using the ITS sequence had distant hits with Cytospora populina (strain CFCC 89644; GenBank KR045640; identities = 499/522 (95.59%), 10 gaps (1%)); Cytospora cenisia (strain CPC 28396; GenBank KY051983; identities = 489/521 (95.59%), 9 gaps (1%)). The closest hits using the RPB2 sequence had the highest similarity to Cytospora gigalocus (strain CFCC 89620; GenBank KU710957; identities = 690/711 (97.05%), 0 gaps (0%)); Cytospora hippophaes (strain CFCC 89637; GenBank KF765711; identities = 686/711 (96.48%), 0 gaps (0%)). The closest hits using the TEF1-α sequence had distant hits with Cytospora coryli (strain CFCC 53162; GenBank MN850758; identities = 397/423 (93.85%), 3 gaps (0%)); Cytospora piceae (strain CFCC 52842; GenBank MH820403; identities = 385/420 (91.67%), 12 gaps (2%)). The closest hits using the TUB2 sequence had distant hits with Cytospora gigalocus (strain CFCC 89620; GenBank KR045669; identities = 400/420 (95.24%), 11 gaps (2%)); Cytospora leucostoma (strain CFCC 53140; GenBank MN861115; identities = 395/419 (94.27%), 10 gaps (2%)).
3.3. Pathogenicity Test
The three Cytospora haidianensis strains (CFCC 54184, CFCC 54056 and CFCC 54057) tested in this study were pathogenic on the Euonymus alatus twigs. No symptoms were observed in the non-inoculated controls. Brown lesions appeared at the inoculated points after 7 days of inoculation. The diseased spots turned brown and lesion areas were up to 16 mm long at 14 days after inoculation. By the third week after inoculation, the length of the brown necrotic lesions ranged from 36 to 45 mm (Figure 4). Koch’s postulates were performed by successful re-isolation of fungal strains from all necrotic twigs inoculated with Cytospora haidianensis. The morphology and DNA sequences of the isolates retrieved from the inoculated twigs were consistent with those of the strains used for inoculation.
3.4. Effects of Temperature and pH on Mycelial Growth
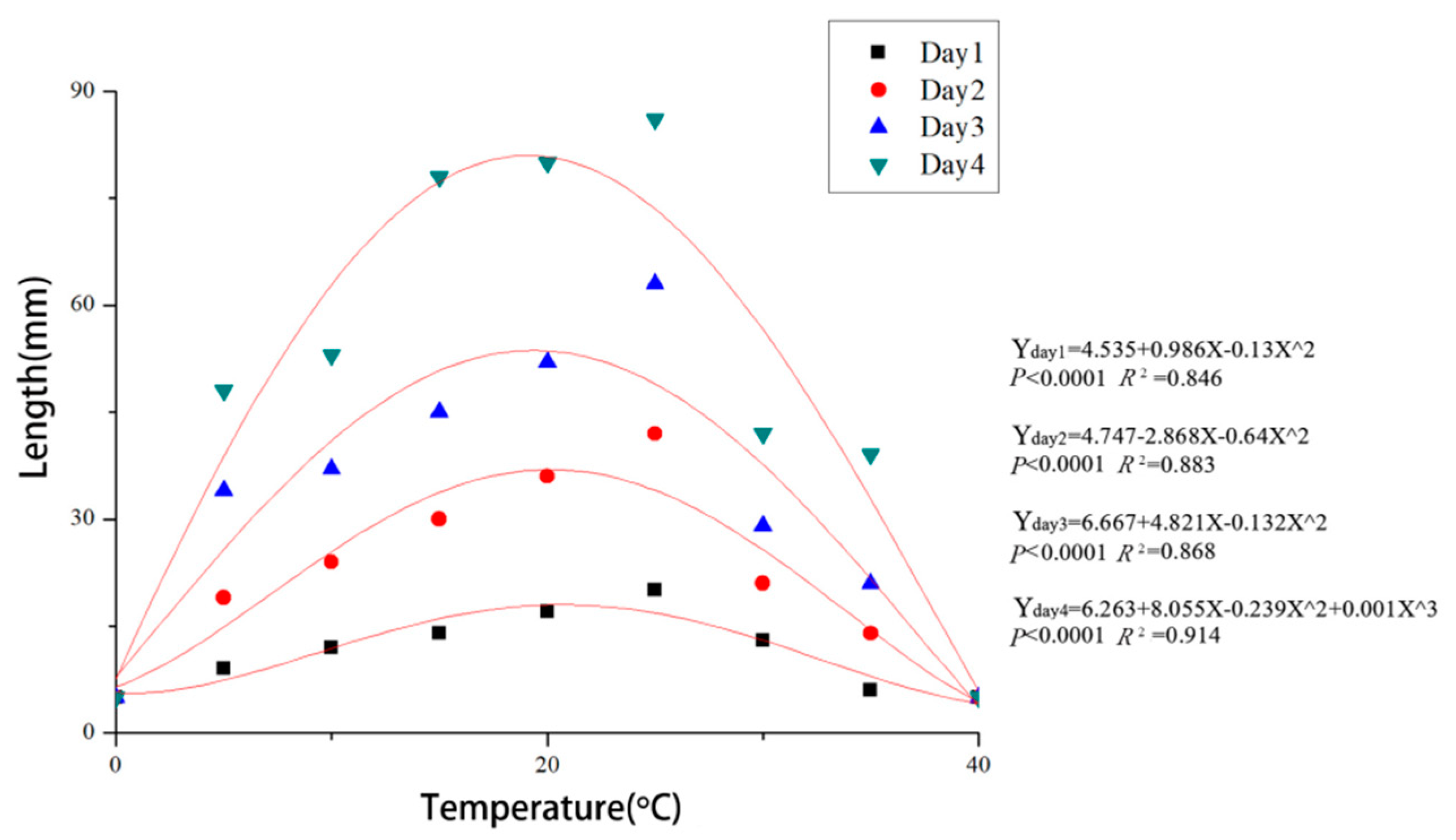
Colonies of C. haidianensis grew on PDA in the temperature range from 5 to 35 °C but not at 0 and 40 °C after 48 h of incubation. The fastest mycelial growth occurred at 19.8 °C, reaching 20 mm after 24 h and 86 mm after 96 h, and the least growth occurred at 5 and 35 °C. The data conform to the regression equations Y = 4.535 + 0.986X − 0.13X² (p < 0.0001, R² = 0.846) at 24 h, Y = 4.747 − 2.868X − 0.64X² (p < 0.0001, R² = 0.883) at 48 h, Y = 6.667 + 4.821X − 0.132X² (p < 0.0001, R² = 0.868) at 72 h and Y = 6.263 + 8.055X − 0.239X² + 0.001X³ (p < 0.0001, R² = 0.914) at 96 h (X = temperature (°C), Y = growth (colony diameter, mm)). Based on the regression analysis, the optimal growth for C. haidianensis after incubation was estimated to occur at 19.8 °C (Figure 5).
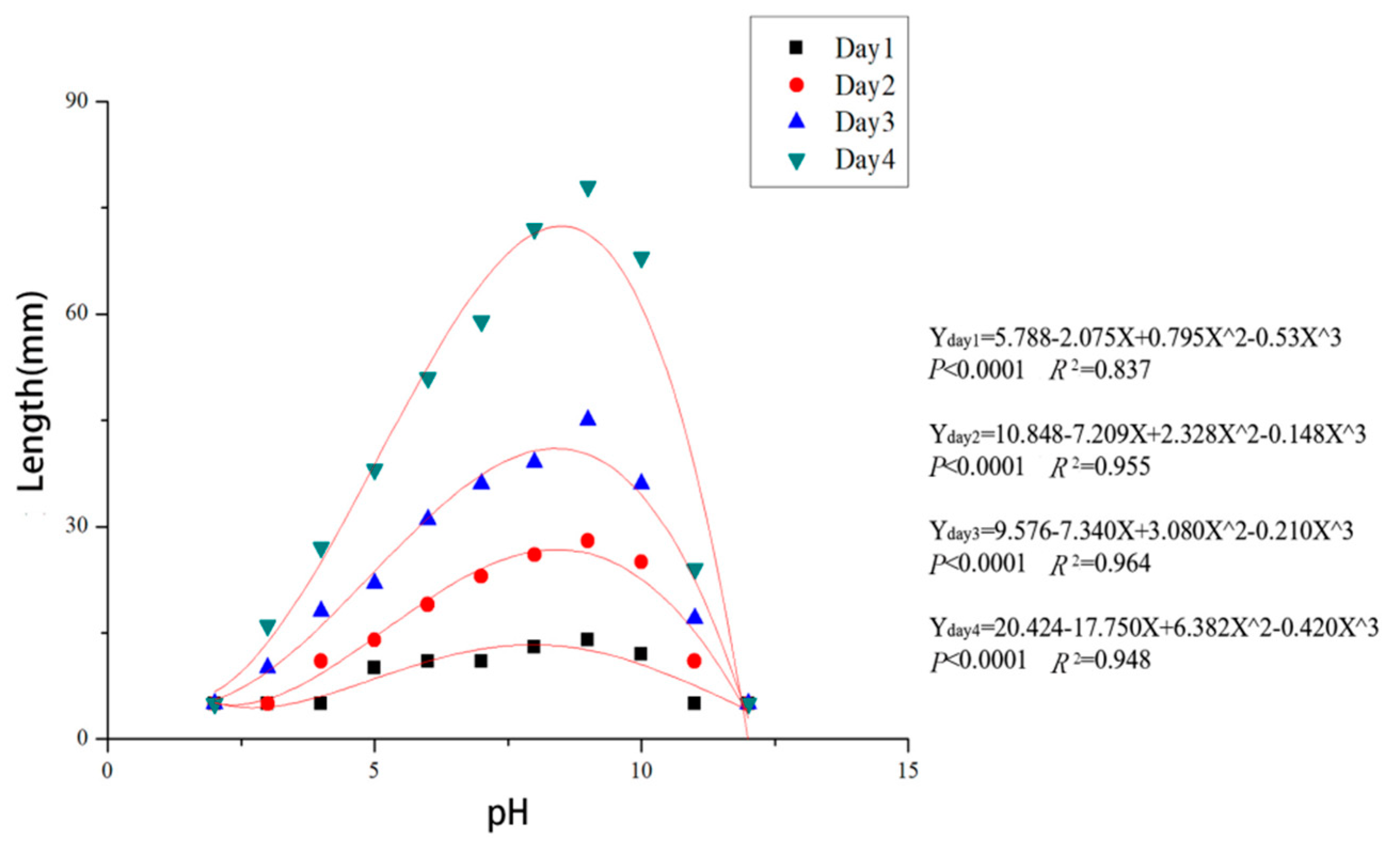
Colonies of C. haidianensis grew on PDA in the pH range from 3.0 to 10.0, but not at pH 2.0 and 12.0. After 48 h, the mycelium of C. haidianensis grew on PDA in the pH range from 3.0 to 10.0, but not at pH 2.0 or 12.0. Mycelium grew most rapidly at pH 9.0 after 24 h, reaching 14 mm, followed by pH 8.0 and 10.0, which gave colony diameters of 13 mm and 12 mm, respectively. The mycelia almost covered the 90 mm dishes after 96 h incubation at pH 8.0 and 9.0, while they grew more slowly at pH 3.0, 4.0, 5.0 and 11.0, attaining colony diameters of no more than 45 mm after 96 h. The data fit the regression equations Y = 5.788 − 2.075X + 0.795X² − 0.53X³ (p < 0.0001, R² = 0.837) at 24 h, Y = 10.848 − 7.209X + 2.328X² − 0.148X³ (p < 0.0001, R² = 0.955) at 48 h, Y = 9.576 − 7.340X + 3.080X² − 0.210X³ (p < 0.0001, R² = 0.964) at 72 h and Y = 20.424 − 17.750X + 6.382X² − 0.420X³ (p < 0.0001, R² = 0.948) at 96 h incubation (X = pH, Y = growth (colony diameter, mm)) (Figure 6). Based on these regression equations, the optimal growth of C. haidianensis after 24 and 48 h incubation was estimated to be at pH 8.3.
3.5. Effects of Carbon Sources on Mycelial Growth
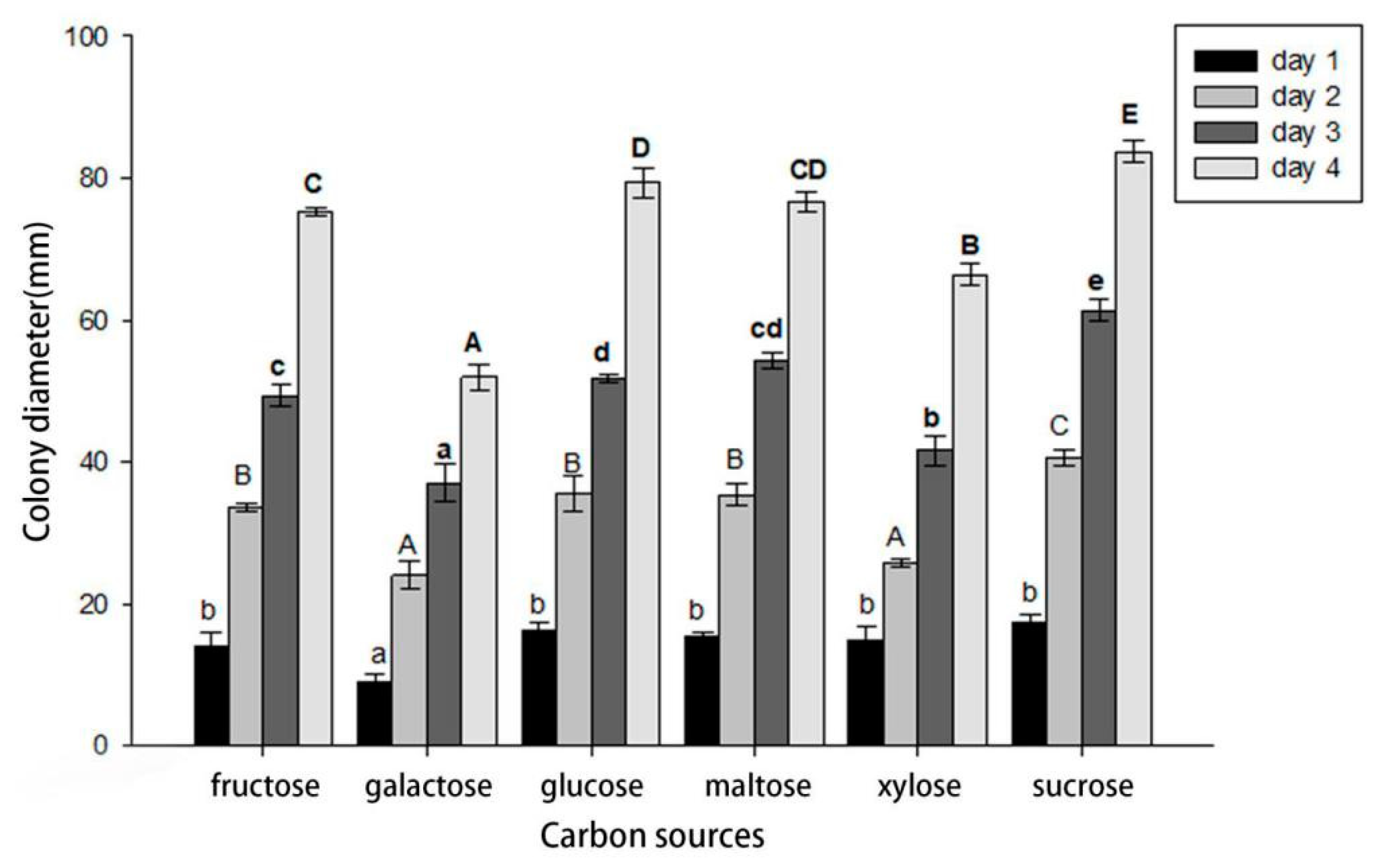
Cytospora haidianensis was able to grow using all six carbon sources tested. After 24 h, the utilization of sucrose was significantly greater than galactose, while there was no difference among fructose, glucose, xylose and maltose, which were slightly less well utilized than the other three carbon sources. The utilization of galactose was significantly lower than that of all other carbon sources tested. However, after 96 h, sucrose utilization was significantly higher than galactose and xylose, while there was no difference between fructose and glucose. Galactose had the lowest level of carbon utilization (Figure 7).
4. Discussion
In the present study, three specimens were collected from symptomatic branches and twigs associated with dieback disease of Euonymus alatus in Beijing, China. A novel fungal species, C. haidianensis, was introduced based on molecular, morphological and physiological data, and confirmed as the causal agent after pathogenic tests.
According to our multilocus phylogenetic analysis, C. haidianensis is a lineage well supported (MP-BS = 100, ML-BS = 100 and BPP = 1.0) and placed in a clade related to C. euonymicola, C. gigalocus and C. coryli (Figure 3). In a recent study, Fan et al. [3] described C. euonymicola and C. euonymina from twigs and branches of Euonymus kiautschovicus in Shaanxi Province, China. Comparing these species with the novel species C. haidianensis, C. euonymicola mainly has small ectostromatic discs (240–350 µm diam) and conidia (4.5–5 × 1 μm) and C. euonymina mainly has small ectostromatic discs (200–230 µm diam) and slightly larger conidia (6.5–7.5 × 1.5–2 μm), but the latter is not phylogenetically related to the new species. Cytospora gigalocus was described by Fan et al. [22] on stems of Juglans regia in Qinghai Province, China, mainly having slightly large ectostromatic discs (330–620 µm diam) and small conidia (4.6–5.6 × 0.8–1.3 µm), differing from C. haidianensis based on these morphological features (see notes for C. haidianensis). Cytospora coryli was recently proposed by Zhu et al. [4] as necrotrophic on branches of Corylus mandshurica in Mount Dongling (China), differing from C. haidianensis based on the size of ectostromatic discs (270–340 μm diam), large locules (1550–1710 μm diam), conidiophores (15.5–18.5 × 1–2 μm), conidiogenous cells (7.5–14 × 1–2 μm) and conidia (5–7 × 1–2 μm), and culture characteristics.
Pathogenicity tests were conducted on 1-year potted E. alatus plants in a greenhouse. The results indicated that C. haidianensis was pathogenic on E. alatus twigs. According to Pan et al. [7], Cytospora species invade the xylem and cause mortality of the whole branch, similar to the results obtained in this study within three weeks, showing the typical stem blight that occurred in the sampled place (Figure 4). The growth temperature for phytopathogenic fungi is generally from 10 to 35 °C, optimally from 20 to 30 °C [33]. For instance, the optimal growth temperature of Penicillium cellarum causing rot in stored sugar beet roots was reported as 22 °C [34]; for Diaporthe neotheicola and D. ambigua causing dieback blueberry in Chile, it was 25 °C; for Diaporthe sp., it was 22 °C [35]; and for Phoma sorghina, which was found to cause twisted leaf disease in sugarcane in China, it was 20–25 °C [36]. The mycelia of C. haidianensis grew from 5 to 35 °C, with an optimal temperature of 19.8 °C (Figure 5).
Most phytopathogenic fungi grow optimally in a pH range between 5 and 6.5 [37]. For Lasiodiplodia vaccinii, the range was 5.0 to 7.0, though it could still grow slowly at pH of 4.0 or 10.0 [33]. Similar results have been reported for L. theobromae, which could grow on media with a pH range from 4.0 to 10.0, with the optimal pH in the range of 5.0 to 7.0 [36]. The optimal pH value for C. haidianensis was from 8.0 to 10.0, though it could still grow slowly at pH of 4.0 or 11.0 (Figure 6). All six carbon sources tested in this study contributed to the growth of C. haidianensis, with less utilization of xylose than all the other carbon sources used (Figure 7).
The dieback in Euonymus alatus caused by C. haidianensis damages the plants. Cytospora haidianensis blights many branches and leaves discolouration, causing gradual death of a large number of E. alatus (Figure 4). This phenomenon is not confined to Beijing; Cytospora euonymicola was also reported as a pathogenic fungus from Euonymus in Shaanxi Province, and Cytospora euonymina was also found in Euonymus in Shanxi Province [3]. A similar phenomenon also happens in other countries; Cytospora euonymi was also associated with the blight of Euonymus twigs in the USA and Europe. Other genera such as Cercospora, Colletotrichum, Coniothyrium and Fusarium were also reported to be pathogenic fungi in Euonymus [38].
To date, C. haidianensis has been found only from Euonymus alatus in Beijing. Management practices, including better ventilation and lighting, might help to alleviate the damage resulting from stem dieback caused by C. haidianensis. The distribution and host spectrum of C. haidianensis need further study.
5. Conclusions
A novel fungal species, Cytospora haidianensis, is an emerging pathogen on Euonymus alatus dieback disease in Beijing, China. The new species is the causal agent for E. alatus by Koch’s postulates that grows best at 19.8 °C, pH 8.3. All the six carbon sources tested support the growth of C. haidianensis with the sucrose utilization is significantly higher than others.
Author Contributions
Experiments: X.Z., M.P. and H.L.; writing—original draft preparation: X.Z.; writing—review and editing: X.F. and C.T. All authors have read and agreed to the published version of the manuscript.
Funding
This study was financed by the Fundamental Research Funds for the Central Universities (2019ZY23) and College Student Research and Career-creation Program of Beijing (X201910022006).
Acknowledgments
We are grateful for the assistance of Xinao Mei (Beijing University of Chemical Technology), Lin Zhao (Beijing Forestry University) and Zhuang An (Shanghai Jiaotong University) during this study.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Dirr, M.A. Winged euonymus is a stalwart shrub for consistent fall color. Nurs. Manag. Prod. 2001, 17, 91–95. [Google Scholar]
- Olive, J.W.; Wick, R.L. Euonymus diseases. In Diseases of Woody Ornamentals and Trees in Nurseries; Jones, R.K., Benson, D.M., Eds.; APS Press: St. Paul, MN, USA, 2001. [Google Scholar]
- Fan, X.L.; Bezerra, J.D.P.; Tian, C.M.; Crous, P.W. Cytospora (Diaporthales) in China. Persoonia 2020, 45, 1–45. [Google Scholar] [CrossRef]
- Zhu, H.Y.; Pan, M.; Bezerra, J.D.P.; Tian, C.M.; Fan, X.L. Discovery of Cytospora species associated with canker disease of tree hosts from Mount Dongling of China. MycoKeys 2020, 68, 97–121. [Google Scholar] [CrossRef]
- Adams, G.C.; Wingfield, M.J.; Common, R.; Roux, J. Phylogenetic relationships and morphology of Cytospora species and related teleomorphs (Ascomycota, Diaporthales, Valsaceae) from Eucalyptus. Stud. Mycol. 2005, 52, 1–144. [Google Scholar]
- Adams, G.C.; Roux, J.; Wingfield, M.J. Cytospora species (Ascomycota, Diaporthales, Valsaceae), introduced and native pathogens of trees in South Africa. Australas. Plant Pathol. 2006, 35, 521–548. [Google Scholar] [CrossRef]
- Pan, M.; Zhu, H.Y.; Tian, C.M.; Alvarez, L.V.; Fan, X.L. Cytospora piceae sp. nov. associated with canker disease of Picea crassifolia in China. Phytotaxa 2018, 383, 181–196. [Google Scholar] [CrossRef]
- Palavouzis, S.C.; Tzamos, S.; Paplomatas, E.; Thomidis, T. First report of Cytospora punicae isolated from pomegranate plants with symptom of collar rot in northern Greece. J. Plant Pathol. 2015, 97, 209–220. [Google Scholar]
- Rayner, R.W. A Mycological Colour Chart; Commonwealth Mycological Institute: Kew, UK, 1970. [Google Scholar]
- Doyle, J.J.; Doyle, J.L. Isolation of plant DNA from fresh tissue. Focus 1990, 12, 13–15. [Google Scholar]
- Carbone, I.; Anderson, J.B.; Kohn, L.M. A method for designing primer sets for the speciation studies in filamentous ascomycetes. Mycologia 1999, 91, 553–556. [Google Scholar] [CrossRef]
- White, T.J.; Bruns, T.; Lee, S.; Taylor, J. Amplification and direct sequencing of fungal ribosomal RNA genes for phylogenetics. PCR Protoc. Guide Methods Appl. 1990, 18, 315–322. [Google Scholar]
- Liu, Y.J.; Whelen, S.; Hall, B.D. Phylogenetic relationships among ascomycetes: Evidence from an RNA polymerse II subunit. Mol. Biol. Evol. 1999, 16, 1799–1808. [Google Scholar] [CrossRef]
- Alves, A.; Crous, P.W.; Correia, A.; Phillips, A.J.L. Morphological and molecular data reveal cryptic speciation in Lasiodiplodia theobromae. Fungal Divers. 2008, 28, 1–13. [Google Scholar]
- Glass, N.L.; Donaldson, G.C. Development of primer sets designed for use with the PCR to amplify conserved genes from filamentous ascomycetes. Appl. Environ. Microbiol. 1995, 61, 1323–1330. [Google Scholar] [CrossRef] [Green Version]
- Katoh, K.; Standley, D.M. MAFFT multiple sequence alignment software version 7: Improvements in performance and usability. Mol. Biol. Evol. 2013, 30, 772–780. [Google Scholar] [CrossRef] [Green Version]
- Tamura, K.; Stecher, G.; Peterson, D.; Filipski, A.; Kumar, S. MEGA6: Molecular evolutionary genetics analysis version 6.0. Mol. Biol. Evol. 2013, 30, 2725–2729. [Google Scholar] [CrossRef] [Green Version]
- Zhu, H.Y.; Tian, C.M.; Fan, X.L. Multigene phylogeny and morphology reveal Cytospora spiraeae sp. nov. (Diaporthales, Ascomycota) in China. Phytotaxa 2018, 338, 49–62. [Google Scholar] [CrossRef]
- Fan, X.L.; Liang, Y.M.; Ma, R.; Tian, C.M. Morphological and phylogenetic studies of Cytospora (Valsaceae, Diaporthales) isolates from Chinese scholar tree, with description of a new species. Mycoscience 2014, 55, 252–259. [Google Scholar] [CrossRef]
- Fan, X.L.; Tian, C.M.; Yang, Q.; Liang, Y.M.; You, C.J.; Zhang, Y.B. Cytospora from Salix in northern China. Mycotaxon 2014, 129, 303–315. [Google Scholar] [CrossRef]
- Fan, X.L.; Hyde, K.D.; Liu, M.; Liang, Y.M.; Tian, C.M. Cytospora species associated with walnut canker disease in China, with description of a new species C. gigalocus. Fungal Biol. 2015, 119, 310–319. [Google Scholar] [CrossRef]
- Fan, X.L.; Hyde, K.D.; Yang, Q.; Liang, Y.M.; Ma, R.; Tian, C.M. Cytospora species associated with canker disease of three anti-desertification plants in northwestern China. Phytotaxa 2015, 197, 227–244. [Google Scholar] [CrossRef] [Green Version]
- Lawrence, D.P.; Holland, L.A.; Nouri, M.T.; Travadon, R.; Abramians, A.; Michailides, T.J.; Trouillas, F.P. Molecular phylogeny of Cytospora species associated with canker diseases of fruit and nut crops in California, with the descriptions of ten new species and one new combination. IMA Fungus 2018, 9, 333–370. [Google Scholar] [CrossRef]
- Lawrence, D.P.; Travadon, R.; Pouzoulet, J.; Rolshausen, P.E.; Wilcox, W.F.; Baumgartner, K. Characterization of Cytospora isolates from wood cankers of declining grapevine in North America, with the descriptions of two new Cytospora species. Plant Pathol. 2017, 66, 713–725. [Google Scholar] [CrossRef]
- Swofford, D.L. PAUP*: Phylogenetic Analysis Using Parsimony. (*and Other Methods), Version 4.0b10; Sinauer Associates: Sunderland, MA, USA, 2003. [Google Scholar]
- Ronquist, F.; Huelsenbeck, J.P. MrBayes 3: Bayesian phylogenetic inference under mixed models. Bioinformatics 2003, 19, 1572–1574. [Google Scholar] [CrossRef] [Green Version]
- Stamatakis, A. RAxML version 8: A tool for phylogenetic analysis and post-analysis of large phylogenies. Bioinformatics 2014, 30, 1312–1313. [Google Scholar] [CrossRef]
- Hillis, D.M.; Bull, J.J. An empirical test of bootstrapping as a method for assessing confidence in phylogenetic analysis. Syst. Biol. 1993, 42, 182–192. [Google Scholar] [CrossRef]
- Rannala, B.; Yang, Z. Probability distribution of molecular evolutionary trees: A new method of phylogenetic inference. J. Mol. Evol. 1996, 43, 304–311. [Google Scholar] [CrossRef]
- Posada, D.; Crandall, K.A. Modeltest: Testing the model of DNA substitution. Bioinformatics 1998, 14, 817–818. [Google Scholar] [CrossRef] [Green Version]
- Rambaut, A.; Drummond, A. FigTree v.1.3.1. Institute of Evolutionary Biology; University of Edinburgh: Edinburgh, UK, 2010. [Google Scholar]
- Xu, C.N.; Zhang, H.J.; Zhou, Z.S.; Hu, T.; Wang, S.T.; Wang, Y.N.; Cao, K.Q. Identification and distribution of Botryosphaeriaceae species associated with blueberry stem blight in China. Eur. J. Plant Pathol. 2015, 143, 737–752. [Google Scholar] [CrossRef]
- Zhao, L.; Wang, Y.; He, W.; Zhang, Y. Stem blight of blueberry caused by Lasiodiplodia vaccinii sp. nov. in China. Plant Dis. 2019, 103, 2041–2050. [Google Scholar] [CrossRef]
- Strausbaugh, C.A.; Dugan, F. A novel Penicillium sp. causes rot in stored sugar beet roots in Idaho. Plant Dis. 2017, 101, 1781–1787. [Google Scholar] [CrossRef]
- Elfar, K.; Torres, R.; Díaz, G.A.; Latorre, B.A. Characterization of Diaporthe australafricana and Diaporthe spp. associated with stem canker of blueberry in Chile. Plant Dis. 2013, 97, 1042–1050. [Google Scholar] [CrossRef] [Green Version]
- Xu, C.N. Etiology, Molecular Detection and Control of Blueberry Stem Blight in China. Ph.D. Thesis, Hebei Agricultural University, Baoding, Hebei, China, 2016. [Google Scholar]
- Lin, Z.Y.; Wei, J.J.; Zhang, M.Q.; Xu, S.Q.; Guo, Q.; Wang, X.; Wang, J.H.; Chen, B.S.; Que, Y.X.; Deng, Z.H.; et al. Identification and characterization of a new fungal pathogen causing twisted leaf disease of sugarcane in China. Plant Dis. 2015, 99, 325–332. [Google Scholar] [CrossRef] [Green Version]
- Spaulding, P. Disease of Foreign Forest Trees Growing in the United States; Department of Agriculture: Washington, DC, USA, 1958.
Figure 1.
Phylogram of the best-parsimonious tree of Cytospora based on combined actin (ACT), internal transcribed spacer (ITS), RNA polymerase II second largest subunit (RPB2), translation elongation factor 1-alpha (TEF1-α) and beta-tubulin (TUB2) genes. Maximum parsimony (MP) and maximum likelihood (ML) bootstrap support values above 70% are shown at the first and second positions, respectively. Thickened branches represent posterior probabilities from Bayesian inference (BI) above 0.95. Scale bar = 200 nucleotide substitutions. Diaporthe vaccinii CBS 160.32 was used as the outgroup. Ex-type strains are in bold. Strains from the current study are in bold and blue.
Figure 1.
Phylogram of the best-parsimonious tree of Cytospora based on combined actin (ACT), internal transcribed spacer (ITS), RNA polymerase II second largest subunit (RPB2), translation elongation factor 1-alpha (TEF1-α) and beta-tubulin (TUB2) genes. Maximum parsimony (MP) and maximum likelihood (ML) bootstrap support values above 70% are shown at the first and second positions, respectively. Thickened branches represent posterior probabilities from Bayesian inference (BI) above 0.95. Scale bar = 200 nucleotide substitutions. Diaporthe vaccinii CBS 160.32 was used as the outgroup. Ex-type strains are in bold. Strains from the current study are in bold and blue.


Figure 2.
Phylogram of the best-parsimonious tree of Cytospora based on combined ACT, ITS, RPB2, TEF1-α and TUB2 genes. MP and ML bootstrap support values above 70% are shown at the first and second positions, respectively. Thickened branches represent posterior probabilities from BI above 0.95. Scale bar = 40 nucleotide substitutions. Diaporthe vaccinii CBS 160.32 was used as the outgroup. Ex-type strains are in bold. Strains from the current study are in bold and blue.
Figure 2.
Phylogram of the best-parsimonious tree of Cytospora based on combined ACT, ITS, RPB2, TEF1-α and TUB2 genes. MP and ML bootstrap support values above 70% are shown at the first and second positions, respectively. Thickened branches represent posterior probabilities from BI above 0.95. Scale bar = 40 nucleotide substitutions. Diaporthe vaccinii CBS 160.32 was used as the outgroup. Ex-type strains are in bold. Strains from the current study are in bold and blue.

Figure 3.
Cytospora haidianensis (CF 20198643). (a) Habitat of conidiomata on stems and branches of Euonymus alatus. (b) Transverse section of conidioma. (c) Longitudinal section through conidioma. (d) Conidiophores and conidiogenous cells. (e) Conidia. (f) Top (left) and bottom (right) sides of colonies on potato dextrose agar (PDA) after 30 days. Scale bars: a: 1 mm; b: 100 µm; c: 200 µm; d,e: 10 µm; f: 1 cm.
Figure 3.
Cytospora haidianensis (CF 20198643). (a) Habitat of conidiomata on stems and branches of Euonymus alatus. (b) Transverse section of conidioma. (c) Longitudinal section through conidioma. (d) Conidiophores and conidiogenous cells. (e) Conidia. (f) Top (left) and bottom (right) sides of colonies on potato dextrose agar (PDA) after 30 days. Scale bars: a: 1 mm; b: 100 µm; c: 200 µm; d,e: 10 µm; f: 1 cm.

Figure 4.
Stem blight symptoms on Euonymus alatus caused by Cytospora haidianensis. (a) Death of the whole plant caused by C. haidianensis on University Road, Beijing, China. (b) Stem blight caused by C. haidianensis in the greenhouse. Symptoms after (c) one week, (d) two weeks and (e) three weeks after inoculation of C. haidianensis. (f) Symptoms on Euonymus alatus twigs three weeks after inoculation of Cytospora haidianensis. (g,h) No symptoms on Euonymus alatus twigs after three weeks of inoculation with agar block (control).
Figure 4.
Stem blight symptoms on Euonymus alatus caused by Cytospora haidianensis. (a) Death of the whole plant caused by C. haidianensis on University Road, Beijing, China. (b) Stem blight caused by C. haidianensis in the greenhouse. Symptoms after (c) one week, (d) two weeks and (e) three weeks after inoculation of C. haidianensis. (f) Symptoms on Euonymus alatus twigs three weeks after inoculation of Cytospora haidianensis. (g,h) No symptoms on Euonymus alatus twigs after three weeks of inoculation with agar block (control).

Figure 5.
Regression curves and equations for mycelial growth of Cytospora haidianensis after incubation for 24, 48, 72 and 96 h at 0, 5, 10, 15, 20, 25, 30, 35 and 40 °C on PDA medium (X = temperature (°C), Y = growth (colony diameter, mm)). Optimal mycelial growth temperature was estimated to be 19.8 °C.
Figure 5.
Regression curves and equations for mycelial growth of Cytospora haidianensis after incubation for 24, 48, 72 and 96 h at 0, 5, 10, 15, 20, 25, 30, 35 and 40 °C on PDA medium (X = temperature (°C), Y = growth (colony diameter, mm)). Optimal mycelial growth temperature was estimated to be 19.8 °C.

Figure 6.
Regression curves and equations for mycelial growth of Cytospora haidianensis after incubation for 24, 48, 72 and 96 h at pH 2.0, 3.0, 4.0, 5.0, 6.0, 7.0, 8.0, 9.0, 10.0, 11.0 and 12.0 on PDA medium (X = pH, Y = growth (colony diameter, mm)). Optimum mycelial growth was estimated to be at pH 8.3.
Figure 6.
Regression curves and equations for mycelial growth of Cytospora haidianensis after incubation for 24, 48, 72 and 96 h at pH 2.0, 3.0, 4.0, 5.0, 6.0, 7.0, 8.0, 9.0, 10.0, 11.0 and 12.0 on PDA medium (X = pH, Y = growth (colony diameter, mm)). Optimum mycelial growth was estimated to be at pH 8.3.

Figure 7.
Effects of carbon source on growth of Cytospora haidianensis. Bars with uppercase or lowercase letters represent significant differences after, respectively, 24, 48, 72 and 96 h incubation, according to HSD tests at the p < 0.05 level.
Figure 7.
Effects of carbon source on growth of Cytospora haidianensis. Bars with uppercase or lowercase letters represent significant differences after, respectively, 24, 48, 72 and 96 h incubation, according to HSD tests at the p < 0.05 level.


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
Strains and GenBank accession numbers of Cytospora species used in the phylogenetic analyses in this study.
Table 1.
Strains and GenBank accession numbers of Cytospora species used in the phylogenetic analyses in this study.
| Species | Strain1 | Host | Origin | GenBank Accession Numbers | ||||
|---|---|---|---|---|---|---|---|---|
| ACT | ITS | RPB2 | TEF1-α | TUB2 | ||||
| Cytospora ailanthicola | CFCC 89970T | Ailanthus altissima | China | MH933526 | MH933618 | MH933592 | MH933494 | MH933565 |
| Cytospora leucosperma | CFCC 89622 | Pyrus bretschneideri | China | KU710988 | KR045616 | KU710944 | KU710911 | KR045657 |
| CFCC 89894 | Pyrus bretschneideri | China | KU710989 | KR045617 | KU710945 | KU710912 | KR045658 | |
| Cytospora ampulliformis | MFLUCC 16-0583T | Sorbus intermedia | Russia | KY417692 | KY417726 | KY417794 | NA | NA |
| MFLUCC 16-0629 | Acer platanoides | Russia | KY417693 | KY417727 | KY417795 | NA | NA | |
| Cytospora amygdali | CBS 144233T | Prunus dulcis | USA | MG972002 | MG971853 | NA | MG971659 | MG971718 |
| Cytospora atrocirrhata | CFCC 89615 | Juglans regia | China | KF498673 | KR045618 | KU710946 | KP310858 | KR045659 |
| CFCC 89616 | Juglans regia | China | KF498674 | KR045619 | KU710947 | KP310859 | KR045660 | |
| Cytospora beilinensis | CFCC 50493T | Pinus armandii | China | MH933527 | MH933619 | NA | MH933495 | MH933561 |
| CFCC 50494 | Pinus armandii | China | MH933528 | MH933620 | NA | MH933496 | MH933562 | |
| Cytospora berberidis | CFCC 89927T | Berberis dasystachya | China | KU710990 | KR045620 | KU710948 | KU710913 | KR045661 |
| CFCC 89933 | Berberis dasystachya | China | KU710991 | KR045621 | KU710949 | KU710914 | KR045662 | |
| Cytospora bungeana | CFCC 50495T | Pinus bungeana | China | MH933529 | MH933621 | MH933593 | MH933497 | MH933563 |
| CFCC 50496 | Pinus bungeana | China | MH933530 | MH933622 | MH933594 | MH933498 | MH933564 | |
| Cytospora californica | CBS 144234T | Juglans regia | USA | MG972083 | MG971935 | NA | MG971645 | NA |
| Cytospora carbonacea | CFCC 89947 | Ulmus pumila | China | KP310842 | KR045622 | KU710950 | KP310855 | KP310825 |
| Cytospora carpobroti | CMW 48981T | Carpobrotus edulis | South Africa | NA | MH382812 | NA | MH411212 | MH411207 |
| Cytosporacastanae | DBT 183T | Castanea sativa | North India | NA | KC963921 | NA | NA | NA |
| Cytospora celtidicola | CFCC 50497T | Celtis sinensis | China | MH933531 | MH933623 | MH933595 | MH933499 | MH933566 |
| CFCC 50498 | Celtis sinensis | China | MH933532 | MH933624 | MH933596 | MH933500 | MH933567 | |
| Cytospora centrivillosa | MFLUCC 16-1206T | Sorbus domestica | Italy | NA | MF190122 | MF377600 | NA | NA |
| MFLUCC 17-1660 | Sorbus domestica | Italy | NA | MF190123 | MF377601 | NA | NA | |
| Cytospora ceratosperma | CFCC 89624 | Juglans regia | China | NA | KR045645 | KU710976 | KP310860 | KR045686 |
| CFCC 89625 | Juglans regia | China | NA | KR045646 | KU710977 | KP31086 | KR045687 | |
| Cytospora ceratospermopsis | CFCC 89626T | Juglans regia | China | KU711011 | KR045647 | KU710978 | KU710934 | KR045688 |
| CFCC 89627 | Juglans regia | China | KU711012 | KR045648 | KU710979 | KU710935 | KR045689 | |
| Cytospora chrysosperma | CFCC 89629 | Salix psammophila | China | NA | KF765673 | KF765705 | NA | NA |
| CFCC 89981 | Populus alba subsp. pyramidalis | China | MH933533 | MH933625 | MH933597 | MH933501 | MH933568 | |
| CFCC 89982 | Ulmus pumila | China | KP310835 | KP281261 | NA | KP310848 | KP310818 | |
| Cytosporacoryli | CFCC 53162T | Corylus mandshurica | China | NA | MN854450 | MN850751 | MN850758 | MN861120 |
| Cytospora cotini | MFLUCC 14-1050T | Cotinus coggygria | Russia | NA | KX430142 | KX430144 | NA | NA |
| Cytospora curvata | MFLUCC 15-0865T | Salix alba | Russia | KY417694 | KY417728 | KY417796 | NA | NA |
| Cytospora davidiana | CXY 1350T | Populus davidiana | China | NA | KM034870 | NA | NA | NA |
| CXY 1374 | Populus davidiana | China | NA | KM034869 | NA | NA | NA | |
| Cytosporadiopuiensis | MFLUCC 18-1419T | Undefined wood | Thailand | MN685819 | MK912137 | NA | NA | NA |
| Cytosporaleucostoma | MFLUCC 15-0864 | Crataegus monogyna | Ukraine | KY417729 | KY417729 | KY41769 | KY417797 | NA |
| Cytospora elaeagni | CFCC 89632 | Elaeagnus angustifolia | China | KU710995 | KR045626 | KU710955 | KU710918 | KR045667 |
| CFCC 89633 | Elaeagnus angustifolia | China | KU710996 | KF765677 | KU710956 | KU710919 | KR045668 | |
| Cytospora elaeagnicola | CFCC 52882T | Elaeagnus angustifolia | China | MK732344 | MK732341 | MK732347 | NA | NA |
| CFCC 52883 | Elaeagnus angustifolia | China | MK732345 | MK732342 | MK732348 | NA | NA | |
| CFCC 52884 | Elaeagnus angustifolia | China | MK732346 | MK732343 | MK732349 | NA | NA | |
| Cytospora erumpens | CFCC 50022 | Prunus padus | China | MH933534 | MH933627 | NA | MH933502 | MH933569 |
| MFLUCC 16-0580T | Salix × fragilis | Russia | KY417699 | KY417733 | KY417801 | NA | NA | |
| Cytospora eucalypti | CBS 144241 | Eucalyptus globulus | USA | MG972056 | MG971907 | NA | MG971617 | MG971772 |
| Cytospora euonymicola | CFCC 50499T | Euonymus kiautschovicus | China | MH933535 | MH933628 | MH933598 | MH933503 | MH933570 |
| CFCC 50500 | Euonymus kiautschovicus | China | MH933536 | MH933629 | MH933599 | MH933504 | MH933571 | |
| Cytospora euonymina | CFCC 89993T | Euonymus kiautschovicus | China | MH933537 | MH933630 | MH933600 | MH933505 | MH933590 |
| CFCC 89999 | Euonymus kiautschovicus | China | MH933538 | MH933631 | MH933601 | MH933506 | MH933591 | |
| Cytospora fraxinigena | MFLUCC 14-0868T | Fraxinus ornus | Italy | NA | MF190133 | NA | NA | NA |
| MFLU 17-0880 | Fraxinus ornus | Italy | NA | MF190134 | NA | NA | NA | |
| Cytospora fugax | CXY 1371 | Populus simonii | China | NA | KM034852 | NA | NA | KM034891 |
| CXY 1381 | Populus ussuriensis | China | NA | KM034853 | NA | NA | KM034890 | |
| Cytosporagalegicola | MFLUCC 18-1199T | Galega officinalis | Italy | MN685810 | MK912128 | MN685820 | NA | NA |
| Cytospora germanica | CXY 1322 | Elaeagnus oxycarpa | China | NA | JQ086563 | NA | NA | NA |
| Cytospora gigalocus | CFCC 89620T | Juglans regia | China | KU710997 | KR045628 | KU710957 | KU710920 | KR045669 |
| CFCC 89621 | Juglans regia | China | KU710998 | KR045629 | KU710958 | KU710921 | KR045670 | |
| Cytospora gigaspora | CFCC 50014 | Juniperus procumbens | China | KU710999. | KR045630 | KU710959 | KU710922 | KR045671 |
| CFCC 89634T | Salix psammophila | China | KU711000 | KF765671 | KU710960 | KU710923 | KR045672 | |
| Cytospora granati | CBS 144237T | Punica granatum | USA | MG971949 | MG971799 | NA | MG971514 | MG971664 |
| Cytosporahaidianensis | CFCC 54056 | Euonymus alatus | China | MT363978 | MT360041 | MT363987 | MT363997 | MT364007 |
| CFCC 54057T | Euonymus alatus | China | MT363979 | MT360042 | MT363988 | MT363998 | MT364008 | |
| CFCC 54184 | Euonymus alatus | China | MT363980 | MT360043 | MT363989 | MT363999 | MT364009 | |
| Cytospora hippophaës | CFCC 89639 | Hippophaë rhamnoides | China | KU711001 | KR045632 | KU710961 | KU710924 | KR045673 |
| CFCC 89640 | Hippophaë rhamnoides | China | KF765730 | KF765682 | KU710962 | KP310865 | KR045674 | |
| Cytospora japonica | CFCC 89956 | Prunus cerasifera | China | KU710993 | KR045624 | KU710953 | KU710916 | KR045665 |
| CFCC 89960 | Prunus cerasifera | China | KU710994 | KR045625 | KU710954 | KU710917 | KR045666 | |
| Cytospora joaquinensis | CBS 144235T | Populus deltoides | USA | MG972044 | MG971895 | NA | MG971605 | MG971761 |
| Cytospora junipericola | BBH 42444 | Juniperus communis | Italy | NA | MF190126 | NA | MF377579 | NA |
| MFLU 17-0882T | Juniperus communis | Italy | NA | MF190125 | NA | MF377580 | NA | |
| Cytospora juniperina | CFCC 50501T | Juniperus przewalskii | China | MH933539 | MH933632 | MH933602 | MH933507 | NA |
| CFCC 50502 | Juniperus przewalskii | China | MH933540 | MH933633 | MH933603 | MH933508 | MH933572 | |
| CFCC 50503 | Juniperus przewalskii | China | MH933541 | MH933634 | MH933604 | MH933509 | NA | |
| Cytospora kantschavelii | CXY 1383 | Populus maximowiczii | China | NA | KM034867 | NA | NA | NA |
| CXY 1386 | Populus maximowiczii | China | NA | KM034867 | NA | NA | NA | |
| Cytospora kuanchengensis | CFCC 52464T | Castanea mollissima | China | MK442940 | MK432616 | MK578076 | NA | NA |
| CFCC 52465 | Castanea mollissima | China | MK442941 | MK432617 | MK578077 | NA | NA | |
| Cytospora leucostoma | CFCC 50015 | Sorbus aucuparia | China | KU711002 | KR045634 | NA | KU710925 | KR045675 |
| CFCC 50016 | Sorbus aucuparia | China | MH820408 | MH820400 | NA | MH820404 | MH820389 | |
| CFCC 50017 | Prunus cerasifera | China | MH933542 | MH933635 | NA | MH933510 | MH933573 | |
| CFCC 50018 | Prunus serrulata | China | MH933543 | MH933636 | NA | MH933511 | MH933574 | |
| CFCC 50019 | Rosa helenae | China | MH933544 | MH933637 | NA | NA | NA | |
| CFCC 50020 | Prunus persica | China | MH933545 | MH933638 | NA | NA | NA | |
| CFCC 50021 | Prunus salicina | China | MH933546 | MH933639 | NA | MH933512 | MH933575 | |
| CFCC 50023 | Cornus alba | China | KU711003 | KR045635 | KU710964 | KU710926 | KR045676 | |
| CFCC 50024 | Prunus pseudocerasus | China | MH933547 | MH933640 | MH933605 | NA | MH933576 | |
| CFCC 50467 | Betula platyphylla | China | NA | KT732948 | NA | NA | NA | |
| CFCC 50468 | Betula platyphylla | China | NA | KT732949 | NA | NA | NA | |
| CFCC 53140 | Prunus sibirica | China | MN850760 | MN854445 | MN850746 | MN850753 | MN861115 | |
| CFCC 53141 | Prunus sibirica | China | MN850761 | MN854446 | MN850747 | MN850754 | MN861116 | |
| CFCC 53156 | Juglans mandshurica | China | MN850762 | MN854447 | MN850748 | MN850755 | MN861117 | |
| MFLUCC 16-0574 | Rosa sp. | Russia | KY417696 | KY417731 | KY417798 | NA | NA | |
| Cytospora longiostiolata | MFLUCC 16-0628T | Salix × fragilis | Russia | KY417700 | KY417734 | KY417802 | NA | NA |
| Cytospora longispora | CBS 144236T | Prunus domestica | USA | MG972054 | MG971905 | NA | MG971615 | MG971764 |
| Cytospora lumnitzericola | MFLUCC 17-0508T | Lumnitzera racernosa | Thailand | MH253457 | MG975778 | MH253453 | NA | NA |
| Cytospora mali | CFCC 50028 | Malus pumila | China | MH933548 | MH933641 | MH933606 | MH933513 | MH933577 |
| CFCC 50029 | Malus pumila | China | MH933549 | MH933642 | MH933607 | MH933514 | MH933578 | |
| CFCC 50030 | Malus pumila | China | MH933550 | MH933643 | MH933608 | MH933524 | MH933579 | |
| CFCC 50031 | Crataegus sp. | China | KU711004 | KR045636 | KU710965 | KU710927 | KR045677 | |
| CFCC 50044 | Malus baccata | China | KU711005 | KR045637 | KU710966 | KU710928 | KR045678 | |
| Cytospora melnikii | CFCC 89984 | Rhus typhina | China | MH933551 | MH933644 | MH933609 | MH933515 | MH933580 |
| MFLUCC 15-0851T | Malus domestica | Russia | KY417701 | KY417735 | KY417803 | NA | NA | |
| MFLUCC 16-0635 | Populus nigra var. italica | Russia | KY417702 | KY417736 | KY417804 | NA | NA | |
| Cytosporamyrtagena | CFCC 52454 | Castanea mollissima | China | MK442938 | MK432614 | MK578074 | NA | NA |
| CFCC 52455 | Castanea mollissima | China | MK442939 | MK432615 | MK578075 | NA | NA | |
| Cytospora nivea | MFLUCC 15-0860 | Salix acutifolia | Russia | KY417703 | KY417737 | KY417805 | NA | NA |
| CFCC 89641 | Elaeagnus angustifolia | China | KU711006 | KF765683 | KU710967 | KU710929 | KR045679 | |
| CFCC 89643 | Salix psammophila | China | NA | KF765685 | KU710968 | KP310863 | KP310829 | |
| Cytosporanotastroma | NE_TFR5 | Populus tremuloides | USA | NA | JX438632 | NA | JX438543 | NA |
| NE_TFR8 | Populus tremuloides | USA | NA | JX438633 | NA | JX438542 | NA | |
| Cytospora oleicola | CBS 144248T | Olea europaea | USA | MG972098 | MG971944 | NA | MG971660 | MG971752 |
| Cytospora palm | CXY 1276 | Cotinus coggygria | China | NA | JN402990 | NA | KJ781296 | NA |
| CXY 1280T | Cotinus coggygria | China | NA | JN411939 | NA | KJ781297 | NA | |
| Cytospora parakantschavelii | MFLUCC 15-0857T | Populus × sibirica | Russia | KY417704 | KY417738 | KY417806 | NA | NA |
| MFLUCC 16-0575 | Pyrus pyraster | Russia | KY417705 | KY417739 | KY417807 | NA | NA | |
| Cytospora parapistaciae | CBS 144506T | Pistacia vera | USA | MG971954 | MG971804 | NA | MG971519 | MG971669 |
| Cytospora parasitica | MFLUCC 15-0507T | Malus domestica | Russia | KY417706 | KY417740 | KY417808 | NA | NA |
| XJAU 2542-1 | Malus sp. | China | NA | MH798884 | NA | MH813452 | NA | |
| Cytospora paratranslucens | MFLUCC 15-0506T | Populus alba var. bolleana | Russia | KY417707 | KY417741 | KY417809 | NA | NA |
| MFLUCC 16-0627 | Populus alba | Russia | KY417708 | KY417742 | KY417810 | NA | NA | |
| Cytosporapiceae | CFCC 52841T | Picea crassifolia | China | MH820406 | MH820398 | MH820395 | MH820402 | MH820387 |
| CFCC 52842 | Picea crassifolia | China | MH820407 | MH820399 | MH820396 | MH820403 | MH820388 | |
| Cytosporapingbianensis | MFLUCC 18-1204T | Undefined wood | China | MN685817 | MK912135 | MN685826 | NA | NA |
| Cytospora pistaciae | CBS 144238T | Pistacia vera | USA | MG971952 | MG971802 | NA | MG971517 | MG971667 |
| Cytospora platanicola | MFLU 17-0327T | Platanus hybrida | Italy | MH253449 | MH253451 | MH253450 | NA | NA |
| Cytospora platyclada | CFCC 50504T | Platycladus orientalis | China | MH933552 | MH933645 | MH933610 | MH933516 | MH933581 |
| CFCC 50505 | Platycladus orientalis | China | MH933553 | MH933646 | MH933611 | MH933517 | MH933582 | |
| CFCC 50506 | Platycladus orientalis | China | MH933554 | MH933647 | MH933612 | MH933518 | MH933583 | |
| Cytospora platycladicola | CFCC 50038T | Platycladus orientalis | China | MH933555 | KT222840 | MH933613 | MH933519 | MH933584 |
| CFCC 50039 | Platycladus orientalis | China | KU711008 | KR045642 | KU710973 | KU710931 | KR045683 | |
| Cytospora plurivora | CBS 144239T | Olea europaea | USA | MG972010 | MG971861 | NA | MG971572 | MG971726 |
| Cytospora populicola | CBS 144240T | Populus deltoides | USA | MG972040 | MG971891 | NA | MG971601 | MG971757 |
| Cytospora populina | CFCC 89644T | Salix psammophila | China | KU711007 | KF765686 | KU710969 | KU710930 | KR045681 |
| Cytospora populinopsis | CFCC 50032T | Sorbus aucuparia | China | MH933556 | MH933648 | MH933614 | MH933520 | MH933585 |
| CFCC 50033 | Sorbus aucuparia | China | MH933557 | MH933649 | MH933615 | MH933521 | MH933586 | |
| Cytospora pruinopsis | CFCC 50034T | Ulmus pumila | China | KP310836 | KP281259 | KU710970 | KP310849 | KP310819 |
| CFCC 50035 | Ulmus pumila | China | KP310837 | KP281260 | KU710971 | KP310850 | KP310820 | |
| CFCC 53153 | Ulmus pumila | China | MN850763 | MN854451 | MN850752 | MN850759 | MN861121 | |
| Cytospora predappioensis | MFLUCC 17-2458T | Platanus hybrida | Italy | NA | MG873484 | NA | NA | NA |
| Cytospora pruinosa | CFCC 50036 | Syringa oblata | China | KP310832 | KP310800 | NA | KP310845 | KP310815 |
| CFCC 50037 | Syringa oblata | China | MH933558 | MH933650 | NA | MH933522 | MH933589 | |
| Cytospora prunicola | MFLU 17-0995T | Prunus sp. | Italy | MG742353 | MG742350 | MG742352 | NA | NA |
| Cytosporapubescentis | MFLUCC 18-1201T | Quercus pubescens | Italy | MN685812 | MK912130 | MN685821 | NA | NA |
| Cytospora punicae | CBS 144244 | Punica granatum | USA | MG972091 | MG971943 | NA | MG971654 | MG971798 |
| Cytospora quercicola | MFLUCC 14-0867T | Quercus sp. | Italy | NA | MF190129 | NA | NA | NA |
| MFLU 17-0881 | Quercus sp. | Italy | NA | MF190128 | NA | NA | NA | |
| Cytospora ribis | CFCC 50026 | Ulmus pumila | China | KP310843 | KP281267 | KU710972 | KP310856 | KP310826 |
| CFCC 50027 | Ulmus pumila | China | KP310844 | KP281268 | NA | KP310857 | KP310827 | |
| Cytospora rosae | MFLU 17-0885 | Rosa canina | Italy | NA | MF190131 | NA | NA | NA |
| Cytospora rostrata | CFCC 89909T | Salix cupularis | China | KU711009 | KR045643 | KU710974 | KU710932 | KR045684 |
| CFCC 89910 | Salix cupularis | China | KU711010 | KR045644 | KU710975 | KU710933 | NA | |
| Cytospora rusanovii | MFLUCC 15-0853 | Populus × sibirica | Russia | KY417709 | KY417743 | KY417811 | NA | NA |
| MFLUCC 15-0854T | Salix babylonica | Russia | KY417710 | KY417744 | KY417812 | NA | NA | |
| Cytospora salicacearum | MFLUCC 15-0861 | Salix × fragilis | Russia | KY417711 | KY417745 | KY417813 | NA | NA |
| MFLUCC 15-0509T | Salix alba | Russia | KY417712 | KY417746 | KY417814 | NA | NA | |
| MFLUCC 16-0576 | Populus nigra var. italica | Russia | KY417707 | KY417741 | KY417809 | NA | NA | |
| MFLUCC 16-0587 | Prunus cerasus | Russia | KY417708 | KY417742 | KY417810 | NA | NA | |
| Cytospora salicicola | MFLUCC 15-0866 | Salix alba | Russia | KY417715 | KY417749 | KY417817 | NA | NA |
| MFLUCC 14-1052T | Salix alba | Russia | KU982637 | KU982636 | NA | NA | NA | |
| Cytospora salicina | MFLUCC 15-0862T | Salix alba | Russia | KY417716 | KY417750 | KY417818 | NA | NA |
| MFLUCC 16-0637 | Salix × fragilis | Russia | KY417717 | KY417751 | KY417819 | NA | NA | |
| Cytospora schulzeri | CFCC 50040 | Malus domestica | China | KU711013 | KR045649 | KU710980 | KU710936 | KR045690 |
| CFCC 50042 | Malus asiatica | China | KU711014 | KR045650 | KU710981 | KU710937 | KR045691 | |
| Cytospora sibiraeae | CFCC 50045T | Sibiraea angustata | China | KU711015 | KR045651 | KU710982 | KU710938 | KR045692 |
| CFCC 50046 | Sibiraea angustata | China | KU711015 | KR045652 | KU710983 | KU710939 | KR045693 | |
| Cytospora sophorae | CFCC 50047 | Styphnolobium japonicum | China | KU711017 | KR045653 | KU710984 | KU710940 | KR045694 |
| CFCC 50048 | Magnolia grandiflora | China | MH820409 | MH820401 | MH820397 | MH820405 | MH820390 | |
| CFCC 89598 | Styphnolobium japonicum | China | KU711018 | KR045654 | KU710985 | KU710941 | KR045695 | |
| Cytospora sophoricola | CFCC 89595T | Styphnolobium japonicum var. pendula | China | KU711019 | KR045655 | KU710986 | KU710942 | KR045696 |
| CFCC 89596 | Styphnolobium japonicum var. pendula | China | KU711020 | KR045656 | KU710987 | KU710943 | KR045697 | |
| Cytospora sophoriopsis | CFCC 89600T | Styphnolobium japonicum | China | KU710992 | KR045623 | KU710951 | KU710915 | KP310817 |
| Cytospora sorbi | MFLUCC 16-0631T | Sorbus aucuparia | Russia | KY417718 | KY417752 | KY417820 | NA | NA |
| Cytospora sorbicola | MFLUCC 16-0584T | Acer pseudoplatanus | Russia | KY417721 | KY417755 | KY417823 | NA | NA |
| MFLUCC 16-0633 | Cotoneaster melanocarpus | Russia | KY417724 | KY417758 | KY417826 | NA | NA | |
| Cytospora spiraeae | CFCC 50049T | Spiraea salicifolia | China | MG708196 | MG707859 | MG708199 | NA | NA |
| CFCC 50050 | Spiraea salicifolia | China | MG708197 | MG707860 | MG708200 | NA | NA | |
| Cytosporaspiraeicola | CFCC 53138T | Spiraea salicifolia | China | NA | MN854448 | MN850749 | MN850756 | MN861118 |
| CFCC 53139 | Tilia nobilis | China | NA | MN854449 | NA | NA | NA | |
| Cytospora tamaricicola | CFCC 50507 | Rosa multifolora | China | MH933559 | MH933651 | MH933616 | MH933525 | MH933587 |
| CFCC 50508T | Tamarix chinensis | China | MH933560 | MH933652 | MH933617 | MH933523 | MH933588 | |
| Cytospora tanaitica | MFLUCC 14-1057T | Betula pubescens | Russia | KT459413 | KT459411 | NA | NA | NA |
| Cytospora thailandica | MFLUCC 17-0262T | Xylocarpus moluccensis | Thailand | MH253459 | MG975776 | MH253455 | NA | NA |
| MFLUCC 17-0263T | Xylocarpus moluccensis | Thailand | MH253460 | MG975777 | MH253456 | NA | NA | |
| Cytospora tibouchinae | CPC 26333T | Tibouchina semidecandra | France | NA | KX228284 | NA | NA | NA |
| Cytospora translucens | CXY 1351 | Populus davidiana | China | NA | KM034874 | NA | NA | KM034895 |
| Cytospora ulmi | MFLUCC 15-0863T | Ulmus minor | Russia | NA | KY417759 | NA | NA | NA |
| Cytospora vinacea | CBS 141585T | Vitis interspecific hybrid ‘Vidal’ | USA | NA | KX256256 | NA | KX256277 | KX256235 |
| Cytosporaxinglongensis | CFCC 52458 | Castanea mollissima | China | MK442946 | MK432622 | MK578082 | NA | NA |
| CFCC 52459 | Castanea mollissima | China | MK442947 | MK432623 | MK578083 | NA | NA | |
| Cytospora viridistroma | CBS 202.36T | Cercis canadensis Castigl. | USA | NA | MN172408 | NA | MN271853 | NA |
| Cytospora viticola | CBS 141586T | Vitis vinifera | USA | NA | KX256239 | NA | KX256260 | KX256218 |
| Cytospora xylocarpi | MFLUCC 17-0251T | Xylocarpus granatum | Thailand | MH253458 | MG975775 | MH253454 | NA | NA |
| Diaporthe vaccinii | CBS 160.32 | Vaccinium macrocarpon | USA | JQ807297 | KC343228 | NA | KC343954 | KC344196 |
BBH, BIOTEC Bangkok Herbarium, National Science and Technology Development Agency, Thailand; CBS, Westerdijk Fungal Biodiversity Institute (CBS-KNAW Fungal Biodiversity Centre), Utrecht, Netherlands; CFCC, China Forestry Culture Collection Centre, Beijing, China; CMW, culture collection of Michael Wingfield, University of Pretoria, South Africa; CPC, culture collection of Pedro Crous, Netherlands; MFLU, Mae Fah Luang University herbarium, Thailand; MFLUCC, Mae Fah Luang University Culture Collection, Thailand; XJAU, Xinjiang Agricultural University, Xinjiang, China; NA, not applicable. All new isolates used in this study are indicated in bold type and strains from generic type species are marked by a superscript T.
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Zhou, X.; Pan, M.; Li, H.; Tian, C.; Fan, X. Dieback of Euonymus alatus (Celastraceae) Caused by Cytospora haidianensis sp. nov. in China. Forests 2020, 11, 524. https://0-doi-org.brum.beds.ac.uk/10.3390/f11050524
AMA Style
Zhou X, Pan M, Li H, Tian C, Fan X. Dieback of Euonymus alatus (Celastraceae) Caused by Cytospora haidianensis sp. nov. in China. Forests. 2020; 11(5):524. https://0-doi-org.brum.beds.ac.uk/10.3390/f11050524
Chicago/Turabian StyleZhou, Xian, Meng Pan, Haoyu Li, Chengming Tian, and Xinlei Fan. 2020. "Dieback of Euonymus alatus (Celastraceae) Caused by Cytospora haidianensis sp. nov. in China" Forests 11, no. 5: 524. https://0-doi-org.brum.beds.ac.uk/10.3390/f11050524
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.