Plasticity of Leaf Traits of Juglans regia L. f. luodianense Liu et Xu Seedlings Under Different Light Conditions in Karst Habitats

Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Site
2.2. Experimental Design
2.3. Measurement and Calculation of Leaf Properties
2.3.1. Anatomical Structure of Leaves
2.3.2. Morphological and Biomass Indicators
2.3.3. Physiological and Biochemical Indices
3. Results
3.1. Anatomical Structure of Leaves
3.2. Leaf Morphology and Biomass
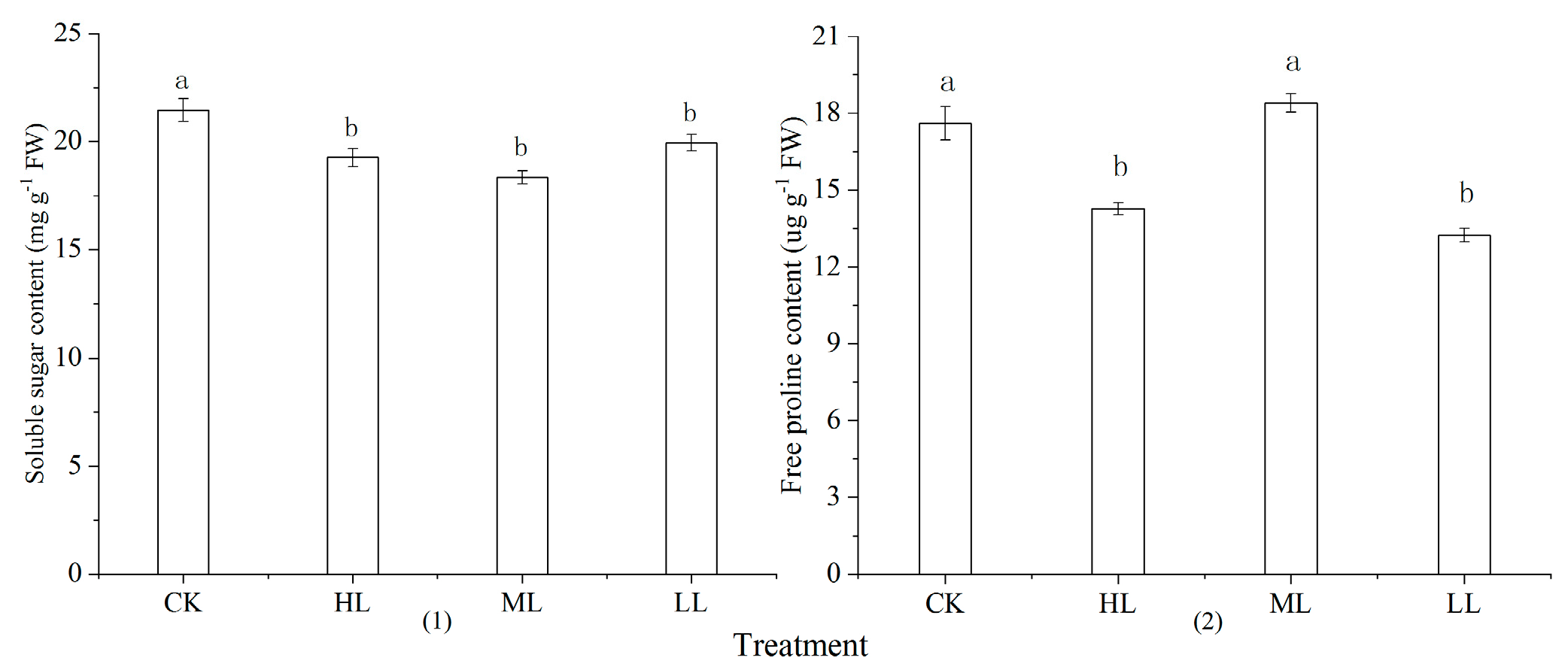
3.3. Soluble Matter Content of Leaves
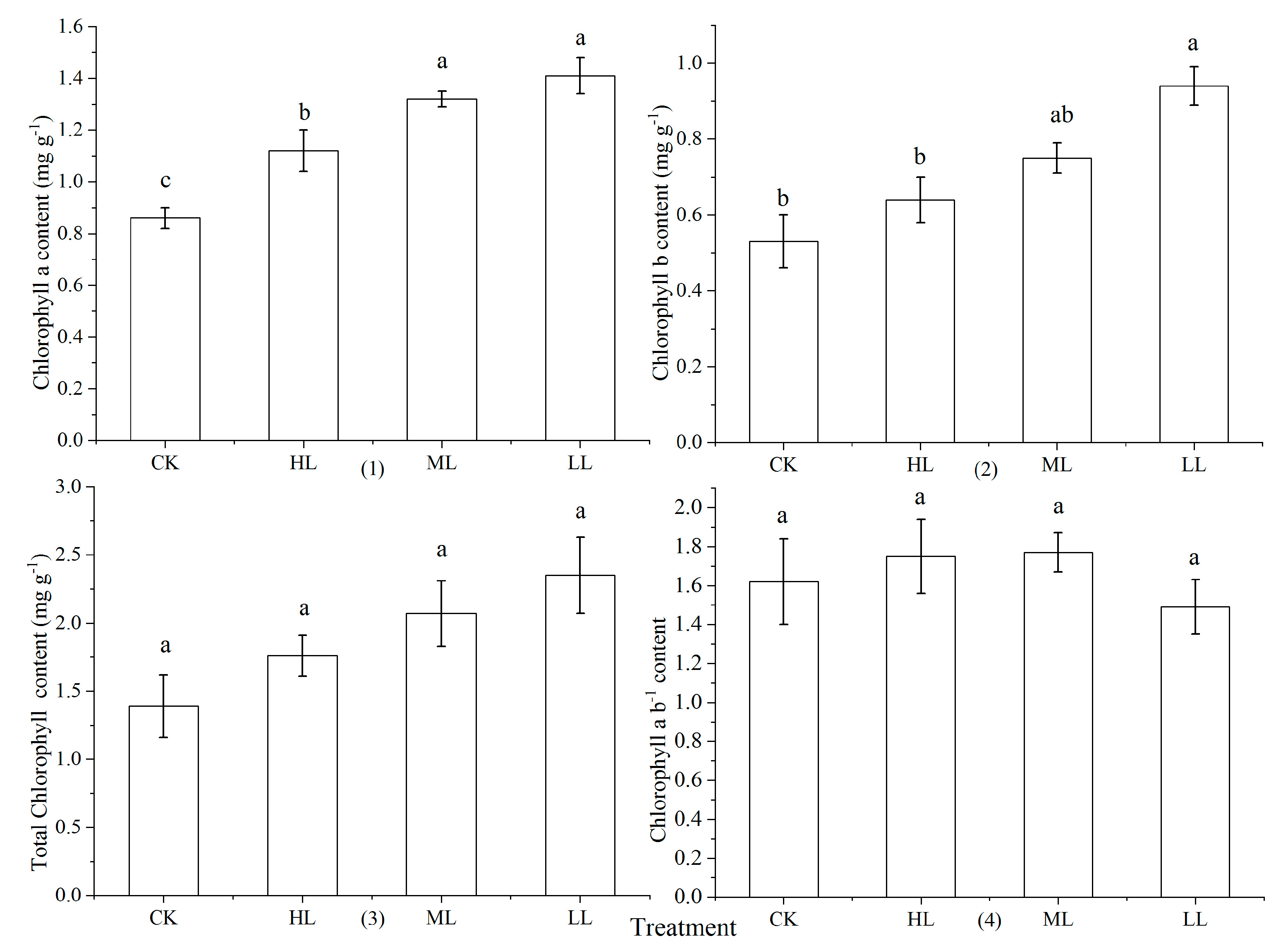
3.4. Chlorophyll Content
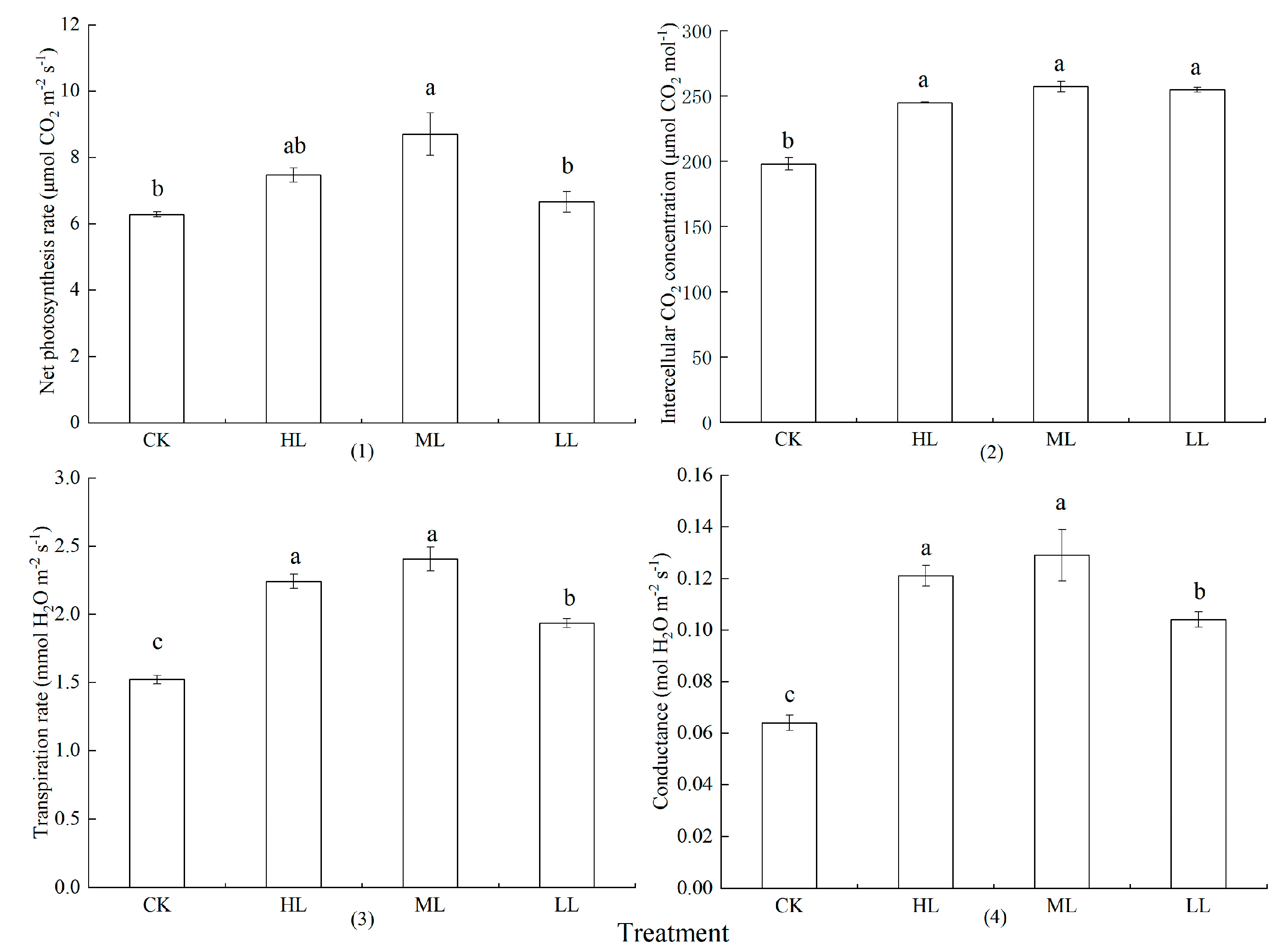
3.5. Photosynthetic Parameters
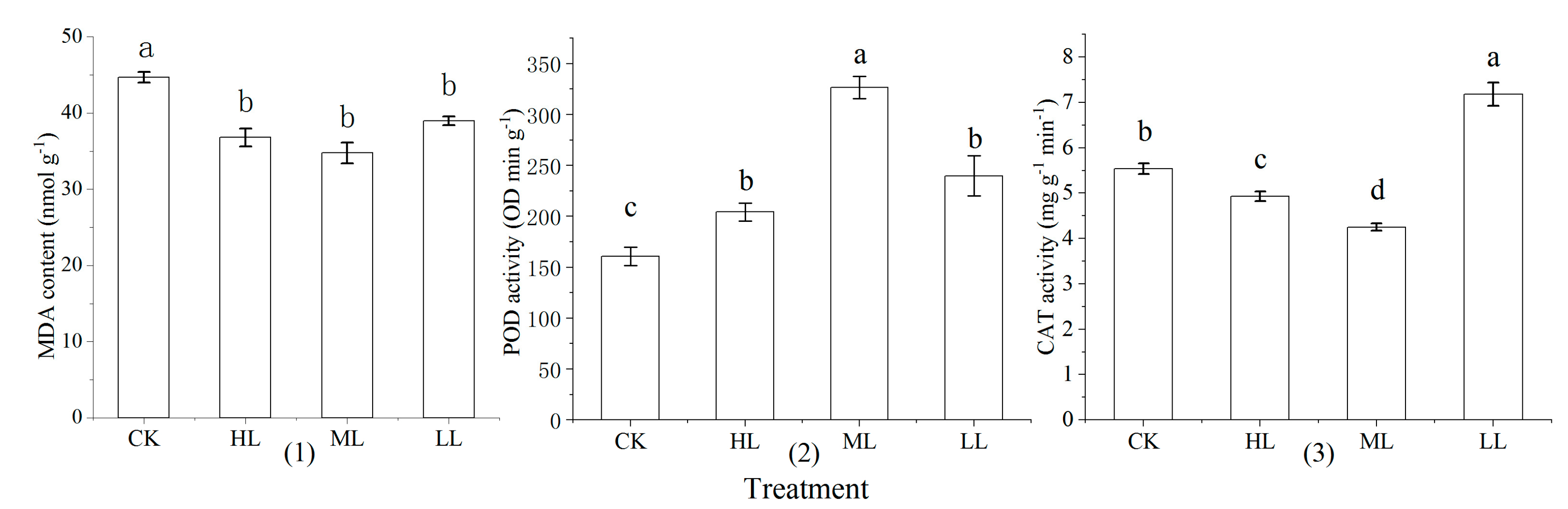
3.6. Malondialdehyde Content and Related Enzyme Activity
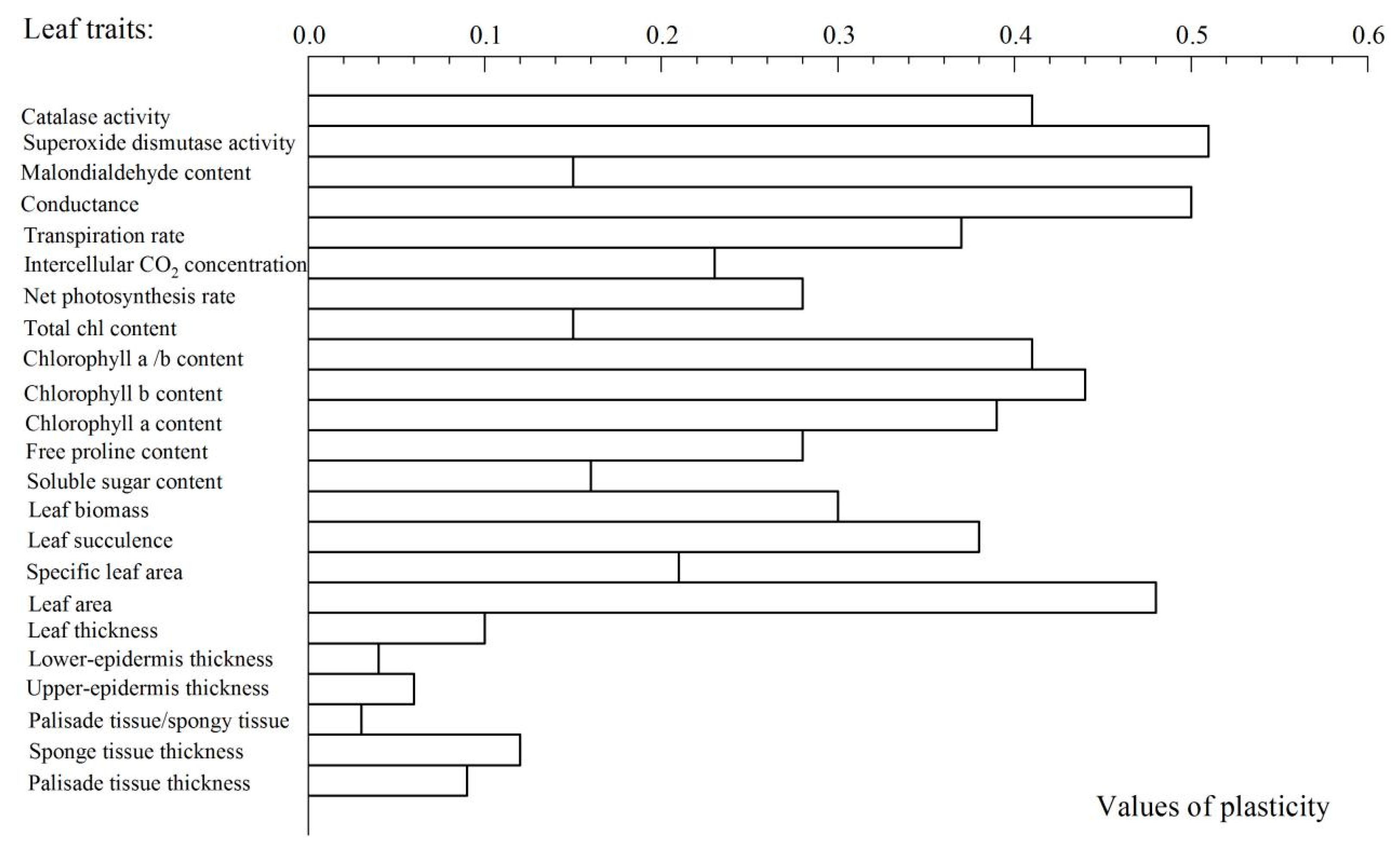
3.7. Evaluation of Leaf Plasticity
4. Discussion
4.1. Effect of Different Light Intensities on the Leaf Characteristics of J. Regia f. Luodianense Seedlings
4.2. Evaluation of the Plasticity of the Seedling Leaves of J. Regia f. Luodianense Under Different Light Intensities
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Kariny, C.N.; Lindamir, H.P.; Mariza, B.R.; de Luiz, A.S. Do eugenia hiemalis seedling leaves under different light conditions develop phenotypic plasticity? Plant Ecol. 2015, 216, 1571–1581. [Google Scholar]
- Sultan, S.E. Promising directions in plant phenotypic plasticity. Perspect. Plant Ecol. Evol. Syst. 2004, 6, 227–233. [Google Scholar] [CrossRef] [Green Version]
- Gianoli, E.; Valladares, F. Studying phenotypic plasticity: The advantages of a broad approach. Biol. J. Llinnean Soc. 2012, 105, 1–7. [Google Scholar] [CrossRef] [Green Version]
- Aubin-Horth, N.; Renn, S.C.P. Genomic reaction norms: Using integrative biology to understand molecular mechanisms of phenotypic plasticity. Mol. Ecol. 2009, 18, 3763–3780. [Google Scholar] [CrossRef]
- Lambers, H.; Chapin, F.S.; Pons, T.L. Plant Physiological Ecology, 2nd ed.; Springer: New York, NY, USA, 2008; pp. 320–374. [Google Scholar]
- Smith, H. Phytochromes and light signal perception by plants-an emerging synthesis. Nature 2000, 407, 585–591. [Google Scholar] [CrossRef]
- Chen, F.S.; Zeng, D.H.; Fahey, T.J.; Yao, C.Y.; Yu, Z.Y. Response of leaf anatomy of Chenopodium acuminatum to soil resource availability in a semi-arid grassland. Plant Ecol. 2010, 209, 375–382. [Google Scholar] [CrossRef]
- Valladares, F.; Arrieta, S.; Aranda, I.; Lorenzo, D.; Sánchez-Gómez, D.; Tena, D.; Suárez, F.; Alberto Pardos, J.A. Shade tolerance, photoinhibition sensitivity and phenotypic plasticity of Ilex aquifolium in continental Mediterranean sites. Tree Physiol. 2005, 25, 1041–1052. [Google Scholar] [CrossRef] [Green Version]
- Cornelissen, J.H.C.; Lavorel, S.; Garnier, E.; Díaz, S.; Buchmann, N.; Gurvich, D.E.; Reich, P.B.; Steege, H.; Morgan, H.D.; Heijden, M.G.; et al. A handbook of protocols for standardised and easy measurement of plant functional traits worldwide. Aust. J. Bot. 2003, 51, 335–380. [Google Scholar] [CrossRef] [Green Version]
- Vendramini, F.; Díaz, S.; Gurvich, D.E.; Wilson, P.J.; Thompson, K.; Hodgson, J.G. Leaf traits as indicators of resource-use strategy in floras with succulent species. New Phytol. 2002, 154, 147–157. [Google Scholar] [CrossRef]
- Pandey, S.K.; Singh, H.; Singh, J.S. Species and site effects on leaf traits of woody vegetation in a dry tropical environment. Curr. Sci. 2009, 96, 1109–1114. [Google Scholar]
- Poorter, H.; Nagel, O.W. The role of biomass allocation in the growth response of plants to different levels of light, CO2, nutrients and water: A quantitative review. Aust. J. Plant Physiol. 2000, 27, 595–607. [Google Scholar]
- Feng, Y.L.; Wang, J.F.; Sang, W.G. Biomass allocation, morphology and photosynthesis of invasive and noninvasive exotic species grown at four irradiance levels. Acta Oecol. 2007, 31, 40–47. [Google Scholar] [CrossRef]
- Rego, G.M.; Possamai, E. Efeito do sombreamento sobre o teor de clorofila e crescimento inicial do Jequitibá-Rosa. Bol. Pesqui. Florestal. 2006, 53, 179–194. [Google Scholar]
- Taiz, L.; Zeiger, E. Fisiologia Vegetal, 5th ed.; Artmed Porto Alegre: Rio Grande do Sul, Brazil, 2013. [Google Scholar]
- Tazoe, Y.; Noguchi, K.; Terashima, I. Effects of growth light and nitrogen nutrition on the organization of the photosynthetic apparatus in leaves of a C4 plant, Amaranthus cruentus. Plant Cell Environ. 2006, 29, 691–700. [Google Scholar] [CrossRef]
- Osada, N.; Onoda, Y.; Hikosaka, K. Effects of atmospheric CO2 concentration, irradiance, and soil nitrogen availability on leaf photosynthetic traits of Polygonum sachalinense around natural CO2 springs in northern Japan. Oecologia 2010, 164, 41–52. [Google Scholar] [CrossRef]
- Crabtree, R.C.; Bazzaz, F.A. Seedling response of four birch species to simulated nitrogen deposition: Ammonium vs. nitrate. Ecol. Appl. Publ. Ecol. Soc. Am. 1993, 3, 315–321. [Google Scholar] [CrossRef]
- Streb, P.; Tel-or, E.; Feierabend, J. Light stress effects and antioxidative protection in two desert plants. Funct. Ecol. 1997, 11, 416–424. [Google Scholar] [CrossRef]
- Valladares, F.; Wright, S.J.; Lasso, E.; Kitajima, K.; Pearcy, R.W. Plastic phenotypic response to light of 16 congeneric shrubs from a panamanina rainforest. Ecology 2000, 81, 1925–1936. [Google Scholar] [CrossRef]
- Tang, G.; Li, X.; Lin, L.; Li, L.; Lu, J. Change of different shading on moisture conditions and the physiological response in Alhagi sparsifolia. Chin. J. Plant Ecol. 2013, 37, 354–364. [Google Scholar] [CrossRef]
- Liu, J.M.; Xu, G.R. A New Forma of Juglans regia L. Guizhou Sci. 2011, 29, 95–96. [Google Scholar]
- Zhu, S.Q.; He, J.X.; Wei, L.M.; Zhang, C.G.; Chen, Z.R. Karst Forest Ecology Research III; Guizhou Science and Technology Press: Guiyang, China, 2003. [Google Scholar]
- Li, A.D.; Jia, S.; Yu, L.F. Microclimates of different microhabitats in Huajiang karst area. J. Zhejiang For. Coll. 2010, 27, 374–378. [Google Scholar]
- Du, X.L.; Wang, S.J. Micro-habitat characteristics in the karst desertification area: A case study of the wangjiazhai catchment in Guizhou Province. Earth Environ. 2010, 38, 255–261. [Google Scholar]
- Klich, M.G. Leaf variations in Elaeagnus angustifolia related to environmental heterogeneity. Environ. Exp. Bot. 2000, 44, 171–183. [Google Scholar] [CrossRef]
- Xu, G.R.; Liu, J.M.; Yan, G.H.; LI, M.J.; He, X.; Xu, X.J. Quantification methods of chlorophyll from Juglans regia L. f. luodianense. J. Mount. Agric. Biol. 2010, 29, 419–423. [Google Scholar]
- Liu, J.M.; Zhao, X.P.; Liao, X.F.; Zhang, D.K.; Yan, G.H.; Wang, M.; Wen, P. Study on protective enzyme system and MDA content of Juglans regia L. f. luodianense Liu et Xu under dry stress. J. Henan Agric. Sci. 2012, 41, 122–126. [Google Scholar]
- Wen, P.; Liu, J.M.; Xu, G.R.; Liao, X.F.; Zhang, D.K.; Yan, G.H.; Wang, M.; Zhao, X.P. Effect of water stress on photosynthesis and transpiration of Juglans regia L. f. luodianense Liu et Xu. Guizhou Agric. Sci. 2013, 41, 57–60. [Google Scholar]
- Wang, D.; Liu, J.M.; Xu, G.R.; Chen, J.Z. Main nutritional ingredients and morphological characters of Juglans regia L. f. luodianense Liu et Xu. Chin. Oils Fats 2019, 44, 95–98. [Google Scholar]
- Lu, Y.F.; Shi, L.; Yan, S.C. Effects of different light intensities on activities of the primary defense proteins in needles of Larix gmelinii. Acta Ecol. Sin. 2012, 32, 3621–3627. [Google Scholar]
- Wang, M.L.; Wei, X.; Tang, H.; Liang, H.L.; Zou, R. Effects of light intensity on growth and photosynthesis of three karst plant seedlings. Chin. J. Ecol. 2015, 34, 604–610. [Google Scholar]
- Gaskin, T.A. Light quality under saran shade cloth. Agron. J. 1965, 57, 313–314. [Google Scholar] [CrossRef]
- Zhou, Y. Experiment of Plant Tissue Anatomy (Revised); Beijing Normal University Press: Beijing, China, 1993. [Google Scholar]
- Lichtenthaler, H.K.; Wellburn, A.R. Determinations of total carotenoids and chlorophylls a and b of leaf extracts in different solvents. Biochem. Soc. Trans. 1983, 11, 591–592. [Google Scholar] [CrossRef] [Green Version]
- Zou, Q. Experimental Guidance of Plant Physiology; China Agriculture Press: Beijing, China, 2000. [Google Scholar]
- Li, H.S. Principles and Techniques of Plant Physiological and Biochemical Experiments; Higher Education Press: Beijing, China, 2000. [Google Scholar]
- Hodges, D.M.; DeLong, J.M.; Forney, C.F.; Prange, R.K. Improving the thiobarbituric acid-reactive-substances assay for estimating lipid peroxidation in plant tissues containing anthocyanin and other interfering compounds. Planta 1999, 207, 604–611. [Google Scholar] [CrossRef]
- Shi, J.; Fu, X.Z.; Peng, T.; Huang, X.S.; Fan, Q.J.; Liu, J.H. Spermine pretreatment confers dehydration tolerance of citrus in vitro plants via modulation of antioxidative capacity and stomatal response. Tree Physiol. 2010, 30, 914–922. [Google Scholar] [CrossRef] [PubMed]
- Aebi, H. Catalase in vitro. Methods Enzymol. 1984, 105, 121–126. [Google Scholar] [PubMed]
- Ashton, P.M.S.; Olander, L.P.; Berlyn, G.P.; Thadani, R.; Cameron, I.R. Changes in leaf structure in relation to crown position and tree size of Betula papyrifera within fire-origin stands of interior cedar-hemlock. Can. J. Bot. 1998, 76, 1180–1187. [Google Scholar]
- Vogelmann, T.C.; Martin, G. The functional significance of palisade tissue: Penetration of directional versus diffuse light. Plant Cell Environ. 1993, 16, 65–72. [Google Scholar] [CrossRef]
- James, S.A.; Bell, D.T. Influence of light availability on leaf structure and growth of two Eucalyptus globulus ssp. globulus provenances. Tree Physiol. 2000, 20, 1007–1018. [Google Scholar]
- Gordon, D.C.; Percy, K.E.; Riding, R.T. Effects of uv-B radiation on epicuticular wax production and chemical composition of four Picea species. New Phytol. 1998, 138, 441–449. [Google Scholar] [CrossRef]
- Dickson, W.C. Integrative Plant Anatomy; Academic Press: San Diego, CA, USA, 2000. [Google Scholar]
- Li, F.L.; Bao, W.K. Response and adaptation of morphological and anatomical structures of plant leaves to environmental changes. Bull. Bot. 2005, 22, 118–127. [Google Scholar]
- Parelle, J.; Roudaut, J.P.; Ducrey, M. Light acclimation and photosynthetic response of beech (Fagus sylvatical) saplings under artificial shading or natural Mediterranean conditions. Ann. For. Sci. 2006, 63, 257–266. [Google Scholar] [CrossRef] [Green Version]
- Espíndola-Júnior, A.; Boeger, M.R.T.; Maccari-Júnior, A. Anatomia foliar de Mikania glomerata Spreng. (Asteraceae) sob diferentes condições de luminosidade. Rev. Bras. Bot. 2009, 32, 749–758. [Google Scholar] [CrossRef] [Green Version]
- Guo, X.; Wang, R.Q.; Wang, C.D.; Xu, F.; Zhao, S.; Guo, W.H. Acer truncatum seedlings are more plastic than Quercus variabilis seedlings in response to different light regimes. Dendrobiology 2016, 76, 35–49. [Google Scholar] [CrossRef]
- Saldaña-Acosta, A.; Meave, J.A.; Sánchez-Velásquez, L.R. Seedling biomass allocation and vital rates of cloud forest tree species: Responses to light in shade house conditions. For. Ecol. Manag. 2009, 258, 1650–1659. [Google Scholar] [CrossRef]
- Xiao, H.; Wang, C.; Liu, J. Insights into the differences in leaf functional traits of heterophyllous Syringa oblata under different light intensities. J. For. Res. 2015, 26, 613–621. [Google Scholar] [CrossRef]
- Gommers, C.M.; Visser, E.J.; Onge, K.R.S.; Voesenek, L.A.; Pierik, R. Shade tolerance: When growing tall is not an option. Trends Plant Sci. 2013, 18, 65–71. [Google Scholar] [CrossRef]
- Zhang, D.L.; Zhang, L.; Ge, W.Z. Phenotypic plasticity and respond of Juglans mandshurica seedling under different light environments. Bull. Bot. Res. 2017, 37, 658–663. [Google Scholar]
- Liang, M.M.; Gong, M.M.; Li, H.; Zhang, S.Y.; Wang, Z.S.; Zhang, X.M.; Guo, S.P.; Qi, G.H. Effects of different shading treatments on the growth and development of ‘LYuling’ Juglans regia. J. Northwest Agric. For. Univ. 2017, 32, 120–124. [Google Scholar]
- Guo, Q.Q.; LI, H.E.; Gao, C.; Yang, R. Leaf traits and photosynthetic characteristics of endangered Sinopodophyllum hexandrum (Royle) Ying under different light regimesin Southeastern Tibet Plateau. Photosynthetica 2019, 57, 548–555. [Google Scholar]
- Xiong, J.; Wang, C.; Xing, W.L.; Wu, M.K.; Cheng, X.R.; Zhang, C. Morphological and physiological responses of Ardisia crenata seedlings under different light intensities. Plant Sci. J. 2018, 36, 736–744. [Google Scholar]
- Yu, S.W.; Tang, Z.C. Plant Physiology and Molecular Biology, 2nd ed.; Science Press: Beijing, China, 1998. [Google Scholar]
- Verbruggen, N.; Hermans, C. Proline accumulation in plants: A review. Amino Acids 2008, 35, 753–759. [Google Scholar] [CrossRef]
- Wang, D.; Chen, J.Z.; Xiong, X.; Wang, S.; Liu, J.M. Allelopathic effects of Cinnamomum migao on seed germination and seedling growth of its associated species Liquidambar formosana. Forests 2019, 10, 535. [Google Scholar] [CrossRef] [Green Version]
- Fang, C.; Zhihui, C. Research progress on the use of plant allelopathy in agriculture and the physiological and ecological mechanisms of allelopathy. Front. Plant Sci. 2015, 6, 1020. [Google Scholar]
- Pintó-Marijuan, M.; Munné-Bosch, S. Photo-oxidative stress markers as a measure of abiotic stress-induced leaf senes-cence: Advantages and limitations. J. Exp. Bot. 2014, 65, 3845–3857. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, G.L.; Chen, M.; Du, G.Z. Response of biomass allocation and morphological characteristics to light and nutrient resources for seedlings of three alpine species. Acta Ecol. Sin. 2010, 30, 60–66. [Google Scholar]
- Cheng, X.R.; Xing, W.L.; Yuan, H.J.; Xiong, J.; Yu, M.K. Phenotypic plasticity of Illicium lanceolatum in response to varied light environments. Acta Ecol. Sin. 2019, 39, 1935–1944. [Google Scholar]
- Wang, Y.; Wei, X.L. The effect of different light intensity on seedling ecological adaptability of two karst tree species. Guizhou Agric. Sci. 2009, 37, 139–142. [Google Scholar]
- Huang, H.T.; Huang, J.J.; Chen, J.; Chen, Y.J.; Guan, D.S. Growth, physiological and biochemical response of Baccaurea ramiflora Lour. seedlings to different shading environments. Chin. J. Ecol. 2020, 39, 1538–1547. [Google Scholar]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Treatments | Thickness of Palisade Tissue (μm) | Thickness of Spongy Tissue (μm) | Palisade Tissue/Spongy Tissue | Thickness of Upper Epidermis (μm) | Thickness of Lower Epidermis (μm) | Thickness of Leaf (μm) |
|---|---|---|---|---|---|---|
| CK | 47.46 ± 0.3 a | 59.96 ± 0.92 a | 0.82 ± 0.01 a | 11.64 ± 0.17 a | 5.25 ± 0.02 a | 124.31 ± 1.95 a |
| HL | 47.04 ± 0.84 ab | 57.43 ± 0.72 b | 0.81 ± 0.01 a | 11.35 ± 0.06 ab | 5.11 ± 0.02 b | 120.93 ± 1.54 a |
| ML | 45.28 ± 0.58 b | 54.18 ± 0.36 c | 0.81 ± 0.01 a | 11.20 ± 0.05 b | 5.07 ± 0.02 b | 115.73 ± 2.68 ab |
| LL | 43.28 ± 0.31 c | 52.78 ± 0.73 c | 0.83 ± 0.01 a | 10.89 ± 0.05 c | 5.06 ± 0.03 b | 112.01 ± 2.37 b |
| Treatments | Maximum Leaf Area (cm2) | Specific Leaf Area | Leaf Biomass (g) | Leaf Succulence |
|---|---|---|---|---|
| CK | 2705.54 ± 46.93 d | 3.52 ± 0.05 a | 9.36 ± 0.11 b | 251.99 ± 6.72 c |
| HL | 3069.02 ± 52.39 c | 3.63 ± 0.07 a | 9.90 ± 0.04 b | 254.54 ± 4.33 c |
| ML | 4690.26 ± 66.02 b | 3.46 ± 0.03 a | 13.03 ± 0.30 a | 299.17 ± 4.25 b |
| LL | 4994.78 ± 55.40 a | 2.24 ± 0.06 b | 12.81 ± 0.41 a | 321.99 ± 4.73 a |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, D.; Huang, X.; Chen, J.; Li, L.; Cheng, J.; Wang, S.; Liu, J. Plasticity of Leaf Traits of Juglans regia L. f. luodianense Liu et Xu Seedlings Under Different Light Conditions in Karst Habitats. Forests 2021, 12, 81. https://0-doi-org.brum.beds.ac.uk/10.3390/f12010081
Wang D, Huang X, Chen J, Li L, Cheng J, Wang S, Liu J. Plasticity of Leaf Traits of Juglans regia L. f. luodianense Liu et Xu Seedlings Under Different Light Conditions in Karst Habitats. Forests. 2021; 12(1):81. https://0-doi-org.brum.beds.ac.uk/10.3390/f12010081
Chicago/Turabian StyleWang, Deng, XiaoLong Huang, JingZhong Chen, LiXia Li, Jing Cheng, Shu Wang, and JiMing Liu. 2021. "Plasticity of Leaf Traits of Juglans regia L. f. luodianense Liu et Xu Seedlings Under Different Light Conditions in Karst Habitats" Forests 12, no. 1: 81. https://0-doi-org.brum.beds.ac.uk/10.3390/f12010081