
Biomass Yield and Economic, Energy and Carbon Balances of Ulmus pumila L., Robinia pseudoacacia L. and Populus × euroamericana (Dode) Guinier Short-Rotation Coppices on Degraded Lands under Mediterranean Climate


Abstract
:1. Introduction
2. Materials and Methods
2.1. Experimental Design and Plant Material
- Bare root nursery seedlings, 50–70 cm in length, of two Robinia pseudoacacia L. taxa (the improved cultivar “Nyírségi” and commercial nursery plants from Germany), Populus nigra L., Ailanthus altissima (Mill) Swingle., and Ulmus pumila L;
- Paulownia plants of 15–20 cm in height (Paulownia fortunei (Seem.) Hemsl., clone “UHU”) coming from root cuttings;
- Bare root nursery plants, 50–70 cm in length, coming from rooted hardwood cuttings of Platanus × hispanica Mill. ex Münchh;
- Poplar hardwood cuttings of 20–25 cm in length belonging to five hybrid clones—four clones of Populus × euroamericana (Dode) Guinier (clones “Adige”, “AF2”, “Oudenberg”, “I214”), and one clone of Populus × interamericana van Broekhuizen (clone “Raspalje”).
2.2. Shoot Growth and Biomass Assessment
2.3. Physical–Chemical Properties of Soil and Plant Biomass
2.4. Valuation of Biomass and Determination of the Economic, Energy and CO2 Balances of the Production Process
2.5. Data Analysis
3. Results
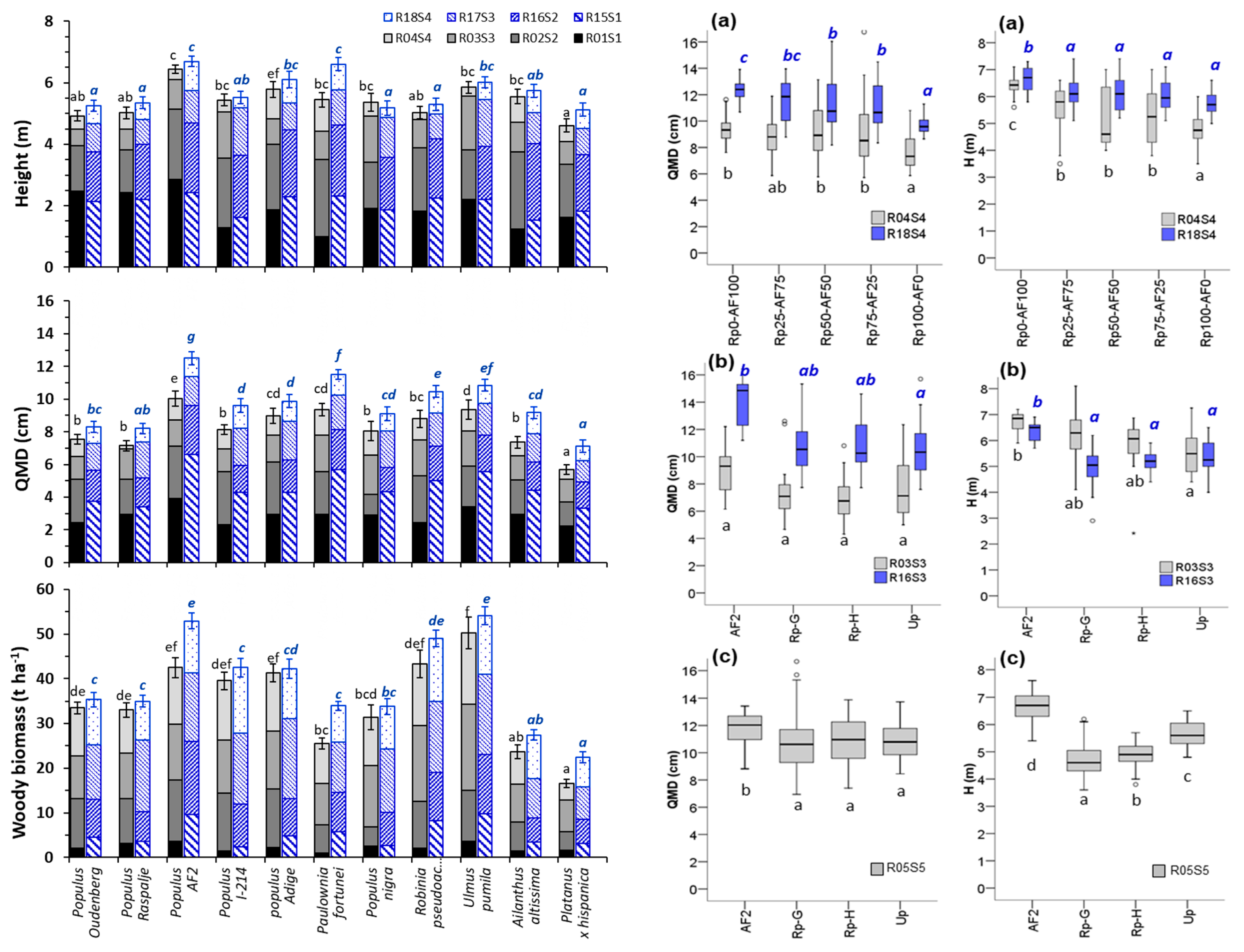
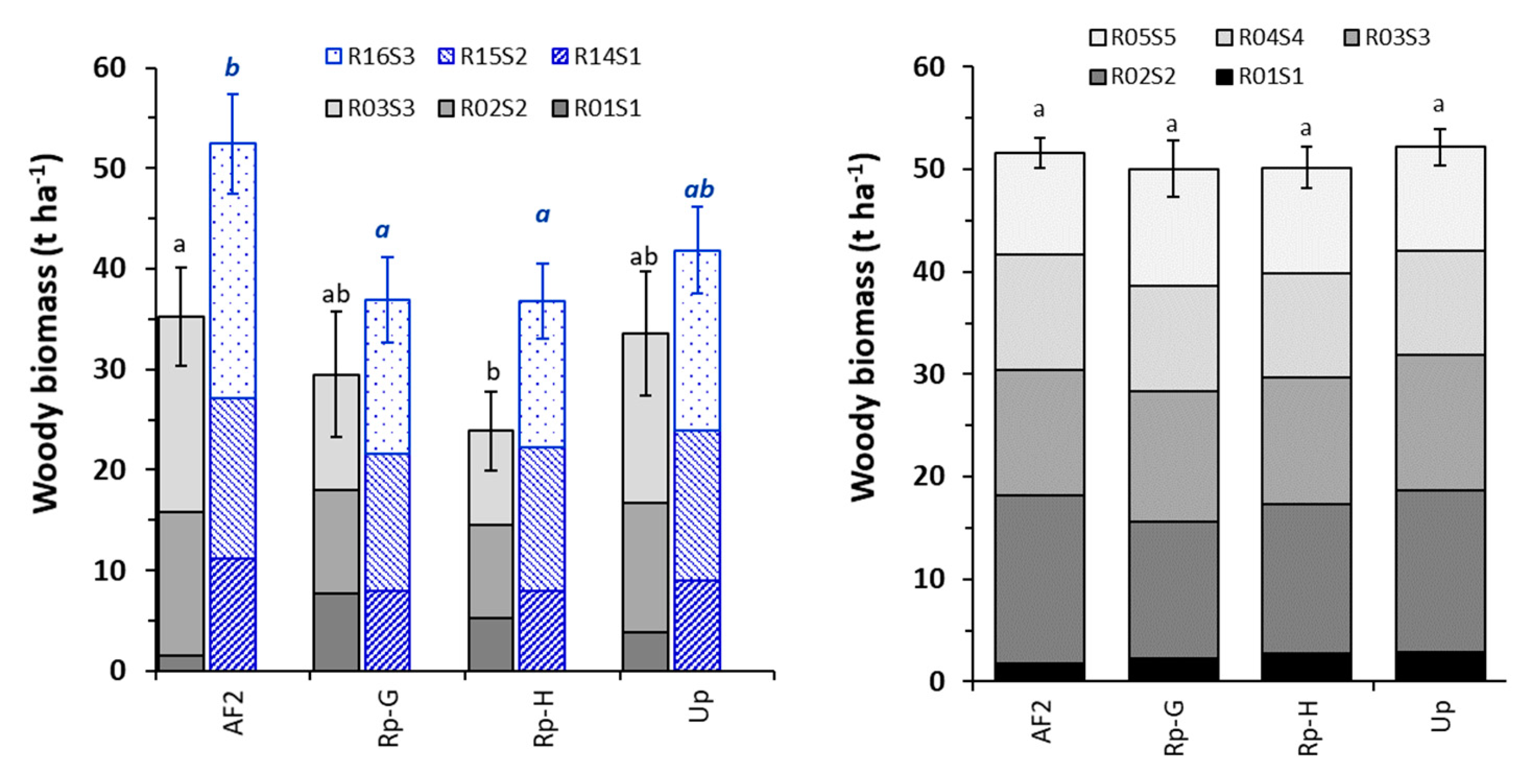
3.1. Shoot Growth and Biomass Production
3.2. Physical–Chemical Characterization of Soil and Plant Biomass
- A significant increase in exchangeable sodium percentage (ESP), organic carbon (OC), N content and available K content. The increase in ESP was evident at the end of the first rotation (3–4 years), but those of OC, N and K took 5–8 years to differentiate from the initial soil;
- The opposite trend was obtained for C/N ratio and the contents of P and active limestone—that is, a significant decrease over time;
- For the rest of the properties analyzed in the initial soils, no significant differences were obtained for the studied period (5–8 years, depending on the experiment);
- In the Ra4 assay, those cases in which differences between the two soil layers (0–15 and 15–30 cm) were significant were due to a higher mineral nutrient content (+ 40–75%), greater OC (+ 130–160%) and lower ESP (−28%) in the shallowest horizon (0–15 cm) than in the deepest.
- No significant differences were found between different trials (p > 0.060), and therefore the mean value from the four trials as a whole has been shown for each taxon;
- Significant differences between taxa were obtained for N, Ca, S, Fe, Cl and Ash contents (p < 0.040), although these differences were not very high in absolute terms. Robinia pseudoacacia stands out for its high N and low Ca contents, and high wood density;
- According to the ISO 17225-4 standard [67], the chipped woody biomass is of high quality, since it is harvested without leaves and has a low bark percentage (12% for Robinia pseudoacacia, 15% for Ulmus pumila, and 14% for Populus × euroamericana “AF2”);
- Due to their physical–mechanical and energetic properties (heating values; bulk density, mechanical durability and moisture of pellets), the highest-quality pellets (i.e., ENplus-A1 for domestic use) could be obtained for the three species studied according to standard ISO 17225-2 [50]. However, the chemical properties (N, S, Cl or ash contents) devalue their quality to commercial ENplus-B or Industrial grade pellets. ENplus-A2 quality pellets could be manufactured from debarked wood of these three species, since it contains ≤0.5% N, ≤0.05% S, ≤0.02% Cl and ≤1.2% ash;
- Considering the Mo1, Mo3 and Ra4 trials as a whole, the average amounts of N, P and K removed with the aboveground woody biomass harvested during the studied period were 53.7, 10.8 and 35.0 kg ha−1 year−1, respectively. The nutrient contents in the litterfall (11.5, 1.3 and 2.2 kg ha−1 year−1, respectively) and roots (47.1, 4.6 and 15.6 kg ha−1 year−1, respectively), as well as the new amounts accumulated in the uppermost layer of the soil (0–30 cm) (57.9, −5.5, 36.7 kg ha−1 year−1, respectively), must be added to those contained in the AGWD, which amounted to 170.2, 11.2 and 89.5 kg ha−1 year−1 during the study period. Since in this study the root biomass has not been evaluated, in order to estimate it, averages value of the AGWB/root dry matter ratio of 2.03, 1.70 and 2.10 have been considered for Robinia pseudoacacia, Ulmus pumila and Populus × euroamericana “AF2”, respectively, according to the results reported by other authors for plants of similar size [68,69,70];
- Taking into account that, on average for all the trials as a whole, the amounts of N, P and K supplied by fertilization (62.3, 27.2 and 51.7 kg ha−1 year−1, respectively) and by the irrigation water (24.4, 0.6 and 0.9 kg ha−1 year−1, respectively) were 86.7, 27.8 and 52.6 kg ha−1 year−1, respectively—for instance, in the case of N there were 83.5 kg ha−1 year−1 in the plants and in the soil layer, whose origin has not been determined.
3.3. Economic, Energy and CO2 Emission Balances of the Production Process
- The average biomass production of Populus × euroamericana “AF2”, Robinia pseudoacacia and Ulmus pumila as a whole in the SRC field trials carried out was considered for the estimates (Table 10);
- Although only one to two rotations have been studied in each field trial, the calculation has been extrapolated to four to five rotations, to complete a cycle of at least 15 years (Table 9). Biomass production and cultivation works from the third to the fourth–fifth rotations have been considered the same as for the second rotation;
- The biomass produced will be harvested, chipped and transported by truck over a distance of 25 km;
- The equivalent selling price of the chipped dry biomass, taken to the factory, has been estimated at EUR 82 t−1;
- The valves, filters, dosing pump, irrigation pump, pressure gauges, plastic pipes (PE, PVC), installation, etc., of the irrigation system have been taken into account in the economic costs. However, only plastic pipes have been considered in the assessment of energy costs and CO2 emissions;
- Diesel combustion releases 2.65 kg of CO2 per liter; the average C content of the harvested biomass is 50.5%; 1 tonne of C is equivalent to 3.67 tonnes of CO2; 1 kg of diesel is equivalent to 42.71 MJ, and its density is 0.85 kg L−1; the energy to CO2 conversion factor, corresponding to the Spanish electricity mix of 2019, is 241 g of CO2 released per kWh generated.
4. Discussion
4.1. Plant Growth and Biomass Production
4.2. Physical–Chemical Characteristics of Soil and Biomass and CO2 Sequestration
4.3. Economic, Energy and CO2 Emission Balances
5. Conclusions
- The biomass yield under a Mediterranean climate for short-rotation coppicing crops of Ulmus pumila, Robinia pseudoacacia and Populus × euroamericana “AF2” ranges from less than 5 to more than 20 t ha−1 year−1 (on average 12–14 t ha−1 year−1), depending on irrigation and soil quality;
- At least 800 mm of water (rainfall + irrigation) and about 60 kg ha−1 of N (fertilization) are required annually;
- This production system not only produces high-quality woody biomass, but, after a cycle of 15–17 years, on average it would also be able to:
- -
- Generate 205–237 GJ ha−1 year−1 net (equivalent to 57–66 MWh ha−1 year−1, or to the replacement of 5647–6528 L ha−1 year−1 of diesel);
- -
- Offer a profit of about EUR 1.5 per euro invested;
- -
- Sequester in the 0–30 cm layer of mineral soil 1.5 kg of C per kilogram released into the atmosphere.
- After the first two rotations evaluated (5–8 years), the upper layer of the mineral soil (0–30 cm) compared to the original soil sequestered 0.36–0.83 t ha−1 year−1 of carbon (equivalent to 1.3–3.0 t ha−1 year−1 of CO2);
- Nutrient inputs and outputs should be taken into consideration by the plantation managers in order to offset the outputs contained in the harvested biomass, and to prevent any loss in soil fertility and productivity;
- Degraded soils could be improved if the crop is properly managed, and any cultivation practice that promotes growth, reduces irrigation, fertilization and the cost of harvesting, and increases the added value of the final marketed product will have a major impact on the economic, energy and CO2 emission balances;
- The use of N-fixing species, such as Robinia pseudoacacia, and mixed plantations with the other two species are important aspects to be considered in future plantations.
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- United Nations. In Proceedings of the Conference of the Parties on Its Twenty-First Session, Paris, France, 30 November–13 December 2015. Available online: http://unfccc.int/resource/docs/2015/cop21/eng/10a01.pdf (accessed on 29 July 2021).
- European Union. Directive (EU) 2018/2001 of the European Parliament and of the Council of 11 December 2018 on the Use of Energy from Renewable Sources. Available online: https://eur-lex.europa.eu/eli/dir/2018/2001/oj (accessed on 25 July 2021).
- REN21. Renewable. Global Status Report, Renewable Energy Policy Network for the 21st Century, France. Available online: https://www.ren21.net/wp-content/uploads/2019/05/GSR2017_Full-Report_English.pdf (accessed on 30 July 2021).
- European Commission. Communication from The Commission to The European Parliament, The Council, The European Economic and Social Committee and The Committee of The Regions Energy Roadmap 2050/* COM/2011/0885 final */. Available online: https://eur-lex.europa.eu/legal-content/EN/ALL/?uri=celex:52011DC0885 (accessed on 27 June 2021).
- International Energy Agency (IEA). World Energy Outlook. Available online: https://iea.blob.core.windows.net/assets/4a50d774-5e8c-457e-bcc9513357f9b2fb/World_Energy_Outlook_2017.pdf (accessed on 30 July 2021).
- Antczak, A.; Swierkosz, R.; Szeniawski, M.; Marchwicka, M.; Akus-Szylberg, F.; Przybysz, P.; Zawadzki, J. The comparison of acid and enzymatic hydrolysis of pulp obtained from poplar wood (Populus sp.) by the Kraft method. Drewno 2019, 63, 1–14. [Google Scholar] [CrossRef]
- Titus, B.D.; Brown, K.; Helmisaari, H.-S.; Vanguelova, E.; Stupak, I.; Evans, A.; Clarke, N.; Guidi, C.; Bruckman, V.J.; Varnagiryte-Kabasinskiene, I.; et al. Sustainable forest biomass: A review of current residue harvesting guidelines. Energy Sustain. Soc. 2021, 11, 1–32. [Google Scholar] [CrossRef]
- Małachowska, E.; Lipkiewicz, A.; Niemczyk, M.; Dubowik, M.; Boruszewski, P.; Przybysz, P. Influences of fiber and pulp properties on papermaking ability of cellulosic pulps produced from alternative fibrous raw materials. J. Nat. Fibers 2019, 18, 1–11. [Google Scholar] [CrossRef]
- BIC. Strategic innovation and research agenda (SIRA) Bio-based and renewable industries for development and growth in Europe. In A Public-Private Partnership on Bio-Based Industries; Biobased Industries Consortium, Ed.; Biobased Industries Consortium: Brussels, Belgium, 2013; Available online: https://ec.europa.eu/research/participants/data/ref/h2020/other/legal_basis/jtis/bbi/bbi-sira_en.pdf (accessed on 30 July 2021).
- Bogdanski, A.; Dubois, O.; Jamieson, C.; Krell, R. Making Integrated Food-Energy Systems Work for People and Climate; FAO: Rome, Italy, 2010; p. 116. Available online: http://www.fao.org/docrep/013/i2044e/i2044e.pdf (accessed on 27 June 2021).
- Fernández, M.; Alaejos, J.; Andivia, E.; Vázquez-Piqué, J.; Ruiz, F.; López, F.; Tapias, R. Eucalyptus x urograndis biomass production for energy purposes exposed to a Mediterranean climate under different irrigation and fertilisation regimes. Biomass Bioenergy 2018, 111, 22–30. [Google Scholar] [CrossRef]
- Fernández, M.; Alaejos, J.; Andivia, E.; Madejón, P.; Díaz, M.; Tapias, R. Short rotation coppice of leguminous tree Leucaena spp. improves soil fertility while producing high biomass yields in Mediterranean environment. Ind. Crop Prod. 2020, 157, 112911. [Google Scholar] [CrossRef]
- Riffell, S.; Verschuyl, J.; Miller, D.; Wigley, T.B. Biofuel harvests, coarse woody debris, and biodiversity—A meta-analysis. For. Ecol. Manag. 2011, 261, 878–887. [Google Scholar] [CrossRef]
- Bouget, C.; Lassauce, A.; Jonsell, M. Effects of fuelwood harvesting on biodiversity—A review focused on the situation in Europe. Can. J. For. Res. 2012, 42, 1421–1432. [Google Scholar] [CrossRef]
- Miettinen, J.; Ollikainen, M.; Nieminen, T.M.; Ukonmaanaho, L.; Laurén, A.; Hynynen, J.; Lehtonen, M.; Valsta, L. Whole-tree harvesting with stump removal versus stem-only harvesting in peatlands when water quality, biodiversity conservation and climate change mitigation matter. For. Policy Econ. 2013, 47, 25–35. [Google Scholar] [CrossRef]
- Bessaad, A.; Bilger, I.; Korboulewsky, N. Assessing Biomass Removal and Woody Debris in Whole-Tree Harvesting System: Are the Recommended Levels of Residues Ensured? Forests 2021, 12, 807. [Google Scholar] [CrossRef]
- Srirangan, K.; Akawi, L.; Moo-Young, M.; Chou, C.P. Towards sustainable production of clean energy carriers from biomass resources. Appl. Energy 2012, 100, 172–186. [Google Scholar] [CrossRef]
- Sixto, H.; Cañellas, I.; van Arendonk, J.; Ciria, P.; Camps, F.; Sánchez, M.; Sánchez-González, M. Growth potential of different species and genotypes for biomass production in short rotation in Mediterranean environments. For. Ecol. Manag. 2015, 354, 291–299. [Google Scholar] [CrossRef]
- Oliveira, N.; del Rio, M.; Forrester, D.; Rodríguez-Soalleiro, R.; Pérez-Cruzado, C.; Canellas, I.; Sixto, H. Mixed short rotation plantations of Populus alba and Robinia pseudoacacia for biomass yield. For. Ecol. Manag. 2018, 410, 48–55. [Google Scholar] [CrossRef]
- Seserman, D.-M.; Pohle, I.; Veste, M.; Freese, D. Simulating Climate Change Impacts on Hybrid-Poplar and Black Locust Short Rotation Coppices. Forests 2018, 9, 419. [Google Scholar] [CrossRef] [Green Version]
- Szabolcs, K.; Varga, D. Physical and mechanical properties of wood from invasive tree species. Maderas Cienc. Tecnol. 2020, 23. [Google Scholar] [CrossRef]
- Parrotta, J. Productivity, nutrient cycling, and succession in single-and mixed-species plantations of Casuarina equisetifolia, Eucalyptus robusta, and Leucaena leucocephala in Puerto Rico. For. Ecol. Manag. 1999, 124, 45–77. [Google Scholar] [CrossRef]
- Durán Zuazo, V.H.; Jiménez Bocanegra, J.A.; Perea Torres, F.; Rodríguez Pleguezuelo, C.R.; Francia Martínez, J.R. Biomass yield potential of Paulownia trees in a semi-arid Mediterranean environment (S Spain). Int. J. Renew. Energy Res. 2013, 3, 789–793. Available online: https://ijrer.org/ijrer/index.php/ijrer/article/view/844 (accessed on 30 August 2021).
- Lauri, P.; Havlík, P.; Kindermann, G.; Forsell, N.; Böttcher, H.; Obersteiner, M. Woody biomass energy potential in 2050. Energy Policy 2014, 66, 19–31. [Google Scholar] [CrossRef]
- Fischer, G.; Prieler, S.; van Velthuizen, H.; Berndes, G.; Faaij, A.; Londo, M.; de Wit, M. Biofuel production potentials in Europe: Sustainable use of cultivated land and pastures, Part II: Land use scenarios. Biomass Bioenergy 2010, 34, 173–187. [Google Scholar] [CrossRef] [Green Version]
- Food and Agriculture Organization of United Nations (FAO). Combating land degradation for food security and provision of soil ecosystem services in Europe and Central Asia. In Proceedings of the 39th Session of the European Commission on Agriculture, Budapest, Hungary, 22–23 September 2015; Available online: http://www.fao.org/europe/commissions/eca/eca-39/en/ (accessed on 27 June 2021).
- Zielenkiewicz, T.; Szadkowski, J.; Drożdżek, M.; Zielenkiewicz, A.; Kłosińska, T.; Antczak, A.; Zawadzki, J.; Gawron, J. Application of X-ray fluorescence technique for determination of heavy metals uptake by different species of poplar. Drewno 2016, 59, 113–126. [Google Scholar] [CrossRef]
- Geyer, W. Biomass production in the Central Great Plains USA under various coppice regimes. Biomass Bioenergy 2006, 30, 778–783. [Google Scholar] [CrossRef]
- O’Neill, M.K.; Shock, C.C.; Lombard, K.A.; Heyduck, R.F.; Feibert, E.B.G.; Smeal, D.; Arnold, R.N. Hybrid poplar (Populus ssp.) selections for arid and semi-arid intermountain regions of the western United States. Agrofor. Syst. 2010, 79, 409–418. [Google Scholar] [CrossRef]
- Pérez, I.; Pérez, J.; Carrasco, J.; Ciria, P. Siberian elm responses to different culture conditions under short rotation forestry in Mediterranean areas. Turk. J. Agric. For. 2014, 38, 652–662. [Google Scholar] [CrossRef]
- Vanbeveren, S.P.; De Francesco, F.; Ceulemans, R.; Spinelli, R. Productivity of mechanized whip harvesting with the Stemster MkIII in a short-rotation coppice established on farmland. Biomass Bioenergy 2018, 108, 323–329. [Google Scholar] [CrossRef]
- García, I.P. Evaluación de Ulmus pumila L. y Populus spp. Como Cultivos Energéticos en Corta Rotación. Ph.D. Thesis, Polytechnic University of Madrid, Madrid, Spain, 2016. [Google Scholar] [CrossRef]
- Sanz Elorza, M.; Dana Sánchez, E.D.; Sobrino Vesperinas, E. Atlas de las plantas alóctonas invasoras en España. In Dirección General Para la Biodiversidad; Ministerio de Medio Ambiente: Madrid, Spain, 2004; p. 378. Available online: https://www.miteco.gob.es/es/biodiversidad/temas/inventarios-nacionales/inventario-especies-terrestres/inventario-nacional-de-biodiversidad/ieet_flora_vasc_aloct_invas.aspx (accessed on 18 August 2021).
- Marron, N.; Gana, C.; Gerant, D.; Maillard, P.; Priault, P.; Epron, D. Estimating symbiotic N2 fixation in Robinia pseudoacacia. J. Plant Nutr. Soil Sci. 2018, 181, 296–304. [Google Scholar] [CrossRef] [Green Version]
- Rédei, K.; Osváth-Bujtás, Z.; Veperdi, I. Black locust (Robinia pseudoacacia L.). Improvement in Hungary: A review. Acta Silv. Lign. Hung. 2008, 4, 127–132. [Google Scholar]
- Rédei, K.; Keserű, Z.; Csiha, I.; Rásó, J.; Végh, K.; Antal, B. Juvenile Growth and Morphological Traits of Micropropagated Black Locust (Robinia pseudoacacia, L.) Clones under Arid Site Conditions. Acta Silv. Lignaria Hung. 2013, 9, 35–42. [Google Scholar] [CrossRef] [Green Version]
- Dini-Papanastasi, O. Effects of clonal selection on biomass production and quality in Robinia pseudoacacia var. monophylla Carr. For. Ecol. Manag. 2008, 256, 849–854. [Google Scholar] [CrossRef]
- Kreiser, M.; Giblin, C.; Murphy, R.; Fiesel, P.; Braun, L.; Johnson, G.; Wyse, D.; Cohen, J.D. Conversion of Indole-3-Butyric Acid to Indole-3-Acetic Acid in Shoot Tissue of Hazelnut (Corylus) and Elm (Ulmus). J. Plant Growth Regul. 2016, 35, 710–721. [Google Scholar] [CrossRef]
- Vanguelova, E.; Pitman, R. Impacts of short rotation forestry on soil sustainability. In Short Rotation Forestry: Review of Growth and Environmental Impacts, Forest Research Monograph, Forest Research; McKay, H., Ed.; Forest Research Alice Holt Lodge: Surrey, UK, 2011; p. 37. ISBN 978-0-85538-827-0. [Google Scholar]
- Augusto, L.; Achat, D.L.; Bakker, M.R.; Bernier, F.; Bert, D.; Danjon, F.; Khlifa, R.; Meredieu, C.; Trichet, P. Biomass and nutrients in tree root systems-sustainable harvesting of an intensively managed Pinus pinaster (Ait.) planted forest. GCB Bioenergy 2014, 7, 231–243. [Google Scholar] [CrossRef]
- Madejón, P.; Alaejos, J.; García-Álbala, J.; Fernández, M.; Madejón, E. Three-year study of fast-growing trees in degraded soils amended with composts: Effects on soil fertility and productivity. J. Environ. Manag. 2016, 169, 18–26. [Google Scholar] [CrossRef]
- Grünewald, H.; Böhm, C.; Quinkenstein, A.; Grundmann, P.; Eberts, J.; Von Wühlisch, G. Robinia pseudoacacia L.: A Lesser known tree species for biomass production. BioEnergy Res. 2009, 2, 123–133. [Google Scholar] [CrossRef]
- Giuntoli, J.; Agostini, A.; Edwards, R.; Marelli, L. Solid and gaseous bioenergy pathways: Input values and GHG emissions. In Calculated According to the Methodology Set in COM (2016) Version 2, EUR 27215 EN; Publications Office of the European Union: Luxembourg, 2017. [Google Scholar] [CrossRef]
- Forrester, D.I.; Bauhus, J.; Cowie, A.; Vanclay, J. Mixed-species plantations of Eucalyptus with nitrogen-fixing trees: A review. For. Ecol. Manag. 2006, 233, 211–230. [Google Scholar] [CrossRef] [Green Version]
- Bouillet, J.; Laclau, J.; Gonçalves, J.L.M.; Moreira, M.Z.; Trivelin, P.; Jourdan, C.; Silva, E.; Piccolo, M.D.C.; Tsai, S.; Galiana, A. Mixed-species plantations of Acacia mangium and Eucalyptus grandis in Brazil. For. Ecol. Manag. 2008, 255, 3918–3930. [Google Scholar] [CrossRef]
- Moshki, A.; Lamersdorf, N.P. Symbiotic nitrogen fixation in black locust (Robinia pseudoacacia L.) seedlings from four seed sources. J. For. Res. 2011, 22, 689–692. [Google Scholar] [CrossRef]
- Verlinden, M.; Broeckx, L.; Bulcke, J.V.D.; Van Acker, J.; Ceulemans, R. Comparative study of biomass determinants of 12 poplar (Populus) genotypes in a high-density short-rotation culture. For. Ecol. Manag. 2013, 307, 101–111. [Google Scholar] [CrossRef]
- Magar, L.B.; Khadka, S.; Joshi, J.R.R.; Pokharel, U.; Rana, N.; Thapa, P.; Sharma, K.R.S.R.; Khadka, U.; Marasini, B.P.; Parajuli, N. Total Biomass Carbon Sequestration Ability Under the Changing Climatic Condition by Paulownia tomentosa Steud. Int. J. Appl. Sci. Biotechnol. 2018, 6, 220–226. [Google Scholar] [CrossRef]
- Nilsen, E.T.; Huebner, C.D.; Carr, D.E.; Bao, Z. Interaction between Ailanthus altissima and Native Robinia pseudoacacia in Early Succession: Implications for Forest Management. Forests 2018, 9, 221. [Google Scholar] [CrossRef] [Green Version]
- UNE. Solid Biofuels-Fuel Specifications and Classes-Part 2: Graded Wood Pellets (ISO 17225-2:2014); UNE-EN ISO 17225-2; ISO: Geneva, Switzerland, 2014; Available online: https://www.une.org/encuentra-tu-norma/busca-tu-norma/norma/?c=N0053748 (accessed on 8 August 2021).
- TRAGSA (Empresa de Transformación Agraria, S.A.). Spain, Tarifas TRAGSA. Available online: https://www.tragsa.es/es/grupo-tragsa/regimen-juridico/tarifas/Paginas/default.aspx (accessed on 27 July 2021).
- ENCE (ENCE, Energía y Celulosa, S.A.). Spain, Tarifa ENCE. Available online: https://compramosmadera.es/images/tarifasPDF/20210301_TARIFAS_MADERA_WEB_Mar2021_v1.pdf (accessed on 27 July 2021).
- Audsley, E. Harmonisation of environmental life cycle assessment for agriculture, final report. In Concerted Action AIR3-CT94; European Commission, Directorate-General VI Agriculture: Brussels, Belgium, 2003; p. 129. Available online: https://cordis.europa.eu/project/id/AIR32028 (accessed on 27 June 2021).
- Boto Fidalgo, J.A.; Pastrana Santamarta, P.; Suárez de Cepeda Martínez, M. Consumos Energéticos en Las Operaciones Agrícolas en España; IDAE, Instituto Para la Diversificación y Ahorro de la Energía, Ministerio de Industria, Turismo y Comercio: Madrid, Spain, 2005; p. 74. Available online: https://www.idae.es/uploads/documentos/documentos_10255_Consumos_energeticos_operaciones_agricolas_Espana_05_d94c1676.pdf (accessed on 30 July 2021).
- López Martínez, J.M.; Sánchez Alejo, J. Energy Consumption and Emissions Associated with Transportation by Car and Truck. Monografías EnerTrans. 2009. n° 12. Available online: https://www.investigacion-ffe.es/documentos/enertrans/EnerTrans_Transporte_coche_y_cami%C3%B3n.pdf (accessed on 30 July 2021).
- López Martínez, J.M.; Sánchez Alejo, J.; Mora Sotomayor, A. Energy Consumption and Emissions Associated with Construction and Maintenance of Vehicles. Monografías EnerTrans. 2009. n° 11. Available online: https://www.investigacion-ffe.es/documentos/enertrans/EnerTrans_Construcci%C3%B3n_Vehiculos.pdf (accessed on 30 July 2021).
- Camacho, E.; Rodríguez, J.A.; Montesinos, P.; Carrillo, T. Ahorro de energía en el riego. Agricultura 2010, 930, 454–457. Available online: https://www.mapa.gob.es/ministerio/pags/biblioteca/revistas/pdf_Agri%5CAgri_2010_930_completa.pdf (accessed on 20 August 2021).
- Manzone, M.; Airoldi, G.; Balsari, P. Energetic and economic evaluation of a poplar cultivation for the biomass production in Italy. Biomass Bioenergy 2009, 33, 1258–1264. [Google Scholar] [CrossRef]
- Di Nasso, O.N.N.; Guidi, W.; Ragaglini, G.; Tozzini, C.; Bonari, E. Biomass production and energy balance of a 12-year-old short-rotation coppice poplar stand under different cutting cycles. GCB Bioenergy 2010, 2, 89–97. [Google Scholar] [CrossRef]
- De Wit, M.; Faaij, A. European biomass resource potential and costs. Biomass Bioenergy 2010, 34, 188–202. [Google Scholar] [CrossRef]
- Dillen, S.; Djomo, S.; Al Afas, N.; Vanbeveren, S.; Ceulemans, R.J. Biomass yield and energy balance of a short-rotation poplar coppice with multiple clones on degraded land during 16 years. Biomass Bioenergy 2013, 56, 157–165. [Google Scholar] [CrossRef] [Green Version]
- Schweier, J.; Becker, G. Economics of poplar short rotation coppice plantations on marginal land in Germany. Biomass Bioenergy 2013, 59, 494–502. [Google Scholar] [CrossRef]
- Miguel, G.S.; Corona, B.; Ruiz, D.; Landholm, D.; Laina, R.; Tolosana, E.; Sixto, H.; Canellas, I. Environmental, energy and economic analysis of a biomass supply chain based on a poplar short rotation coppice in Spain. J. Clean. Prod. 2015, 94, 93–101. [Google Scholar] [CrossRef]
- Vanbeveren, S.P.; Spinelli, R.; Eisenbies, M.; Schweier, J.; Mola-Yudego, B.; Magagnotti, N.; Acuna, M.; Dimitriou, I.; Ceulemans, R. Mechanised harvesting of short-rotation coppices. Renew. Sustain. Energy Rev. 2017, 76, 90–104. [Google Scholar] [CrossRef]
- McFarquhar, M. Modeling Group-level repeated measurements of neuroimaging data using the univariate general linear model. Front. Neurosci. 2019, 13, 352. [Google Scholar] [CrossRef]
- Römheld, V. Diagnosis of deficiency and toxicity of nutrients. In Mineral Nutrition of Higher Plants, 3rd ed.; Marschner, P., Ed.; Elsevier Ltd.: Amsterdam, The Netherlands, 2012; pp. 299–312. [Google Scholar] [CrossRef]
- UNE. UNE-EN ISO 17225-4: Solid Biofuels-Fuel Specifications and Classes-Part 4: Graded Wood Chips (ISO 17225-4:2014). Available online: https://www.une.org/encuentra-tu-norma/busca-tu-norma/norma/?c=N0053750 (accessed on 8 August 2021).
- Montero, G.; Ruiz-Peinado, R.; Muñoz, M. Producción de biomasa y fijación de CO2 por los bosques españoles. In Instituto Nacional de Investigación y Tecnología Agraria y Alimentaria; Ministerio de Educación y Ciencia: Madrid, Spain, 2005; p. 270. Available online: http://libros.inia.es/libros/product_info.php?products_id=64 (accessed on 10 August 2021).
- Mokany, K.; Raison, R.J.; Prokushkin, A.S. Critical analysis of root: Shoot ratios in terrestrial biomes. Glob. Chang. Biol. 2005, 12, 84–96. [Google Scholar] [CrossRef]
- Luo, Y.; Wang, X.; Zhang, X.; Booth, T.H.; Lu, F. Root:shoot ratios across China’s forests: Forest type and climatic effects. For. Ecol. Manag. 2012, 269, 19–25. [Google Scholar] [CrossRef]
- Pérez, S.; Renedo, C.; Ortiz, A.; Mañana, M.; Delgado, F.; Tejedor, C. Energetic density of different forest species of energy crops in Cantabria (Spain). Biomass Bioenergy 2011, 35, 4657–4664. [Google Scholar] [CrossRef]
- Pérez-Cruzado, C.; Merino, A.; Rodríguez-Soalleiro, R. A management tool for estimating bioenergy production and carbon sequestration in Eucalyptus globulus and Eucalyptus nitens grown as short rotation woody crops in north-west Spain. Biomass Bioenergy 2011, 35, 2839–2851. [Google Scholar] [CrossRef]
- Stolarski, M.J.; Krzyżaniak, M.; Szczukowski, S.; Tworkowski, J.; Bieniek, A. Dendromass derived from agricultural land as energy feedstock. Pol. J. Environ. Stud. 2013, 22, 511–520. [Google Scholar]
- Berdón, J.B.; Calvo, A.J.M.; Barroso, L.R.; Alcobendas, A.I.P.; Cortés, J.G. Study of Paulownia’s Biomass Production in Mérida (Badajoz), Southwestern Spain. Environ. Ecol. Res. 2017, 5, 521–527. [Google Scholar] [CrossRef] [Green Version]
- Paris, P.; Mareschi, L.; Sabatti, M.; Pisanelli, A.; Ecosse, A.; Nardin, F.; Scarascia-Mugnozza, G. Comparing hybrid Populus clones for SRF across northern Italy after two biennial rotations: Survival, growth and yield. Biomass Bioenergy 2011, 35, 1524–1532. [Google Scholar] [CrossRef]
- Navarro, A.; Portillo-Estrada, M.; Arriga, N.; Vanbeveren, S.P.P.; Ceulemans, R. Genotypic variation in transpiration of coppiced poplar during the third rotation of a short-rotation bio-energy culture. GCB Bioenergy 2018, 10, 592–607. [Google Scholar] [CrossRef] [PubMed]
- Bloemen, J.; Fichot, R.; Horemans, J.A.; Broeckx, L.S.; Verlinden, M.; Zenone, T.; Ceulemans, R.J. Water use of a multigenotype poplar short-rotation coppice from tree to stand scale. GCB Bioenergy 2016, 9, 370–384. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bautista, I.; Mauri, P.V.; Plaza, A.; Ruiz-Fernández, J.; Amorós, M.C.; Sanz, M.; Curt, M.D. Vegetative propagation of Ulmus pumila L. by stem cuttings with a view to the development of bred lines for woody biomass plantations. In Proceedings of the 25th European Biomass Conference and Exhibition, Session 1DV.2.6, Stockholm, Sweden, 12–15 June 2017; ETA-Florence Renewable Energies: Firenze, Italy, 2017; pp. 253–257. Available online: http://www.etaflorence.it/proceedings/?detail=13655 (accessed on 8 August 2021). [CrossRef]
- Amorós, M.D.L.C.; Mauri, P.V.; Curt, M.D. The influence of tree management practices on phenological growth stages of Ulmus pumila L. (Siberian elm). Ann. Appl. Biol. 2021, 179, 1–14. [Google Scholar] [CrossRef]
- Qin, J.; Xi, W.; Rahmlow, A.; Kong, H.; Zhang, Z.; Shangguan, Z. Effects of forest plantation types on leaf traits of Ulmus pumila and Robinia pseudoacacia on the Loess Plateau, China. Ecol. Eng. 2016, 97, 416–425. [Google Scholar] [CrossRef]
- Bessaad, A.; Terreaux, J.-P.; Korboulewsky, N. Assessing the land expectation value of even-aged vs coppice-with-standards stand management and long-term effects of whole-tree harvesting on forest productivity and profitability. Ann. For. Sci. 2021, 78, 1–15. [Google Scholar] [CrossRef]
- Parrotta, J.A. Leucaena leucocephala (Lam.) de Wit: Leucaena, tantan. Res. Note SO-ITFSM-USDA Forest Service; Southern Forest Experiment Estation: New Orleans, LA, USA, 1992; p. 8. Available online: https://www.fs.usda.gov/treesearch/pubs/50724 (accessed on 8 August 2021).
- Vanbeveren, S.; Schweier, J.; Berhongaray, G.; Ceulemans, R.J. Operational short rotation woody crop plantations: Manual or mechanised harvesting? Biomass Bioenergy 2015, 72, 8–18. [Google Scholar] [CrossRef]
- Jug, A.; Hofmann-Schielle, C.; Makeschin, F.; Rehfuess, K. Short-rotation plantations of balsam poplars, aspen and willows on former arable land in the Federal Republic of Germany. II. Nutritional status and bioelement export by harvested shoot axes. For. Ecol. Manag. 1999, 121, 67–83. [Google Scholar] [CrossRef]
- Romanyà, J.; Rovira, P.; Vallejo, R. Análisis del carbono en los suelos agrícolas de España. Aspectos relevantes en relación a la reconversión a la agricultura ecológica en el ámbito mediterráneo. Ecosistemas 2007, 16, 50–57. Available online: https://www.revistaecosistemas.net/index.php/ecosistemas/article/view/138 (accessed on 27 August 2021).
- González, I.; Sixto, H.; Rodríguez-Soalleiro, R.; Oliveira, N. Nutrient Contribution of Litterfall in a Short Rotation Plantation of Pure or Mixed Plots of Populus alba L. and Robinia pseudoacacia L. Forests 2020, 11, 1133. [Google Scholar] [CrossRef]
- Buerkert, A.; Joergensen, R.G.; Ludwig, B.; Schlecht, E. Nutrient and Carbon Fluxes in Terrestrial Agro-Ecosystems. In Mineral Nutrition of Higher Plants, 3rd ed.; Marschner, P., Ed.; Elsevier Ltd.: Amsterdam, The Netherlands, 2012; pp. 473–482. [Google Scholar] [CrossRef]
- Ferré, C.; Comolli, R. Comparison of soil CO2 emissions between short-rotation coppice poplar stands and arable lands. Iforest 2018, 11, 199–205. [Google Scholar] [CrossRef] [Green Version]
- Campos, R.; Pires, G.F.; Costa, M.H. Soil Carbon Sequestration in Rainfed and Irrigated Production Systems in a New Brazilian Agricultural Frontier. Agriculture 2020, 10, 156. [Google Scholar] [CrossRef]
- Berhongaray, G.; Verlinden, M.; Broeckx, L.S.; Janssens, I.; Ceulemans, R. Soil carbon and belowground carbon balance of a short-rotation coppice: Assessments from three different approaches. GCB Bioenergy 2016, 9, 299–313. [Google Scholar] [CrossRef] [Green Version]
- Andivia, E.; Rolo, V.; Jonard, M.; Formánek, P.; Ponette, Q. Tree species identity mediates mechanisms of top soil carbon sequestration in a Norway spruce and European beech mixed forest. Ann. For. Sci. 2016, 73, 437–447. [Google Scholar] [CrossRef] [Green Version]
- Jha, P.; Mohapatra, K.P. Leaf litterfall, fine root production and turnover in four major tree species of the semi-arid region of India. Plant Soil 2009, 326, 481–491. [Google Scholar] [CrossRef]
- George, E.; Horst, W.J.; Neumann, E. Adaptation of Plants to Adverse Chemical Soil Conditions. In Mineral Nutrition of Higher Plants, 3rd ed.; Marschner, P., Ed.; Elsevier Ltd.: Amsterdam, The Netherlands, 2012; pp. 409–472. [Google Scholar] [CrossRef]
- Li, K.; Han, X.; Ni, R.; Shi, G.; De-Miguel, S.; Li, C.; Shen, W.; Zhang, Y.; Zhang, X. Impact of Robinia pseudoacacia stand conversion on soil properties and bacterial community composition in Mount Tai, China. For. Ecosyst. 2021, 8, 1–12. [Google Scholar] [CrossRef]
- Laker, M.C.; Nortjé, G.P. Review of existing knowledge on soil crusting in South Africa. Adv. Agron. 2019, 115, 189–242. [Google Scholar] [CrossRef]
- Rice, S.K.; Westerman, B.; Federici, R. Impacts of the exotic, nitrogen-fixing black locust (Robinia pseudoacacia) on nitrogen cycling in a pine-oak ecosystem. Plant Ecol. 2004, 174, 97–107. [Google Scholar] [CrossRef]
- Castro-Díez, P.; Fierro-Brunnenmeister, N.; González-Muñoz, N.; Gallardo, A. Effects of exotic and native tree leaf litter on soil properties of two contrasting sites in the Iberian Peninsula. Plant Soil 2011, 350, 179–191. [Google Scholar] [CrossRef]
- Carmona, R.; Nuñez, T.; Alonso, M. Biomass yield and quality of an energy dedicated crop of poplar (Populus spp.) clones in the Mediterranean zone of Chile. Biomass Bioenergy 2015, 74, 96–102. [Google Scholar] [CrossRef]
- European Pellet Council (EPC). Available online: https://epc.bioenergyeurope.org/about-pellets/pellets-basics/pellet-standards (accessed on 27 August 2021).
- Morhart, C.; Sheppard, J.; Spiecker, H. Above Ground Leafless Woody Biomass and Nutrient Content within Different Compartments of a P. maximowicii × P. trichocarpa Poplar Clone. Forests 2013, 4, 471–487. [Google Scholar] [CrossRef] [Green Version]
- Fernández, M.; García-Albalá, J.; Andivia, E.; Alaejos, J.; Tapias, R.; Menéndez, J.; Martín, R.T. Sickle bush (Dichrostachys cinerea L.) field performance and physical–chemical property assessment for energy purposes. Biomass Bioenergy 2015, 81, 483–489. [Google Scholar] [CrossRef]
- Vasiliki, K.; Ioannis, B. Utilization perspectives of wood and bark of the invasive species of Ailanthus and Acacia in the production of pellets. In Proceedings of the International Forestry and Environment Symposium, Trabzon, Turkey, 7–10 November 2017; Available online: http://users.auth.gr/jbarb/Publications/IFES2017-209_Utilization.pdf (accessed on 8 August 2021).
- Nunes, L.; Rodrigues, A.; Loureiro, L.; Sá, L.; Matias, J. Energy Recovery from Invasive Species: Creation of Value Chains to Promote Control and Eradication. Recycling 2021, 6, 21. [Google Scholar] [CrossRef]
- Holm, J.K.; Henriksen, U.B.; Hustad, J.E.; Sørensen, L.H. Toward an Understanding of Controlling Parameters in Softwood and Hardwood Pellets Production. Energy Fuels 2006, 20, 2686–2694. [Google Scholar] [CrossRef]
- Stelte, W.; Holm, J.K.; Sanadi, A.R.; Barsberg, S.; Ahrenfeldt, J.; Henriksen, U.B. A study of bonding and failure mechanisms in fuel pellets from different biomass resources. Biomass Bioenergy 2011, 35, 910–918. [Google Scholar] [CrossRef] [Green Version]
- TRADERSbiomass. 2021. Available online: https://tradersbiomass.com/online-biomass-exchangetrading-with-tradersbiomass/trading-with-tradersbiomass/ (accessed on 27 August 2021).
- Jenkins, B.M.; Baxter, L.L.; Miles, T.R., Jr.; Miles, T.R. Combustion properties of biomass. Fuel Process. Technol. 1998, 54, 17–46. [Google Scholar] [CrossRef]
- Verlinden, M.; Broeckx, L.; Zona, D.; Berhongaray, G.; De Groote, T.; Camino-Serrano, M.; Janssens, I.; Ceulemans, R.J. Net ecosystem production and carbon balance of an SRC poplar plantation during its first rotation. Biomass Bioenergy 2013, 56, 412–422. [Google Scholar] [CrossRef] [Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Variable | Field Trials Mo1 and Mo2 | Field Trial Mo3 | Field Trial Ra4 |
|---|---|---|---|
| Time period (years) | 2011–2018 | 2015–2019 | 2015–2020 |
| Absolute maximum temperature (°C) | 44.8 | 44.8 | 40.6 |
| Warmest month’s average maximum temperature (°C) | 34.1 | 34.7 | 31.9 |
| Mean annual temperature (°C) | 13.9 | 14.1 | 16.8 |
| Coldest month’s average minimum temperature (°C) | −0.7 | −0.8 | 4.4 |
| Absolute minimum temperature (°C) | −13.7 | −8.4 | −4.0 |
| Annual precipitation (mm) | 319.8 | 332.8 | 483.6 |
| Range of annual precipitation | 214.2–476.3 | 214.2–461.9 | 236.6–646.2 |
| Summer precipitation from June to August (mm) | 31.0 | 22.1 | 7.6 |
| Vegetative period (>12.5 °C mean monthly temperature) (month) | 6.2 | 6.2 | 8.3 |
| Annual reference crop evapotranspiration, ETo (mm) | 1396.8 | 1399.9 | 1074.4 |
| Variable | Field Trials Mo1 and Mo2 | Field Trial Mo3 | Field Trial Ra4 | |
|---|---|---|---|---|
| Soil sampling depth (cm) | 0–30 | 0–30 | 0–15 | 15–30 |
| Exchangeable Sodium Percentage (%) | 3.9 (0.2) | 5.4 (0.1) | 9.1 (0.3) | 12.7 (0.1) |
| Organic Matter (%) | 2.93 (0.09) | 2.67 (0.03) | 1.22 (0.02) | 0.53 (0.05) |
| C/N ratio | 10.64 (0.46) | 9.16 (0.04) | 11.27 (0.04) | 13.74 (0.76) |
| Organic Carbon (%) | 1.51 (0.04) | 1.46 (0.03) | 0.71 (0.02) | 0.31 (0.03) |
| Active limestone (%) | 7.57 (0.31) | 3.31 (0.03) | 1.27 (0.05) | 1.71 (0.08) |
| N (%) | 0.16 (0.01) | 0.15 (0.01) | 0.06 (0.01) | 0.04 (0.01) |
| Total P (mg kg−1) (1) | 88.8 (2.5) | 78.4 (5.0) | 59.6 (2.0) | 42.9 (1.5) |
| Available K (meq/100 g) | 1.37 (0.09) | 1.40 (0.07) | 0.27 (0.01) | 0.21 (0.01) |
| Variable | Field Trial Mo1 | Field Trial Mo2 | Field Trial Mo3 | Field Trial Ra4 |
|---|---|---|---|---|
| Study period (years) | 2011–2018 | 2011–2018 | 2015–2019 | 2015–2020 |
| Number of taxa/treatments used | 11 taxa | 5 treatments | 4 taxa | 4 taxa |
| Taxa/treatments codes * | Adige; AF2; Oudenberg; I214; Raspalje; Pn; Pf; Aa; Pxh; Rp-H; Up. | Rp0-AF100; Rp25-AF75; Rp50-AF50; Rp75-AF25; Rp100-AF0. | Rp-H; Rp-G; AF2; Up. | Rp-H; Rp-G; AF2; Up. |
| Number of years per rotation | 4 | 4 | 5 | 3 |
| Rotations studied (number of years) | 2 (4 + 4) | 2 (4 + 4) | 1 (5) | 2 (3 + 3) |
| Experimental unit (ExpU): | ||||
| Length (m) | 9.6 | 9.6 | 24 | 6 |
| Number of lines | 3 | 3 | 4 | 3 |
| Number of plants per line | 16 | 16 | 30 | 6 |
| Space between lines (m) | 3.0 | 3.0 | 3.3 | 2.0 |
| Space between plants into a line (m) | 0.6 | 0.6 | 0.8 | 1.0 |
| Number of plants in the ExpU | 48 | 48 | 120 | 18 |
| Area of the ExpU (m2) | 86.4 | 86.4 | 316.8 | 36 |
| Plantation density (plants ha−1) | 5556 | 5556 | 3788 | 5000 |
| Number of plants measured per ExpU and year | 10 | 10 | 20 | 4 |
| Taxon/treatment replicates (number of ExpU) | 4 | 4 | 4 | 4 |
| Total number of plants in the plot | 2112 | 960 | 1920 | 288 |
| Taxon | a | b | R2 | n |
|---|---|---|---|---|
| Populus × euroamericana “Adige” | 0.043 | 2.7074 | 0.981 | 30 |
| Populus × euroamericana “AF2” | 0.034 | 2.7013 | 0.986 | 38 |
| Populus × euroamericana “Oudenberg” | 0.076 | 2.6321 | 0.978 | 30 |
| Populus × euroamericana “I214” | 0.051 | 2.690 | 0.891 | 30 |
| Populus × interamericana “Raspalje” | 0.05 | 2.747 | 0.981 | 30 |
| Populus nigra L. | 0.045 | 2.700 | 0.996 | 30 |
| Paulownia fortunei “UHU” (D < 32.3 mm) | 0.005 | 3.033 | 0.970 | 15 |
| Paulownia fortunei “UHU” (D > 32.3 mm) | (1) | (1) | 0.972 | 17 |
| Robinia pseudoacacia | 0.113 | 2.482 | 0.980 | 32 |
| Ulmus pumila | 0.065 | 2.612 | 0.978 | 32 |
| Ailanthus altissima | 0.009 | 3.042 | 0.980 | 30 |
| Platanus × hispanica | 0.073 | 2.630 | 0.989 | 30 |
| Cultivation Work/Material | Tractor | Cost | Fuel | Energy (9) | CO2 (9) | Efficiency |
|---|---|---|---|---|---|---|
| (kW) | (EUR ha−1) | (L h−1) | (MJ h−1) | (kg h−1) | (h ha−1) | |
| 50 cm deep ploughing (1) | 110 | 115 | 15.0 | 78.5 | 4.04 | 2.00 |
| Disk harrowing (1) | 74 | 45 | 8.7 | 61.5 | 3.10 | 1.40 |
| Rotary harrowing (1) | 74 | 49 | 8.7 | 61.5 | 3.10 | 1.50 |
| Mechanical fertilizing (broadcaster) (2) | 52 | 55 | 3.5 | 32.4 | 1.49 | 0.33 |
| Chemical weeding (sprayer) (2) | 52 | 54 | 3.5 | 32.4 | 1.49 | 0.33 |
| Mechanical planting (1) | 110 | 250 | 12.0 | 78.5 | 4.04 | 2.50 |
| Mechanical weeding (2) | 52 | 30 | 3.5 | 32.4 | 1.49 | 1.25 |
| Harvesting (cutting and field stocking) (2) | 74 | 90 | 8.0 | 61.5 | 3.10 | 1.70 |
| Harvesting (chipping and trailer loading) (1)(8) | 132 | 385 | 20.5 | 78.5 | 4.04 | 2.30 |
| Grubbing and land clearing (1)(4) | 132 | 475 | 20.5 | 78.5 | 4.04 | 2.50 |
| (EUR t−1) | (L km−1) | (MJ km−1) | (kg km−1) | (t payload−1) | ||
| Chip transportation (truck with trailer, 25 km) (3) | 257 | 4.5 | 0.425 | 1.15 | 0.06 | 22.0 |
| (EUR kg−1) | (MJ kg−1) | (kg kg−1) | (kg ha−1) | |||
| Fertilizer N/P/K (5) | 0.5 | 45/30/5 | 3.0/2.0/0.3 | 500/333.3 | ||
| Chelated fertilizer | 6.2 | 100 | 6.7 | |||
| (EUR L−1) | (MJ L−1) | (kg L−1) | (L ha−1 y−1) | |||
| Herbicide | 10.5 | 106 | 7.1 | 2.50 | ||
| (EUR ha−1) | (MJ kg−1) | (kg kg−1) | (kg ha−1 y−1) | |||
| Irrigation (material) (PE/PVC) (6) | 1490 | 78.6/57 | 5.3/3.8 | 523.13 | ||
| (EUR h−1) | (MJ m−3) | (kg m−3) | (m3 ha−1 y−1) | |||
| Irrigation (water pumping) (7) | 0.21 | 0.54 | 0.036 | 4000 | ||
| (EUR plant−1) | ||||||
| Plants | 0.35 |
| Variable | Field | Main Effect | Effect (1) | ||
|---|---|---|---|---|---|
| Trial | TX or TR | Year | SLY | Interactions | |
| ESP (%) | Mo1 | <0.001 | <0.001 | 0.133 | |
| Mo2 | 0.027 | <0.001 | 0.215 | ||
| Mo3 | 0.010 | <0.001 | 0.052 | ||
| Ra4 | <0.001 | <0.001 | <0.001 | ≥0.200 | |
| C/N ratio | Mo1 | 0.051 | <0.001 | 0.390 | |
| Mo2 | 0.010 | 0.001 | 0.076 | ||
| Mo3 | 0.414 | 0.002 | 0.638 | ||
| Ra4 | 0.016 | <0.001 | 0.910 | ≥0.076 | |
| Organic carbon (%) | Mo1 | 0.026 | <0.001 | 0.376 | |
| Mo2 | 0.132 | <0.001 | 0.748 | ||
| Mo3 | 0.519 | <0.001 | 0.781 | ||
| Ra4 | 0.907 | 0.010 | <0.001 | ≥0.289 | |
| Active limestone (%) | Mo1 | 0.220 | <0.001 | 0.750 | |
| Mo2 | 0.459 | <0.001 | 0.593 | ||
| Mo3 | 0.736 | <0.001 | 0.933 | ||
| Ra4 | 0.041 | 0.002 | 0.302 | ≥0.202 | |
| N (%) | Mo1 | 0.011 | 0.001 | 0.301 | |
| Mo2 | 0.005 | 0.004 | 0.100 | ||
| Mo3 | 0.487 | <0.001 | 0.752 | ||
| Ra4 | 0.003 | 0.009 | <0.001 | ≥0.054 | |
| Total P (mg kg−1) | Mo1 | 0.941 | <0.001 | 0.873 | |
| Mo2 | 0.829 | <0.001 | 0.785 | ||
| Mo3 | 0.626 | <0.001 | 0.962 | ||
| Ra4 | 0.004 | <0.001 | <0.001 | ≥0.051 | |
| Available K (meq/100 g) | Mo1 | 0.051 | <0.001 | 0.055 | |
| Mo2 | 0.068 | 0.045 | 0.068 | ||
| Mo3 | 0.789 | 0.040 | 0.959 | ||
| Ra4 | 0.292 | <0.001 | <0.001 | ≥0.063 | |
| Variable | Rotation (1) | Field Trial | |||
|---|---|---|---|---|---|
| Mo1 | Mo2 | Mo3 | Ra4 | ||
| ESP (%) | 1st | 5.1 * (+) | 5.3 * (+) | 6.8 * (+) | 14.1 * (+) |
| 2nd | 6.4 * (+) | 7.0 * (+) | 6.7 * (+) | 13.6 * (+) | |
| C/N ratio | 1st | 10.56 ns | 10.62 ns | 9.16 ns | 8.31 * (−) |
| 2nd | 9.49 * (−) | 9.48 * (−) | 10.35 * (+) | 8.59 * (−) | |
| Organic carbon (%) | 1st | 1.68 * (+) | 1.70 * (+) | 1.36 ns | 0.38 ns |
| 2nd | 1.71 * (+) | 1.70 * (+) | 1.59 * (+) | 0.57 (+) | |
| Active limestone (%) | 1st | 6.35 * (−) | 6.29 * (−) | 2.67 * (−) | 1.28 * (−) |
| 2nd | 4.91 * (−) | 4.85 * (−) | 2.88 * (−) | 1.30 * (−) | |
| N (%) | 1st | 0.16 ns | 0.16 ns | 0.15 ns | 0.05 ns |
| 2nd | 0.17 * (+) | 0.17 * (+) | 0.16 * (+) | 0.06 * (+) | |
| Total P (mg kg−1) | 1st | 83.8 * (−) | 83.4 * (−) | 79.4 * (−) | 35.0 * (−) |
| 2nd | 74.4 * (−) | 70.7 * (−) | 79.4 * (−) | 32.0 * (−) | |
| Available K (meq/100 g) | 1st | 1.42 ns | 1.57 * (+) | 1.54 ns | 0.31 * (+) |
| 2nd | 1.63 * (+) | 1.27 ns | 1.64 * (+) | 0.28 * (+) | |
| Variable | Populus × euroamericana “AF2” | Robinia pseudoacacia | Ulmus pumila | ||||||
|---|---|---|---|---|---|---|---|---|---|
| AGWB | Roots | Litterfall | AGWB | Roots | Litterfall | AGWB | Roots | Litterfall | |
| C (%) | 50.9 (1.1) | 48.8 (1.1) | 54.1 (0.9) | 50.7 (0.8) | 49.4 (0.7) | 53.0 (1.3) | 50.1 (0.8) | 48.5 (0.9) | 52.6 (2.0) |
| N (%) | 0.38 (0.04) a | 0.79 (0.05) | 1.10 (0.06) | 0.54 (0.03) b | 0.74 (0.17) | 1.22 (0.06) | 0.47 (0.07) ab | 0.80 (0.13) | 1.10 (0.05) |
| P (%) | 0.09 (0.01) | 0.09 (0.03) | 0.18 (0.03) | 0.06 (0.06) | 0.10 (0.07) | 0.11 (0.06) | 0.08 (0.04) | 0.08 (0.06) | 0.11 (0.05) |
| K (%) | 0.30 (0.06) | 0.28 (0.04) | 0.23 (0.05) | 0.31 (0.03) | 0.24 (0.04) | 0.22 (0.03) | 0.29 (0.03) | 0.24 (0.04) | 0.20 (0.04) |
| Ca (%) | 0.57 (0.10) b | 1.26 (0.11) | 2.99 (0.35) | 0.38 (0.02) a | 0.48 (0.05) | 2.41 (0.08) | 0.57 (0.11) b | 0.39 (0.08) | 2.87 (0.16) |
| Mg (%) | 0.13 (0.04) | 0.17 (0.05) | 0.37 (0.04) | 0.14 (0.05) | 0.17 (0.04) | 0.22 (0.04) | 0.11 (0.04) | 0.22 (0.04) | 0.33 (0.06) |
| S (%) | 0.06 (0.01) b | 0.08 (0.02) | 0.02 (0.01) | 0.04 (0.01) a | 0.08 (0.02) | 0.02 (0.01) | 0.04 (0.01) a | 0.10 (0.02) | 0.02 (0.01) |
| Fe (mg kg−1) | 78 (12) b | 78 (21) | 303 (23) | 58 (10) ab | 82 (12) | 301 (15) | 53 (9) a | 66 (11) | 272 (23) |
| Mn (mg kg−1) | 7 (1) | 19 (2) | 34 (2) | 6 (1) | 12 (2) | 25 (3) | 5 (2) | 13 (2) | 26 (3) |
| Zn (mg kg−1) | 40 (10) | 29 (8) | 77 (27) | 16 (5) | 28 (4) | 46 (20) | 25 (5) | 28 (4) | 45 (15) |
| Cl (mg kg−1) | 890 (40) b | 817 (77) | 3360 (220) | 744 (23) a | 812 (64) | 2966 (123) | 869 (40) b | 843 (65) | 3132(160) |
| Ash (%) | 2.05 (0.16) a | na | na | 2.04 (0.10) a | na | na | 2.67 (0.13) b | na | na |
| HHV (MJ kg−1) | 19.2 (0.2) | na | na | 19.3 (0.2) | na | na | 19.4 (0.2) | na | na |
| LHV (MJ kg−1) | 17.8 (0.3) | na | na | 18.0 (0.2) | na | na | 18.1 (0.3) | na | na |
| Bdp (kg m−3) (1) | 671 (13) | na | na | 651 (12) | na | na | 636 (14) | na | na |
| MDup (%) (1) | 97.7 (1.5) | na | na | 98.5 (1.7) | na | na | 97.4 (1.8) | na | na |
| Moisturep (%) (1) | 6.6 (0.5) | na | na | 6.8 (0.4) | na | na | 6.8 (0.5) | na | na |
| LHVp (MJ kg−1) (2) | 16.5 (0.2) | na | na | 16.6 (0.2) | na | na | 16.7 (0.2) | na | na |
| WdB (kg m−3) | 0.42 (0.05) a | na | na | 0.77 (0.07) b | na | na | 0.68 (0.06) b | na | na |
| Crop Assumption | Mo1 and Mo2 | Mo3 | Ra4 | |||
|---|---|---|---|---|---|---|
| Year per cycle | 16 | 17 | 15 | |||
| 1st rotation (years) | 4 | 5 | 3 | |||
| 2nd and following rotations (years per rotation) | 4 | 4 | 3 | |||
| Rotations per cycle | 4 | 4 | 5 | |||
| Frequency(depending on the rotation: the 1st or subsequent rotations) | ||||||
| 1st | 2nd to 4th | 1st | 2nd to 4th | 1st | 2nd to 5th | |
| 50 cm-deep ploughing | 1 | 0 | 1 | 0 | 1 | 0 |
| Disk harrowing | 1 | 0 | 1 | 0 | 1 | 0 |
| Rotary harrowing | 1 | 0 | 1 | 0 | 1 | 0 |
| Mechanical fertilizing (broadcaster) | 1 | 0 | 1 | 0 | 1 | 0 |
| Chemical weeding (sprayer) | 1 | 0 | 1 | 0 | 1 | 0 |
| Mechanical planting | 1 | 0 | 1 | 0 | 1 | 0 |
| Mechanical weeding | 1 | 0 | 1 | 0 | 1 | 0 |
| Harvesting (cutting with saw disc, field stocking) | 1 | 3 | 1 | 3 | 1 | 4 |
| Harvesting (chipping and trailer loading) | 1 | 3 | 1 | 3 | 1 | 4 |
| Grubbing and land clearing | 0 | 1 | 0 | 1 | 0 | 1 |
| Chip transportation (truck with trailer, 25 km) | 1 | 3 | 1 | 3 | 1 | 4 |
| Fertilizer N/P/K (mechanical fertilizing) | 1 | 0 | 1 | 0 | 1 | 0 |
| Fertilizer N/P/K (fertigation) | 1 × 4 | 3 × 4 | 1 × 5 | 3 × 4 | 1 × 3 | 4 × 3 |
| Chelated fertilizer | 1 × 4 | 3 × 4 | 1 × 5 | 3 × 4 | 1 × 3 | 4 × 3 |
| Herbicide | 1 | 0 | 1 | 0 | 1 | 0 |
| Irrigation (material) (PE/PVC) | 1 | 0 | 1 | 0 | 1 | 0 |
| Irrigation (water pumping) | 1 × 4 | 3 × 4 | 1 × 5 | 3 × 4 | 1 × 3 | 4 × 3 |
| Plants | 1 | 0 | 1 | 0 | 1 | 0 |
| Crop Assumption | Mo1 (Cycle = 16 Years) | Mo3 (Cycle = 17 Years) | Ra4 (Cycle = 15 Years) | |||
|---|---|---|---|---|---|---|
| Biomass yielded according to rotation | 1st | 2nd to 4th | 1st | 2nd to 4th | 1st | 2nd to 5th |
| AGWB (t ha−1) (1) | 42.40 | 151.92 | 51.35 | 151.92 | 31.80 | 174.36 |
| AGWB (t ha−1 year−1) (1) | 10.60 | 12.66 | 10.27 | 12.66 | 10.60 | 14.53 |
| Economic balance | Incomes | Costs | Incomes | Costs | Incomes | Costs |
| Gross incomes and costs (EUR ha−1) | 15,934 | 10,687 | 16,668 | 10,923 | 16,905 | 11,020 |
| Difference (Inc.–Cos.) (EUR ha−1) | 5247 | 5745 | 5885 | |||
| Ratio (Incomes/Costs) | 1.491 | 1.526 | 1.534 | |||
| Gross incomes and costs (EUR ha−1 year−1) | 996 | 668 | 980 | 643 | 1127 | 735 |
| Difference (Inc.–Cos.) (EUR ha−1 year−1) | 328 | 337 | 392 | |||
| Energy balance | Generated | Consumed | Generated | Consumed | Generated | Consumed |
| Energy (GJ ha−1) (2) | 3498 | 159 | 3659 | 166 | 3711 | 156 |
| Difference (Gen.–Con.) (GJ ha−1) | 3339 | 3493 | 3555 | |||
| Ratio (Generated/Consumed) | 22.00 | 22.04 | 23.79 | |||
| Energy (GJ ha−1 year−1) (1) | 218.6 | 10.0 | 215.2 | 9.8 | 247.4 | 10.4 |
| Difference (Gen.–Con.) (GJ ha−1 year−1) | 208.6 | 205.4 | 237.0 | |||
| CO2 emission balance | Fixed | Released | Fixed | Released | Fixed | Released |
| CO2 fixed and released (t ha−1) | 564.0 | 10.5 | 596.1 | 11.0 | 572.6 | 10.3 |
| Difference (Fix.–Rel.) (t ha−1) (3) | 553.5 | 585.1 | 562.3 | |||
| Difference (Fix.–Rel.) (t ha−1) (3) | 193.4 | 208.4 | 180.2 | |||
| Ratio (Fixed/Released) (3) | 53.71 | 54.19 | 55.59 | |||
| Ratio (Fixed/Released) (3) | 1.522 | 1.538 | 1.459 | |||
| CO2 emission balance (t ha−1 year−1) | 35.25 | 0.66 | 35.07 | 0.65 | 38.17 | 0.69 |
| Difference (Fix.–Rel.) (t ha−1 year−1) (3) | 34.59 | 34.42 | 37.48 | |||
| Difference (Fix.–Rel.) (t ha−1 year−1) (3) | 12.08 | 12.26 | 12.01 | |||
| Cost of energy (EUR GJ−1) (4) | 3.20 | 3.13 | 3.10 | |||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Alesso, S.P.; Tapias, R.; Alaejos, J.; Fernández, M. Biomass Yield and Economic, Energy and Carbon Balances of Ulmus pumila L., Robinia pseudoacacia L. and Populus × euroamericana (Dode) Guinier Short-Rotation Coppices on Degraded Lands under Mediterranean Climate. Forests 2021, 12, 1337. https://0-doi-org.brum.beds.ac.uk/10.3390/f12101337
Alesso SP, Tapias R, Alaejos J, Fernández M. Biomass Yield and Economic, Energy and Carbon Balances of Ulmus pumila L., Robinia pseudoacacia L. and Populus × euroamericana (Dode) Guinier Short-Rotation Coppices on Degraded Lands under Mediterranean Climate. Forests. 2021; 12(10):1337. https://0-doi-org.brum.beds.ac.uk/10.3390/f12101337
Chicago/Turabian StyleAlesso, Silvia Patricia, Raúl Tapias, Joaquín Alaejos, and Manuel Fernández. 2021. "Biomass Yield and Economic, Energy and Carbon Balances of Ulmus pumila L., Robinia pseudoacacia L. and Populus × euroamericana (Dode) Guinier Short-Rotation Coppices on Degraded Lands under Mediterranean Climate" Forests 12, no. 10: 1337. https://0-doi-org.brum.beds.ac.uk/10.3390/f12101337