
Phenotypic Diversity of Almond-Leaved Pear (Pyrus spinosa Forssk.) along Eastern Adriatic Coast

 , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Material and Studied Leaf Traits
2.2. Environmental Data
2.3. Population Diversity and Phenotypic Traits
2.4. Correlations between Geographic, Environmental, and Morphometric Data
2.5. Population Structure
3. Results
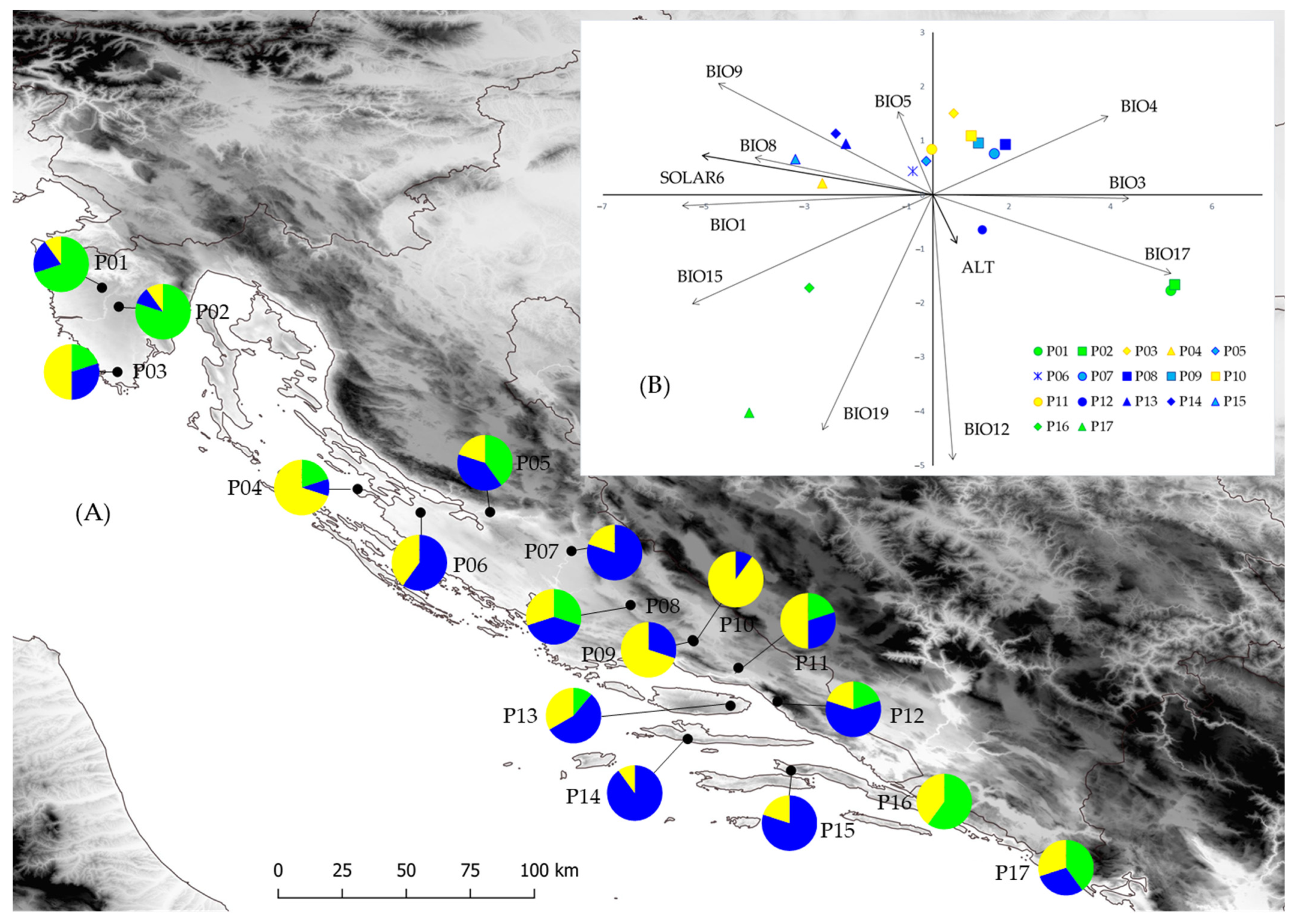
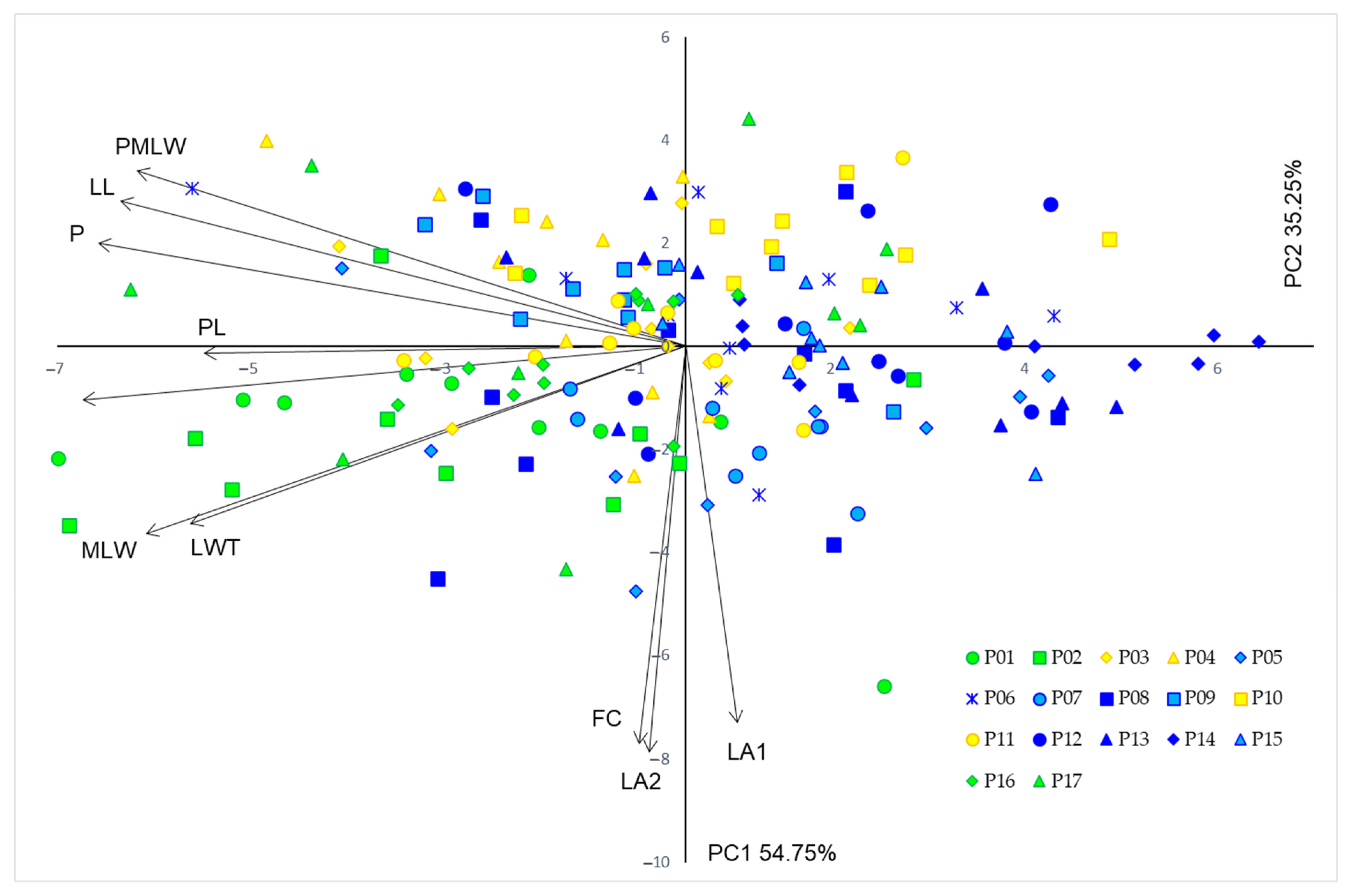
3.1. Population Diversity and Phenotypic Traits
3.2. Climate Differences among Sampling Sites
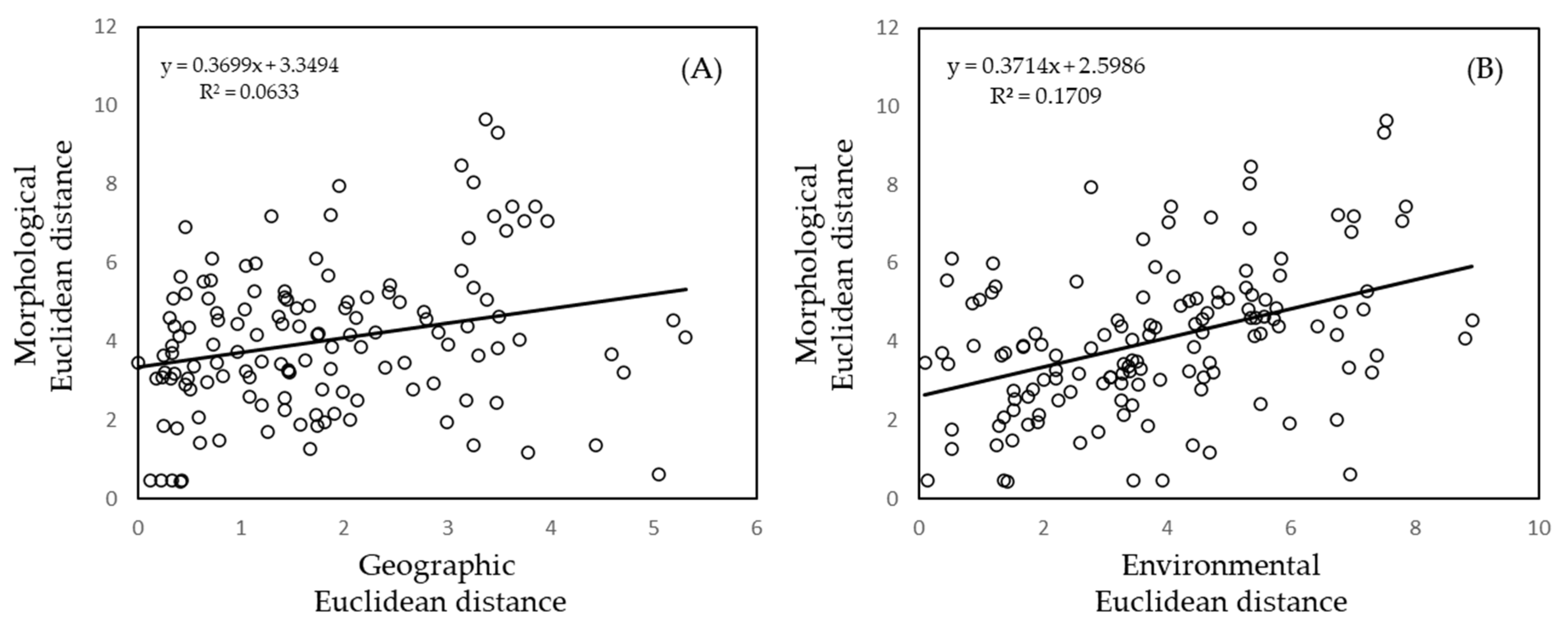
3.3. Correlations between Geographic, Environmental, and Morphometric Data
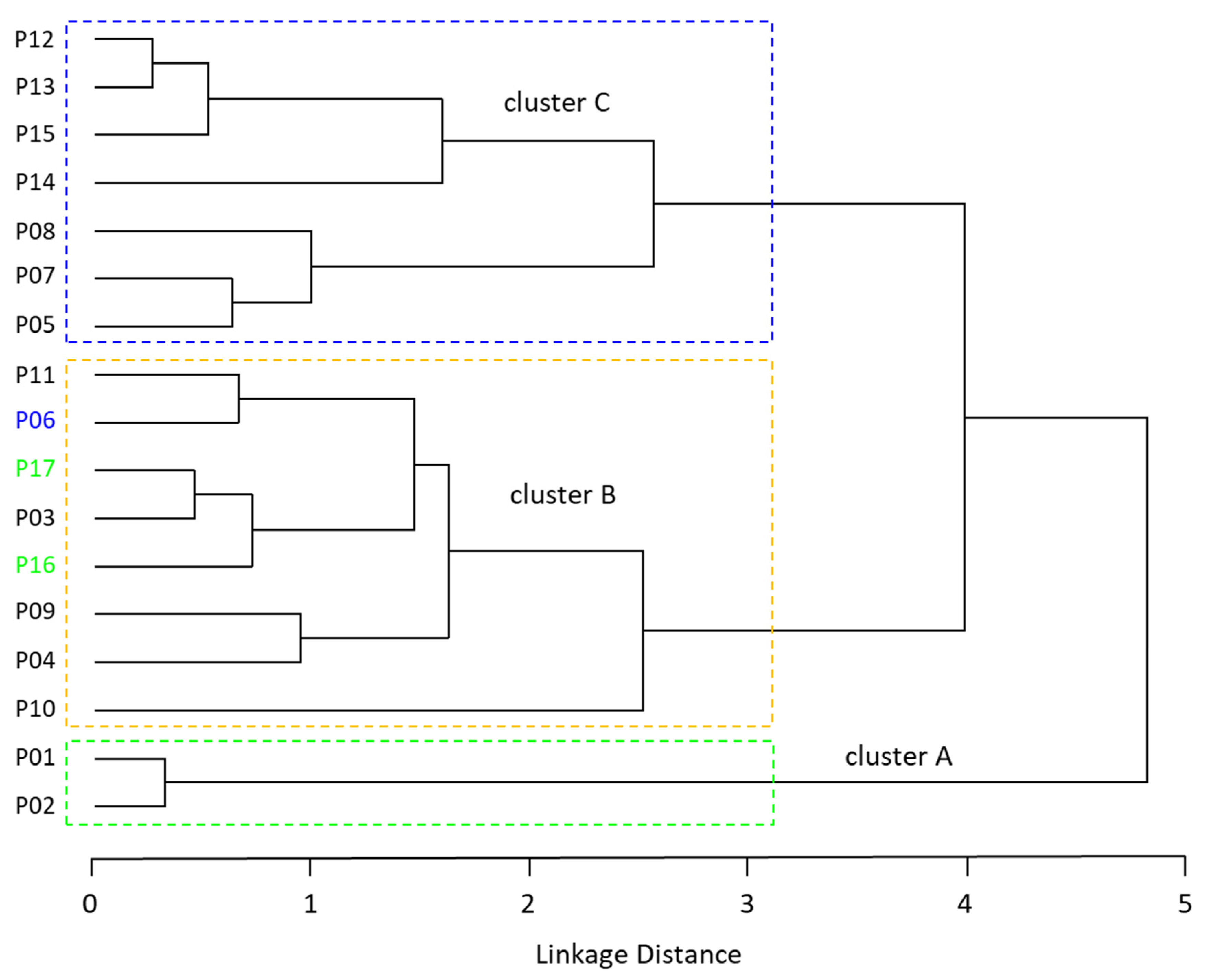
3.4. Population Structure
4. Discussion
4.1. Population Diversity and Phenotypic Traits
4.2. Correlations between Geographic, Environmental, and Morphometric Data
4.3. Population Structure
5. Conclusions and Practical Implications
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Potter, D.; Eriksson, T.; Evans, R.C.; Oh, S.; Smedmark, J.E.E.; Morgan, D.R.; Kerr, M.; Robertson, K.R.; Arsenault, M.; Dickinson, T.A.; et al. Phylogeny and classification of Rosaceae. Plant Syst. Evol. 2007, 266, 5–43. [Google Scholar] [CrossRef]
- Campbell, C.S.; Evans, R.C.; Morgan, D.R.; Dickinson, T.A.; Arsenault, M.P. Phylogeny of subtribe Pyrinae (formerly the Maloideae, Rosaceae): Limited resolution of a complex evolutionary history. Plant Syst. Evol. 2007, 266, 119–145. [Google Scholar] [CrossRef]
- Watkins, R. Apple and Pear. In Evolution of Crop Plants; Simmonds, N.W., Ed.; Longman Group Ltd.: London, UK, 1976; p. 339. [Google Scholar]
- Bell, R.L.; Itai, A. Pyrus. In Wild Crop Relatives: Genomic and Breeding Resources; Kole, C., Ed.; Springer: Berlin/Heidelberg, Germany, 2010; pp. 147–177. [Google Scholar] [CrossRef]
- Volk, G.M.; Cornille, A. Genetic diversity and domestication history in Pyrus. In The Pear Genome: Compendium of Plant Genomes; Korban, S., Ed.; Springer Nature Switzerland AG: Cham, Switzerland, 2019. [Google Scholar] [CrossRef]
- Erfani-Moghadam, J.; Zarei, A. Assessment of genetic structure among different pear species (Pyrus spp.) using apple-derived SSR and evidence of duplications in the pear genome. Biotechnol. Biotechnol. Equip. 2018, 32, 591–601. [Google Scholar] [CrossRef] [Green Version]
- Hancock, J.; Lobos, G. Pears. In Temperate Fruit Crop Breeding; Hancock, J.F., Ed.; Springer: Dordrecht, The Netherlands, 2008; pp. 299–336. [Google Scholar] [CrossRef]
- Bell, R.L.; Quamme, H.A.; Layne, R.E.C.; Skirvin, R.M. Pears. In Fruit Breeding: Tree and Tropical Fruits; Janick, J., Moore, J.N., Eds.; Wiley: New York, NY, USA, 1996; Volume 1, pp. 441–514. [Google Scholar]
- Aldasoro, J.J.; Aedo, C.; Muñoz Garmendia, F. The genus Pyrus L. (Rosaceae) in south-west Europe and North Africa. Bot. J. Linn. Soc. 1996, 121, 143–158. [Google Scholar] [CrossRef] [Green Version]
- Karimi, H.; Eslami, B.; Dehpouri, A.A.; Rad, A.S. Quantitative and qualitative analyses of genus Pyrus L. (Pear Tree) in West Azerbaijan province, Iran. Iran J. Sci. Technol. Trans. A Sci. 2019, 43, 715–724. [Google Scholar] [CrossRef]
- Kumar, S.; Kirk, C.; Deng, C.; Wiedow, C.; Knaebel, M.; Brewer, L. Genotyping-by-sequencing of pear (Pyrus spp.) accessions unravels novel patterns of genetic diversity and selection footprints. Hortic. Res. 2017, 4, 17015. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Challice, J.S.; Westwood, M.N. Numerical taxonomic studies of the genus Pyrus using both chemical and botanical characters. Bot. J. Linn. Soc. 1973, 67, 121–148. [Google Scholar] [CrossRef]
- Browicz, K. Concept and chorology of the genus Pyrus L. Arbor. Kórnickie 1993, 38, 17–33. [Google Scholar]
- Bassil, N.; Postman, J.D. Identification of European and Asian pears using EST-SSRs from Pyrus. Genet. Resour. Crop Evol. 2009, 57, 357–370. [Google Scholar] [CrossRef]
- Zamani, A.; Attar, F.; Maroofi, H. A synopsis of the genus Pyrus (Rosaceae) in Iran. Nord. J. Bot. 2012, 30, 310–332. [Google Scholar] [CrossRef]
- Reim, S.; Lochschmidt, F.; Proft, A.; Wolf, H.; Wolf, H. Species delimitation, genetic diversity and structure of the European indigenous wild pear (Pyrus pyraster) in Saxony, Germany. Genet. Resour. Crop Evol. 2016, 64, 1075–1085. [Google Scholar] [CrossRef]
- Idžojtić, M. Dendrology—Leaf; University of Zagreb, Faculty of Forestry: Zagreb, Croatia, 2009. [Google Scholar]
- Jovanović, B. Fam. Malaceae Lois.-Deslongch. In Flora Srbije; Josifović, M., Ed.; Serbian Academy of Sciences and Arts: Belgrade, Serbia, 1972; Volume 4, pp. 127–178. [Google Scholar]
- Procopiou, J.; Wallace, A. A wild pear native to calcareous soils that has a possible as a pear rootstock. J. Plant Nutr. 2000, 23, 1969–1972. [Google Scholar] [CrossRef]
- Idžojtić, M. Dendrology—Flower, Cone, Fruit, Seed; Academic Press: London, UK, 2013. [Google Scholar]
- Krüssmann, G. Handbuch der Laubgehölze; Paul Parkey in Berlin und Hamburg, Verlag für Landwirtschaft, Veterinärmedizin, Gartenbau und Forstwesen: Berlin, Germany, 1976; Volume 3. [Google Scholar]
- Dostálek, J. Pyrus spinosa und ihre Hybriden in Südwestbulgarien. Folia Geobot. Phytotax. 1980, 15, 59–73. [Google Scholar] [CrossRef]
- Kurtto, A. Rosaceae (pro Parte Majore). Euro+Med Plantbase—The Information Resource for Euro-Mediterranean Plant Diversity. 2009. Available online: http://ww2.bgbm.org/EuroPlusMed/ (accessed on 30 July 2021).
- Zohary, D. Wild apples and wild pears. Bocconea 1997, 7, 409–416. [Google Scholar]
- Fedorov, A.A. Caucasian Representatives of the Genus Pyrus L. (Morphological-Geographical Investigation); The Armenian Branch of the USSR Academy of Sciences: Yerevan, Armenia, 1943. [Google Scholar]
- Korotkova, N.; Parolly, G.; Khachatryan, A.; Ghulikyan, L.; Sargsyan, H.; Akopian, J.; Borsch, T.; Gruenstaeudl, M. Towards resolving the evolutionary history of Caucasian pears (Pyrus, Rosaceae)—Phylogenetic relationships, divergence times and leaf trait evolution. J. Systemat. Evol. 2018, 56, 35–47. [Google Scholar] [CrossRef]
- Falcinelli, F.; Moraldi, M. Il pero mandorlino (Pyrus amygdaliformis Villars.). For. Alberi Oggi 2001, 68, 39–42. [Google Scholar]
- Tsoulpha, P.; Alexandri, S.; Tsaktsira, M. Critical factors affecting an efficient micropropagation protocol for Pyrus spinosa Forskk. J. Appl. Hortic. 2018, 20, 190–195. [Google Scholar] [CrossRef]
- Riera Sensat, F.J.; Vloebergh, A. Las formas geográficas de Pyrus amygdaliformis (Willd. y Vild.). Anal. Esc. Peritos Agríc. Espec. Agropecu. Serv. Técn. Agric. ESAB 1948, 7, 87–96. [Google Scholar]
- Matsumoto, K.; Tamura, F.; Chun, J.-P.; Tanabe, K. Native Mediterranean Pyrus rootstock, P. Amygdaliformis and P. Elaeagrifolia, present higher tolerance to salinity stress compared with Asian natives. J. Jpn. Soc. Hortic. Sci. 2006, 75, 450–457. [Google Scholar] [CrossRef] [Green Version]
- Volk, G.M.; Richards, C.M.; Henk, A.D.; Reilley, A.A.; Bassil, N.V.; Postman, J.D. Diversity of wild Pyrus communis based on microsatellite analyses. J. Am. Soc. Hortic. Sci. 2006, 131, 408–417. [Google Scholar] [CrossRef] [Green Version]
- Wolko, Ł.; Antkowiak, W.; Lenartowicz, E.; Bocianowski, J. Genetic diversity of European pear cultivars (Pyrus communis L.) and wild pear (Pyrus pyraster (L.) Burgsd.) inferred from microsatellite markers analysis. Genet. Resour. Crop Evol. 2010, 57, 801–806. [Google Scholar] [CrossRef] [Green Version]
- Wolko, L.; Bocianowski, J.; Antkowiak, W.; Słomski, R. Genetic diversity and population structure of wild pear (Pyrus pyraster (L.) Burgsd.) in Poland. Open Life Sci. 2015, 10, 19–29. [Google Scholar] [CrossRef] [Green Version]
- Wünsch, A.; Hormaza, J.I. Characterization of variability and genetic similarity of European pear using microsatellite loci developed in apple. Sci. Hortic. 2007, 113, 37–43. [Google Scholar] [CrossRef]
- Kalkışım, O.; Okcu, M.; Okcu, Z.; Karabulut, B.; Yildirim, N.; Agar, G. Relationships among some pears genotypes (Pyrus communis L.) based on ISSR and RAPD analysis. Erwerbs Obstbau 2016, 58, 259–264. [Google Scholar] [CrossRef]
- Queiroz, A.; Assunção, A.; Ramadas, I.; Viegas, W.; Veloso, M.M. Molecular characterization of Portuguese pear landraces (Pyrus communis L.) using SSR markers. Sci. Hortic. 2015, 183, 72–76. [Google Scholar] [CrossRef]
- Queiroz, Á.; Bagoin Guimarães, J.; Sánchez, C.; Simões, F.; Maia de Sousa, R.; Viegas, W.; Veloso, M.M. Genetic diversity and structure of the Portuguese pear (Pyrus communis L.) germplasm. Sustainability 2019, 11, 5340. [Google Scholar] [CrossRef] [Green Version]
- Kocsisné, G.M.; Bolla, D.; Anhalt-Brüderl, U.C.M.; Forneck, A.; Taller, J.; Kocsis, L. Genetic diversity and similarity of pear (Pyrus communis L.) cultivars in Central Europe revealed by SSR markers. Genet. Resour. Crop Evol. 2020, 67, 1755–1763. [Google Scholar] [CrossRef]
- Bao, L.; Chen, K.; Zhang, D.; Cao, Y.; Yamamoto, T.; Teng, Y. Genetic diversity and similarity of pear (Pyrus L.) cultivars native to East Asia revealed by SSR (simple sequence repeat) markers. Genet. Resour. Crop Evol. 2007, 54, 959–971. [Google Scholar] [CrossRef]
- Song, Y.; Fan, L.; Chen, H.; Zhang, M.; Ma, Q.; Zhang, S.; Wu, J. Identifying genetic diversity and a preliminary core collection of Pyrus pyrifolia cultivars by a genome-wide set of SSR markers. Sci. Hortic. 2014, 167, 5–16. [Google Scholar] [CrossRef]
- Cao, Y.; Tian, L.; Gao, Y.; Liu, F. Genetic diversity of cultivated and Ussurian pear (Pyrus ussuriensis Maxim.) in China evaluated with M13-tailed SSR markers. Genet. Resour. Crop Evol. 2012, 59, 9–17. [Google Scholar] [CrossRef]
- Wuyun, T.; Amo, H.; Xu, J.; Ma, T.; Uematsu, C.; Katayama, H. Population structure of and conservation strategies for wild Pyrus ussuriensis Maxim. In China. PLoS ONE 2015, 10, e0133686. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bennici, S.; Las Casas, G.; Distefano, G.; Di Guardo, M.; Continella, A.; Ferlito, F.; Gentile, A.; La Malfa, S. Elucidating the contribution of wild related species on autochthonous pear germplasm: A case study from Mount Etna. PLoS ONE 2018, 13, e0198512. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Debussche, M.; Isenmann, P. Fleshy fruit characters and the choices of bird and mammal seed dispersers in a Mediterrenean region. OIKOS 1989, 56, 327–338. [Google Scholar] [CrossRef]
- Pigozzi, G. Frugivory and seed dispersal by the European badger in a Mediterrenean habitat. J. Mammal. 1992, 73, 630–639. [Google Scholar] [CrossRef]
- Pikelj, K.; Juračić, M. Eastern Adriatic coast (EAC): Geomorphology and coastal vulnerability of a karstic coast. J. Coast. Res. 2013, 29, 944–957. [Google Scholar] [CrossRef]
- Joffre, R.; Rambal, S. Mediterranean Ecosystems; Macmillan Publishers Ltd.; Nature Publishing Group: London, UK, 2001. [Google Scholar] [CrossRef]
- Bednorz, L. Morphological variability of leaves of Sorbus torminalis (L.) Crantz in Poland. Acta Soc. Bot. Pol. 2006, 3, 233–243. [Google Scholar] [CrossRef] [Green Version]
- WinFolia TM, version PRO 2005b; Regent Instruments Inc.: Quebec City, QC, Canada, 2001.
- Hijmans, R.J.; Cameron, S.E.; Parra, J.L.; Jones, P.G.; Jarvis, A. Very high resolution interpolated climate surfaces for global land areas. Int. J. Climatol. 2005, 25, 1965–1978. [Google Scholar] [CrossRef]
- Fick, S.E.; Hijmans, R.J. WorldClim 2: New 1km spatial resolution climate surfaces for global land areas. Int. J. Climatol. 2017, 37, 4302–4315. [Google Scholar] [CrossRef]
- Temunović, M.; Franjić, J.; Šatović, Z.; Grgurev, M.; Frascaria-Lacoste, N.; Fernández-Manjarrés, J.F. Environmental heterogeneity explains the genetic structure of continental and Mediterranean populations of Fraxinus angustifolia Vahl. PLoS ONE 2012, 7, e42764. [Google Scholar] [CrossRef] [Green Version]
- Sokal, R.R.; Rohlf, F.J. Biometry: The Principles and Practice of Statistics in Biological Research, 4th ed.; W.H. Freeman and Co.: New York, NY, USA, 2012; p. 937. [Google Scholar]
- Koutecký, P. MorphoTools: A set of R functions for morphometric analysis. Plant Syst. Evol. 2015, 301, 1115–1121. [Google Scholar] [CrossRef]
- Mantel, N. The detection of disease clustering and a generalized regression approach. Cancer Res. 1967, 27, 209–220. [Google Scholar] [PubMed]
- Legendre, P.; Fortin, M.J. Spatial pattern and ecological analysis. Vegetatio 1989, 80, 107–138. [Google Scholar] [CrossRef]
- Oksanen, J.; Blanchet, F.G.; Friendly, M.; Kindt, R.; Legendre, P.; McGlinn, D.; Minchin, P.R.; O’Hara, R.B.; Simpson, G.L.; Solymos, P.; et al. Package ‘Vegan’: Community Ecology Package; Version 2.4-2. 2017. Available online: https://github.com/vegandevs/vegan (accessed on 24 June 2021).
- McGarigal, K.; Cushman, S.; Stafford, S. Multivariate Statistics for Wildlife and Ecology Research; Springer: New York, NY, USA, 2000; p. 283. [Google Scholar]
- Douaihy, B.; Sobierajska, K.; Jasińska, A.K.; Boratyńska, K.; Ok, T.; Romo, A.; Machon, N.; Didukh, Y.; Dagher-Kharrat, M.B.; Boratyński, A. Morphological versus molecular markers to describe variability in Juniperus excelsa subsp. excelsa (Cupressaceae). AOB Plants 2012, 2012, pls013. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Boratyński, A.; Jasińska, A.K.; Marcysiak, K.; Mazur, M.; Romo, A.M.; Boratyńska, K.; Sobierajska, K.; Iszkuło, G. Morphological differentiation supports the genetic pattern of the geographic structure of Juniperus thurifera (Cupressaceae). Plant Syst. Evol. 2013, 299, 773–784. [Google Scholar] [CrossRef] [Green Version]
- Sobierajska, K.; Boratyńska, K.; Jasińska, A.; Dering, M.; Ok, T.; Douaihy, B.; Dagher-Kharrat, M.B.; Romo, A.; Boratyński, A. Effect of the Aegean Sea barrier between Europe and Asia on differentiation in Juniperus drupacea (Cupressaceae). Bot. J. Linn. Soc. 2016, 180, 365–385. [Google Scholar] [CrossRef] [Green Version]
- Poljak, I.; Idžojtić, M.; Šapić, I.; Korijan, P.; Vukelić, J. Diversity and structure of Croatian continental and Alpine-Dinaric populations of grey alder (Alnus incana /L./ Moench subsp. incana); Isolation by distance and environment explains phenotypic divergence. Šumar. List 2018, 142, 19–31. [Google Scholar] [CrossRef]
- Statistica (Data Analysis Software System), version 13; TIBCO Software Inc.: Palo Alto, CA, USA, 2018; Available online: http://www.statsoft.com. (accessed on 15 September 2021).
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2016; Available online: http://www.R-project.org/. (accessed on 15 September 2021).
- Yilmaz, K.U.; Uzun, A.; Cam, M.; Ercisli, S. Some morphological and fruit characteristics of naturally grown Pyrus elaeagnifolia Pall. of Kayseri province (Central Anatolia, Turkey). Gen. Resour. Crop. Evol. 2015, 62, 711–720. [Google Scholar] [CrossRef]
- Terpó, J.; de Amaral Franco, J. Pyrus L. In Flora Europaea; Tutin, T.G., Heywood, V.H., Burges, N.A., Moore, D.M., Valentine, D.H., Walters, S.M., Webb, D.A., Eds.; Cambridge University Press: Cambridge, UK, 1968; pp. 65–66. [Google Scholar]
- Paganová, V. Taxonomic reliability of leaf and fruit morphological characteristics of the Pyrus L. taxa in Slovakia. Hortic. Sci 2003, 30, 98–107. [Google Scholar] [CrossRef] [Green Version]
- Čolić, S.; Rakonjac, V.; Zec, G.; Nikolić, D.; Fotirić Akšić, M. Morphological and biochemical evaluation of selected almond (Prunus dulcis (Mill.) D.A.Webb.) genotypes in Northern Serbia. Turk. J. Agric. For. 2012, 36, 429–438. [Google Scholar] [CrossRef]
- Khadivi-Khub, A.; Zamani, Z.; Reza Fatahi, M. Multivariate analysis of Prunus subgen. Cerasus germplasm in Iran using morphological variables. Genet. Resour. Crop Evol. 2012, 59, 909–926. [Google Scholar] [CrossRef]
- Farsad, A.; Esna-Ashari, M. Genetic diversity of some Iranian sweet cherry (Prunus avium) cultivars using microsatellite markers and morphological traits. Cytol. Genet. 2016, 50, 8–19. [Google Scholar] [CrossRef]
- Miljković, D.; Stefanović, M.; Orlović, S.; Stanković Neđić, M.; Kesić, L.; Stojnić, S. Wild cherry (Prunus avium (L.) L.) leaf shape and size variations in natural populations at different elevations. Alp. Bot. 2019, 129, 163–174. [Google Scholar] [CrossRef]
- Kumar, C.; Singh, S.K.; Pramanick, K.K.; Verma, M.K.; Srivastav, M.; Rakesh, S.; Bharadwaj, C.; Naga, K.C. Morphological and biochemical diversity among the Malus species including indigenous Himalayan wild apples. Sci. Hortic. 2018, 233, 204–219. [Google Scholar] [CrossRef]
- Kidner, C.A.; Umbreen, S. Why is leaf shape so variable? Int. J. Plant Dev. Biol. 2010, 4, 64–75. [Google Scholar]
- Niinemets, U. Adjustment of foliage structure and function to a canopy light gradient in two co-existing deciduous trees; Variability in leaf inclination angles in relation to petiole morphology. Trees 1998, 12, 446–451. [Google Scholar] [CrossRef]
- Kessler, S.; Sinha, N. Shaping up: The genetic control of leaf shape. Curr. Opin. Plant Biol. 2004, 7, 65–72. [Google Scholar] [CrossRef] [PubMed]
- Tsukaya, H. Leaf shape: Genetic controls and environmental factors. Int. J. Dev. Biol. 2005, 49, 547–555. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Popović, V.; Kerkez, I. Population variability of wild cherry (Prunus avium L.) in Serbia according to the leaf morphology. Šumar. List 2016, 140, 347–355. [Google Scholar] [CrossRef] [Green Version]
- Brus, R.; Ballian, D.; Bogunić, F.; Bobinac, M.; Idžojtić, M. Leaflet morphometric variation of service tree (Sorbus domestica L.) in the Balkan Peninsula. Plant Biosyst. 2011, 145, 278–285. [Google Scholar] [CrossRef]
- Said, A.A.; Oukabli, A.; Gaboun, F.; Simard, M.H.; El Modafar, C. Phenotypic biodiversity of an endemic wild pear, Pyrus mamorensis Trab. in North-western Morocco using morphological descriptors. Genet. Resour. Crop Evol. 2013, 60, 927–938. [Google Scholar] [CrossRef]
- Zarei, A.; Erfani-Moghadam, J.; Jalilian, H. Assessment of variability within and among four Pyrus species using multivariate analysis. Flora 2019, 250, 27–36. [Google Scholar] [CrossRef]
- Nadjat, T.; Tayeb, S. Effect of environmental conditions on morphological variability of leaves and fruits of five populations of Pistacia atlantica Desf. in North Algeria. Biodiv. Res. Conserv. 2020, 58, 1–12. [Google Scholar] [CrossRef]
- Garland, T., Jr.; Kelly, S.A. Phenotypic plasticity and experimental evolution. J. Exp. Biol. 2006, 209, 2344–2361. [Google Scholar] [CrossRef] [Green Version]
- Gratani, L. Plant phenotypic plasticity in response to environmental factors. Adv. Bot. 2014, 2014, 208747. [Google Scholar] [CrossRef] [Green Version]
- Benito Garzón, M.; Alía, R.; Robson, T.M.; Zavala, M.A. Intra-specific variability and plasticity influence potential tree species distributions under climate change. Glob. Ecol. Biogeogr. 2011, 20, 766–778. [Google Scholar] [CrossRef] [Green Version]
- Fox, R.J.; Donelson, J.M.; Schunter, C.; Ravasi, T.; Gaitán-Espitia, J.D. Beyond buying time: The role of plasticity in phenotypic adaptation to rapid environmental change. Philos. Trans. B 2019, 374, 20180174. [Google Scholar] [CrossRef] [PubMed]
- Savolainen, O.; Pyhäjärvi, T.; Knürr, T. Gene flow and local adaptation in trees. Ann. Rev. Ecol. Evol. Syst. 2007, 38, 595–619. [Google Scholar] [CrossRef]
- Wright, S. Isolation by distance. Genetics 1943, 28, 114–138. [Google Scholar] [CrossRef] [PubMed]
- Wang, I.J.; Bradburd, G.S. Isolation by environment. Mol. Ecol. 2014, 23, 5649–5662. [Google Scholar] [CrossRef]
- Nosil, P. Ecological Speciation; Oxford University Press: Oxford, UK, 2012. [Google Scholar] [CrossRef]
- Cortés, A.J.; Garzón, L.N.; Valencia, J.B.; Madriñán, S. On the causes of rapid diversification in the Páramos: Isolation by ecology and genomic divergence in Espeletia. Front. Plant Sci. 2018, 9, 1700. [Google Scholar] [CrossRef]
- Orsini, L.; Vanoverbeke, J.; Swillen, I.; Mergeay, J.; De Meester, L. Drivers of population genetic differentiation in the wild: Isolation by dispersal limitation, isolation by adaptation and isolation by colonization. Mol. Ecol. 2013, 22, 5983–5999. [Google Scholar] [CrossRef] [PubMed]
- Dewoody, J.; Trewin, H.; Taylor, G. Genetic and morphological differentiation in Populus nigra L.: Isolation by colonization or isolation by adaptation? Mol. Ecol. 2015, 24, 2641–2655. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ribbens, E.; Silander, J.A.; Pacala, S.W. Seedling recruitment in forests: Calibrating models to predict patterns of tree seedling dispersion. Ecology 1994, 75, 1794–1806. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Trait | Acronyms | M | SD | Min | Max | CV (%) |
|---|---|---|---|---|---|---|
| Leaf area (cm2) | LA | 3.31 | 1.51 | 0.42 | 12.53 | 45.62 |
| Perimeter (cm) | P | 8.22 | 1.99 | 3.31 | 17.09 | 24.16 |
| Form coefficient | FC | 0.60 | 0.11 | 0.27 | 0.95 | 18.02 |
| Leaf length (cm) | LL | 3.41 | 0.86 | 1.34 | 7.32 | 25.23 |
| Maximal leaf width (cm) | MLW | 1.32 | 0.36 | 0.40 | 3.26 | 27.36 |
| Position of maximal leaf width (cm) | PMLW | 1.73 | 0.48 | 0.54 | 3.97 | 27.69 |
| Leaf width top (cm) | LWT | 0.67 | 0.20 | 0.20 | 1.86 | 29.81 |
| Leaf angle 1 (°) | LA1 | 36.15 | 8.18 | 14.00 | 67.00 | 22.61 |
| Leaf angle 2 (°) | LA2 | 29.28 | 6.38 | 12.00 | 55.00 | 21.81 |
| Petiole length (cm) | PL | 1.13 | 0.49 | 0.06 | 3.62 | 43.22 |
| Trait | PC—Principal Component | ||
|---|---|---|---|
| PC1 | PC2 | PC3 | |
| Leaf area | −0.972374 | −0.131015 | −0.101500 |
| Perimeter | −0.948275 | 0.250623 | 0.000705 |
| Form coefficient | −0.075091 | −0.965928 | −0.072853 |
| Leaf length | −0.911706 | 0.352156 | 0.005661 |
| Maximal leaf width | −0.870284 | −0.456680 | −0.098129 |
| Position of maximal leaf width | −0.885013 | 0.426979 | −0.102035 |
| Leaf width top | −0.799420 | −0.430650 | −0.229308 |
| Leaf angle 1 | 0.083034 | −0.915972 | 0.117808 |
| Leaf angle 2 | −0.058789 | −0.986012 | 0.068873 |
| Petiole length | −0.776832 | −0.016807 | 0.596121 |
| Eigenvalue | 5.47 | 3.52 | 0.46 |
| % Total variance | 54.75 | 35.25 | 4.62 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Vidaković, A.; Liber, Z.; Šatović, Z.; Idžojtić, M.; Volenec, I.; Zegnal, I.; Pintar, V.; Radunić, M.; Poljak, I. Phenotypic Diversity of Almond-Leaved Pear (Pyrus spinosa Forssk.) along Eastern Adriatic Coast. Forests 2021, 12, 1630. https://0-doi-org.brum.beds.ac.uk/10.3390/f12121630
Vidaković A, Liber Z, Šatović Z, Idžojtić M, Volenec I, Zegnal I, Pintar V, Radunić M, Poljak I. Phenotypic Diversity of Almond-Leaved Pear (Pyrus spinosa Forssk.) along Eastern Adriatic Coast. Forests. 2021; 12(12):1630. https://0-doi-org.brum.beds.ac.uk/10.3390/f12121630
Chicago/Turabian StyleVidaković, Antonio, Zlatko Liber, Zlatko Šatović, Marilena Idžojtić, Ida Volenec, Ivana Zegnal, Valentino Pintar, Mira Radunić, and Igor Poljak. 2021. "Phenotypic Diversity of Almond-Leaved Pear (Pyrus spinosa Forssk.) along Eastern Adriatic Coast" Forests 12, no. 12: 1630. https://0-doi-org.brum.beds.ac.uk/10.3390/f12121630