
Growth Response to Weed Control and Fertilisation in Mid-Rotation Plantations of Eucalyptus pellita in South Sumatra, Indonesia


Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Site
2.2. Site Preparation and Plantation Establishment
2.3. Experimental Design and Treatments
- A factorial matrix of three rates of N (N0, N125, and N250 kg ha−1) by two levels of weed control (up to age 1 year and for the full rotation). No additional P was applied in this matrix. In total, there were six treatments in the factorial mixture (Treatments 1–6).
- Additional treatments to explore the potential for further response to P fertiliser: (N125P30 and N0P30). Fertilisers were applied at age 1 year, and weed control was conducted throughout the rotation (Treatments 7–8).
- Additional treatments to test responses to two levels of N fertiliser (N125 and N250) were given at age 2 years. These treatments received full weed control throughout the rotation (Treatments 9–10).
2.4. Measurements and Data Analysis
3. Results
3.1. Response to Fertiliser and Weed-Control Treatment in Mid-Rotation
3.2. Response to Weed Control and P Applied at Planting Time
4. Discussion
4.1. Influence of Fertilisation in Mid-Rotation on Productivity
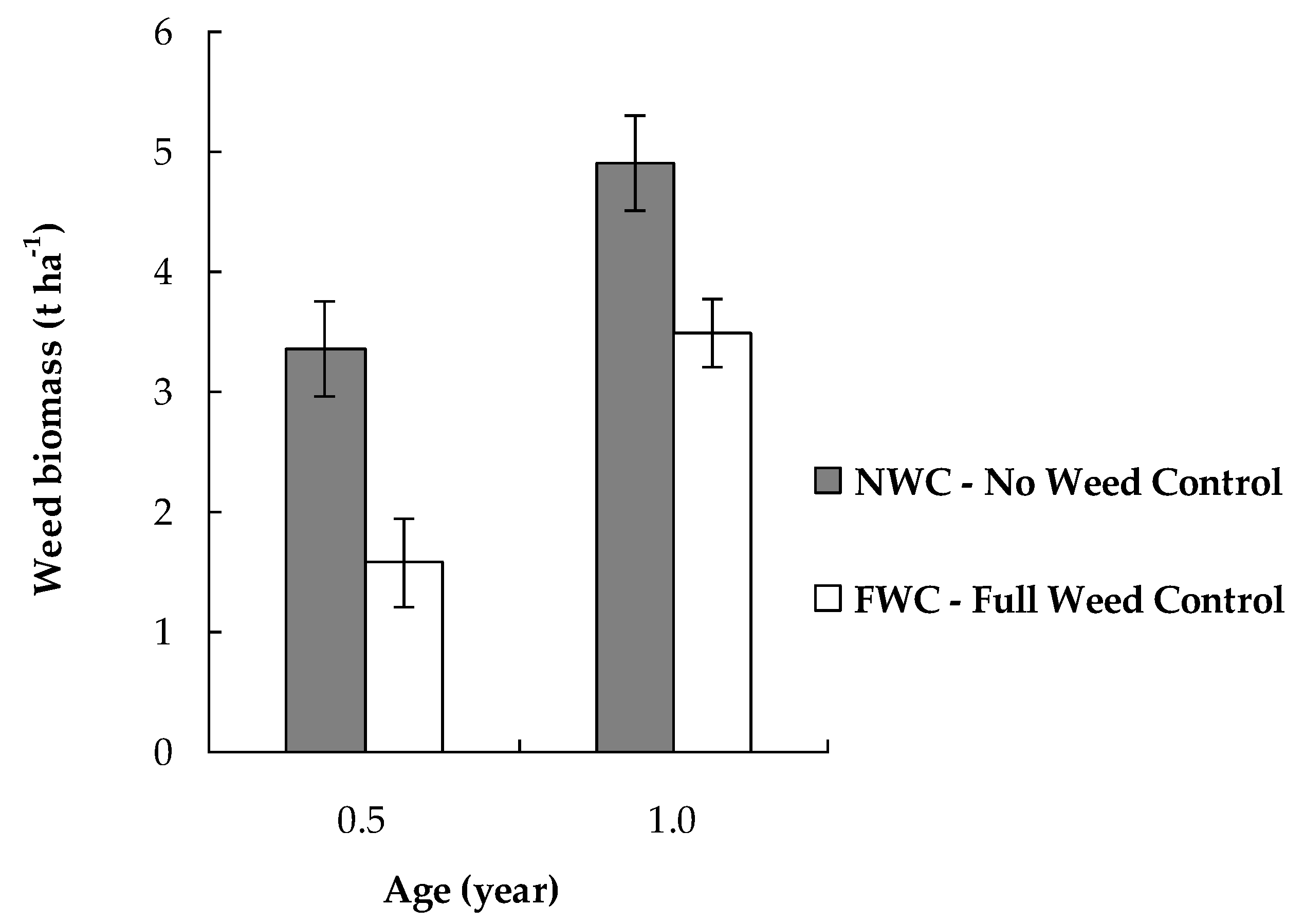
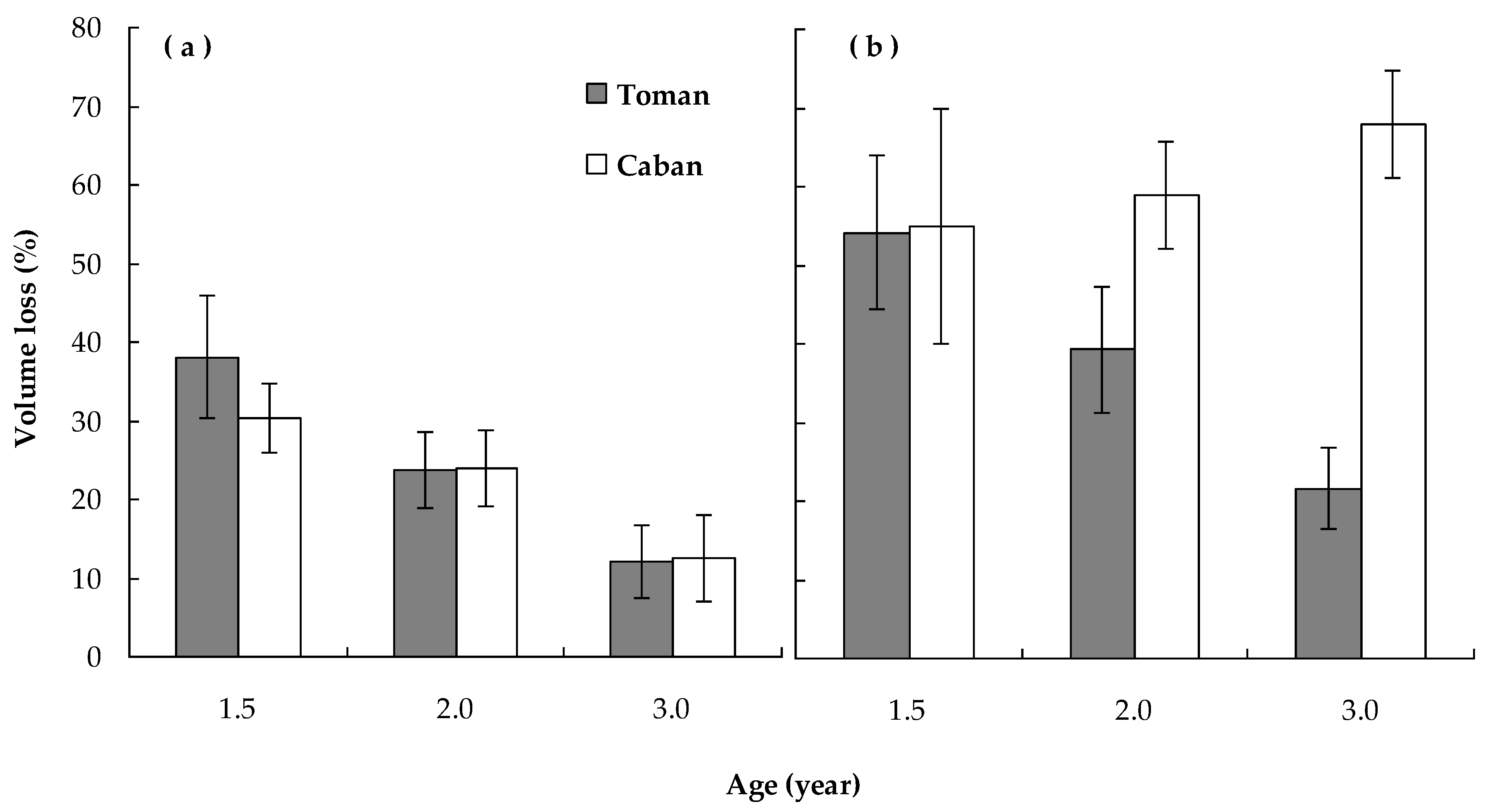
4.2. Effect of Weed Competition on Tree Growth
4.3. Implications and Challenges for Operational Plantations
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Tarigan, M.; Roux, J.; van Week, M.; Tjahjono, B.; Wingfield, M.J. A new wilt and die-back disease of A. mangium associated with Ceratocystis manginecans and C. acaciicvora sp. nov. in Indonesia. S. Afr. J. Bot. 2010, 77, 292–304. [Google Scholar] [CrossRef] [Green Version]
- Nasution, A.; Glen, M.; Beadle, C.; Mohammed, C. Ceratocystis wilt and canker—A disease that compromises the growing of commercial Acacia-based plantations in the tropics. Aust. For. 2019, 82, 80–93. [Google Scholar] [CrossRef] [Green Version]
- Inail, M.A.; Hardiyanto, E.B.; Mendham, D.S. Growth responses of Eucalyptus pellita F. Muell plantations in South Sumatra to macronutrient fertilisers following several rotations of Acacia mangium willd. Forests 2019, 10, 1054. [Google Scholar] [CrossRef] [Green Version]
- Hardiyanto, E.B.; Inail, M.A.; Nambiar, E.K.S. Productivity of Eucalyptus pellita in Sumatra: Acacia mangium Legacy, Response to Phosphorus, and Site Variables for Guiding Management. Forests 2021, 12, 1186. [Google Scholar] [CrossRef]
- Harwood, C.E.; Nambiar, E.K.S. Productivity of acacia and eucalypt plantations in Southeast Asia. 2. Trends and variations. Int. For. Rev. 2014, 16, 249–260. [Google Scholar] [CrossRef]
- Sunarminto, B.H.; Kertonogoro, B.D.; Purwanto, B.H.; Shiddieq, D.; Syukur, A.; Sulakhudin Hatta, M.; Rif’an, M. Soil Mapping and Survey of Region Forest Group PT.MHP South Sumatera; Soil Science Departement, Faculty of Agriculture, Gadjah Mada University: Yogyakarta, Indonesia, 2012; p. 10. [Google Scholar]
- Soil Survey Staff. Keys to Soil Taxonomy, 12th ed.; USDA-Natural Resources Conservation Service: Washington, DC, USA, 2014; p. 359.
- Supriadi, B. Form factor of Eucalyptus pellita. Technical Notes R&D. PT Musi Hutan Persada 2011, 21, 1–5. [Google Scholar]
- Wibisono, M.G.; Veneklaas, E.; Mendham, D.S.; Hardiyanto, E.B. Nitrogen fixation of Acacia mangium Willd. from two seed sources grown at different levels of phosphorus in an Ultisol, South Sumatra, Indonesia. Southern Forests. J. For. Sci. 2015, 77, 59–64. [Google Scholar]
- Mendham, D.S.; Greenhill, M.; Beadle, C.; Hardiyanto, E.; Wibisono, G.; Rimbawanto, A.; Fauzi, A.; Dong, T.L.; Huong, V.D.; Hai, T.A.; et al. Maximising Productivity and Profitability of Eucalypts and Acacias in Indonesia and Vietnam; Final Report of FST/2014/064; Australian Centre for Agricultural Research: Canberra, QT, Australia, 2020; p. 30.
- Pulito, A.P.; Goncalves, J.L.M.; Smethurst, P.J.; Arthur-Junior, J.C.; Alvares, C.A.; Rocha, J.H.T.; Hubner, A.; de Moraes, L.F.; Miranda, A.C.; Kamogawa, M.Y.; et al. Available nitrogen and responses to nitrogen ferilizer in Brazilian Eucalypt plantations on Soils of contrasting texture. Forests 2015, 6, 973–991. [Google Scholar] [CrossRef] [Green Version]
- Albaugh, T.J.; Rubilar, R.A.; Fox, T.R.; Lee Allen, H.; Urrego, J.B.; Zapata, M.; Stape, J.L. Response of Eucalyptus grandis in Colombia to mid-rotation fertilization is dependent on site and rate but not frequency of application. For. Ecol. Manag. 2015, 350, 30–39. [Google Scholar] [CrossRef] [Green Version]
- Du Toit, B.; Arbuthnot, A.; Oscroft, D.; Job, R.A. The effect of remedial fertilizer treatments on growth and pulp properties of Eucalyptus grandis stands established on infertile soils of the Zululand coastal plain. S. Afr. For. J. 2001, 192, 9–19. [Google Scholar]
- NC State University. Forest Productivity Cooperative, Shaping the Future of Plantation Forestry. In Effects of N and P Fertilization Post-Establishment on Growth of Eucalyptus nitens Plantations in the South-Central Chile; Eucalyptus Forestry Summaries 2014; NC State University: Raleigh, NC, USA, 2019; Volume 2, pp. 15–16. [Google Scholar]
- Carneiro, M.; Fabiao, A.; Martins, M.C.; Fabiao, A.; da Silva, M.A.; Hilario, L.; Lousa, M.; Madeira, M. Effects of harrowing and fertilisation on understory vegetation and timber production of a Eucalyptus globulus Labill. Plantatation in Central Portugal. For. Ecol. Manag. 2008, 255, 591–597. [Google Scholar] [CrossRef]
- Laclau, J.P.; Ranger, J.; Gonçalves, J.L.M.; Maquere, V.; Krusche, A.V.; M’bou, A.T.; Nouvellon, Y.; Saint-Andre, L.; Bouillet, J.P.; Piccolo, M.C.; et al. Biogeochemical cycles of nutrients in tropical Eucalyptus plantations Main features shown by intensive monitoring in Congo and Brazil. For. Ecol. Manag. 2010, 259, 1771–1785. [Google Scholar] [CrossRef]
- Inail, A.M.; Hardiyanto, E.B. Growth of Eucalyptus pellita under different harvest residue management after three rotations of Acacia mangium at PT Musi Hutan Persada, South Sumatra. Technical Notes R&D. PT Musi Hutan Persada 2021, 30, 1–8. [Google Scholar]
- Inail, A.M.; Juniarso, S.; Pertiwi, P.S.; Al-Hanan, R.M. Litter decomposition of Eucalyptus pellita in Subanjeriji, South Sumatra. Technical Notes R&D. PT Musi Hutan Persada 2021, 31, 1–5. [Google Scholar]
- Adams, P.R.; Beadle, C.L.; Mendham, N.J.; Smethurst, P.J. The impact of timing and duration of grass control on growth of a young Eucalyptus globulus Labill. plantation. New For. 2003, 26, 147–165. [Google Scholar] [CrossRef]
- George, B.H.; Brennan, P.D. Herbicides are more cost-effective than alternative weed control methods for increasing early growth of Eucalyptus dunnii and Eucalyptus saligna. New For. 2002, 24, 147–163. [Google Scholar] [CrossRef]
- Little, K.M.; van Standen, J. Effect of vegetation control on Eucalyptus grandis x E. camaldulensis volume and economics. S. Afr. J. Bot. 2005, 7, 418–425. [Google Scholar] [CrossRef] [Green Version]
- Little, K.M. Performance of Eucalyptus dunnii as influenced by vegetation control when felled at nine years in KwaZulu-Natal, South Africa. South. For. 2008, 70, 183–191. [Google Scholar] [CrossRef]
- Garau, A.M.; Ghersa, C.M.; Lemcoff, J.H.; Barañao, J.J. Weeds in Eucalyptus globulus subsp. maidenii (F. Muell) establishment: Effects of competition on sapling growth and survivorship. New For. 2009, 37, 251–264. [Google Scholar] [CrossRef]
- Eyles, A.; Worledge, D.; Sands, P.; Ottenschlaeger, M.L.; Paterson, S.C.; Mendham, D.; O’Grady, A.P. Ecophysiological responses of a young blue gum (Eucalyptus globulus) plantation to weed control. Tree Physiol. 2012, 32, 1008–1020. [Google Scholar] [CrossRef] [Green Version]
- Vargas, F.; Rubiar, R.; Gonzales-Benecke, C.A.; Sanchez-Olate, M.; Acaenana, P. Long-term response to area of competition control in Eucalyptus globulus plantations. New For. 2018, 49, 383–398. [Google Scholar] [CrossRef]
- Nurudin, M.; Ohta, S.; Hardiyanto, E.B.; Mendham, D.S.; Wicaksono, A.; Heriyanto, J.; Watanabe, M. Relationships between soil characteristics and productivity of Acacia mangium in South Sumatra. Tropics 2013, 22, 1–12. [Google Scholar] [CrossRef]
- Hardie, M.; Mendham, D.S.; Corkrey, R.; Hardiyanto, E.; Maydra, A.; Siregar, S.; Marolop, R.; Wibowo, A. Effects of eucalypt and acacia plantations on soil water in Sumatra. New For. 2018, 49, 87–104. [Google Scholar] [CrossRef]
- Inail, A.M.; Supriadi, B.; Thaher, E.; Juniarso, S. Productivity of Clone Site Interaction Group 70 in Various Site Character. Technical Notes R&D. PT Musi Hutan Persada 2019, 29, 1–5. [Google Scholar]
- Inail, A.M.; Thaher, E. Potensi dan Respons Pemupukan Coppice di ex. Rotasi pertama Eucalyptus pellita. Technical Notes R&D. PT Musi Hutan Persada 2021, 30, 1–5. (In Indonesian) [Google Scholar]
- Inail, A.M.; Supriadi, B.; Thaher, E. Respon Berbagai Jarak Tanam dalam Memaksimalkan Produktivitas Eucalyptus pellita. Technical Notes R&D. PT Musi Hutan Persada 2016, 26, 1–6. (In Indonesian) [Google Scholar]




{kind=link}
{kind=link}
{kind=link}
| Variable | Site | |||
|---|---|---|---|---|
| Core-Sodong (Subanjeriji) | Satellite-Toman (Subanjeriji) | Satellite-Caban (Subanjeriji) | Core-Lagan | |
| Slope (%) | 1.75 | 1.40 | 5.77 | 9.01 |
| Bulk density 0–20 (g cm−3) | 0.87 | 1.26 | 1.05 | 1.06 |
| Depth to plinthite layer (cm) | >100 | 76 | >100 | 50 |
| Depth of A-horizon (cm) | 18 | 17 | 12 | 15 |
| pH (H2O) | 3.8 | 4.3 | 4.7 | 4.18 |
| Soil organic carbon (%) | 3.23 | 4.23 | 3.00 | 1.80 |
| N total (%) | 0.22 | 0.26 | 0.16 | 0.15 |
| Soil textures (%) | ||||
| - Clay | 53.2 | 38.3 | 45.0 | 43.6 |
| - Silt | 29.7 | 30.4 | 7.3 | 42.1 |
| - Sand | 17.0 | 31.2 | 47.7 | 14.3 |
| Cation exchange capacity (CEC) (cmol kg−1) | 12.1 | 13.3 | 9.5 | 12.2 |
| Exchangeable K (mg kg−1) | 65.2 | 62.6 | 66.5 | 66.9 |
| Extractable P (mg kg−1) | 2.5 | 6.0 | 3.6 | 1.8 |
| Treatment | Fertilisation | Weed Control * | |||
|---|---|---|---|---|---|
| Planting Time | At Age (Year) | ||||
| 1 | 2 | ||||
| P Fertiliser | P Fertiliser | N Fertiliser | N Fertiliser | ||
| (kg P ha−1) | (kg P ha−1) | (kg N ha−1) | (kg N ha−1) | ||
| W1N0P0 | 12 | 0 | 0 | 0 | Full |
| W1N125P0 | 12 | 0 | 125 | 0 | Full |
| W1N250P0 | 12 | 0 | 250 | 0 | Full |
| W0N0P0 | 12 | 0 | 0 | 0 | Up to 1 year |
| W0N125P0 | 12 | 0 | 125 | 0 | Up to 1 year |
| W0N250P0 | 12 | 0 | 250 | 0 | Up to 1year |
| W1N125P30 | 12 | 30 | 125 | 0 | Full |
| W1N0P30 | 12 | 30 | 0 | 0 | Full |
| W1N125Age2P0 | 12 | 0 | 0 | 125 | Full |
| W1N250Age2P0 | 12 | 0 | 0 | 250 | Full |
| Site | Treatment | Level | Height, H (m) | DBH, D (cm) | Volume, V (m3 ha−1) | Survival (%) |
|---|---|---|---|---|---|---|
| Sodong | Application of N at 1 year and weed control | |||||
| N fertiliser | 0 | 19.6 | 15.4 | 193.3 | 76.0 | |
| (kg ha−1) | 125 | 19.2 | 14.8 | 190.4 | 76.5 | |
| 250 | 20.2 | 15.6 | 203.7 | 74.1 | ||
| p-value | 0.15 | 0.13 | 0.60 | 0.82 | ||
| Weed control | <1 year (W0) | 19.4 | 15.0 | 190.6 | 77.0 | |
| Full (W1) | 19.9 | 15.5 | 201.1 | 74.0 | ||
| p-value | 0.24 | 0.18 | 0.39 | 0.36 | ||
| Application of P at 1 year with full weed control | ||||||
| P fertiliser | 0 | 19.2 | 15.2 | 201.5 | 78.3 | |
| (kg ha−1) | 30 | 20.0 | 15.7 | 210.8 | 75.1 | |
| p-value | 0.11 | 0.23 | 0.47 | 0.60 | ||
| Timing of N application | ||||||
| Age (year) | 1 | 20.0 | 15.3 | 193.7 | 74.5 | |
| 2 | 19.9 | 15.5 | 211.0 | 73.1 | ||
| p-value | 0.92 | 0.51 | 0.42 | 0.74 | ||
| Lagan | Application of N at 1 year and weed control | |||||
| N fertiliser | 0 | 18.0 | 14.4 | 151.4 | 73.5 | |
| (kg ha−1) | 125 | 17.7 | 14.0 | 164.4 | 83.0 | |
| 250 | 18.2 | 14.0 | 155.9 | 77.5 | ||
| p-value | 0.54 | 0.31 | 0.36 | 0.01 | ||
| Weed control | <1 year (W0) | 17.6 | 14.0 | 146.0 | 76.2 | |
| Full (W1) | 18.2 | 14.3 | 168.5 | 79.8 | ||
| p-value | 0.15 | 0.27 | 0.01 | 0.14 | ||
| Application of P at 1 year with full weed control | ||||||
| P fertiliser | 0 | 18.3 | 14.4 | 171.5 | 78.5 | |
| (kg ha−1) | 30 | 18.7 | 14.7 | 165.1 | 73.1 | |
| p-value | 0.40 | 0.41 | 0.70 | 0.13 | ||
| Timing of N application | ||||||
| Age (year) | 1 | 18.1 | 14.1 | 173.8 | 79.9 | |
| 2 | 18.6 | 15.1 | 169.6 | 77.1 | ||
| p-value | 0.13 | 0.09 | 0.74 | 0.63 | ||
| Site | Variable | Level | Height, H | DBH, D | Volume, V |
|---|---|---|---|---|---|
| (m) | (cm) | (m3 ha−1) | |||
| Toman | Weed control | NWC | 11.1 a | 9.6 a | 56.2 a |
| (W) | MWC | 11.6 ab | 10.0 a | 63.0 ab | |
| FWC | 12.1 b | 10.9 b | 71.7 b | ||
| p-value | 0.048 | <0.001 | <0.001 | ||
| P fertiliser | 0 | 10.8 | 9.5 | 53.1 | |
| (kg ha−1) | 12 | 12.4 | 10.9 | 74.1 | |
| p-value | <0.001 | <0.001 | <0.001 | ||
| Caban | Weed control | NWC | 8.7 a | 5.9 a | 25.1 a |
| (W) | MWC | 11.9 b | 9.4 b | 68.6 b | |
| FWC | 12.8 b | 9.8 b | 78.4 b | ||
| p-value | <0.001 | <0.001 | <0.001 | ||
| P fertiliser | 0 | 9.7 | 7.1 | 39.4 | |
| (kg ha−1) | 33 | 12.6 | 9.6 | 75.3 | |
| p-value | <0.001 | <0.001 | <0.001 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Inail, M.A.; Hardiyanto, E.B.; Mendham, D.S.; Thaher, E. Growth Response to Weed Control and Fertilisation in Mid-Rotation Plantations of Eucalyptus pellita in South Sumatra, Indonesia. Forests 2021, 12, 1653. https://0-doi-org.brum.beds.ac.uk/10.3390/f12121653
Inail MA, Hardiyanto EB, Mendham DS, Thaher E. Growth Response to Weed Control and Fertilisation in Mid-Rotation Plantations of Eucalyptus pellita in South Sumatra, Indonesia. Forests. 2021; 12(12):1653. https://0-doi-org.brum.beds.ac.uk/10.3390/f12121653
Chicago/Turabian StyleInail, Maydra A., Eko B. Hardiyanto, Daniel S. Mendham, and Erlanda Thaher. 2021. "Growth Response to Weed Control and Fertilisation in Mid-Rotation Plantations of Eucalyptus pellita in South Sumatra, Indonesia" Forests 12, no. 12: 1653. https://0-doi-org.brum.beds.ac.uk/10.3390/f12121653