
Root Foraging Ability for Phosphorus in Different Genotypes Taxodium ‘Zhongshanshan’ and Their Parents under Phosphorus Deficiency

Abstract
:1. Introduction
2. Materials and Methods
2.1. Experimental Materials
2.2. Experimental Design
2.3. Plants Measurements
2.4. Statistical Analysis
3. Results
3.1. Interactive Effects of Experimental Materials and P Levels
3.2. Changes of Root Morphology
3.3. Changes of Plant Growth
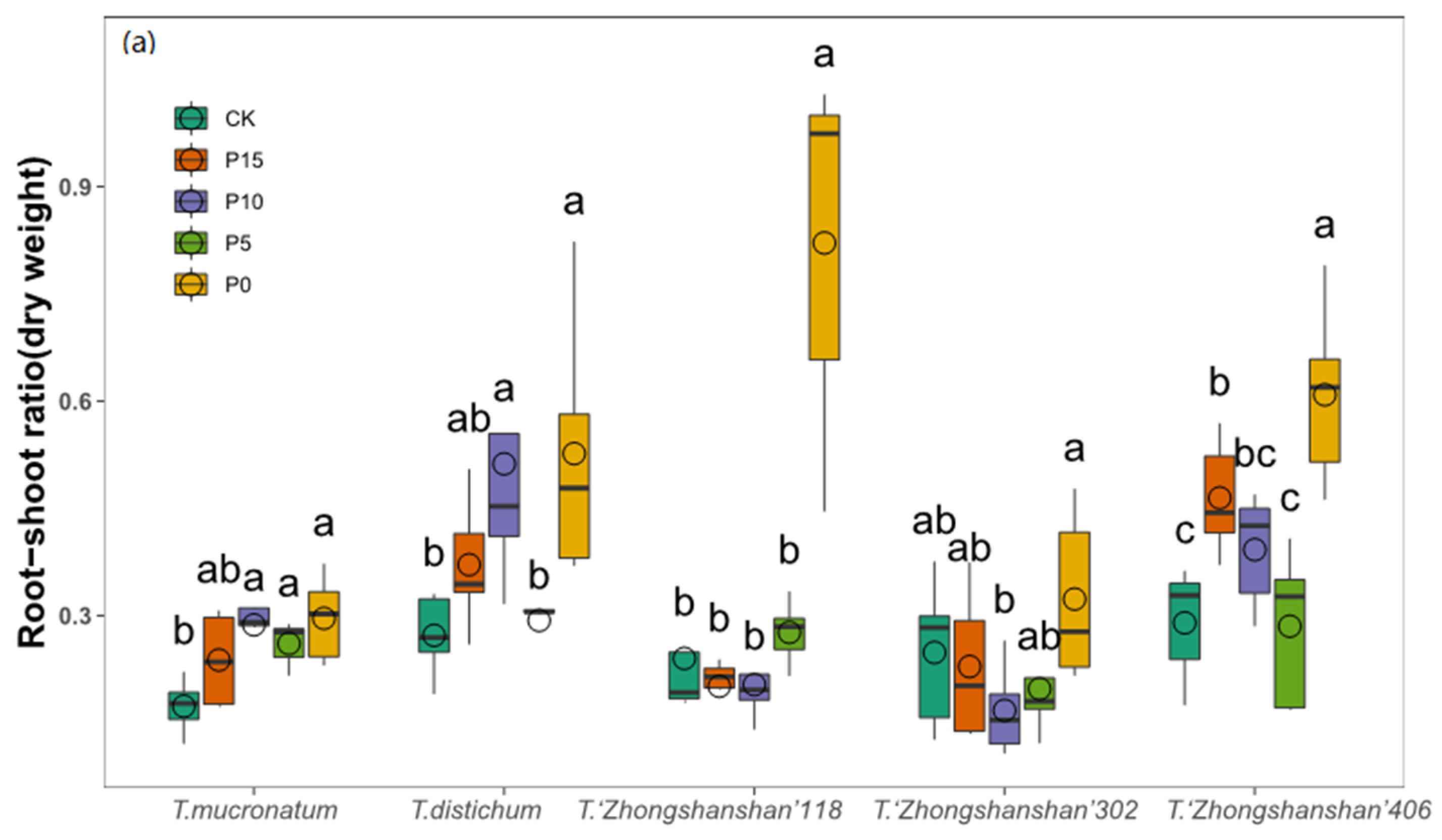
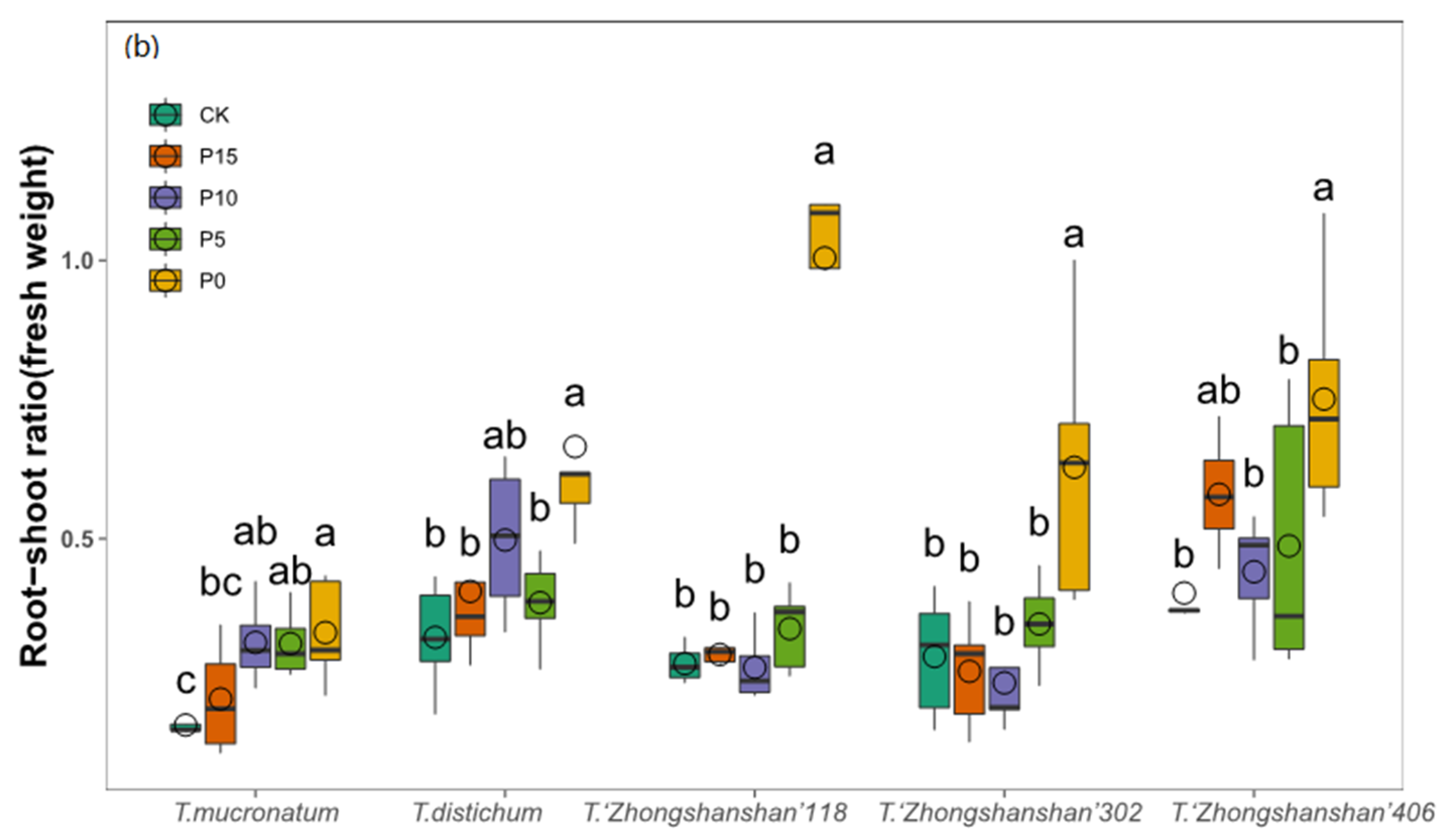
3.4. Changes of Root–Shoot Ratio
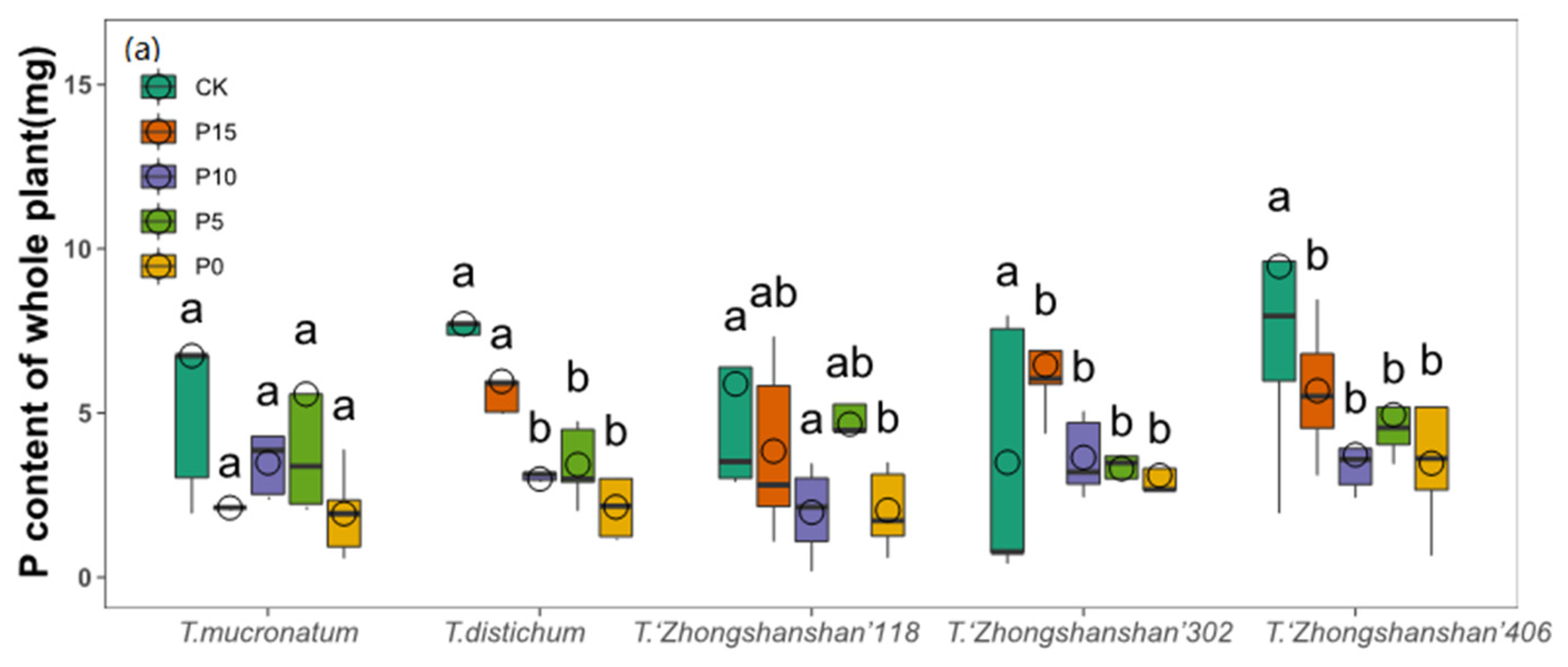
3.5. Changes of Plant P Contents
3.6. Correlation Analysis
3.7. Comprehensive Evaluation of Root P-Foraging Ability
3.8. Stepwise Regression Analysis
4. Discussions
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Kvakić, M.; Pellerin, S.; Ciais, P.; Achat, D.L.; Augusto, L.; Denoroy, P.; Gerber, J.S.; Goll, D.; Mollier, A.; Mueller, N.D.; et al. Quantifying the Limitation to World Cereal Production Due To Soil Phosphorus Status. Glob. Biogeochem. Cycles 2018, 32, 143–157. [Google Scholar] [CrossRef]
- Van der Salm, C.; Kros, J.; de Vries, W. Evaluation of different approaches to describe the sorption and desorption of phosphorus in soils on experimental data. Sci. Total Environ. 2016, 571, 292–306. [Google Scholar] [CrossRef]
- Xu, X.; Qiu, H.; Zhou, X. The absorption, Translocation and Metabolism of Phosphorus of Plant. J. Shandong Agric. Univ. 2001, 3, 397–400. [Google Scholar]
- Zhao, Q.; Zeng, D. Phosphorus cycling in terrestrial ecosystems and its controlling factors. Acta Phytoecol. Sin. 2005, 1, 153–163. [Google Scholar]
- Miguel, M.A.; Postma, J.A.; Lynch, J.P. Phene Synergism between Root Hair Length and Basal Root Growth Angle for Phosphorus Acquisition. Plant Physiol. 2015, 167, 1430–1439. [Google Scholar] [CrossRef]
- Yu, J.; Yin, D.; Wu, J.; Zhou, C.; Ma, X. A review of Adaptation Mechanism of Trees under low Phosphorus stress. World For. Res. 2007, 30, 18–23. [Google Scholar]
- Gu, X. Effects of Planting Methods and Nitrogen Application Rate on Soil Environment and Yield of Winter Rapeseed. Ph.D. Thesis, Northwest Agriculture and Forestry University, Xianyang, China, 2018. [Google Scholar]
- Cathcart, J.B. World phosphate reserves and resources. In The Role of Phosphorus in Agriculture; Khasawneh, F.E., Sample, E.C., Kamprath, E.J., Eds.; American Society of Agronomy: Madison, WI, USA, 1980; pp. 1–18. [Google Scholar]
- Li, Y.; Xiao, Z. China’s Forestland Soil Pollution, Degradation, Erosion Problems and Countermeasures. For. Econ. 2015, 37, 3–15. [Google Scholar]
- Kale, R.R.; Anila, M.; Swamy, H.M.; Bhadana, V.P.; Rani, C.V.D.; Senguttuvel, P.; Sundaram, R.M. Morphological and molecular screening of rice germplasm lines for low soil P tolerance.Journal of plant biochemistry and biotechnology. J. Plant Biochem. Biotechnol. 2020, 10, 1–12. [Google Scholar]
- Lambers, H.; Plaxton, W.C. Phosphorus:Back to the roots. Annu. Plant Rev. 2015, 48, 3–22. [Google Scholar]
- Da Silva, A.; Bruno, I.P.; Franzini, V.I.; Marcante, N.C.; Benitiz, L.; Muraoka, T. Phosphorus uptake efficiency, root morphology and architecture in Brazilian wheat cultivars. J. Radioanal. Nucl. Chem. 2016, 7, 1055–1063. [Google Scholar] [CrossRef]
- Yao, J.; Zhou, Z.; Chu, X. Effect of neighborhood competition on dry matter accumulation, nitrogen and phosphorus efficiency of three provenances of Schima superba in a heterogeneous nutrient environment. Acta Ecol. Sin. 2018, 38, 1780–1788. [Google Scholar]
- He, X.; Qi, B.; Wang, M.; Sun, Y. Differences in Biomass and Phosphorus Nutrition of Oats with Different Phosphorus Efficiency under Low Phosphorus Stress. Mol. Plant Breed. 2019, 38, 1780–1788. [Google Scholar]
- Xu, X.; Zhang, Y. Research progress on the root adaptation mechanism of plants under low phosphorus stress. Jiangsu J. Agric. Sci. 2018, 34, 1425–1429. [Google Scholar]
- Liu, P.; Wu, A.; Wang, J. Study on Phosphorus Use Efficiency and Phosphorus Remobilization Characteristics of Four Different Sorghum Genotypes. J. Shanxi Agric. Sci. 2018, 46, 344–349. [Google Scholar]
- Li, T.; Ye, D.; Zhang, X.; Guo, J. Research advances on response characteristics of plants to different forms of phosphorus. J. Plant Nutr. Fertil. 2017, 23, 1536–1546. [Google Scholar]
- Wu, W.; Wang, P.; Chen, N. Root foraging ability for phosphorus of different Chinese fir family seedlings under low phosphorus supply. J. Fujian Agric. For. Univ. 2019, 48, 174–181. [Google Scholar]
- Aziz, T.; Steffens, D.; Rahmatullah; Schubert, S. Variation in phosphorus efficiency among brassica cultivars II:Changes in root morphology and carboxylate exudation. J. Plant Nutr. 2011, 34, 2127–2138. [Google Scholar] [CrossRef]
- Wang, Z.; Hua, J.; Yin, Y. An Integrated Transcriptome and Proteome Analysis Reveals Putative Regulators of Adventitious Root Formation in Taxodium ‘Zhongshanshan’. Int. J. Mol. Sci. 2019, 20, 1225. [Google Scholar] [CrossRef] [Green Version]
- Yu, C.; Xu, S.; Yin, Y. Transcriptome analysis of the Taxodium ‘Zhongshanshan 405’ roots in response to salinity stress. Plant Physiol. Biochem. 2016, 100, 156–165. [Google Scholar] [CrossRef]
- Wang, Z.; Gu, C.; Xuan, L.; Hua, J.; Shi, Q.; Fan, W.; Yin, Y.; Yu, F. Identification of suitable reference genes in Taxodium ‘Zhongshanshan’ under abiotic stresses. Trees 2017, 31, 1519–1530. [Google Scholar] [CrossRef]
- Qi, B.; Yang, Y.; Yin, Y.; Xu, M.; Li, H. De novo sequencing, assembly, and analysis of the Taxodium‘Zhongshansa’roots and shoots transcriptome in response to short-term waterlogging. BMC Plant Biol. 2014, 14, 201. [Google Scholar] [CrossRef] [Green Version]
- Jiang, H.; Zhang, L.; Zhang, Q. Effects of waterlogging stress on photosynthesis and fluorescence of Taxodium hybrid. J. Fujian Agric. For. Univ. 2019, 48, 447–452. [Google Scholar]
- Shi, Q.; Yin, Y.; Wang, Z. Response in cuttings of Taxodium hybrid‘Zhongshanshan’and their parents to drought and re-hydration. Chin. J. Appl. Ecol. 2016, 27, 3435–3443. [Google Scholar]
- Yu, C.; Xu, J.; Lu, Z.; Yin, Y. A regional experiment of new cultivars of Taxodium mucronatum hybrid. South China For. Sci. 2015, 43, 27–30. [Google Scholar] [CrossRef]
- Yin, Y.; Chen, Y.; Chaoguang, Y.U. Breeding and popularization of new hybrid varieties of Taxodium Rich. Trees Sci. Technol. Achiev. 2013. [Google Scholar]
- Xu, J.; Yin, Y.; Hua, J. Analysis on Growth Characteristics of Three Taxodium mucronatum Hybrids. Mod. Agric. Sci. Technol. 2015, 24, 144–146, 150. [Google Scholar]
- Li, C.; Wang, J.; Chen, X. Experimental study on introduction of Taxodium hybrid‘Zhongshanshan’. J. Hebei For. Sci. Technol. 2012, 2, 9–12. [Google Scholar]
- Guo, J.; Shi, Q.; Xiong, Y.; Yin, Y.; Hua, J. Effects of salt-alkaline mixed stress on growth and photosynthetic characteristics of Taxodium hybrid ‘Zhongshanshan 406’. J. Nanjing For. Univ. 2019, 43, 61–68. [Google Scholar]
- Shen, J.; Mao, D. Methods of Plant Nutrition Research; China Agricultural University Press: Beijing, China, 2011. [Google Scholar]
- Yang, S.; Zhang, H.X.; Yang, X.Y.; Chen, Q.X.; Wu, H.W. Differential Growth Performance of Elaeagnus angustifolia Provenances under NaCl Stress. Sci. Silvae Sin. 2015, 51, 51–58. [Google Scholar]
- Chen, W.; OuYang, Z.; OuYang, S.; Li, Z.; He, Y. Effects of low phosphorus stress on growth and biomass of phyllostachys alba. Huannan For. Sci. Technol. 2020, 47, 25–30. [Google Scholar]
- Xu, J.; Li, Q.; Wu, W.; Rashid Muhammad, H.U.; Ma, X.; Wu, P. Effects of vertical phosphor competition on root growth and biomass allocation of Chinese fir. Acta Ecol. Sin. 2019, 39, 2071–2081. [Google Scholar]
- Yu, C.; Yin, Y.; Xu, J. Four hybrid varieties of Taxodium. Sci. Silvae Sin. 2011, 47, 181–182. (In Chinese) [Google Scholar]
- Hua, J.; Yin, Y.; Zhou, D.; Yu, C.; Xu, J. Effects of soil water conditions on growth and physiology of Taxodium ‘Zhongshanshan 406’. J. Ecol. Rural Environ. 2011, 6, 10. [Google Scholar]
- Yuan, J. Study on Adaptive Mechanism of Camellia Oleifera to Low-Phosphorus Environment. Ph.D. Thesis, Beijing Forestry University, Beijing, China, 2013. [Google Scholar]
- Li, B.; Xiao, K.; Li, Y. The genotypic differences in photosynthetic characteristics of wheat to low- phosphorous stress. J. Agric. Univ. Hebei. 2002, 1, 5–9. [Google Scholar]
- Haynes, R.J. Active ion uptake and maintenance of cationanion balance: A critical examination of their role in regulating rhizosphere pH. Plant Soil 1990, 126, 247–264. [Google Scholar] [CrossRef]
- Jones, D.L.; Darrah, P.R. Role of root derived organic acids in the mobilization of nutrients from the rhizosphere. Plant Soil 1994, 166, 247–257. [Google Scholar] [CrossRef]
- Hinsinger, P. Bioavailability of soil inorganic P in the rhizosphere as affected by root-induced chemical changes: A review. Plant Soil 2001, 237, 173–195. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Plants | Basal Diameter/mm | Plant Height/cm | Biomass/g |
|---|---|---|---|
| T.mucronatum | 3.21 ± 0.81 | 18.42 ± 2.7 | 1.87 ± 0.6 |
| T.distichum | 4.26 ± 0.9 | 23.31 ± 4.25 | 3.6 ± 1.76 |
| T.‘Zhongshanshan’118 | 3.39 ± 0.69 | 13.33 ± 3.75 | 2.42 ± 1.31 |
| T.‘Zhongshanshan’302 | 4.37 ± 1.00 | 19.07 ± 3.19 | 3.24 ± 1.79 |
| T.‘Zhongshanshan’406 | 4.68 ± 1.19 | 23.18 ± 4.22 | 4.82 ± 1.72 |
| Mean value | 4 ± 1.09 | 19.54 ± 5.13 | 3.21 ± 1.81 |
| Indicators | Materials | P Level | Materials × P Level | |||
|---|---|---|---|---|---|---|
| F | p | F | p | F | p | |
| Root length | 32.720 | 0.000 | 2.248 | 0.070 | 0.637 | 0.846 |
| Root surface area | 26.770 | 0.000 | 2.668 | 0.037 | 0.703 | 0.784 |
| Root volume | 27.036 | 0.000 | 0.886 | 0.476 | 1.232 | 0.259 |
| Net biomass | 13.512 | 0.000 | 7.496 | 0.000 | 1.308 | 0.209 |
| Net basal diameter | 7.316 | 0.000 | 1.284 | 0.282 | 2.106 | 0.014 |
| Net plant height | 5.103 | 0.001 | 15.109 | 0.000 | 2.571 | 0.002 |
| Root-shoot ratio (fresh weight) | 12.724 | 0.000 | 32.825 | 0.000 | 3.719 | 0.000 |
| Root-shoot ratio (dry weight) | 14.362 | 0.000 | 25.047 | 0.000 | 5.179 | 0.000 |
| P content of aboveground | 2.451 | 0.051 | 14.717 | 0.000 | 1.315 | 0.203 |
| P content of underground | 3.076 | 0.020 | 12.224 | 0.000 | 2.249 | 0.008 |
| P content of whole plant | 1.528 | 0.200 | 5.373 | 0.001 | 2.010 | 0.019 |
| Indicators | Measuring Range | T.Mucronatum | T.Distichum | T.‘Zhongshanshan’118 | T.‘Zhongshanshan’302 | T.‘Zhongshanshan’406 | |||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| F | p | F | p | F | p | F | p | F | p | ||
| Root length (cm) | 0 < Root length ≤ 0.500 | 0.593 | 0.672 | 0.973 | 0.445 | 1.29 | 0.311 | 2.086 | 0.121 | 3.609 | 0.023 |
| 0.500 < Root length ≤ 1.000 | 3.900 | 0.018 | 0.718 | 0.590 | 0.320 | 0.861 | 0.279 | 0.888 | 1.467 | 0.250 | |
| 1.000 < Root length ≤ 1.500 | 1.548 | 0.229 | 0.988 | 0.438 | 0.696 | 0.604 | 1.589 | 0.216 | 1.070 | 0.398 | |
| 1.500 < Root length ≤ 2.000 | 0.72 | 0.589 | 2.13 | 0.117 | 0.624 | 0.651 | 1.449 | 0.255 | 2.171 | 0.109 | |
| 2.000 < Root length ≤ 2.500 | 0.787 | 0.548 | 1.639 | 0.206 | 0.727 | 0.585 | 0.878 | 0.495 | 1.808 | 0.167 | |
| 2.500 < Root length ≤ 3.000 | 0.173 | 0.949 | 2.068 | 0.125 | 0.607 | 0.662 | 1.022 | 0.420 | 1.173 | 0.352 | |
| Root length > 3.000 | 0.668 | 0.622 | 1.632 | 0.207 | 0.785 | 0.550 | 0.986 | 0.438 | 1.807 | 0.167 | |
| Root surface area (cm2) | 0 < Root surface area ≤ 0.500 | 0.702 | 0.600 | 0.929 | 0.468 | 1.422 | 0.267 | 1.952 | 0.141 | 2.82 | 0.053 |
| 0.500 < Root surface area ≤ 1.000 | 3.829 | 0.019 | 0.743 | 0.574 | 0.317 | 0.863 | 0.275 | 0.890 | 1.418 | 0.264 | |
| 1.000 < Root surface area ≤ 1.500 | 1.435 | 0.261 | 0.963 | 0.45 | 0.698 | 0.604 | 1.611 | 0.210 | 1.168 | 0.355 | |
| 1.500 < Root surface area ≤ 2.000 | 0.731 | 0.582 | 2.125 | 0.117 | 0.644 | 0.638 | 1.449 | 0.255 | 2.140 | 0.113 | |
| 2.000 < Root surface area ≤ 2.500 | 0.778 | 0.553 | 1.646 | 0.204 | 0.716 | 0.592 | 0.851 | 0.510 | 1.798 | 0.169 | |
| 2.500 < Root surface area ≤ 3.000 | 0.161 | 0.955 | 2.019 | 0.133 | 0.598 | 0.669 | 1.002 | 0.430 | 1.179 | 0.350 | |
| Root surface area > 3.000 | 0.636 | 0.643 | 1.648 | 0.204 | 0.757 | 0.567 | 0.989 | 0.436 | 1.785 | 0.171 | |
| Root volume (cm3) | 0 < Root volume ≤ 0.500 | 0.941 | 0.462 | 0.892 | 0.488 | 1.383 | 0.279 | 1.827 | 0.163 | 2.105 | 0.118 |
| 0.500 < Root volume ≤ 1.000 | 3.717 | 0.021 | 0.76 | 0.564 | 0.315 | 0.864 | 0.291 | 0.881 | 1.352 | 0.286 | |
| 1.000 < Root volume ≤ 1.500 | 1.319 | 0.299 | 0.935 | 0.465 | 0.698 | 0.603 | 1.634 | 0.205 | 1.276 | 0.312 | |
| 1.500 < Root volume ≤ 2.000 | 0.743 | 0.575 | 2.117 | 0.118 | 0.663 | 0.625 | 1.453 | 0.254 | 2.110 | 0.117 | |
| 2.000 < Root volume ≤ 2.500 | 0.767 | 0.560 | 1.660 | 0.201 | 0.706 | 0.598 | 0.836 | 0.519 | 1.786 | 0.171 | |
| 2.500 < Root volume ≤ 3.000 | 0.151 | 0.960 | 1.968 | 0.140 | 0.588 | 0.675 | 0.988 | 0.437 | 1.184 | 0.348 | |
| Root volume > 3.000 | 0.603 | 0.665 | 1.679 | 0.196 | 0.736 | 0.580 | 0.993 | 0.434 | 1.762 | 0.176 | |
| Indicators | Net Plant Height | Net Biomass | Net Basal Diameter | Root Volume | Root Surface Area | Root Length | Root–Shoot Ratio (Fresh Weight) | Root–Shoot Ratio (Dry Weight) | P Content of Whole Plant | P Content of Aboveground | P Content of Underground |
|---|---|---|---|---|---|---|---|---|---|---|---|
| Net plant height | 1 | ||||||||||
| Net biomass | 0.635 ** | 1 | |||||||||
| Net basal diameter | 0.206 | 0.114 | 1 | ||||||||
| Root volume | 0.097 | 0.634 ** | 0.050 | 1 | |||||||
| Root surface area | 0.228 | 0.515 ** | 0.449 * | 0.453 * | 1 | ||||||
| Root length | −0.208 | 0.200 | −0.390 | 0.374 | 0.077 | 1 | |||||
| Root–shoot ratio (fresh weight) | −0.648 ** | −0.560 ** | 0.001 | −0.044 | −0.289 | −0.248 | 1 | ||||
| Root–shoot ratio (dry weight) | −0.651 ** | −0.509 ** | 0.041 | −0.099 | −0.245 | −0.313 | 0.913 ** | 1 | |||
| P content of whole plant | −0.492 * | 0.746 ** | −0.120 | 0.371 | 0.298 | 0.292 | −0.530 ** | −0.561 ** | 1 | ||
| P content of aboveground | 0.398 * | 0.753 ** | 0.216 | 0.595 ** | 0.515 ** | 0.469 * | −0..345 | −0.416 * | 0.828 ** | 1 | |
| P content of underground | 0.386 | 0.391 | −0.506 ** | 0.088 | −0.176 | −0.112 | −0.402 * | −0.274 | 0.501 * | 0.039 | 1 |
| Plants | Evaluation D | ||||
|---|---|---|---|---|---|
| P15 | P10 | P5 | P0 | Mean Value (D) | |
| T.mucronatum | 0.516 | 0.659 | 0.708 | 0.513 | 0.599 |
| T.distichum | 0.588 | 0.305 | 0.326 | 0.571 | 0.448 |
| T.‘Zhongshanshan’118 | 0.481 | 0.560 | 0.605 | 0.108 | 0.439 |
| T.‘Zhongshanshan’302 | 0.424 | 0.443 | 0.268 | 0.738 | 0.468 |
| T.‘Zhongshanshan’406 | 0.210 | 0.303 | 0.265 | 0.627 | 0.351 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Xie, R.; Hua, J.; Yin, Y.; Wan, F. Root Foraging Ability for Phosphorus in Different Genotypes Taxodium ‘Zhongshanshan’ and Their Parents under Phosphorus Deficiency. Forests 2021, 12, 215. https://0-doi-org.brum.beds.ac.uk/10.3390/f12020215
Xie R, Hua J, Yin Y, Wan F. Root Foraging Ability for Phosphorus in Different Genotypes Taxodium ‘Zhongshanshan’ and Their Parents under Phosphorus Deficiency. Forests. 2021; 12(2):215. https://0-doi-org.brum.beds.ac.uk/10.3390/f12020215
Chicago/Turabian StyleXie, Rongxiu, Jianfeng Hua, Yunlong Yin, and Fuxu Wan. 2021. "Root Foraging Ability for Phosphorus in Different Genotypes Taxodium ‘Zhongshanshan’ and Their Parents under Phosphorus Deficiency" Forests 12, no. 2: 215. https://0-doi-org.brum.beds.ac.uk/10.3390/f12020215