
From Acid Rain to Low Precipitation: The Role Reversal of Norway Spruce, Silver Fir, and European Beech in a Selection Mountain Forest and Its Implications for Forest Management

Abstract
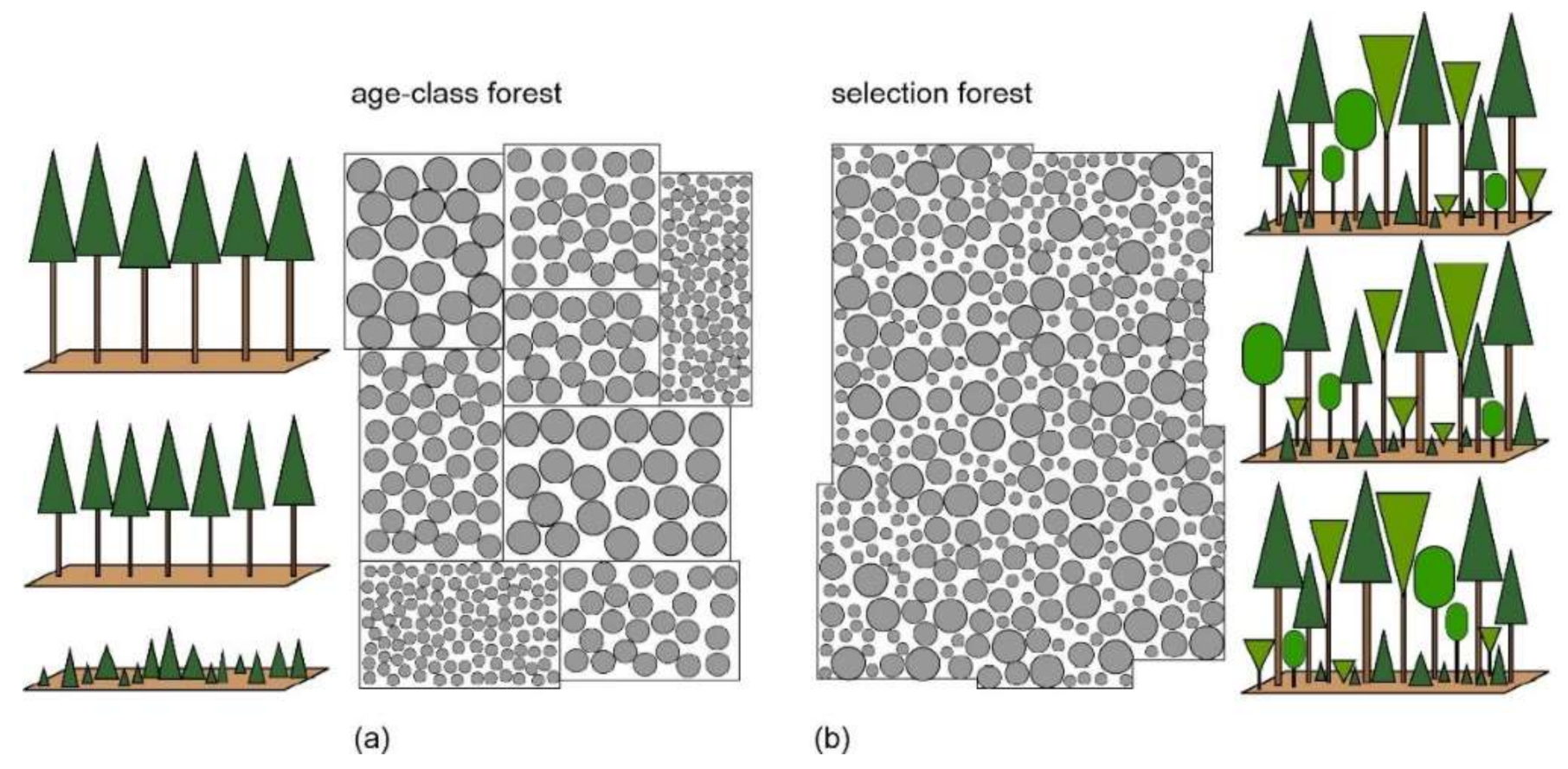
:1. Introduction
2. Materials and Methods
2.1. Material
2.2. Measurements, Metrics, Objective Variables
2.3. Methods
2.3.1. Stand Level Evaluation
2.3.2. Species Level Evaluation
2.3.3. Diameter Distribution Level
2.3.4. Tree Level Evaluation
2.4. Statistical Models
3. Results
3.1. Tree and Stand Characteristics
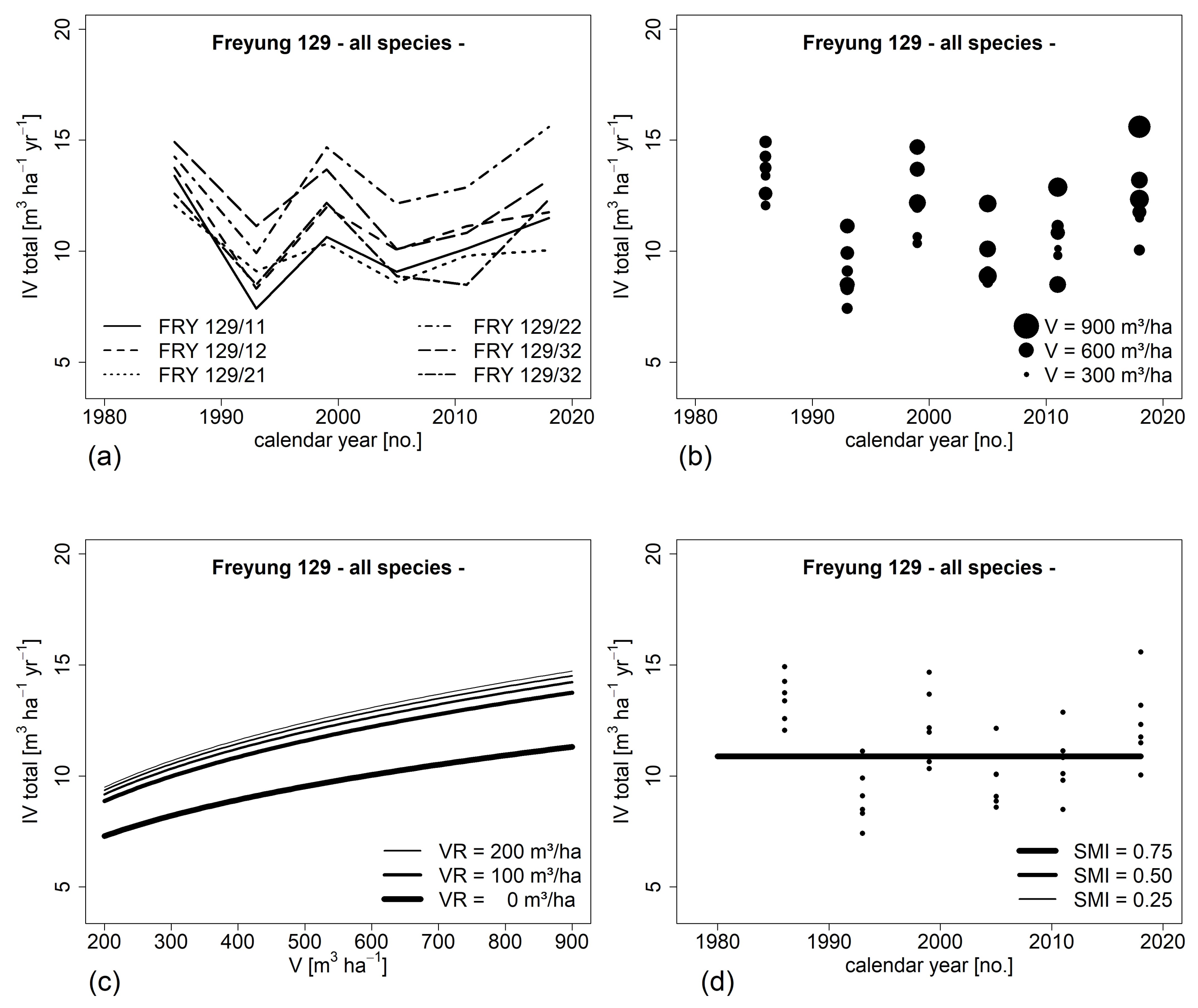
3.2. Development of Stand Level Growth (H I)
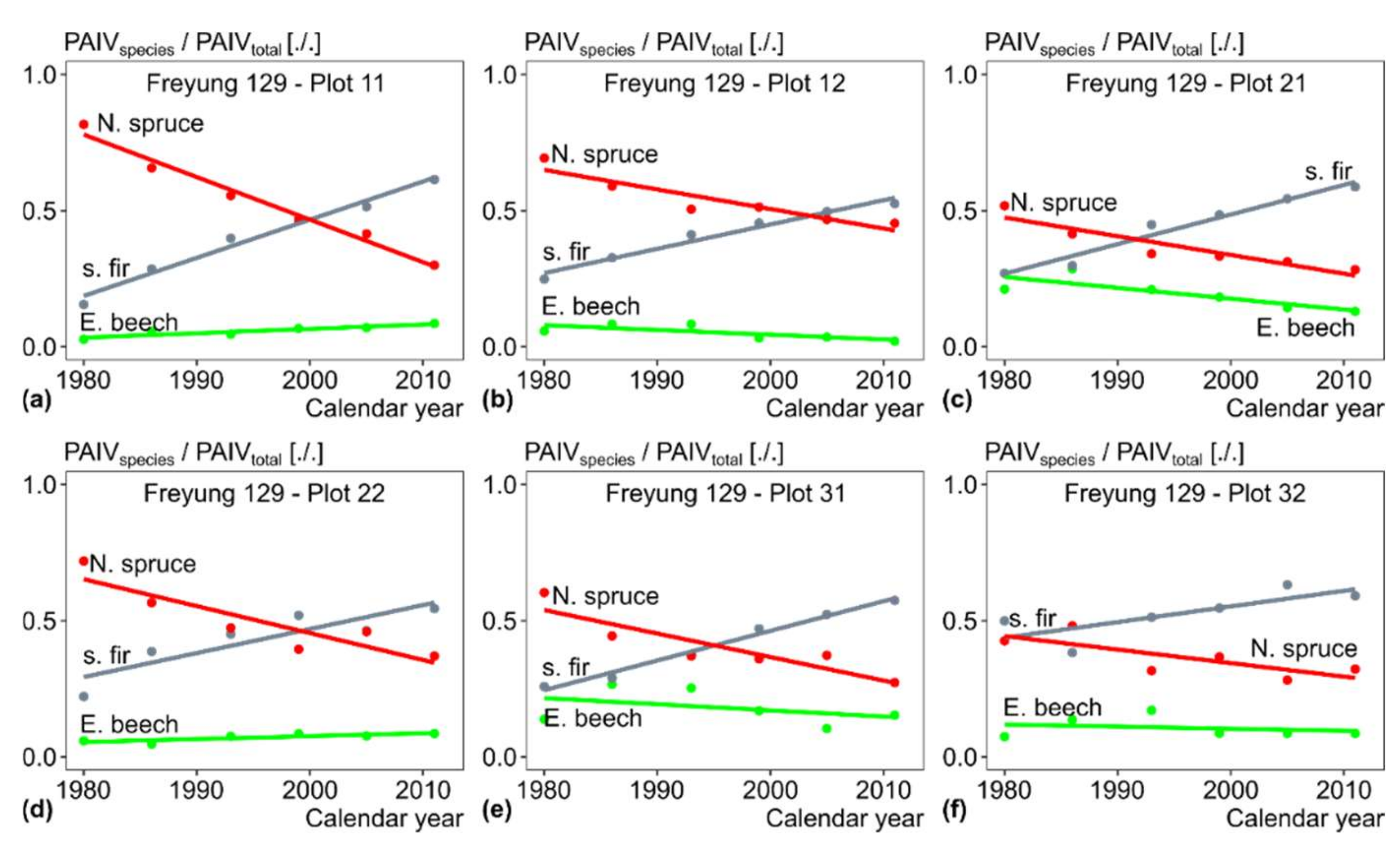
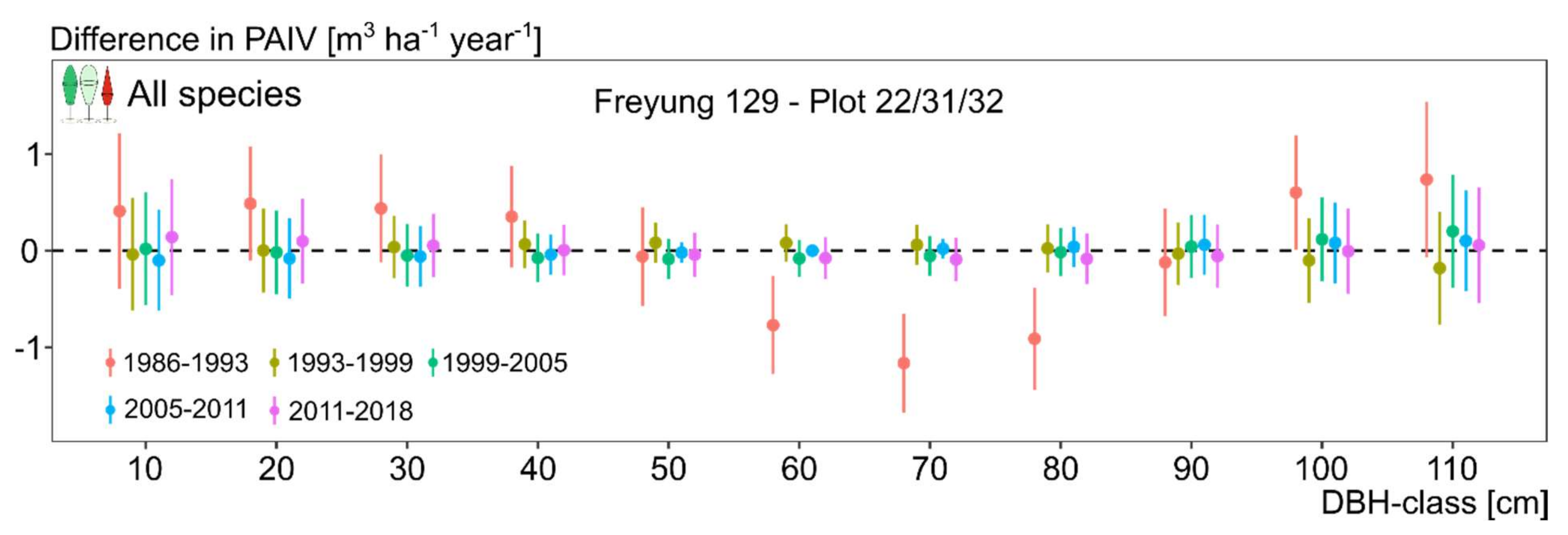
3.3. Size Class Contribution to Stand Growth over Time (H II)
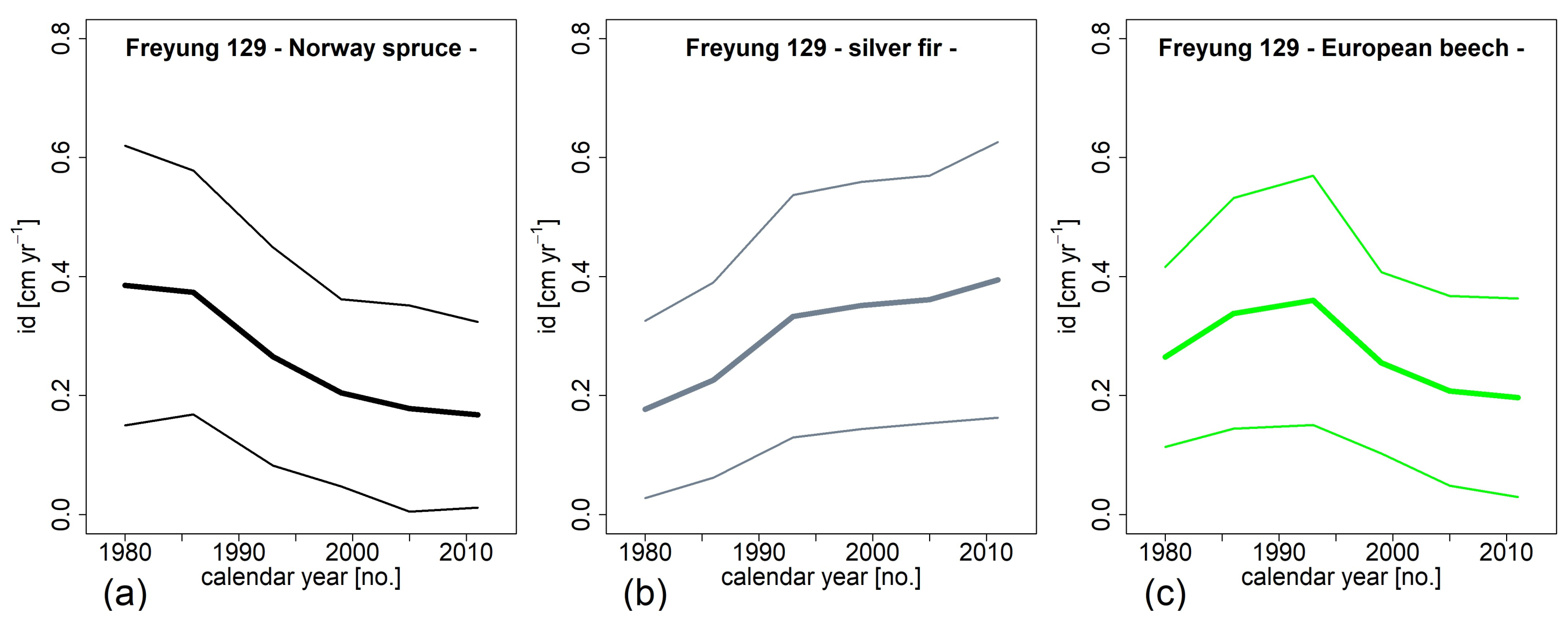
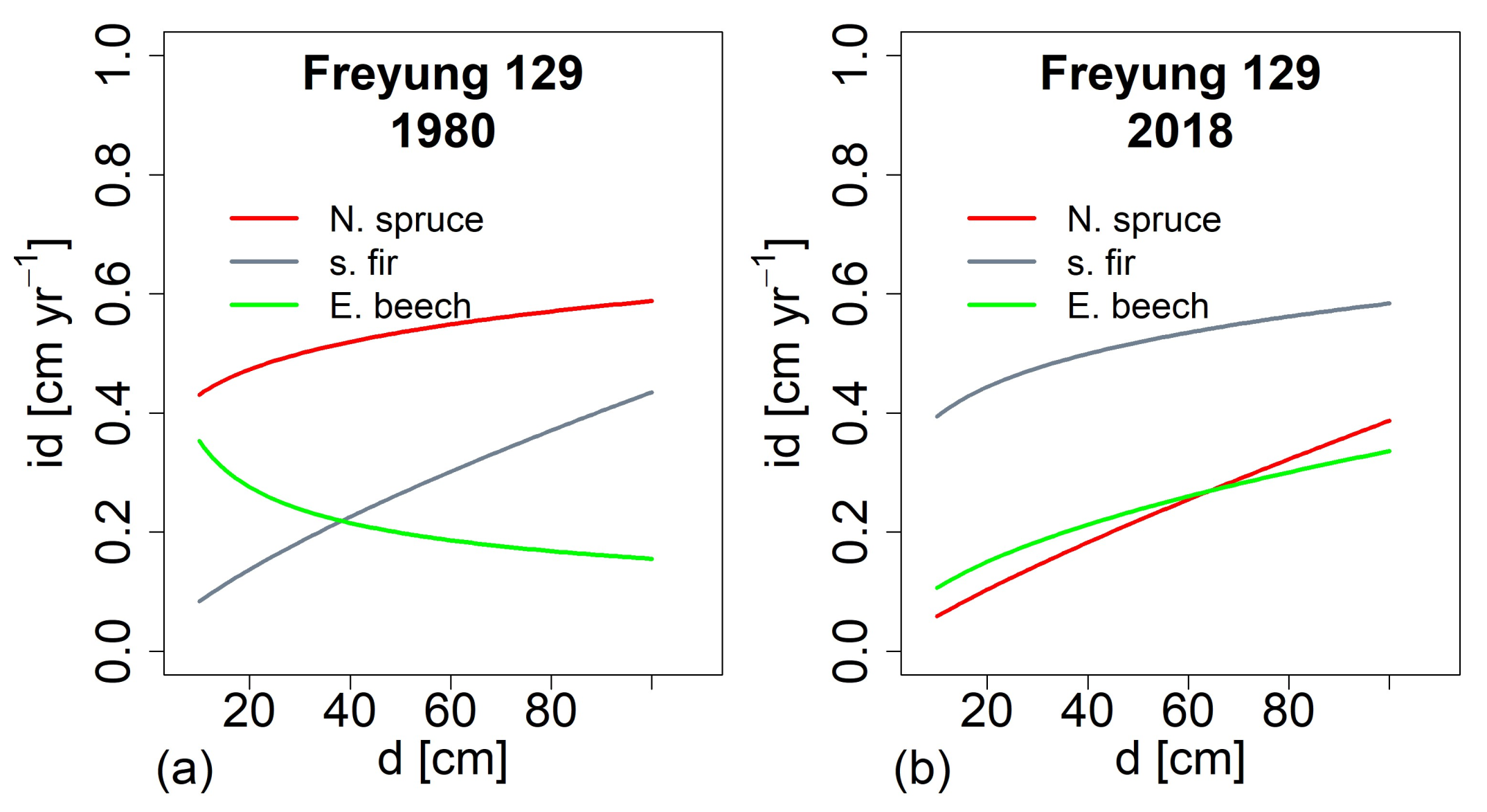
3.4. Change of Growth and Growth Partitioning from 1980ies to Present (H III)
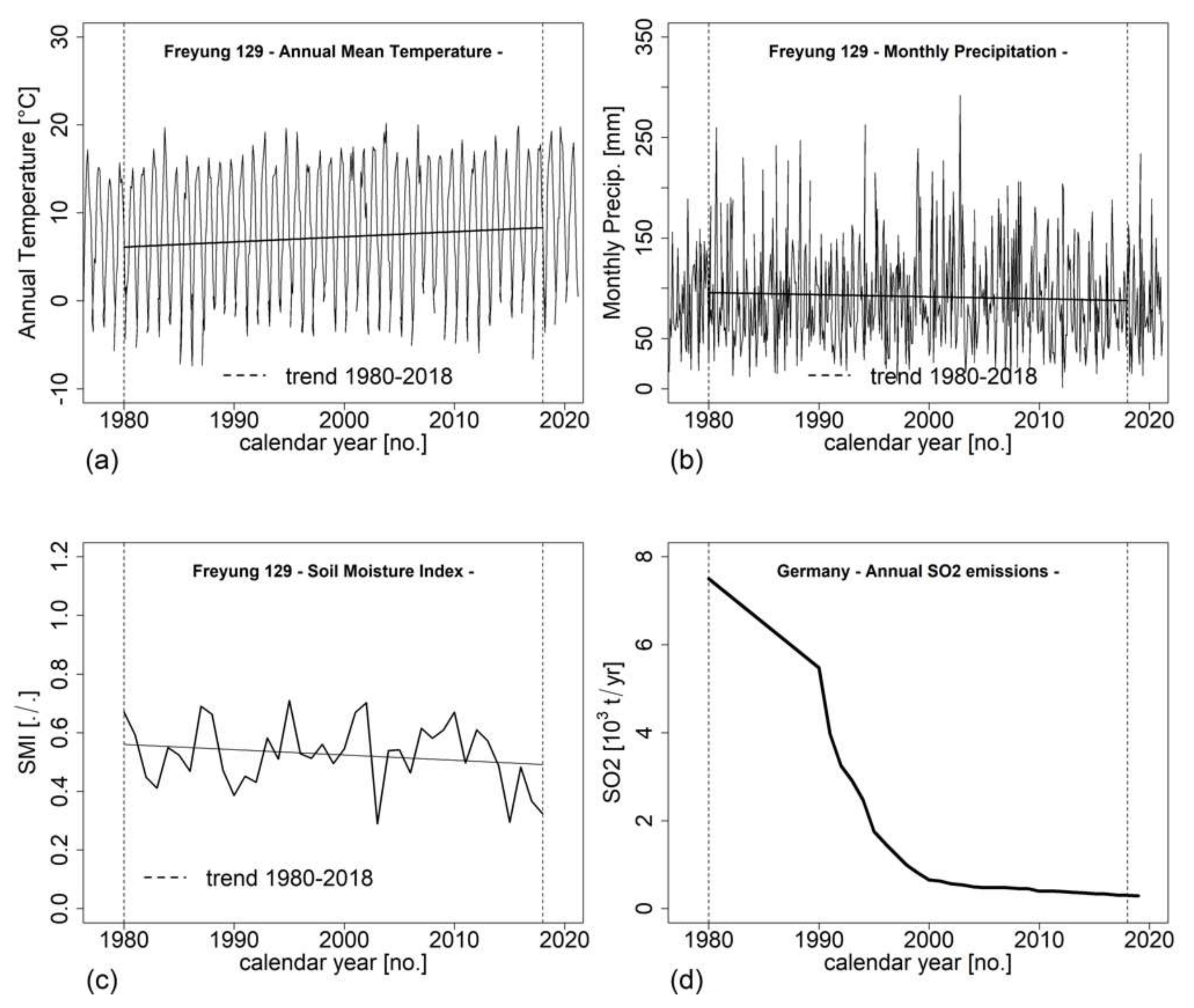
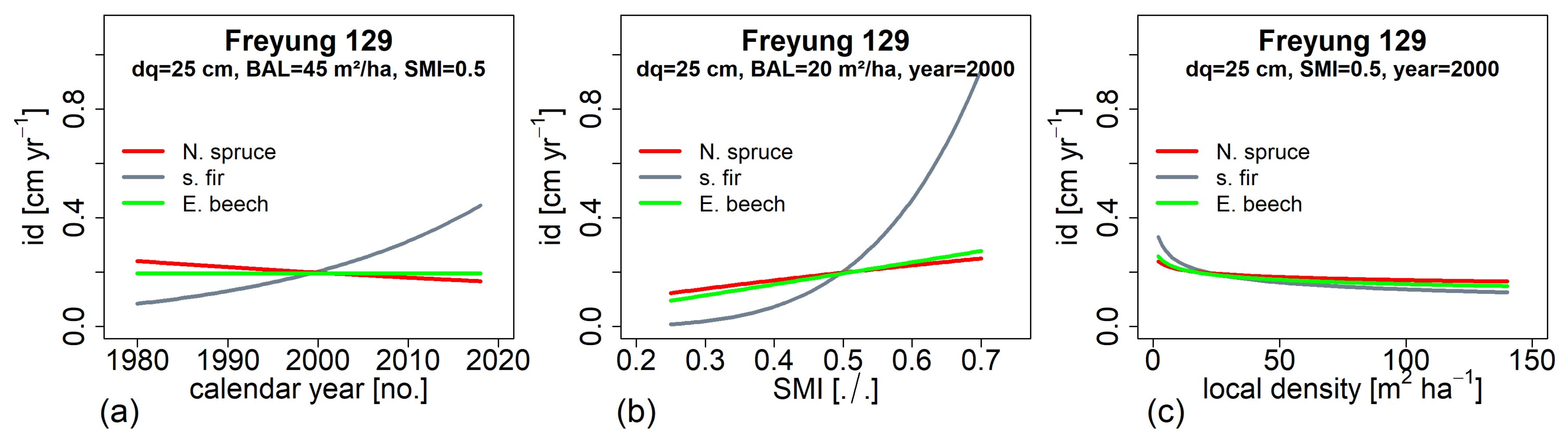
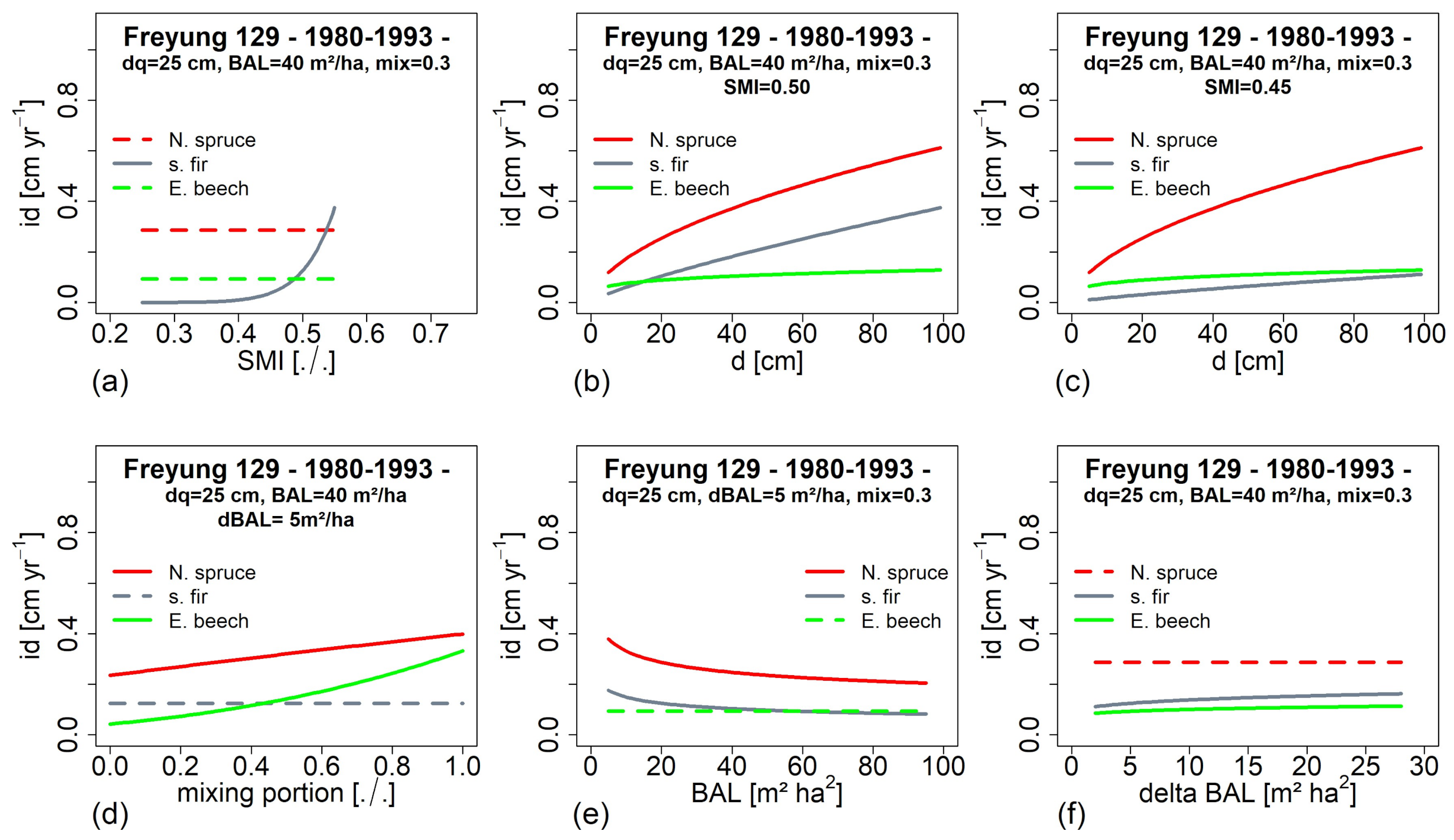
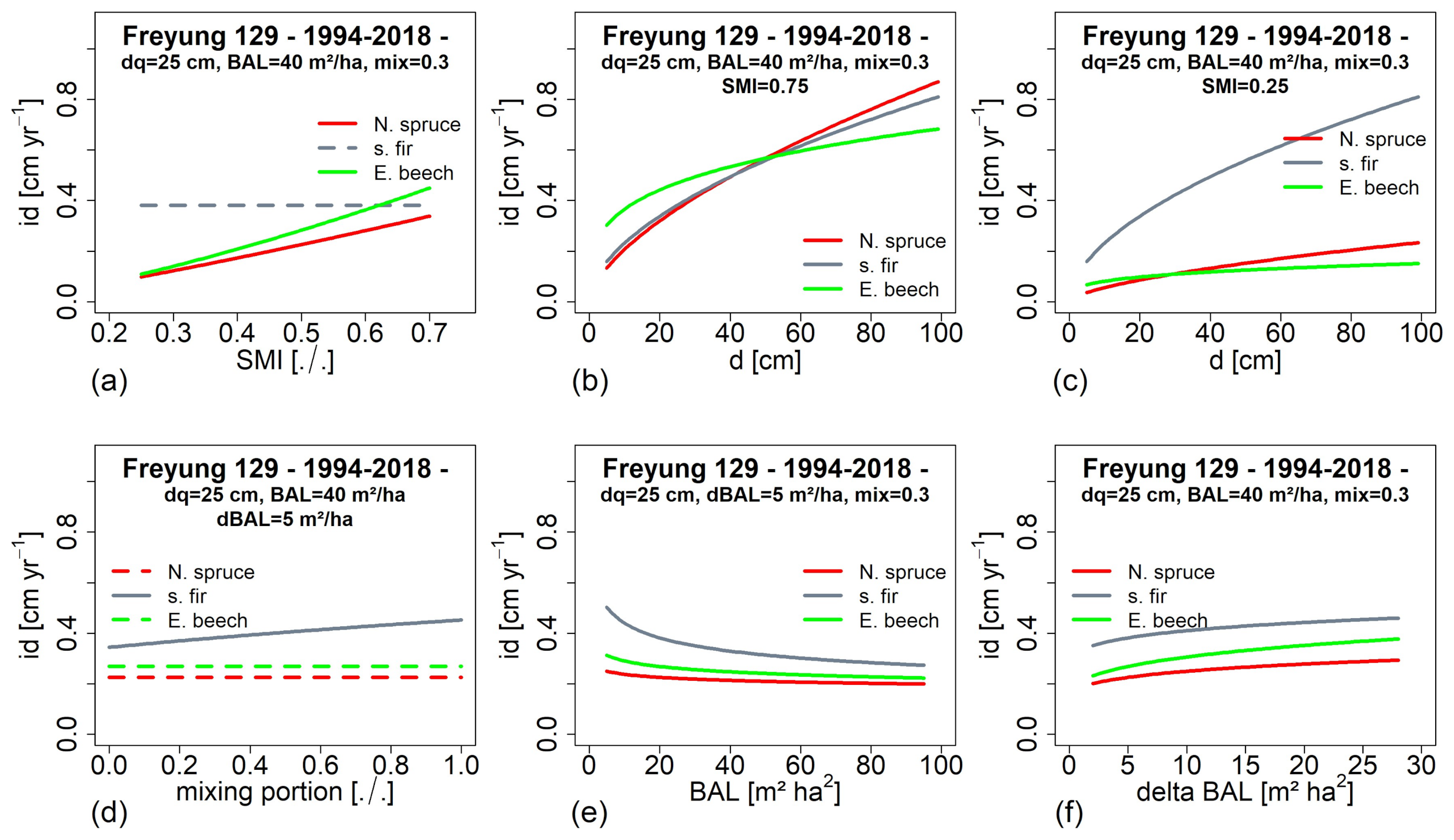
3.5. Impact of Environmental Factors on Growth Development (H IV)
3.6. Tree Growth Depending on Environemnatl Conditions in the Past and at Present (H V)
4. Discussion
4.1. Individual Tree Resilience versus Stand Level Resilience
4.2. Diversity Promotes Stability
4.3. Management of Diversity
4.4. Methodological Consideration
4.5. Relevance for Climate Smart Forest Management
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Reventlow, D.O.J.; Nord-Larsen, T.; Biber, P.; Hilmers, T.; Pretzsch, H. Simulating Conversion of Even-Aged Norway Spruce into Uneven-Aged Mixed Forest: Effects of Different Scenarios on Production, Economy and Heterogeneity. Eur. J. For. Res. 2021. [Google Scholar] [CrossRef]
- Hilmers, T.; Biber, P.; Knoke, T.; Pretzsch, H. Assessing Transformation Scenarios from Pure Norway Spruce to Mixed Uneven-Aged Forests in Mountain Areas. Eur. J. For. Res. 2020, 139, 567–584. [Google Scholar] [CrossRef] [Green Version]
- Hanewinkel, M.; Pretzsch, H. Modelling the Conversion from Even-Aged to Uneven-Aged Stands of Norway Spruce (Picea abies Karst.) with a Distance-Dependent Growth Simulator. For. Ecol. Manag. 2000, 134, 55–70. [Google Scholar] [CrossRef]
- Spathelf, P.; Bolte, A.; van der Maaten, E.C.D. Is Close-to-Nature Silviculture (CNS) an Adequate Concept to Adapt Forests to Climate Change? N Waldbehandlungskonzepts “Neue Multifunktionalität”. Landbauforsch. Appl. Agric. For. Res. 2015, 161–170. [Google Scholar] [CrossRef]
- Brang, P.; Spathelf, P.; Larsen, J.B.; Bauhus, J.; Boncčìna, A.; Chauvin, C.; Drössler, L.; García-Güemes, C.; Heiri, C.; Kerr, G.; et al. Suitability of Close-to-Nature Silviculture for Adapting Temperate European Forests to Climate Change. For. Int. J. For. Res. 2014, 87, 492–503. [Google Scholar] [CrossRef] [Green Version]
- Puettmann, K.J.; Coates, K.D.; Messier, C.C. A Critique of Silviculture: Managing for Complexity; Island Press: Washington, DC, USA, 2012; ISBN 978-1-61091-123-8. [Google Scholar]
- Walentowski, H. Handbuch der Natürlichen Waldgesellschaften Bayerns: Ein auf Geobotanischer Grundlage Entwickelter Leitfaden für die Praxis in Forstwirtschaft und Naturschutz; Geobotanica-Verlag: Freising, Germany, 2004. [Google Scholar]
- Schütz, J.-P.; Diez, C. Der Plenterbetrieb: Unterlage zur Vorlesung Waldbau III (Waldverjüngung) und zu SANASILVA-Fortbildungskursen; ETH: Zurich, Switzerland, 1989. [Google Scholar]
- Pretzsch, H. Die Fichten-Tannen-Buchen-Plenterwaldversuche in den ostbayerischen Forstämtern Freyung und Bodenmais. Forstarchiv 1985, 56, 3–9. [Google Scholar]
- Dittmar, O. Untersuchungen im Buchen-Plenterwald Keula. Forst Holz. 1990, 45, 419–423. [Google Scholar]
- Reisch, J. Das ertragskundliche Verhalten eines Fichtenbestandes auf Hochmoor im Forstamt St. Andreasberg (Harz). Forstw. Cbl. 1950, 69, 466–482. [Google Scholar] [CrossRef]
- Indermühle, M.P. Struktur-, Alters- und Zuwachsuntersuchungen in Einem Fichten-Plenterwald der Subalpinen Stufe: (Sphagno-Piceetum Calamagrostietosum Villosae); ETH: Zurich, Switzerland, 1978; p. 98S. [Google Scholar]
- Guldin, J.M.; Bragg, D.C.; Zingg, A. Plentering with pines—Results from the United States. Schweiz. Z. Forstwes. 2017, 168, 75–83. [Google Scholar] [CrossRef] [Green Version]
- Yamahata, K. Untersuchungen Über Den Plenterwaid von Kiefern (P. thunbergii). J. Jpn. For. Soc. 1965, 47, 377–383. [Google Scholar]
- Mayer, H. Waldbau: Auf soziologisch-ökologischer Grundlage; Fischer: Stuttgart, Germany, 1980. [Google Scholar]
- Schütz, J.-P. Sylviculture 2: La Gestion Des Forêts Irrégulières et Mélangées; PPUR Presses Polytechniques et Universitaires Romandes: Lausann, Switzerland, 1997. [Google Scholar]
- Pretszch, H. Transitioning monocultures to complex forest stands in Central Europe: Principles and practice. In Achieving Sustainable Management of Boreal and Temperate Forests; Stanturf, J.A., Ed.; Burleigh Dodds Science Publishing: Cambridge, UK, 2019; pp. 355–396. ISBN 978-1-78676-292-4. [Google Scholar]
- Schmidt-Vogt, H. Die Fichte, Band, I. Taxonomie. Verbreitung. Morphologie. Ökologie. Waldgesellschaft; Verlag Paul Parey: Hamburg/Berlin, Germany, 1977; Volume 647. [Google Scholar]
- Elling, W.; Heber, U.; Polle, A.; Beese, F. Schädigung von Waldökosystemen. Auswirkungen Anthropogener Umweltänderungen und Schutzmaßnahmen; Elsevier: München, Germany, 2007. [Google Scholar]
- Pretzsch, H.; Grams, T.; Häberle, K.H.; Pritsch, K.; Bauerle, T.; Rötzer, T. Growth and Mortality of Norway Spruce and European Beech in Monospecific and Mixed-Species Stands under Natural Episodic and Experimentally Extended Drought. Results of the KROOF Throughfall Exclusion Experiment. Trees 2020, 34, 957–970. [Google Scholar] [CrossRef]
- Grams, T.E.E.; Hesse, B.D.; Gebhardt, T.; Weikl, F.; Rötzer, T.; Kovacs, B.; Hikino, K.; Hafner, B.D.; Brunn, M.; Bauerle, T.; et al. The Kroof Experiment: Realization and Efficacy of a Recurrent Drought Experiment plus Recovery in a Beech/Spruce Forest. Ecosphere 2021, 12, e03399. [Google Scholar] [CrossRef]
- Liziniewicz, M. The Development of Beech in Monoculture and Mixtures; SLU, Southern Swedish Forest Research Centre: Alnarp, Sweden, 2009. [Google Scholar]
- Kraj, W.; Sztorc, A. Genetic Structure and Variability of Phenological Forms in the European Beech (Fagus sylvatica L.). Ann. For. Sci. 2009, 66, 1–7. [Google Scholar] [CrossRef] [Green Version]
- Remmert, H. The Mosaic-Cycle Concept of Ecosystems—An Overview. In The Mosaic-Cycle Concept of Ecosystems; Remmert, H., Ed.; Ecological Studies; Springer: Berlin/Heidelberg, Germany, 1991; Volume 85, pp. 1–21. ISBN 978-3-642-75652-8. [Google Scholar]
- Fischer, A. Vegetation dynamics in european beech forests. Ann. Bot. 1997, 55. [Google Scholar] [CrossRef]
- Rothe, A.; Kreutzer, K.; Küchenhoff, H. Influence of Tree Species Composition on Soil and Soil Solution Properties in Two Mixed Spruce-Beech Stands with Contrasting History in Southern Germany. Plant Soil 2002, 240, 47–56. [Google Scholar] [CrossRef]
- Schmid, I. The Influence of Soil Type and Interspecific Competition on the Fine Root System of Norway Spruce and European Beech. Basic Appl. Ecol. 2002, 3, 339–346. [Google Scholar] [CrossRef]
- Goisser, M.; Geppert, U.; Rötzer, T.; Paya, A.; Huber, A.; Kerner, R.; Bauerle, T.; Pretzsch, H.; Pritsch, K.; Häberle, K.; et al. Does Belowground Interaction with Fagus Sylvatica Increase Drought Susceptibility of Photosynthesis and Stem Growth in Picea Abies? For. Ecol. Manag. 2016, 375, 268–278. [Google Scholar] [CrossRef]
- Nagel, T.A.; Svoboda, M.; Kobal, M. Disturbance, Life History Traits, and Dynamics in an Old-Growth Forest Landscape of Southeastern Europe. Ecol. Appl. 2014, 24, 663–679. [Google Scholar] [CrossRef]
- Leuschner, C.; Ellenberg, H. Ecology of Central European Forests: Vegetation Ecology of Central Europe; Springer: New York, NY, USA, 2017; Volume 1. [Google Scholar]
- Schmidt-Vogt, H. Untersuchungen zur Bedeutung des Lichtfaktors bei Femelschlagverjüngung von Tannen-Buchen-Fichten-Wäldern im westlichen Hochschwarzwald. Forstw. Cbl. 1972, 91, 238–247. [Google Scholar] [CrossRef]
- Magin, R.; Mayer, H. Struktur und Leistung Mehrschichtiger Mischwälder in den Bayerischen Alpen; Mitteilungen aus der Staatsforstverwaltung Bayerns: Munich, Germany, 1959; p. 30. [Google Scholar]
- Preuhsler, T. Ertragskundliche Merkmale oberbayerischer Bergmischwald-Verjüngungsbestände auf kalkalpinen Standorten im Forstamt Kreuth. Forstw. Cbl. 1981, 100, 313–345. [Google Scholar] [CrossRef]
- Bachofen, H. Gleichgewicht, Struktur und Wachstum in Plenterbeständen| Equilibrium, Structure and Increment in Selection Forest Stands. Schweiz. Z. Forstwes. 1999, 150, 157–170. [Google Scholar] [CrossRef] [Green Version]
- Knoke, T. The Economics of Continuous Cover Forestry. In Continuous Cover Forestry; Pukkala, T., von Gadow, K., Eds.; Managing Forest Ecosystems; Springer Netherlands: Dordrecht, The Netherlands, 2012; pp. 167–193. ISBN 978-94-007-2202-6. [Google Scholar]
- Knoke, T. Zur finanziellen Attraktivität von Dauerwaldwirtschaft und Überführung: Eine Literaturanalyse|On the Financial Attractiveness of Continuous Cover Forest Management and Transformation: A Review. Schweiz. Z. Forstwes. 2009, 160, 152–161. [Google Scholar] [CrossRef]
- Yachi, S.; Loreau, M. Biodiversity and Ecosystem Productivity in a Fluctuating Environment: The Insurance Hypothesis. Proc. Natl. Acad. Sci. USA 1999, 96, 1463–1468. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kunert, N.; Cárdenas, A.M. Are Mixed Tropical Tree Plantations More Resistant to Drought than Monocultures? Forests 2015, 6, 2029–2046. [Google Scholar] [CrossRef] [Green Version]
- Merlin, M.; Perot, T.; Perret, S.; Korboulewsky, N.; Vallet, P. Effects of Stand Composition and Tree Size on Resistance and Resilience to Drought in Sessile Oak and Scots Pine. For. Ecol. Manag. 2015, 339, 22–33. [Google Scholar] [CrossRef] [Green Version]
- D’Amato, A.W.; Bradford, J.B.; Fraver, S.; Palik, B.J. Forest Management for Mitigation and Adaptation to Climate Change: Insights from Long-Term Silviculture Experiments. For. Ecol. Manag. 2011, 262, 803–816. [Google Scholar] [CrossRef]
- Coll, L.; Ameztegui, A.; Collet, C.; Löf, M.; Mason, B.; Pach, M.; Verheyen, K.; Abrudan, I.; Barbati, A.; Barreiro, S.; et al. Knowledge Gaps about Mixed Forests: What Do European Forest Managers Want to Know and What Answers Can Science Provide? For. Ecol. Manag. 2018, 407, 106–115. [Google Scholar] [CrossRef] [Green Version]
- Hilmers, T.; Avdagić, A.; Bartkowicz, L.; Bielak, K.; Binder, F.; Bončina, A.; Dobor, L.; Forrester, D.I.; Hobi, M.L.; Ibrahimspahić, A.; et al. The Productivity of Mixed Mountain Forests Comprised of Fagus Sylvatica, Picea Abies, and Abies Alba across Europe. Forestry 2019, 92, 512–522. [Google Scholar] [CrossRef] [Green Version]
- Menz, F.C.; Seip, H.M. Acid Rain in Europe and the United States: An Update. Environ. Sci. Policy 2004, 7, 253–265. [Google Scholar] [CrossRef]
- Elkin, C.; Giuggiola, A.; Rigling, A.; Bugmann, H. Short- and Long-Term Efficacy of Forest Thinning to Mitigate Drought Impacts in Mountain Forests in the European Alps. Ecol. Appl. 2015, 25, 1083–1098. [Google Scholar] [CrossRef]
- Zohner, C.M.; Mo, L.; Renner, S.S.; Svenning, J.-C.; Vitasse, Y.; Benito, B.M.; Ordonez, A.; Baumgarten, F.; Bastin, J.-F.; Sebald, V.; et al. Late-Spring Frost Risk between 1959 and 2017 Decreased in North America but Increased in Europe and Asia. Proc. Natl. Acad. Sci. USA 2020, 117, 12192–12200. [Google Scholar] [CrossRef]
- Pretzsch, H.; Hilmers, T.; Biber, P.; Avdagić, A.; Binder, F.; Bončina, A.; Bosela, M.; Dobor, L.; Forrester, D.I.; Lévesque, M.; et al. Evidence of Elevation-Specific Growth Changes of Spruce, Fir, and Beech in European Mixed Mountain Forests during the Last Three Centuries. Can. J. For. Res. 2020. [Google Scholar] [CrossRef]
- Lindner, M.; Maroschek, M.; Netherer, S.; Kremer, A.; Barbati, A.; Garcia-Gonzalo, J.; Seidl, R.; Delzon, S.; Corona, P.; Kolström, M.; et al. Climate Change Impacts, Adaptive Capacity, and Vulnerability of European Forest Ecosystems. For. Ecol. Manag. 2010, 259, 698–709. [Google Scholar] [CrossRef]
- Knoke, T. Analysis and optimization of wood production in a selection forest—On forest management planning in uneven-aged forests. Forstl. Forsch. Munch. 1998, 170, 182. [Google Scholar]
- Knoke, T. Zur betriebswirtschaftlichen Optimierung der Vorratshöhe in einem Plenterwald. Forst Holz. 1999, 483–488. Available online: https://mediatum.ub.tum.de/doc/625061/625061.pdf (accessed on 2 July 2021).
- Oberdorfer, E. Pflanzensoziologische Exkursionsflora für Deutschland und die angrenzenden Gebiete. Eugen Ulm. Stuttg. Pflanzengeogr. Angaben Florenelemente 1970, 3, 18–21. [Google Scholar]
- Uhl, E.; Ammer, C.; Spellmann, H.; Schölch, M.; Pretzsch, H. Zuwachstrend und Stressresilienz von Tanne und Fichte im Vergleich. Allg. Forst Jagdztg. 2013, 11–12, 278–292. [Google Scholar]
- Rothe, A.; Dittmar, C.; Zang, C. Tanne–Vom Sorgenkind Zum Hoffnungsträger. LWF Wissen 2011, 66, 59–63. [Google Scholar]
- DWD Climate Data Center (CDC). Raster der Monatsmittel der Lufttemperatur (2m) Für Deutschland, Version v1.0; Deutscher Wetterdienst: Offenbach am Main, Germany; Available online: https://opendata.dwd.de/climate_environment/CDC/grids_germany/monthly/air_temperature_mean/ (accessed on 15 April 2021).
- DWD Climate Data Center (CDC). Raster der Monatssumme der Niederschlagshöhe Für Deutschland, Version v1.0; Deutscher Wetterdienst: Offenbach am Main, Germany; Available online: https://opendata.dwd.de/climate_environment/CDC/grids_germany/monthly/precipitation/ (accessed on 15 April 2021).
- Zink, M.; Samaniego, L.; Kumar, R.; Thober, S.; Mai, J.; Schäfer, D.; Marx, A. The German Drought Monitor. Environ. Res. Lett. 2016, 11, 074002. [Google Scholar] [CrossRef]
- Umweltbundesamt. Daten zur Umwelt. Der Zustand der Umwelt in Deutschland.; Erich Schmidt Verlag: Berlin, Germany, 2005. [Google Scholar]
- Umweltbundesamt. Nationale Tabellen für die Deutsche Berichterstattung Atmosphärischer Emissionen Seit 1990, Emissionsentwicklung 1990 sis 2018 (Stand Februar 2020). Available online: https://www.umweltbundesamt.de/daten/luft/luftschadstoff-emissionen-in-deutschland/schwefeldioxid-emissionen#entwicklung-seit-1990 (accessed on 15 April 2021).
- Biber, P. Continuity by Flexibility-Standardised Data Evaluation within a Scientific Growth and Yield Information System. Allg. Forst Jagdztg. 2013, 184, 167–177. [Google Scholar]
- Pretzsch, H. Forest Dynamics, Growth, and Yield. In Forest Dynamics, Growth and Yield: From Measurement to Model; Pretzsch, H., Ed.; Springer: Berlin/Heidelberg, Germany, 2009; pp. 1–39. ISBN 978-3-540-88307-4. [Google Scholar]
- Johann, K. DESER-Norm 1993. Normen der Sektion Ertragskunde im Deutschen Verband Forstlicher Forschungsanstalten zur Aufbereitung von Waldwachstumskundlichen Dauerversuchen. Proc. Dt. Verb. Forstl. Forsch. Sek Ertragskd Unterreichenbach-Kapfenhardt 1993, 96–104. [Google Scholar]
- Wood, S.N. Generalized Additive Models: An Introduction with R, 2nd ed.; Chapman and Hall/CRC: Boca Raton, FL, USA, 2017. [Google Scholar]
- Pommerening, A.; Stoyan, D. Edge-Correction Needs in Estimating Indices of Spatial Forest Structure. Can. J. For. Res. 2011. [Google Scholar] [CrossRef] [Green Version]
- Radtke, P.J.; Burkhart, H.E. A Comparison of Methods for Edge-Bias Compensation. Can. J. For. Res. 2011. [Google Scholar] [CrossRef]
- Prodan, M. Forest Biometrics; Pergamon Press: Oxford, UK, 1968. [Google Scholar]
- Pretzsch, H.; del Río, M. Density Regulation of Mixed and Mono-Specific Forest Stands as a Continuum: A New Concept Based on Species-Specific Coefficients for Density Equivalence and Density Modification. For. Int. J. For. Res. 2020, 93, 1–15. [Google Scholar] [CrossRef]
- Dirnberger, G.; Sterba, H.; Condés, S.; Ammer, C.; Annighöfer, P.; Avdagić, A.; Bielak, K.; Brazaitis, G.; Coll, L.; Heym, M.; et al. Species Proportions by Area in Mixtures of Scots Pine (Pinus sylvestris L.) and European Beech (Fagus sylvatica L.). Eur. J. For. Res. 2017, 136, 171–183. [Google Scholar] [CrossRef] [Green Version]
- Pretzsch, H.; Zenner, E.K. Toward Managing Mixed-Species Stands: From Parametrization to Prescription. For. Ecosyst. 2017, 4, 19. [Google Scholar] [CrossRef] [Green Version]
- Pretzsch, H.; Biber, P. Tree Species Mixing Can Increase Maximum Stand Density. Can. J. For. Res. 2016, 46, 1179–1193. [Google Scholar] [CrossRef] [Green Version]
- Samaniego, L.; Kumar, R.; Zink, M. Implications of Parameter Uncertainty on Soil Moisture Drought Analysis in Germany. J. Hydrometeorol. 2013, 14, 47–68. [Google Scholar] [CrossRef]
- Marx, A. (Ed.) Klimaanpassung in Forschung und Politik; Springer Fachmedien Wiesbaden: Wiesbaden, Germany, 2017; ISBN 978-3-658-05577-6. [Google Scholar]
- Marx, A.; Samaniego, L.; Kumar, R.; Thober, S.; Mai, J.; Zink, M. Der Dürremonitor—Aktuelle Information zur Bodenfeuchte in Deutschland. In Forum für Hydrologie und Wasserbewirtschaftun:g Wasserressourcen—Wissen im Flussgebieten Vernetzen. Beiträge zum Tag der Hydrologie am 17./18. März 2016 in Koblenz, Ausgerichtet von der Hochschule Koblenz und der Bundesanstalt für Gewässerkunde; Wernecke, G., Ebner von Eschenbach, A.-D., Strunck, Y., Kirschbauer, L., Müller, H., Eds.; Deutsche Vereinigung für Wasserwirtschaft, Abwasser und Abfall (DWA): Hennef, Germany, 2016; Volume 37, pp. 131–142. [Google Scholar]
- Schwarz, J.; Skiadaresis, G.; Kohler, M.; Kunz, J.; Schnabel, F.; Vitali, V.; Bauhus, J. Quantifying Growth Responses of Trees to Drought—A Critique of Commonly Used Resilience Indices and Recommendations for Future Studies. Curr. For. Rep. 2020, 6, 185–200. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2021. [Google Scholar]
- Wickham, H.; Averick, M.; Bryan, J.; Chang, W.; McGowan, L.D.; François, R.; Grolemund, G.; Hayes, A.; Henry, L.; Hester, J.; et al. Welcome to the Tidyverse. J. Open Source Softw. 2019, 4, 1686. [Google Scholar] [CrossRef]
- Pinheiro, J.; Bates, D.; DebRoy, S.; Sarkar, D.; R Core Team. Nlme: Linear and Nonlinear Mixed Effects Models. R Package Version 3.1-152. 2021. Available online: https://CRAN.R-project.org/package=nlme (accessed on 2 July 2021).
- Bates, D.; Mächler, M.; Bolker, B.; Walker, S. Fitting Linear Mixed-Effects Models Using Lme4. J. Stat. Softw. 2015, 67, 1–48. [Google Scholar] [CrossRef]
- Pretzsch, H.; Forrester, D.I. Stand Dynamics of Mixed-Species Stands Compared with Monocultures. In Mixed-Species Forests; Pretzsch, H., Forrester, D.I., Bauhus, J., Eds.; Springer: Berlin/Heidelberg, Germany, 2017; pp. 117–209. ISBN 978-3-662-54551-5. [Google Scholar]
- Bitterlich, W. Die Winkelzählprobe. Forstwiss. Cent. 1952, 71, 215–225. [Google Scholar] [CrossRef]
- Pienaar, L.V.; Rheney, J.W. Modeling Stand Level Growth and Yield Response to Silvicultural Treatments. For. Sci. 1995, 41, 629–638. [Google Scholar] [CrossRef]
- Biging, G.S.; Dobbertin, M. Evaluation of Competition Indices in Individual Tree Growth Models. For. Sci. 1995, 41, 360–377. [Google Scholar] [CrossRef]
- Bowditch, E.; Santopuoli, G.; Binder, F.; del Río, M.; La Porta, N.; Kluvankova, T.; Lesinski, J.; Motta, R.; Pach, M.; Panzacchi, P.; et al. What Is Climate-Smart Forestry? A Definition from a Multinational Collaborative Process Focused on Mountain Regions of Europe. Ecosyst. Serv. 2020, 43, 101113. [Google Scholar] [CrossRef]
- Del Río, M.; Pretszch, H.; Boncina, A.; Avdagić, A.; Bielak, K.; Binder, F.; Coll, L.; Kasanin-Grubin, M.; Klopčič, M.; Hilmers, T.; et al. Assessment of Indicators for Climate Smart Management in Mountain Forests. In Climate-Smart Forestry in Mountain Regions; Tognetti, R., Smith, M., Panzacchi, P., Eds.; Springer Nature Switzerland AG: Cham, Switzerland, in review.
- Forest Europe Sustainable Forest Management, Criteria and Indicators. Available online: http://www.foresteurope.org/sfm_criteria/criteria (accessed on 11 April 2021).












{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Variables’ and Metrics’ Names | Abbreviation | Explanation and Indication |
|---|---|---|
| (i) Tree level variables | ||
| stem diameter | d | indication of tree present size |
| tree height | h | determination of radius for competition analysis |
| height to crown base, to lowest branch | hcb | indication of bole length, used for visualization |
| crown radius | cr | , for visualization |
| crown length | cl | , used for visualization |
| search radius for neighborhood analysis | sr | for analyzing |
| BALpre | m2 ha−1 | local basal area in the circle before selection cut |
| BALpost | m2 ha−1 | local basal area in the circle after selection cut |
| ΔBAL | m2 ha−1 | removed local basal area within circle by selection cut |
| mixing proportion in the reference circle around a tree | mportion | m = 0, i.e., mono-specific stand, 0.1, 0.2… mixing proportions based on standardized stand density indices |
| annual stem diameter increment | id | periodical diameter increment/period length |
| (ii) Stand level variables | ||
| quadratic mean stem diameter | dq | calculated species-overarching |
| standing stem volume | V | merchantable volume > 7 cm at the smaller end |
| stand stem volume growth | IV | periodical mean annual stem volume growth |
| Variable | Unit | Mean | Sd. Dev. | Min | Max |
|---|---|---|---|---|---|
| Tree number | ha−1 | 848.67 | 160.55 | 688.00 | 1080.00 |
| Mean height | m | 19.59 | 1.44 | 17.40 | 21.22 |
| Mean diameter | cm | 25.24 | 3.01 | 21.46 | 28.87 |
| Stand basal area | m2 ha−1 | 42.48 | 10.84 | 29.11 | 59.87 |
| Standing volume | m3 ha−1 | 610.63 | 188.05 | 373.57 | 873.11 |
| Volume proportion N. spruce | ./. | 0.45 | 0.08 | 0.35 | 0.56 |
| Volume proportion s. fir | ./. | 0.46 | 0.05 | 0.41 | 0.53 |
| Volume proportion E. beech | ./. | 0.09 | 0.05 | 0.01 | 0.17 |
| Basal area growth | m2 ha−1 yr−1 | 0.80 | 0.09 | 0.68 | 0.90 |
| Stem volume growth | m3 ha−1 yr−1 | 12.40 | 1.88 | 10.04 | 15.59 |
| Variable | Unit | Mean | Sd. Dev. | Min | Max |
|---|---|---|---|---|---|
| N. spruce, n = 2919 | |||||
| id | cm yr−1 | 0.39 | 0.23 | 0.02 | 1.15 |
| d | cm | 38.94 | 25.10 | 7.00 | 89.50 |
| cd | m | 5.50 | 2.03 | 2.27 | 12.32 |
| height | m | 24.09 | 12.67 | 4.20 | 42.80 |
| crown length | m | 14.77 | 8.24 | 2.00 | 31.40 |
| crown ratio | m m−1 | 0.61 | 0.10 | 0.23 | 0.90 |
| h/d-ratio | m cm−1 | 0.69 | 0.15 | 0.42 | 1.12 |
| BALpre | m2 ha−1 | 40.00 | 33.27 | 0.52 | 196.68 |
| BALpost | m2 ha−1 | 37.14 | 31.97 | 0.52 | 196.68 |
| ΔBAL | m2 ha−1 | 2.87 | 10.85 | 0.00 | 187.45 |
| mportion | ./. | 0.50 | 0.34 | 0.00 | 1.00 |
| s. fir, n = 2360 | |||||
| id | cm yr−1 | 0.17 | 0.15 | 0.01 | 0.73 |
| d | cm | 21.55 | 14.59 | 7.00 | 91.00 |
| cd | m | 4.94 | 1.52 | 1.65 | 9.88 |
| height | m | 16.27 | 8.98 | 4.80 | 41.40 |
| crown length | m | 9.16 | 6.09 | 1.10 | 28.40 |
| crown ratio | m m−1 | 0.54 | 0.15 | 0.16 | 0.87 |
| h/d-ratio | m cm−1 | 0.78 | 0.12 | 0.45 | 1.12 |
| BALpre | m2 ha−1 | 50.46 | 27.10 | 0.48 | 199.23 |
| BALpost | m2 ha−1 | 45.63 | 26.52 | 0.47 | 199.23 |
| ΔBAL | m2 ha−1 | 4.82 | 11.38 | 0.00 | 122.30 |
| mportion | ./. | 0.56 | 0.30 | 0.00 | 1.00 |
| E. beech, n = 1216 | |||||
| id | cm yr−1 | 0.27 | 0.15 | 0.01 | 0.70 |
| d | cm | 33.44 | 14.81 | 7.50 | 70.00 |
| cd | m | 9.53 | 2.35 | 4.00 | 16.18 |
| height | m | 21.85 | 6.53 | 5.10 | 34.80 |
| crown length | m | 16.22 | 4.96 | 2.50 | 24.90 |
| crown ratio | m m−1 | 0.75 | 0.11 | 0.49 | 0.92 |
| h/d-ratio | m cm−1 | 0.71 | 0.20 | 0.24 | 1.24 |
| BALpre | m2 ha−1 | 47.44 | 32.51 | 0.69 | 184.90 |
| BALpost | m2 ha−1 | 44.78 | 31.67 | 0.69 | 184.41 |
| ΔBAL | m2 ha−1 | 2.66 | 9.34 | 0.00 | 119.15 |
| mportion | ./. | 0.75 | 0.27 | 0.00 | 1.00 |
| Model | Species | n | Std | p-Value | Std | p-Value | Std | p-Value | Std | p-Value | Std | p-Value | Std | p-Value | Std | p-Value | |||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 2 | all species | 216 | 0.44 | 0.53 | 0.405 | 0.29 | 0.08 | <0.001 | 0.04 | 0.01 | <0.001 | ||||||||||||
| 3 | all species | 216 | 3.19 | 16.15 | 0.843 | −0.13 | 0.08 | 0.134 | 0.01 | 2.12 | 1.00 | 0.01 | 0.15 | 1.00 | |||||||||
| 4 | N. spruce | 276 | 1351.66 | 281.99 | <0.001 | −259.19 | 74.82 | 0.009 | −178.24 | 37.09 | <0.001 | 34.17 | 9.84 | <0.001 | |||||||||
| 4 | s. fir | 382 | −1068.54 | 278.95 | <0.001 | 207.60 | 79.75 | <0.001 | 140.24 | 36.72 | <0.001 | 27.26 | 10.50 | 0.009 | |||||||||
| 4 | E. beech | 179 | 1202.78 | 455.42 | 0.009 | −325.06 | 136.49 | 0.018 | −158.50 | 59.90 | 0.008 | 42.78 | 17.95 | 0.018 | |||||||||
| 5 | N. spruce | 675 | 144.31 | 43.69 | <0.001 | 0.60 | 0.04 | <0.001 | −0.09 | 0.05 | 0.073 | 0.10 | 0.03 | <0.001 | −19.35 | 5.76 | <0.001 | 0.69 | 0.37 | 0.063 | |||
| 5 | s. fir | 893 | 3828.77 | 1870.84 | <0.041 | 0.65 | 0.04 | <0.001 | −0.25 | 0.05 | <0.001 | 0.13 | 0.02 | <0.001 | −503.69 | 246.03 | <0.040 | 6489.43 | 2819.47 | <0.022 | −853.17 | 370.75 | <0.021 |
| 5 | E. beech | 285 | −1.44 | 0.47 | 0.003 | 0.30 | 0.06 | <0.001 | −0.14 | 0.09 | <0.001 | 0.16 | 0.04 | 0.100 | 1.04 | 0.51 | 0.042 | ||||||
| 6a | N. spruce | 246 | −2.54 | 0.43 | <0.001 | 0.55 | 0.07 | <0.001 | −0.22 | 0.11 | 0.042 | 0.76 | 0.27 | 0.005 | |||||||||
| 6a | s. fir | 421 | 3.90 | 1.97 | 0.049 | 0.80 | 0.07 | <0.001 | −0.28 | 0.08 | 0.001 | 0.17 | 0.04 | <0.001 | 11.55 | 3.02 | <0.001 | ||||||
| 6a | E. beech | 89 | −4.13 | 0.62 | <0.001 | 0.23 | 0.17 | 0.184 | 0.13 | 0.06 | <0.001 | 2.95 | 0.52 | <0.001 | |||||||||
| 6b | N. spruce | 435 | −2.72 | 0.33 | <0.001 | 0.67 | 0.05 | <0.001 | −0.08 | 0.07 | 0.201 | 0.17 | 0.04 | <0.001 | |||||||||
| 6b | s. fir | 494 | −2.37 | 0.30 | <0.001 | 0.55 | 0.05 | <0.001 | −0.22 | 0.07 | 0.002 | 0.12 | 0.03 | <0.001 | 1.20 | 0.37 | 0.002 | ||||||
| 6b | E. beech | 194 | −1.25 | 0.48 | 0.009 | 0.27 | 0.07 | <0.001 | −0.12 | 0.09 | 0.188 | 0.22 | 0.05 | <0.001 | 0.39 | 0.16 | 0.014 |
| Fixed Effect Variable | t Value | p Value |
|---|---|---|
| Intercept | 10.601 | <0.001 |
| period 1986 | −1.905 | 0.058 |
| period 1993 | −0.244 | 0.807 |
| period 1999 | −0.623 | 0.534 |
| period 2005 | −0.699 | 0.485 |
| period 2011 | −0.707 | 0.481 |
| Smooth terms | F value | p value |
| DBH-class | 13.649 | <0.001 |
| DBH-class: period 1980 | 4.335 | <0.001 |
| DBH-class: period 1986 | 0.041 | 0.851 |
| DBH-class: period 1993 | 0.526 | 0.665 |
| DBH-class: period 1999 | 0.019 | 0.898 |
| DBH-class: period 2005 | 0.036 | 0.860 |
| DBH-class: period 2011 | 0.219 | 0.788 |
| Fixed Effect Variable | t Value | p Value |
|---|---|---|
| Intercept | 4.631 | <0.001 |
| Smoother terms | F value | p value |
| DBH-class | 0.772 | 0.492 |
| period | 2.966 | 0.034 |
| DBH-class, period | 0.786 | 0.001 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Uhl, E.; Hilmers, T.; Pretzsch, H. From Acid Rain to Low Precipitation: The Role Reversal of Norway Spruce, Silver Fir, and European Beech in a Selection Mountain Forest and Its Implications for Forest Management. Forests 2021, 12, 894. https://0-doi-org.brum.beds.ac.uk/10.3390/f12070894
Uhl E, Hilmers T, Pretzsch H. From Acid Rain to Low Precipitation: The Role Reversal of Norway Spruce, Silver Fir, and European Beech in a Selection Mountain Forest and Its Implications for Forest Management. Forests. 2021; 12(7):894. https://0-doi-org.brum.beds.ac.uk/10.3390/f12070894
Chicago/Turabian StyleUhl, Enno, Torben Hilmers, and Hans Pretzsch. 2021. "From Acid Rain to Low Precipitation: The Role Reversal of Norway Spruce, Silver Fir, and European Beech in a Selection Mountain Forest and Its Implications for Forest Management" Forests 12, no. 7: 894. https://0-doi-org.brum.beds.ac.uk/10.3390/f12070894