Evaluation of Long-Term Shortleaf Pine Progeny Tests in the Ouachita and Ozark National Forests, USA

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
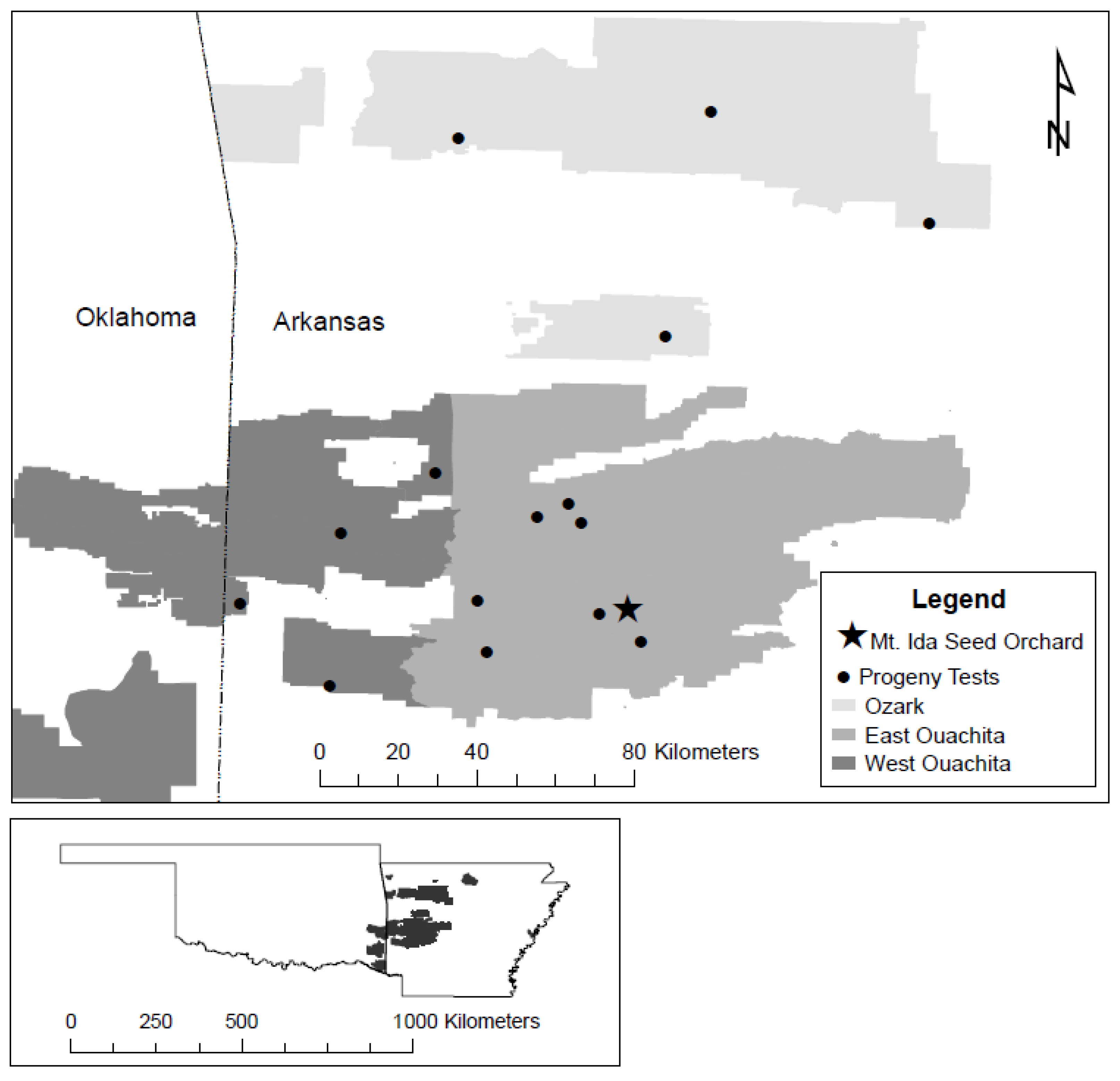
2.1. Study Area
2.2. Seed Sources
2.3. Original Design of Crosses and Test Plantations
2.4. Current Sampling and Data Collection
2.5. Data Analysis
3. Results and Discussion
3.1. Tree Size Differences
3.2. Survival
3.3. Trait Heritability
4. Management Implications
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Little, E.L., Jr. Atlas of United States Trees. Volume 1. Conifers and Important Hardwoods; Miscellaneous Publication 1146; U.S. Department of Agriculture: Washington, DC, USA, 1971; p. 9.
- Sutter, R. Ecological and Social History of Shortleaf Pine; Shortleaf Pine Initiative: Lexington, KY, USA, 2019; Available online: http://shortleafpine.net/why-shortleaf/history (accessed on 16 December 2019).
- Mohr, C.T.; Roth, F. The Timber Pines of the Southern United States; Bulletin 13 (Revised); USDA Division of Forestry: Washington, DC, USA, 1897; p. 176.
- Mattoon, W.R. The Life History of Shortleaf Pine; Bulletin 244; USDA Forest Service: Washington, DC, USA, 1915; p. 46.
- Pickens, B. Silvics of Shortleaf Pine; Shortleaf Pine Initiative: Lexington, KY, USA, 2019; Available online: http://shortleafpine.net/why-shortleaf/silvics/silvics-of-shortleaf-pine/ (accessed on 16 December 2019).
- Walker, L.C.; Wiant, H.V. Silviculture of Shortleaf Pine; School of Forestry Bulletin 9; Stephen F. Austin State College: Nacogdoches, TX, USA, 1966; p. 60. [Google Scholar]
- Lawson, E.R. Pinus echinata Mill. Shortleaf pine. In Silvics of North America. Volume 1. Conifers; Agriculture Handbook 654; Burns, R.M., Honkala, B.H., Eds.; USDA Forest Service: Washington, DC, USA, 1990; pp. 316–326. [Google Scholar]
- Studyvin, C.; Gwaze, D. Genetic Improvement of Shortleaf Pine on the Mark Twain, Ouachita, and Ozark National Forests; General Technical Report NRS-P-15; USDA Forest Service, Northern Research Station: Newtown Square, PA, USA, 2007; pp. 84–88.
- Oswalt, C.M. Spatial and temporal trends of the shortleaf pine resource in the eastern United States. In Proceedings of the Shortleaf Pine Conference: East Meets West; Special Report 11; Kush, J., Barlow, R.J., Gilbert, J.C., Eds.; Alabama Agricultural Experiment Station: Auburn, AL, USA, 2012; pp. 33–37. [Google Scholar]
- Stewart, J.F.; Tauer, C.G.; Guldin, J.M.; Nelson, C.D. Hybridization in naturally regenerated shortleaf pine near stands of artificially regenerated stands of loblolly pine. South. J. Appl. For. 2013, 37, 102–107. [Google Scholar] [CrossRef]
- Stewart, J.F.; Will, R.E.; Robertson, K.M.; Nelson, C.D. Frequent fire protects shortleaf pine (Pinus echinata) from introgression by loblolly pine (P. taeda). Conserv. Genet. 2015, 16, 491–495. [Google Scholar] [CrossRef]
- Tauer, C.G.; Stewart, J.F.; Will, R.E.; Lilly, C.J.; Guldin, J.M.; Nelson, C.D. Hybridization leads to loss of genetic integrity in shortleaf pine: Unexpected consequences of pine management and fire suppression. J. For. 2012, 110, 216–224. [Google Scholar] [CrossRef]
- Rosson, J.F. Forests of Arkansas, 2015; Resource Update FS-81; USDA Forest Service: Washington, DC, USA, 2016; p. 4.
- Bragg, D.C. Silviculture’s impact on the historical shortleaf component of pine forests in the Upper West Gulf Coastal Plain. In Proceedings of the 18th Biennial Southern Silvicultural Research Conference, Knoxville, TN, USA, 2–5 March 2015; Schweitzer, C.J., Clatterbuck, W.K., Oswalt, C.M., Eds.; e-Gen. Tech. Rep. SRS-212. USDA Forest Service, Southern Research Station: Asheville, NC, USA, 2016; pp. 432–443. [Google Scholar]
- Anderson, M.L.; Hayes, P.D.; Keyser, C.M.; Lituma, R.D.; Sutter, R.; Zollner, D. Shortleaf Pine Restoration Plan: Restoring an American Forest Legacy; Shortleaf Pine Initiative: Lexington, KY, USA, 2016; Available online: http://shortleafpine.net/tools-and-resources/restoration-plan/shortleaf-pine-restoration-plan (accessed on 21 February 2020).
- Guldin, J.M. Restoration and Management of Shortleaf Pine in Pure and Mixed Stands—Science, Empirical Observation, and the Wishful Application of Generalities; General Technical Report NRS-P-15; USDA Forest Service: Newtown Square, PA, USA, 2007; pp. 47–57.
- Studyvin, C.; Gwaze, D. Differences among Shortleaf Pine Seed Sources on the Ozark and Ouachita National Forest at Age Ten; General Technical Report SRS-156; USDA Forest Service, Southern Research Station: Asheville, NC, USA, 2012; pp. 329–333.
- Crane, B.; Hipkins, V.; Josserand, S.; Echt, C. Genetic integrity of longleaf and shortleaf pine seed orchards and seed banks. Tree Plant. Notes 2019, 62, 95–103. [Google Scholar]
- Guldin, J.M. Silvicultural options in forests of the southern United States under changing climate conditions. New For. 2019, 50, 71–87. [Google Scholar] [CrossRef]
- Kitchens, R.N. Trends in Shortleaf Pine Tree Improvement; Arkansas Cooperative Extension Service, University of Arkansas at Monticello: Monticello, AR, USA, 1986; pp. 89–100. [Google Scholar]
- Schmidtling, R.C. Southern Pine Seed Sources; General Technical Report SRS-44; USDA Forest Service: Asheville, NC, USA, 2001; 25p.
- Stewart, J.F.; Will, R.E.; Crane, B.S.; Nelson, C.D. The genetics of shortleaf pine (Pinus echinata Mill.) with implications for restoration and management. Tree Genet. Genomes 2016, 12, 15. [Google Scholar] [CrossRef]
- Wells, O.O.; Wakeley, P.C. Variation in shortleaf pine from several geographic sources. For. Sci. 1970, 16, 415–423. [Google Scholar]
- Bragg, D.C.; Hossain, S.M. Revisiting shortleaf outplantings of the Southwide Pine Seed Source Study on the Crossett Experimental Forest. In Proceedings of the 20th Biennial Southern Silvicultural Research Conference, Shreveport, LA, USA, 12–14 March 2019; Bragg, D.C., Koerth, N.E., Holley, A.G., Eds.; Gen. Tech. Rep. SRS-253. USDA Forest Service, Southern Research Station: Asheville, NC, USA, 2020; pp. 318–324. [Google Scholar]
- Zarnoch, S.J.; Ruehle, J.L.; Belanger, R.P.; Marx, D.H.; Bryan, W.C. Growth and Crown Vigor of 25-Year-Old Shortleaf Pine Progenies on a Littleleaf Disease Site; Research Paper SE-289; USDA Forest Service, Southeastern Forest Experiment Station: Asheville, NC, USA, 1994; 12p.
- La Farge, T. Are there significant differences among the three shortleaf pine seed sources on the National Forests in Arkansas? In Proceedings of the 1st Northern Forest Genetics Association Conference, Burlington, VT, USA, 23–25 July 1991; DeHayes, D.H., Hawley, G.J., Hanover, J.W., Eds.; The University of Vermont: Burlington, VT, USA, 1991; pp. 40–47. [Google Scholar]
- Crane, B. Shortleaf Pine Genetic Resources to Support Restoration in the Southern Region; Shortleaf Pine Workshop: Ecology and Management for Multiple Objectives in the Interior Highlands, Online Webinar. 2014. Available online: http://shortleafpine.net/media/videos-and-webinars/webinar (accessed on 15 April 2021).
- Crane, B. From tree improvement to species improvement: Restoration and conservation efforts on the Forest Service’s Southern National Forests. In Proceedings of the 28th Southern Forest Tree Improvement Conference, Raleigh, NC, USA, 21–23 June 2005; North Carolina State University: Raleigh, NC, USA, 2005; p. 168. [Google Scholar]
- Woods, A.J.; Foti, T.L.; Chapman, S.S.; Omernik, J.M.; Wise, J.A.; Murray, E.O.; Prior, W.L.; Pagan, J.B.; Comstock, J.A., Jr.; Radford, M. Ecoregions of Arkansas (Color Poster with Map, Descriptive Text, Summary Tables, and Photographs); U.S. Geological Survey (map scale 1:1,000,000): Reston, VA, USA, 2004.
- PRISM Climate Group. 30-yr Normal Mean Temperature [Map]: Annual (Period: 1981–2010); PRISM Climate Group: Oregon State University, Corvallis, OR, USA, 2013; Available online: http://www.prism.oregonstate.edu/normals/ (accessed on 9 May 2019).
- Pugh, A.L.; Westerman, D.A. Mean Annual, Seasonal, and Monthly Precipitation and Runoff in Arkansas, 1951–2011; Science Investigation Report 2014-5006; U.S. Department of the Interior Geological Survey: Reston, VA, USA, 2014; p. 40.
- Rowland, B.; Ouachita National Forest, Mount Ida, AR, USA. Personal communication, 2019.
- La Farge, T.; Gates, J.E. The use of best linear prediction to obtain breeding values for height and survival in 37 full-sib progeny tests of shortleaf pine (Pinus echinata Mill.) on the Ouachita and Ozark-St. Francis National Forests. In Proceedings of the 21st Southern Forest Tree Improvement Conference, Knoxville, TN, USA, 17–20 June 1991; Tennessee Division of Forestry: Knoxville, TN, USA, 1991; pp. 144–151. [Google Scholar]
- Web Soil Survey. Natural Resources; Web Soil Survey: Fayetteville, AR, USA, 2020. Available online: https://websoilsurvey.sc.egov.usda.gov/App/WebSoilSurvey.aspx (accessed on 21 January 2020).
- SAS Institute Inc. SAS/STAT User’s Guide, Version 9.4 ed; SAS Institute Inc.: Cary, NC, USA, 2018; p. 846. [Google Scholar]
- Isik, F.; Holland, J.; Maltecca, C. Genetic Data Analysis for Plant and Animal Breeding; Springer: Cham, Switzerland, 2017; p. 400. [Google Scholar] [CrossRef]
- Lynch, M.; Walsh, B. Genetics and Analysis of Quantitative Traits, 1st ed.; Sinauer Associates, Inc.: Sunderland, MA, USA, 1998. [Google Scholar]
- Gwaze, D.P.; Melick, R.; Studyvin, C.; Coggeshall, M. Genetic control of growth traits in shortleaf pine in Missouri. South. J. Appl. For. 2005, 29, 2000–2004. [Google Scholar] [CrossRef] [Green Version]
- Schmidtling, R.C. Seed Transfer and Genecology in Shortleaf Pine; General Technical Report SRS-1; USDA Forest Service: Asheville, NC, USA, 1995; pp. 373–378.
- Schmidtling, R.C. Determining seed transfer guidelines for southern pines. In National Proceedings: Forest and Conservation Nursery Associations-2002; Proceedings RMRS-P-28; Riley, L.E., Dumroese, R.K., Landis, T.D., Tech Cords, Eds.; USDA Forest Service, Rocky Mountain Research Station: Ogden, UT, USA, 2003; pp. 8–11. [Google Scholar]
- McCullough, R.B. Phenotypic Variation and Relationships of Several Variables in Shortleaf Pine in Oklahoma. Doctoral Thesis, Oklahoma State University, Stillwater, OK, USA, 1972; p. 53.
- Jayawickrama, K.J.S.; McKeand, S.E.; Jett, J.B. Phenological variation in height and diameter growth in provenances and families of loblolly pine. New For. 1998, 16, 11–25. [Google Scholar] [CrossRef]
- Fox, T.R.; Jokela, E.J.; Allen, H.L. The development of pine plantation silviculture in the southern United States. J. For. 2007, 105, 337–347. [Google Scholar]
- Bragg, D.C.; Shelton, M.G.; Zeide, B. Impacts and management implications of ice storms on forests in the southern United States. For. Ecol. Manag. 2003, 186, 99–123. [Google Scholar] [CrossRef]
- Pike, C.; Potter, K.M.; Berrang, P.; Crane, B.; Baggs, J.; Leites, L.; Luther, T. New seed-collection zones for the Eastern United States: The Eastern Seed Zone Forum. J. For. 2020, 118, 444–451. [Google Scholar] [CrossRef] [Green Version]
- Bridgewater, F.S., Jr. Genotypic Variation among Shortleaf Pine Seedlings within Oklahoma. Doctoral Thesis, Oklahoma State University, Stillwater, OK, USA, 1969; p. 52. [Google Scholar]
- Wells, O.O.; McConnell, J.L. Breeding populations in the R-8 tree improvement program. In Progeny Testing: Proceedings of the a Servicewide Genetics Workshop, Charleston, SC, USA, 5–9 December 1983; Miller, R., Ed.; USDA Forest Service: Washington, DC, USA, 1984; pp. 61–67. Available online: https://www.fs.usda.gov/treesearch/pubs/55399 (accessed on 16 December 2019).



{kind=link}
{kind=link}
| Test # | Ranger District | Source Region | SI50 (m) a | -------Families------- | Date Planted | ------------------------As Measured in 2018–2019------------------------ | ||||
|---|---|---|---|---|---|---|---|---|---|---|
| Unique b | Shared | Age (Years) c | % Storm Damage d | Live # of Trees | Percent Alive | |||||
| 21 | Oden | EOU | 21.3 | 15 | 8 | 1984 | 34 | 88.8 | 660 | 49.2 |
| 29 | Oden | EOU | 19.8 | 24 | 3 | 1985 | 33 | 65.9 | 772 | 58.9 |
| 48 | Oden | EOU | 19.8 | 26 | 6 | 1987 | 31 | 88.3 | 706 | 47.7 |
| 1 | Womble | EOU | 18.2 | 7 | 12 | 1978 | 40 | 38.5 | 437 | 48 |
| 5 | Womble | EOU | 21.3 | 28 | 6 | 1981 | 38 | 75.7 | 778 | 58.3 |
| 19 | Womble | EOU | 21.3 | 1 | 27 | 1984 | 34 | 76.1 | 625 | 36.3 |
| 31 | Womble | EOU | 21.3 | 11 | 18 | 1985 | 33 | 65.3 | 578 | 51.7 |
| 25 | Cold Springs | WOU | 19.8 | 25 | 9 | 1984 | 35 | 68.9 | 1086 | 63.8 |
| 6 | Mena | WOU | 19.8 | 3 | 28 | 1981 | 38 | 93.3 | 691 | 43.1 |
| 10 | Mena | WOU | 21.3 | 21 | 16 | 1982 | 37 | 34.2 | 724 | 53.2 |
| 38 | Mena | WOU | 19.8 | 6 | 15 | 1986 | 33 | 36.9 | 563 | 46.5 |
| 14 | Big Piney | OZ | 15.2 | 15 | 15 | 1984 | 35 | 8 | 755 | 47.1 |
| 28 | Boston Mountains | OZ | 15.2 | 27 | 9 | 1987 | 32 | 8.8 | 871 | 54.4 |
| 33 | Mount Magazine | OZ | 18.2 | 10 | 12 | 1988 | 31 | 4.1 | 947 | 56 |
| 32 | Pleasant Hill | OZ | 18.2 | 30 | 7 | 1988 | 31 | 2.2 | 755 | 69.9 |
| Averages | 50.4 | 52.3 | ||||||||
| Totals | 249 | 96 | 11,058 | |||||||
| Family Type a | Number of Families | ---Live Trees--- | ----------d.b.h. (cm)----------- | ----------Total Height (m)--------- | |||
|---|---|---|---|---|---|---|---|
| No. | Percent b | Mean (Min.–Max.) | CV | Mean (Min.–Max.) | CV | ||
| Unique | 249 | 5593 | 55.9 | 23.1 (7.80–43.7) | 16.7 | 17.4 (9.14–24.9) | 9.88 |
| Shared-1 | 88 | 3630 | 49.5 | 24.4 (8.63–44.6) | 18.3 | 18.7 (8.83–30.5) | 12.5 |
| Shared-2 | 8 | 1835 | 51.6 | 23.5 (9.39–42.7) | 21.3 | 18.3 (10.4–27.4) | 15.7 |
| All | 345 | 11058 | 52.3 | 23.6 (7.80–44.6) | 18.8 | 18.1 (8.83–30.5) | 12.8 |
| Family a | Parent Seed Source(s) | No. of Tests | ---Live Trees--- | ---------d.b.h. (cm)-------- | ------Total Height (m)----- | |||
|---|---|---|---|---|---|---|---|---|
| No. | Percent b | Mean (Min.–Max.) | CV | Mean (Min.–Max.) | CV | |||
| 103_201 | EOU | 13 | 292 | 53.1 | 24.3 (10.4–40.6) | 18.3 | 18.7 (10.4–25.9) | 16.0 |
| 115_312 | EOU & WOU | 14 | 302 | 52.7 | 23.1 (11.6–34.1) | 18.2 | 17.8 (10.9–25.6) | 15.8 |
| 120_333 | EOU | 9 | 211 | 57.4 | 24.1 (10.2–35.8) | 20.8 | 17.8 (11.5–24.4) | 16.8 |
| 135_233 | EOU | 3 | 47 | 36.6 | 28.1 (16.5–39.1) | 20.8 | 19.9 (14.6–25.1) | 15.3 |
| 322_901 | EOU & WOU | 14 | 277 | 48.9 | 23.2 (11.4–38.3) | 20.7 | 17.8 (11.6–24.1) | 15.5 |
| 620_901 | EOU & OZ | 14 | 267 | 46.0 | 23.1 (9.39–42.7) | 23.1 | 17.7 (11.3–23.5) | 15.0 |
| 319_913 | EOU | 14 | 350 | 59.8 | 23.3 (10.2–35.8) | 20.7 | 17.8 (11.3–24.3) | 17.1 |
| 888_999 | EOU | 5 | 89 | 49.4 | 21.8 (10.2–34.3) | 22.9 | 19.1 (15.5–27.4) | 14.2 |
| Variable | Covariance Parameter | Estimate | Z Value | p > Z |
|---|---|---|---|---|
| d.b.h. | Test | 0.63 | 2.53 | 0.0057 |
| Replication(test) | 0.06 | 2.83 | 0.0024 | |
| Family | 0.23 | 4.54 | 0.0001 | |
| Test*family | 0.04 | 1.43 | 0.0771 | |
| Rep*family(test) | 0.06 | 1.53 | 0.0629 | |
| Residual | 2.88 | 43.18 | 0.0001 | |
| Height | Test | 37.2 | 2.41 | 0.0079 |
| Replication(test) | 13.5 | 4.58 | 0.0001 | |
| Family | 1.89 | 2.33 | 0.0096 | |
| Test*family | 1.15 | 1.40 | 0.0809 | |
| Rep*family(Test) | 10.3 | 7.74 | 0.0001 | |
| Residual | 27.1 | 23.31 | 0.0001 | |
| Survival | Test | 0.09 | 2.11 | 0.0213 |
| Replication(test) | 0.03 | 2.51 | 0.0731 | |
| Family | 0.04 | 1.95 | 0.1843 | |
| Test*family | 0.05 | 2.64 | 0.0673 | |
| Rep*family(test) | 0.06 | 2.66 | 0.0746 | |
| Residual | 0.97 | 71.21 | 0.0001 |
| Trait | Test | ------------Family a-------------- ---------Replication a----------- ----Replication × Family a--- | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| no. | Estimate | Z Val. | p > Z | Estimate | Z Val. | p > Z | Estimate | Z Val. | p > Z | |
| d.b.h. | ||||||||||
| 1 | 0.23 | 1.53 | 0.062 | n.a. | n.a. | n.a. | 0.40 | 2.30 | 0.011 | |
| 5 | 0.33 | 2.76 | 0.002 | 0.12 | 1.07 | 0.142 | 0.03 | 0.42 | 0.337 | |
| 6 | 0.36 | 2.86 | 0.002 | 0.09 | 1.13 | 0.129 | 0.01 | 0.03 | 0.486 | |
| 10 | 0.16 | 1.97 | 0.024 | 0.02 | 0.67 | 0.252 | 0.09 | 1.01 | 0.156 | |
| 14 | 0.06 | 1.04 | 0.149 | n.a. | n.a. | n.a. | 0.16 | 1.34 | 0.090 | |
| 19 | 0.25 | 1.89 | 0.029 | 0.04 | 0.75 | 0.226 | 0.25 | 1.72 | 0.042 | |
| 21 | 0.18 | 2.16 | 0.015 | 0.03 | 0.82 | 0.206 | n.a. | n.a. | n.a. | |
| 25 | 0.11 | 1.90 | 0.028 | 0.04 | 0.94 | 0.173 | 0.20 | 2.69 | 0.003 | |
| 28 | 0.44 | 3.03 | 0.001 | n.a. | n.a. | n.a. | n.a. | n.a. | n.a. | |
| 29 | 0.01 | 0.50 | 0.308 | 0 | 0.45 | 0.326 | n.a. | n.a. | n.a. | |
| 31 | 0.14 | 1.81 | 0.035 | 0.06 | 1.02 | 0.154 | 0.13 | 1.40 | 0.080 | |
| 32 | 0.16 | 1.94 | 0.026 | 0.02 | 0.63 | 0.265 | 0.11 | 1.19 | 0.116 | |
| 33 | 0.32 | 2.89 | 0.001 | 0.03 | 0.90 | 0.185 | 0.06 | 0.67 | 0.250 | |
| 38 | 0.20 | 1.68 | 0.045 | 0.09 | 1.05 | 0.147 | 0.06 | 0.50 | 0.310 | |
| 48 | 0.11 | 1.79 | 0.036 | n.a. | n.a. | n.a. | 0 | 0.11 | 0.454 | |
| Height | ||||||||||
| 1 | 2.65 | 1.32 | 0.056 | 41.80 | 1.20 | 0.115 | 12.10 | 2.61 | 0.004 | |
| 5 | 1.18 | 0.61 | 0.271 | 32.10 | 1.20 | 0.114 | 8.76 | 2.61 | 0.004 | |
| 6 | 0.29 | 0.35 | 0.365 | 9.48 | 1.35 | 0.088 | 5.75 | 2.92 | 0.001 | |
| 10 | 0 | n.a. | n.a. | 12.00 | 1.19 | 0.117 | 5.91 | 3.27 | 0.001 | |
| 14 | 4.86 | 2.29 | 0.011 | 1.92 | 1.10 | 0.135 | 3.33 | 1.21 | 0.113 | |
| 19 | 0 | n.a. | n.a. | 10.50 | 1.34 | 0.089 | 7.05 | 2.69 | 0.003 | |
| 21 | 2.33 | 1.71 | 0.043 | 1.24 | 0.96 | 0.169 | 3.02 | 1.53 | 0.063 | |
| 25 | 0 | n.a. | n.a. | 20.10 | 1.37 | 0.085 | 10.00 | 3.54 | 0 | |
| 28 | 4.83 | 2.37 | 0.008 | 0.75 | 0.89 | 0.186 | 5.58 | 2.65 | 0.004 | |
| 29 | 0 | n.a. | n.a. | 29.50 | 1.39 | 0.083 | 9.24 | 4.27 | 0 | |
| 31 | 3.51 | 1.76 | 0.038 | 28.00 | 1.38 | 0.083 | 5.60 | 1.87 | 0.030 | |
| 32 | 0 | n.a. | n.a. | n.a. | n.a. | n.a. | 5.99 | 1.69 | 0.045 | |
| 33 | 6.01 | 1.47 | 0.050 | 11.40 | 1.25 | 0.104 | 29.30 | 4.59 | 0 | |
| 38 | 3.95 | 1.18 | 0.118 | 22.90 | 1.34 | 0.090 | 9.51 | 1.67 | 0.047 | |
| 48 | 1.69 | 1.71 | 0.043 | 1.48 | 1.15 | 0.125 | 0.96 | 0.57 | 0.285 | |
| Survival | ||||||||||
| 1 | 0.01 | 0.53 | 0.296 | 0.01 | 0.64 | 0.259 | n.a. | n.a. | n.a. | |
| 5 | 0.01 | 1.65 | 0.049 | 0 | 0.09 | 0.465 | 0.01 | 2.51 | 0.006 | |
| 6 | 0.01 | 0.77 | 0.220 | 0 | 0.28 | 0.391 | n.a. | n.a. | n.a. | |
| 10 | 0.01 | 1.31 | 0.095 | 0 | 0.87 | 0.193 | 0.01 | 2.26 | 0.011 | |
| 14 | 0.01 | 1.61 | 0.053 | 0 | 0.70 | 0.243 | 0.01 | 2.10 | 0.018 | |
| 19 | 0.01 | 2.25 | 0.012 | 0 | 0.13 | 0.446 | 0.01 | 1.57 | 0.058 | |
| 21 | 0.01 | 0.78 | 0.217 | 0 | 0.85 | 0.198 | 0.01 | 1.92 | 0.027 | |
| 25 | 0.01 | 1.49 | 0.068 | n.a. | n.a. | n.a. | 0.01 | 3.26 | 0 | |
| 28 | 0.02 | 2.50 | 0.006 | 0 | 0.43 | 0.333 | 0.01 | 2.75 | 0.003 | |
| 29 | 0.01 | 1.80 | 0.035 | 0 | 0.56 | 0.286 | 0 | 0.89 | 0.187 | |
| 31 | 0.01 | 2.29 | 0.011 | 0 | 0.46 | 0.324 | 0 | 1.27 | 0.102 | |
| 32 | 0.01 | 1.70 | 0.044 | 0 | 0.24 | 0.404 | 0 | 1.23 | 0.108 | |
| 33 | 0.01 | 1.40 | 0.081 | 0 | 0.82 | 0.206 | 0.01 | 2.92 | 0.001 | |
| 38 | 0.02 | 2.28 | 0.011 | 0 | 0.52 | 0.300 | 0 | 1.53 | 0.063 | |
| 48 | 0.02 | 2.27 | 0.011 | n.a. | n.a. | n.a. | 0 | 0.92 | 0.179 | |
| Variable | Source of Variation | df | F | p |
|---|---|---|---|---|
| d.b.h. | Seed source | 2 | 4.20 | 0.050 |
| Height | Seed source | 2 | 0.43 | 0.644 |
| Survival | Seed source | 2 | 2.33 | 0.023 |
| Seed Source | d.b.h. | CV | Total Height | CV | Survival | CV |
|---|---|---|---|---|---|---|
| East Ouachita (EOU) | 24.6 a | 6.34 | 18.4 a | 9.56 | 50.0 a | 4.78 |
| West Ouachita (WOU) | 23.7 a | 6.49 | 18.2 a | 8.41 | 51.4 a | 3.97 |
| Ozark (OZ) | 22.4 b | 6.38 | 17.7 a | 5.98 | 56.7 b | 5.11 |
| Trait | Individual Tree Heritability (h2i) | Standard Error of h2i Estimate | Full-Sibling Family Mean Heritability (h2FS) | Standard Error of h2FS Estimate |
|---|---|---|---|---|
| d.b.h. | 0.146 | 0.030 | 0.720 | 0.054 |
| Height | 0.094 | 0.039 | 0.406 | 0.118 |
| Survival | 0.067 | 0.034 | 0.520 | 0.267 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hossain, S.M.; Bragg, D.C.; McDaniel, V.L.; Pike, C.C.; Crane, B.S.; Nelson, C.D. Evaluation of Long-Term Shortleaf Pine Progeny Tests in the Ouachita and Ozark National Forests, USA. Forests 2021, 12, 953. https://0-doi-org.brum.beds.ac.uk/10.3390/f12070953
Hossain SM, Bragg DC, McDaniel VL, Pike CC, Crane BS, Nelson CD. Evaluation of Long-Term Shortleaf Pine Progeny Tests in the Ouachita and Ozark National Forests, USA. Forests. 2021; 12(7):953. https://0-doi-org.brum.beds.ac.uk/10.3390/f12070953
Chicago/Turabian StyleHossain, Shaik M., Don C. Bragg, Virginia L. McDaniel, Carolyn C. Pike, Barbara S. Crane, and C. Dana Nelson. 2021. "Evaluation of Long-Term Shortleaf Pine Progeny Tests in the Ouachita and Ozark National Forests, USA" Forests 12, no. 7: 953. https://0-doi-org.brum.beds.ac.uk/10.3390/f12070953