
Analyses of Microstructure and Dynamic Deposition of Cell Wall Components in Xylem Provide Insights into Differences between Two Black Poplar Cultivars


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Materials
2.2. Sample Preparation and Light Microscopy
2.3. Measurement of Cell Wall Thickness and Anatomical Characteristics
2.4. SRS Microscopy
2.5. Quantitative Analysis of SRS Image
2.6. Immunolocalization of Pectin
3. Results and Discussion
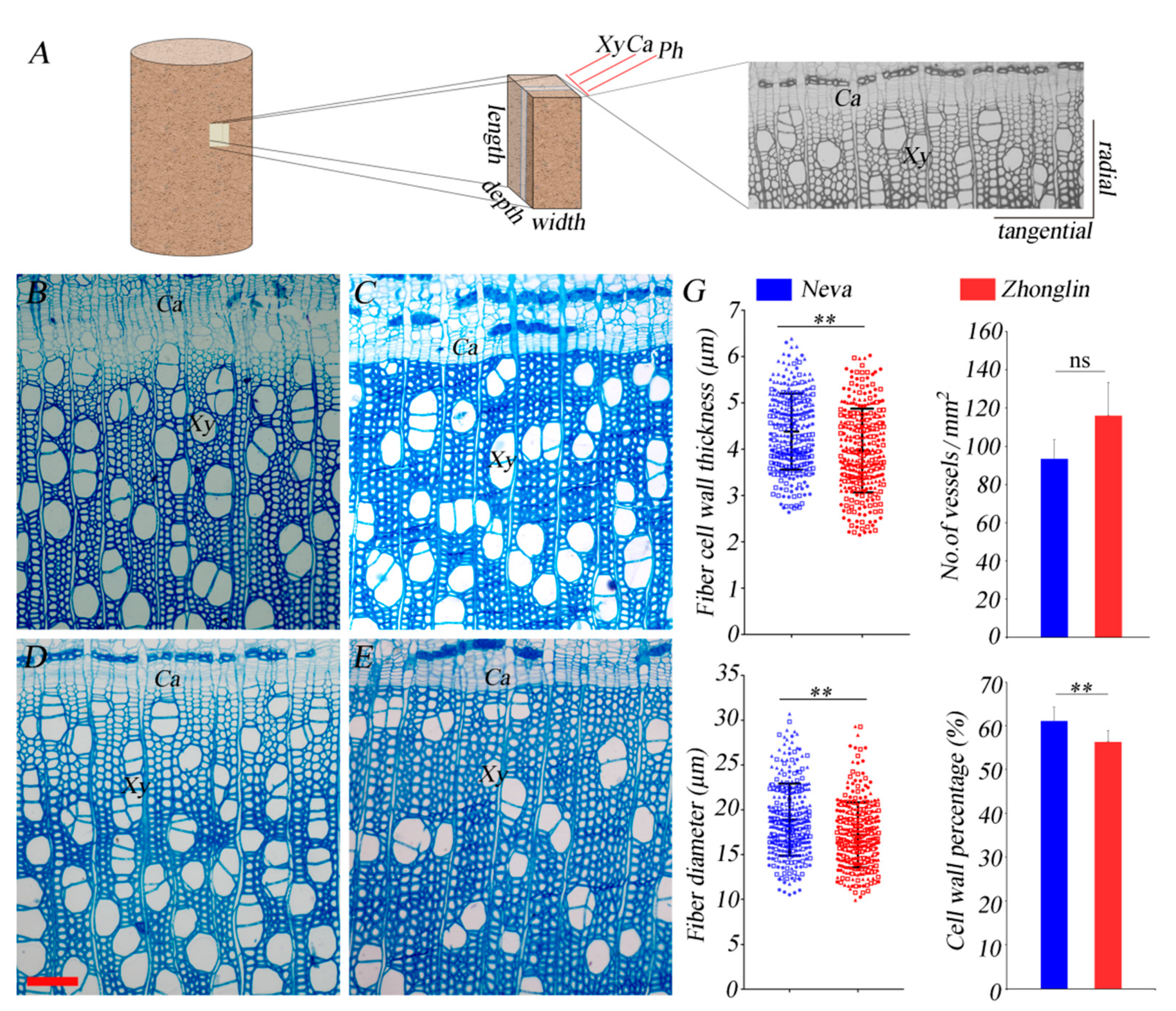
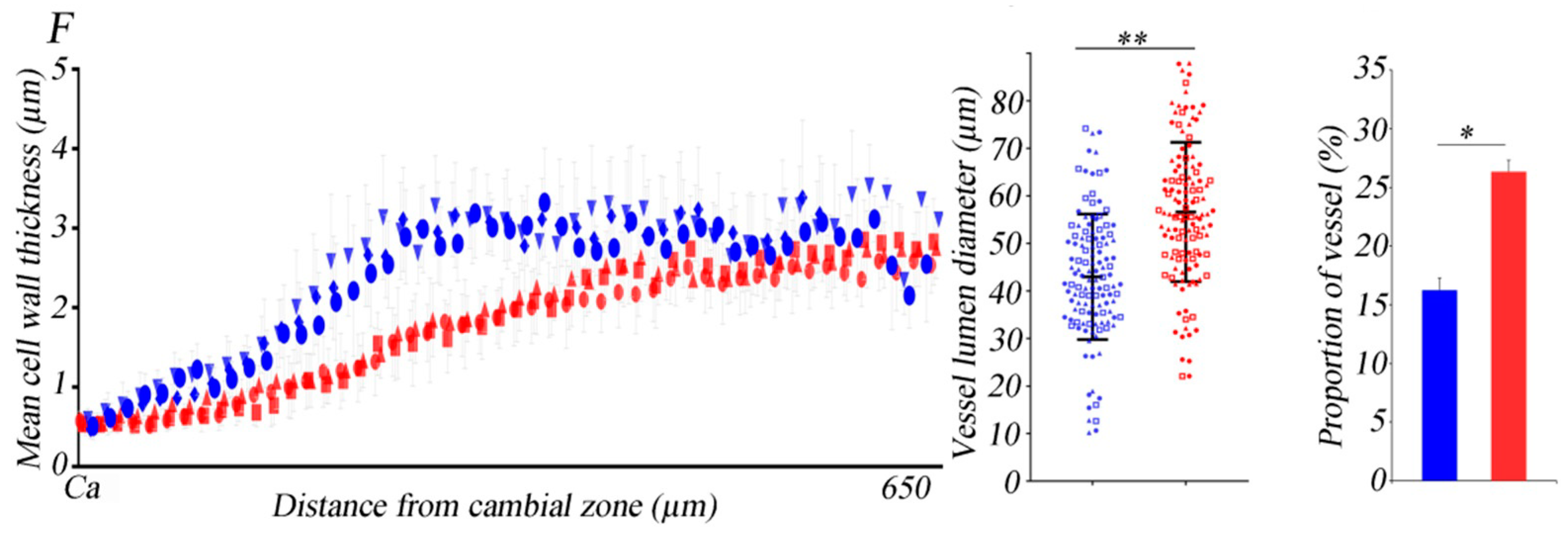
3.1. Differences in Anatomical Characteristics between the Two Black Poplar Cultivars
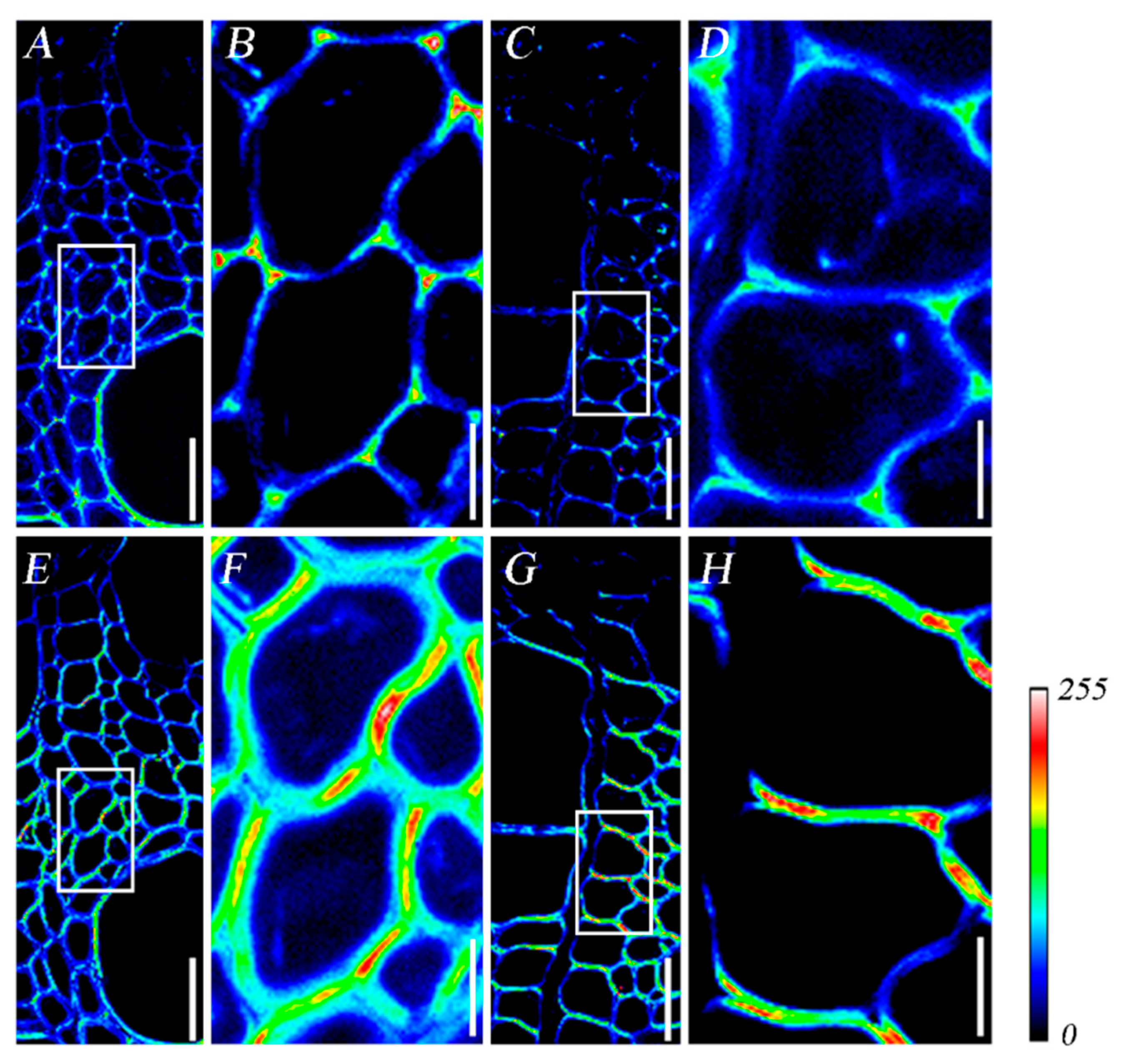
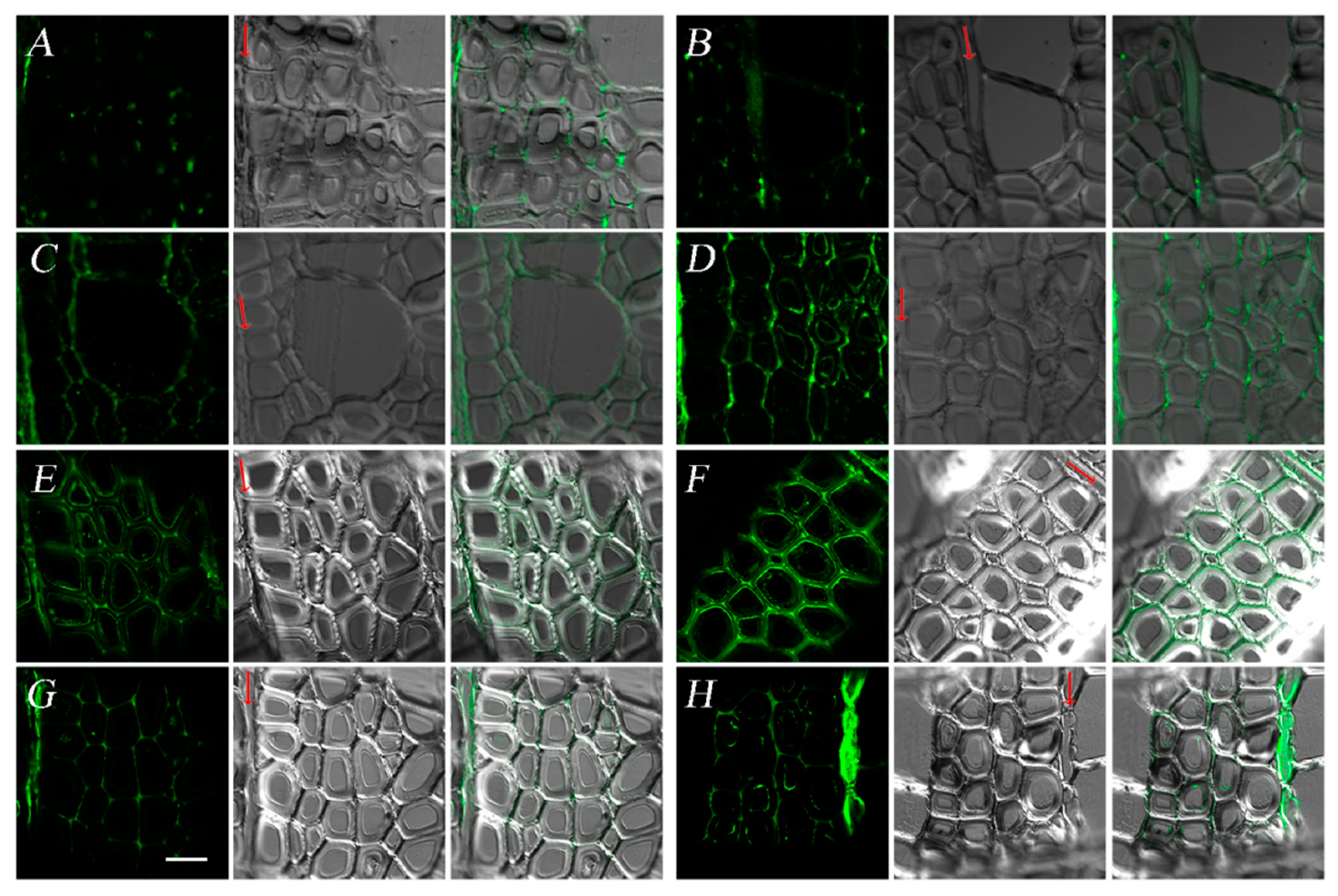
3.2. Heterogeneous Deposition of Lignin and Cellulose during Xylem Development
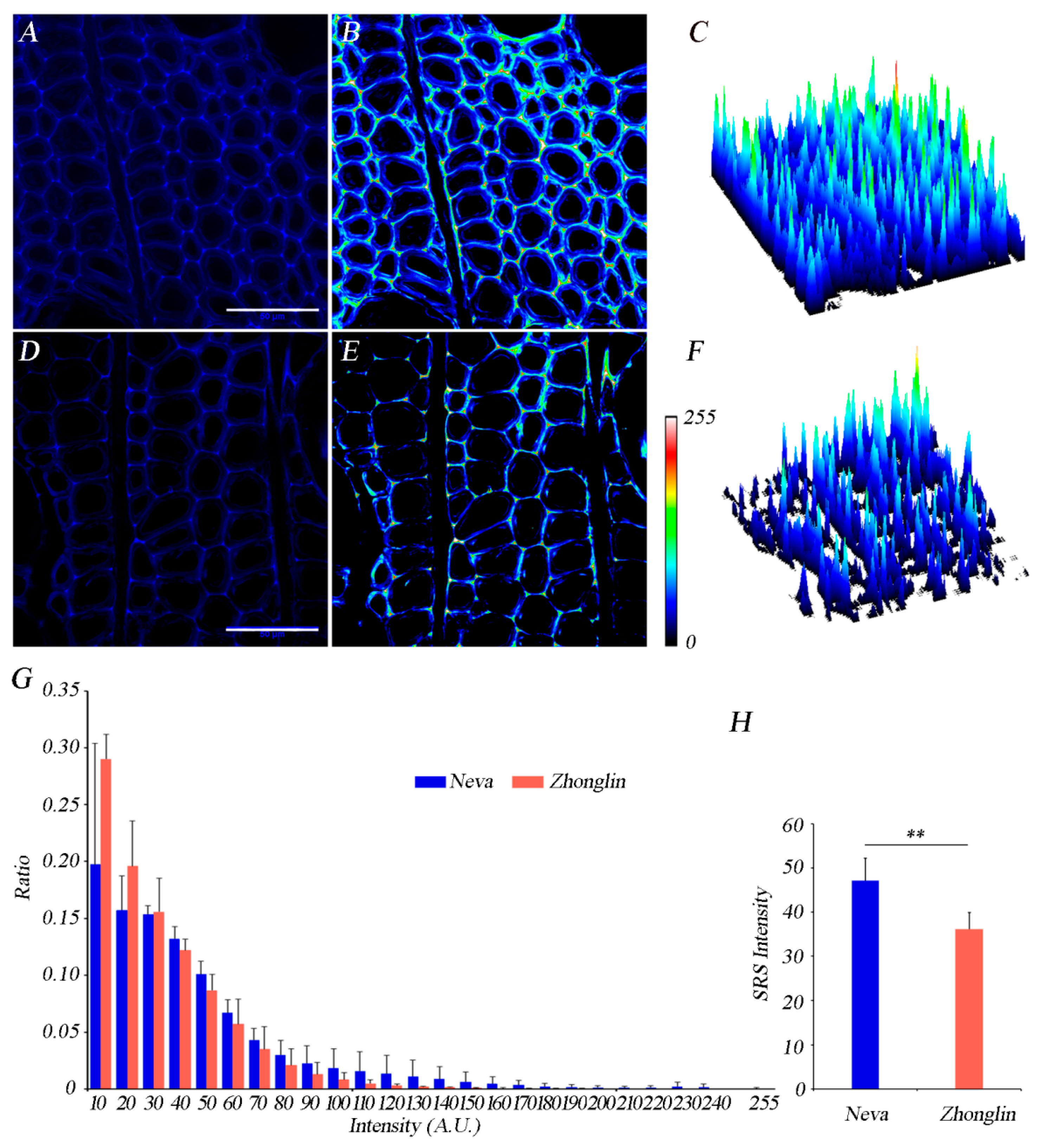
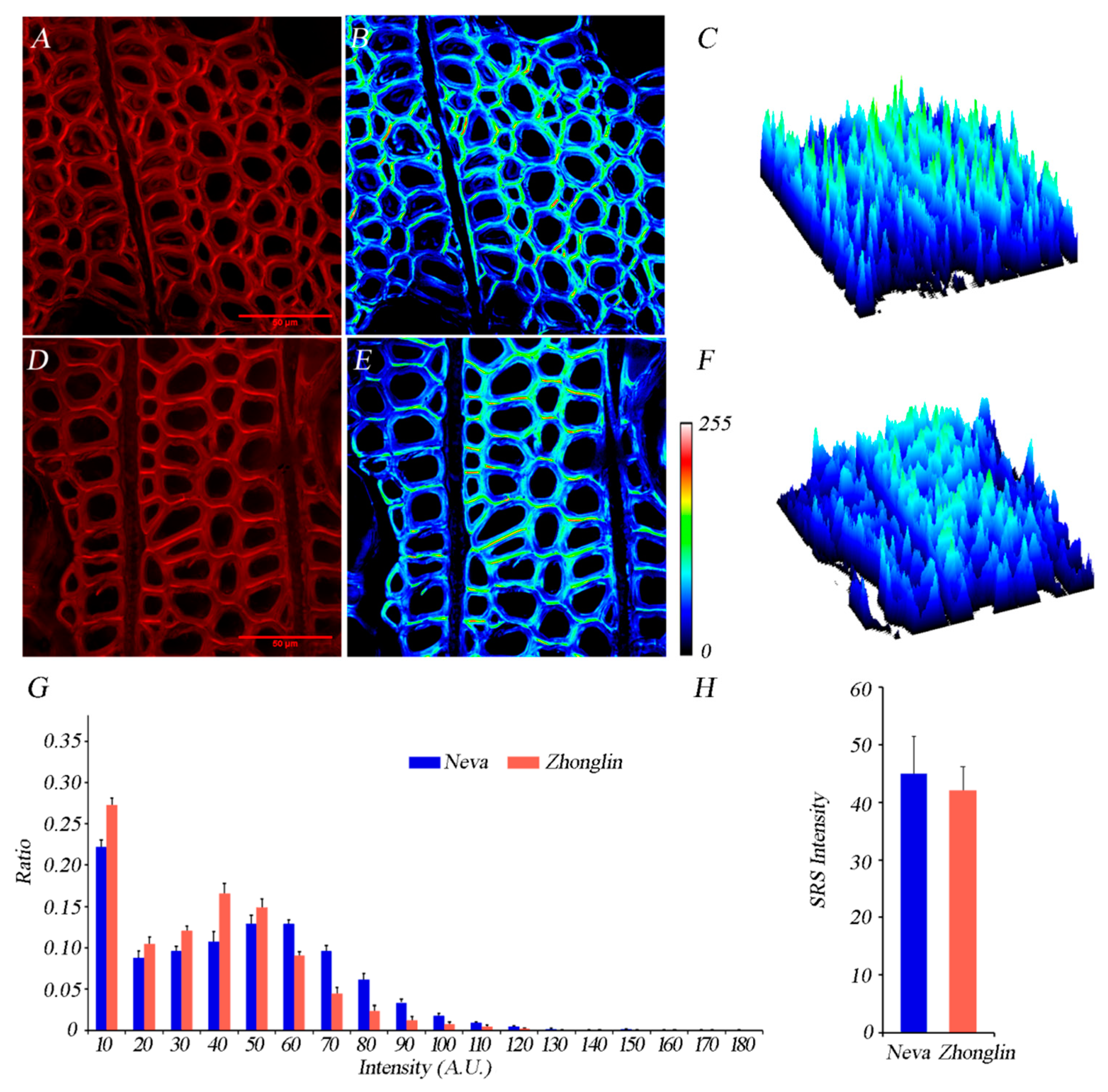
3.3. Differences of Lignin and Cellulose Deposition in Matured Xylem in the Two Cultivars
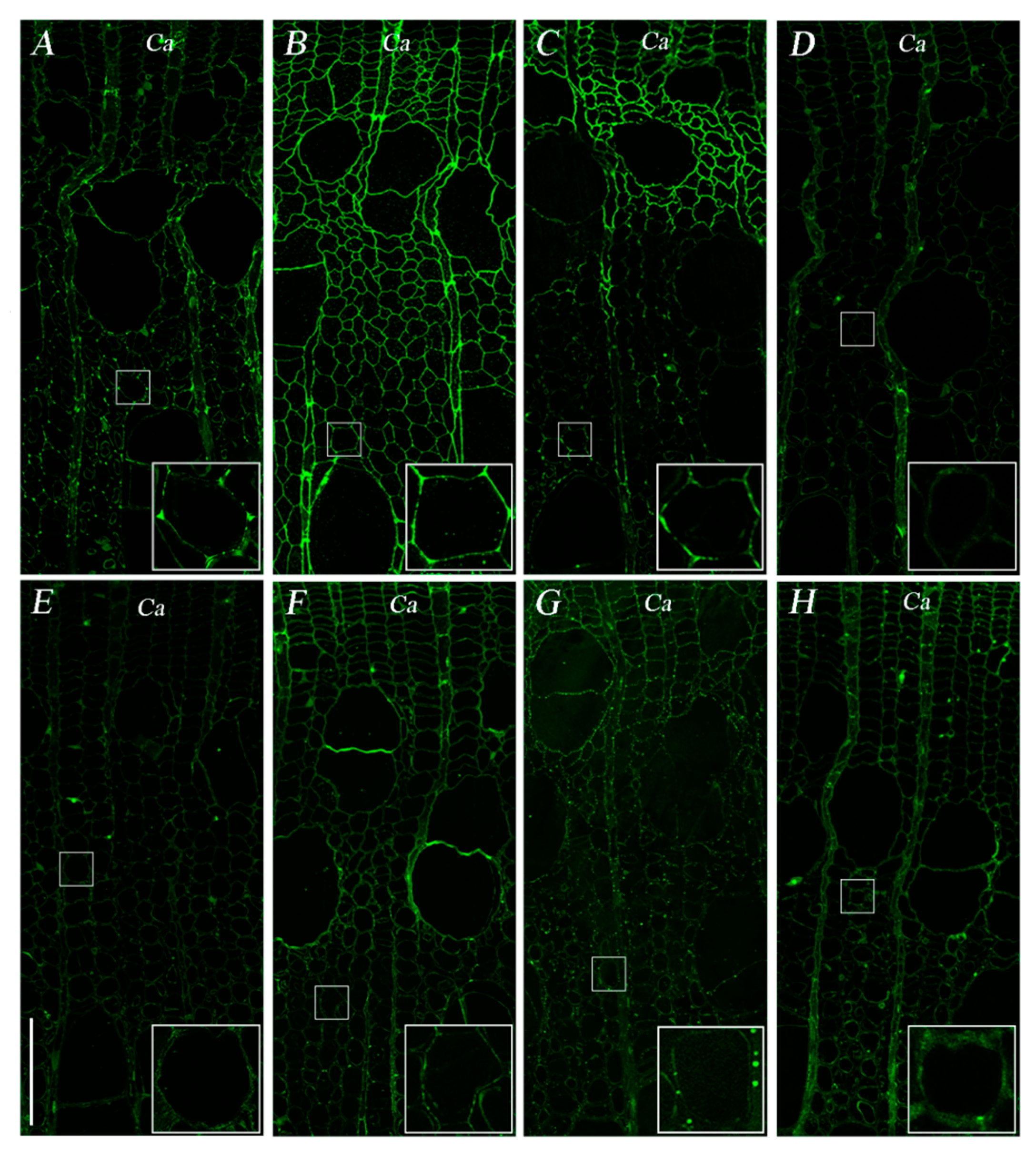
3.4. Difference in Pectin Patterns between the Two Cultivars
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Samuels, A.L.; Samuels, A.L.; Kaneda, M.; Kaneda, M.; Rensing, K.H.; Rensing, K.H. The cell biology of wood formation: From cambial divisions to mature secondary xylem. Can. J. Bot. 2006, 84, 631–639. [Google Scholar] [CrossRef]
- Lucas, W.J.; Groover, A.; Lichtenberger, R.; Furuta, K.; Yadav, S.R.; Helariutta, Y.; He, X.Q.; Fukuda, H.; Kang, J.; Brady, S.M. The plant vascular system: Evolution, development and functions. J. Integr. Plant Biol. 2013, 55, 294–388. [Google Scholar] [CrossRef]
- Meents, M.J.; Watanabe, Y.; Samuels, A.L. The cell biology of secondary cell wall biosynthesis. Ann. Bot. 2018, 121, 107–1125. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lampugnani, E.R.; Khan, G.A.; Somssich, M.; Persson, S. Building a plant cell wall at a glance. J. Cell Sci. 2018, 131, jcs207373. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cosgrove, D.J.; Jarvis, M.C. Comparative structure and biomechanics of plant primary and secondary cell walls. Front. Plant Sci. 2012, 3, 204. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Höfte, H.; Voxeur, A. Plant cell walls. Curr. Biol. 2017, 27, R865. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, B.; Gao, Y.; Zhang, L.; Zhou, Y. The plant cell wall: Biosynthesis, construction, and functions. J. Integr. Plant Biol. 2021, 63, 251–272. [Google Scholar] [CrossRef] [PubMed]
- Vanholme, R.; Demedts, B.; Morreel, K.; Ralph, J.; Boerjan, W. Lignin biosynthesis and structure. Plant Physiol. 2010, 153, 895–905. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Voxeur, A.; Höfte, H. Cell wall integrity signaling in plants: “To grow or not to grow that’s the question”. Glycobiology 2016, 26, 950–960. [Google Scholar] [CrossRef]
- Plomion, C.; Leprovost, G.; Stokes, A. Wood formation in trees. Plant Physiol. 2001, 127, 1513–1523. [Google Scholar] [CrossRef]
- Abedi, T.; Castilleux, R.; Nibbering, P.; Niittylä, T. The spatio-temporal distribution of cell wall-associated glycoproteins during wood formation in Populus. Front. Plant Sci. 2020, 11, 611607. [Google Scholar] [CrossRef] [PubMed]
- Pauly, M.; Keegstra, K. Plant cell wall polymers as precursors for biofuels. Curr. Opin. Plant Biol. 2010, 13, 305–312. [Google Scholar] [CrossRef]
- Zeng, Y.; Himmel, M.E.; Ding, S.Y. Visualizing chemical functionality in plant cell walls. Biotechnol. Biofuels 2017, 10, 263. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zeng, Y.; Zhao, S.; Yang, S.; Ding, S.Y. Lignin plays a negative role in the biochemical process for producing lignocellulosic biofuels. Curr. Opin. Biotechnol. 2014, 27, 38–45. [Google Scholar] [CrossRef]
- Xu, H.; Zhao, Y.; Suo, Y.; Guo, Y.; Man, Y.; Jing, Y.; He, X.; Lin, J. A label-free, fast and high-specificity technique for plant cell wall imaging and composition analysis. Plant Methods 2021, 17, 29. [Google Scholar] [CrossRef] [PubMed]
- Guedes, F.T.P.; Laurans, F.; Quemener, B.; Assor, C.; Lainé-Prade, V.; Boizot, N.; Vigouroux, J.; Lesage-Descauses, M.C.; Leplé, J.C.; Déjardin, A.; et al. Non-cellulosic polysaccharide distribution during G-layer formation in poplar tension wood fibers: Abundance of rhamnogalacturonan I and arabinogalactan proteins but no evidence of xyloglucan. Planta 2017, 246, 857–878. [Google Scholar] [CrossRef] [PubMed]
- Jin, K.; Liu, X.; Wang, K.; Jiang, Z.; Tian, G.; Yang, S.; Shang, L.; Ma, J. Imaging the dynamic deposition of cell wall polymer in xylem and phloem in Populus × euramericana. Planta 2018, 248, 849–858. [Google Scholar] [CrossRef] [PubMed]
- Soukup, A. Selected simple methods of plant cell wall histochemistry and staining for light microscopy. Methods Mol. Biol. 2019, 1992, 27–42. [Google Scholar] [PubMed]
- Wu, H.; Xu, H.; Li, H.; Wei, D.; Lin, J.; Li, X. Seasonal development of cambial activity in relation to xylem formation in Chinese fir. J. Plant Physiol. 2016, 195, 23–30. [Google Scholar] [CrossRef] [PubMed]
- Fromm, J.; Rockel, B.; Lautner, S.; Windeisen, E.; Wanner, G. Lignin distribution in wood cell walls determined by TEM and backscattered SEM techniques. J. Struct. Biol. 2003, 143, 77–84. [Google Scholar] [CrossRef]
- Donaldson, L.A.; Knox, J.P. Localization of cell wall polysaccharides in normal and compression wood of radiata pine: Relationships with lignification and microfibril orientation. Plant Physiol. 2012, 158, 642–653. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Man, Y.; Zhao, Y.Y.; Ye, R.; Lin, J.X.; Jing, Y.P. In vivo cytological and chemical analysis of Casparian strips using stimulated Raman scattering microscopy. J. Plant Physiol. 2018, 220, 136–144. [Google Scholar] [CrossRef]
- Zhang, L.; Liu, B.; Zhang, J.; Hu, J. Insights of molecular mechanism of xylem development in five black poplar cultivars. Front. Plant Sci. 2020, 11, 620. [Google Scholar] [CrossRef]
- Song, X.L.; Yao, C.L.; Le, W.; Zou, X. Analysis of wood property of four species of Populus euramericanas and studies on their pulping performance. China Puplp Pap. Ind. 2010, 31, 34–38. [Google Scholar]
- Liu, Y.; Zhou, L.; Zhu, Y.; Liu, S. Anatomical features and its radial variations among different catalpa bungei clones. Forests 2020, 11, 824. [Google Scholar] [CrossRef]
- Istikowati, W.I.; Aiso, H.; Ishiguri, F.; Sutiya, B.; Ohshima, J.; Iizuka, K.; Yokota, S. Study of radial variation in anatomical characteristics of three native fast-growing tree species of a secondary forest in South Kalimantan. Indones. Appita. J. 2016, 69, 49–56. [Google Scholar]
- Chen, H.M.; Han, J.J.; Cui, K.M.; He, X.Q. Modification of cambial cell wall architecture during cambium periodicity in Populus tomentosa Carr. Trees 2010, 24, 533–540. [Google Scholar] [CrossRef]
- Li, H.; Jiang, X.; Ramaswamy, H.S.; Zhu, S.; Yu, Y. High-pressure treatment effects on density profile, surface roughness, hardness, and abrasion resistance of paulownia wood boards. Trans. ASABE 2018, 61, 1181–1188. [Google Scholar] [CrossRef]
- Ververis, C.; Georghiou, K.; Christodoulakis, N.; Santas, P.; Santas, R. Fiber dimensions, lignin, and cellulose content of various plant materials and their suitability for paper production. Ind. Crop. Prod. 2003, 19, 245–254. [Google Scholar] [CrossRef]
- Francis, R.C.; Brown, A.F.; Bosshart, R.R.; Balch, S.; Waite, D.M. Four additional hybrid poplars with papermaking properties superior to aspen. TAPPI J. 2005, 4, 13–17. [Google Scholar]
- Seth, R.S.; Page, D.H. Fiber properties and tearing resistance. Tappi. J. 1988, 71, 103–107. [Google Scholar]
- Ona, T.; Sonoda, T.; Ito, K.; Shibata, M.; Tamai, Y.; Kojima, Y.; Ohshima, J.; Yokota, S.; Yoshizawa, N. Investigation of relationship between cell and pulp properties in Eucalyptus by examination of within-tree property variations. Wood. Sci. Technol. 2001, 35, 229–243. [Google Scholar] [CrossRef]
- Freudiger, C.W.; Min, W.; Saar, B.G.; Lu, S.; Holtom, G.R.; He, C.; Tsai, J.C.; Kang, J.X.; Xie, X.S. Label-free biomedical imaging with high sensitivity by stimulated Raman scattering microscopy. Science 2008, 322, 1857–1861. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, X.; Ma, J.; Zhe, J.; Yang, G.H.; Zhou, X.; Xu, F. Method for automatically identifying spectra of different wood cell wall layers in Raman imaging data set. Microsc. Res. Technol. 2014, 77, 609–618. [Google Scholar] [CrossRef] [PubMed]
- Gierlinger, N.; Schwanninger, M. Chemical imaging of poplar wood cell walls by confocal Raman microscopy. Plant Physiol. 2006, 140, 1246–1254. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, L.; Yao, C.L.; Song, X.L.; Zou, X. Infection of the different soils and different tree-ages of chemical compositions in Populus×euramericana Cv.‘74/76′. Pap. Pap. Mak. 2010, 29, 25–28. [Google Scholar]
- De Meester, B.; Calderón, B.M.; de Vries, L.; Pollier, J.; Goeminne, G.; Van Doorsselaere, J.; Chen, M.; Ralph, J.; Vanholme, R.; Boerjan, W. Tailoring poplar lignin without yield penalty by combining a null and haploinsufficient CINNAMOYL-CoA REDUCTASE2 allele. Nat. Commun. 2020, 11, 5020. [Google Scholar] [CrossRef]
- Hu, J.Q.; Qi, Q.; Zhao, Y.L.; Tian, X.M.; Lu, H.; Gai, Y.; Jiang, X.N. Unraveling the impact of Pto4CL1 regulation on the cell wall components and wood properties of perennial transgenic Populus tomentosa. Plant Physiol. Biochem. 2019, 139, 672–680. [Google Scholar] [CrossRef]
- Shafrin, F.; Ferdous, A.S.; Sarkar, S.K.; Ahmed, R.; Amin, A.; Hossain, K.; Sarker, M.; Rencoret, J.; Gutiérrez, A.; Del Rio, J.C.; et al. Modification of monolignol biosynthetic pathway in Jute: Different gene, different consequence. Sci. Rep. 2017, 7, 39984. [Google Scholar] [CrossRef] [PubMed]
- Bai, Y.; Wu, D.; Liu, F.; Li, Y.; Chen, P.; Lu, M.; Zheng, B. Characterization and functional analysis of the poplar Pectate Lyase-Like Gene PtPL1-18 reveal its role in the development of vascular tissues. Front. Plant Sci. 2017, 8, 1123. [Google Scholar] [CrossRef] [Green Version]
- Willats, W.G.; Gilmartin, P.M.; Mikkelsen, J.D.; Knox, J.P. Cell wall antibodies without immunization: Generation and use of de-esterified homogalacturonan block-specific antibodies from a naive phage display library. Plant J. 1999, 18, 57–65. [Google Scholar] [CrossRef] [PubMed]
- Wolf, S.; Mouille, G.; Pelloux, J. Homogalacturonan methyl-esterification and plant development. Mol. Plant 2009, 2, 851–860. [Google Scholar] [CrossRef] [PubMed]
- Leśniewska, J.; Öhman, D.; Krzesłowska, M.; Kushwah, S.; Barciszewska-Pacak, M.; Kleczkowski, L.A.; Sundberg, B.; Moritz, T.; Mellerowicz, E.J. Defense responses in aspen with altered pectin methylesterase activity reveal the hormonal inducers of tyloses. Plant Physiol. 2017, 173, 1409–1419. [Google Scholar] [CrossRef] [PubMed]







Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sun, N.; Bu, Y.; Pan, C.; Wu, X.; Cao, Y.; Jing, Y. Analyses of Microstructure and Dynamic Deposition of Cell Wall Components in Xylem Provide Insights into Differences between Two Black Poplar Cultivars. Forests 2021, 12, 972. https://0-doi-org.brum.beds.ac.uk/10.3390/f12080972
Sun N, Bu Y, Pan C, Wu X, Cao Y, Jing Y. Analyses of Microstructure and Dynamic Deposition of Cell Wall Components in Xylem Provide Insights into Differences between Two Black Poplar Cultivars. Forests. 2021; 12(8):972. https://0-doi-org.brum.beds.ac.uk/10.3390/f12080972
Chicago/Turabian StyleSun, Na, Yufen Bu, Chen Pan, Xinyuan Wu, Yuan Cao, and Yanping Jing. 2021. "Analyses of Microstructure and Dynamic Deposition of Cell Wall Components in Xylem Provide Insights into Differences between Two Black Poplar Cultivars" Forests 12, no. 8: 972. https://0-doi-org.brum.beds.ac.uk/10.3390/f12080972