
Tissue Structure Changes of Aquilaria sinensis Xylem after Fungus Induction


1
Forestry College, Guangxi University, East University Road No. 100, Nanning 530000, China
2
School of Resources, Environment and Materials of Guangxi University, East University Road No. 100, Nanning 530000, China
*
Author to whom correspondence should be addressed.
Forests 2022, 13(1), 43; https://0-doi-org.brum.beds.ac.uk/10.3390/f13010043
Submission received: 24 November 2021
/
Revised: 23 December 2021
/
Accepted: 27 December 2021
/
Published: 1 January 2022
(This article belongs to the Special Issue Wood Growth and Structure)
Abstract
:In this study, we analyzed the mechanism and the process of fungal-induced agarwood formation in Aquilaria sinensis and studied the functional changes in the xylem structure after the process. The microscopic structure of the white zone, transition zone, agarwood zone, and decay zone of 12-and 18-months of inoculation A. sinensis xylem was studied. The distribution of nuclei, starch grains, soluble sugars, sesquiterpenes, fungal propagules, and mycelium in xylem tissues was investigated by histochemical analysis. The results show that the process of agarwood formation was accompanied by apoptosis of parenchyma cells such as interxylary phloem, xylem rays, and axial parenchyma. Regular changes in the conversion of starch grains to soluble sugars, the production of sesquiterpenoids, and other characteristic components of agarwood in various types of parenchyma cells were also observed. The material transformation was concentrated in the interxylary phloem, providing a structural and material basis for the formation of agarwood. It is the core part of the production of sesquiterpenoids and other characteristic products of agarwood. Compared with the A. sinensis inoculated for 12 months, the xylem of the A. sinensis inoculated for 18 months was more vigorous. There were no significant differences between the 12 and 18 months of inoculation in terms of sugars and agarwood characteristic products. In production, harvesting after 12 months of inoculation can improve harvesting efficiency.
1. Introduction
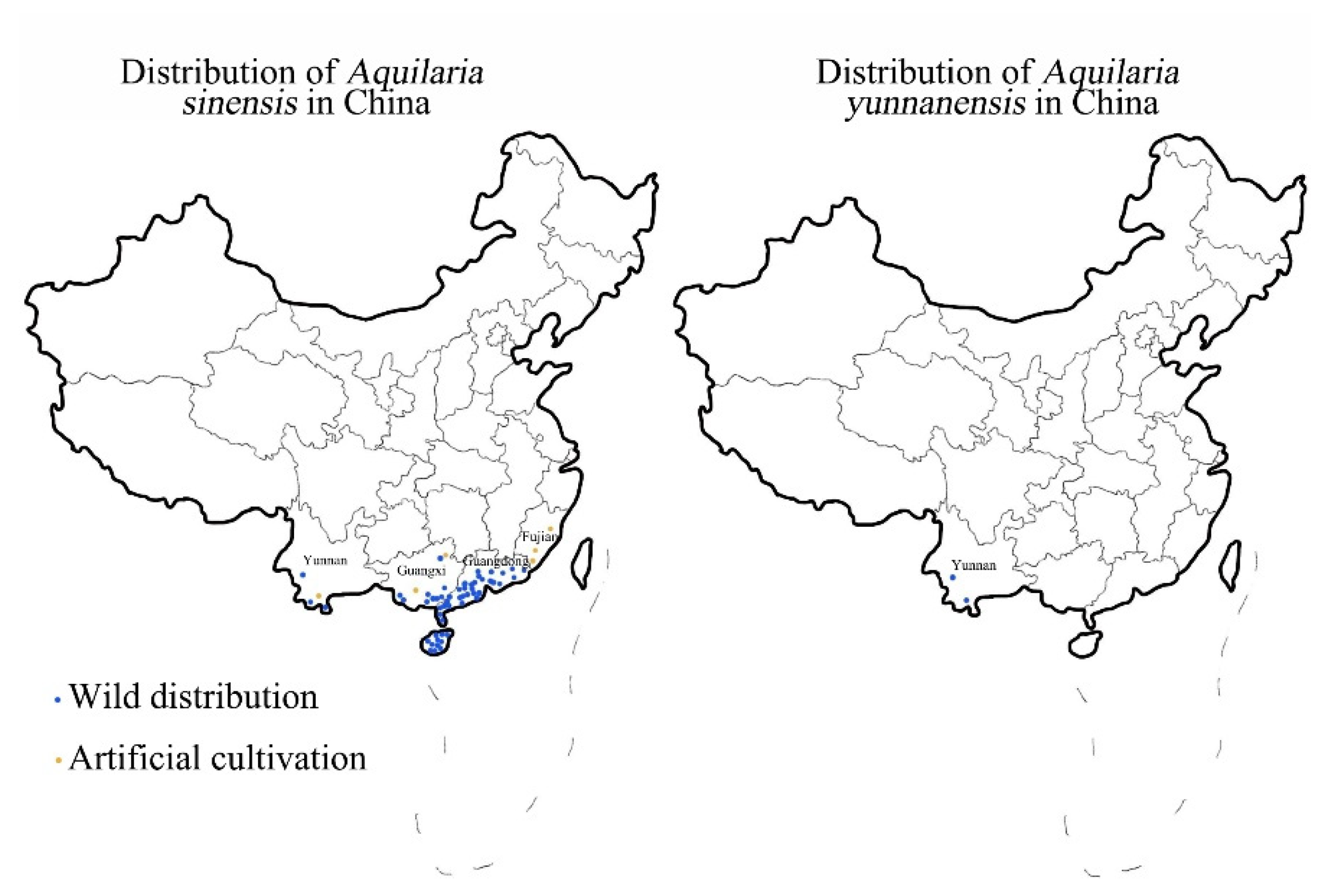
When Aquilaria trees of the Thymelaeaceae family are subjected to external stress or injury, a solid-state coagulation of resin and lignin is produced at the wound site, which along with the xylem, is termed agarwood. Agarwood is a precious natural component that has medicinal properties for treating chest pains and asthma. The native species of Aquilaria trees in China are Aquilaria sinensis and Aquilaria yunnanensis [1], the distribution of which is shown in Figure 1. Compared with A. yunnanensis, A. sinensis has a larger distribution area and stronger adaptability to the climate [2], making it the most common tree in China for agarwood production. There is a great demand for agarwood because of its medicinal and aromatic properties which have led to the predatory logging of wild resources. A. sinensis was included in the List of National Key Protected Wild Plants in 1999 and listed as a Class II endangered plant in China [3]. All Aquilaria species are listed in Appendix II of the Convention on International Trade in Endangered Species of Wild Fauna and Flora [4]. There are only 130,000 wild A. sinensis plants left, and the annual production of natural agarwood is 118 tons, accounting for only 23.6% of the market demand. Due to the scarcity of naturally occurring agarwood, artificial techniques have been effective for increasing the production of agarwood and meeting market demands [5]. The differences between artificial agarwood and natural agarwood are mainly in the types and relative contents of sesquiterpenoids and 2-(2-phenylethyl) chromones. Short-term formation of artificial agarwood and natural agarwood formed over the years is still a certain gap but has been able to meet the national standards and enough to apply in production. Due to the rarity and high cost of natural agarwood, almost all of the agarwood currently for medicine, daily use, and spices is artificial agarwood. It can be seen that artificial agarwood has been able to replace natural agarwood to a certain extent.
Agarwood-inducing techniques can be divided into physical trauma, chemical induction, and biological induction methods depending on the treatment method. The physical trauma method refers to cutting and burning, which are more traditional, less efficient, and uncontrollable. Chemical induction methods include ordinary chemical methods, such as injecting inorganic salts and other chemicals that can promote the formation of agarwood, and whole-tree agarwood-inducing techniques (Agar-Wit) [6,7]. There are drawbacks with both methods, such as complicated operation processes, chemical residues, and unstable quality [8,9,10]. Biological induction methods usually refer to fungus induction technology, in which mycelium or fungal endophyte culture supernatants gathered from the agarwood-containing A. sinensis that produces agarwood is inoculated into a healthy A. sinensis trunk [6,7,11,12]. The quality of agarwood produced by fungus induction is comparable to that of naturally occurring agarwood [13]. At present, biological induction methods are widely used in production and have a greater prospect of development. However, the quality of agarwood obtained from different culture conditions and different strain treatments vary greatly [14]. Moreover, characteristic agarwood components such as baimuxinal and α-copaen-11-ol were lower than those of the physical trauma method [15], whereas the requirements for the operating environment and operating techniques were higher. The current A. sinensis plantations usually use traditional agarwood techniques, such as physical and chemical agarwood methods, that generally require older trees (generally more than ten years) with labor-intensive operation and low yield, and some methods even produce chemical pollution and other problems. Therefore, artificial agarwood cannot form large-scale industrialization at present.
In recent years, many studies have been conducted on the structural and functional changes in the xylem of A. sinensis during fungal infection-induced agarwood by histochemical analysis, scanning electron microscopy and transmission electron microscopy. It was found that all physiological metabolic activities of A. sinensis, including agarwood formation, were mainly performed by parenchyma cells. The parenchyma cells in the xylem of A. sinensis include interxylary phloem, xylem rays, and axial parenchyma. The various types of parenchyma cells form a network of living cells in the wood [16]. Agarwood first appears in the parenchyma cells and then enters the surrounding vessels and wood fibers [17]. Although the interxylary phloem, xylem rays, and axial parenchyma are all parenchyma cells that can store and transport and are capable of respiration and metabolism, they have different physiological functions [18,19,20]. The interxylary phloem of A. sinensis xylem differentiates into meristem tissue when subjected to external stimuli and cornifies the cells in the middle of the interxylary phloem, initiating the physical defense process [21,22]. Axial parenchyma is important to the distribution of nutrients, thickens the cytoderm defensively under external stress, and produces agarwood [23,24].
In the previous study, the contents of alcohol-soluble extracts and essential oils of the agarwood induced by the fungal inducer were analyzed and its chemical composition was analyzed by gas chromatography-mass spectrometry (GC–MS). The characteristic substances were quantitatively analyzed by high-performance liquid chromatography (HPLC), and ultraviolet-visible spectrophotometry (UV–vis) was used to analyze the total chromone content. Through these experiments, we found that the content of the characteristic products of agarwood in the tested samples met the requirements of industry standards. This suggests that fungal inoculation had the effect of inducing the formation of agarwood from A. sinensis [25]. Furthermore, we found that the artificial agarwood had similar biological activity to the wild agarwood [26]. This proved that the fungus-induced agarwood had medicinal value.
The role played by various types of parenchyma cell tissue types during agarwood formation is unclear. The mode of movement and the action site of the fungus after infecting the tree are not well known.
In this study, the microstructural characteristics of A. sinensis at 12 and 18 months of inoculation were observed to investigate the differences and changes in various types of parenchyma cells, as well as the movement trends of mycelium in the xylem. The aim of this study was to analyze the effect of fungal induction on the structure of parenchyma cells in the xylem of A. sinensis, and to lay the foundation for elucidating the action sites and mechanisms of fungal induction of agarwood formation.
2. Materials and Methods
2.1. Fungus Induction Operation and Wood Collection
A natural agarwood stump with fragrant ingredients for years, high oil content, good quality, and a bare cross-section was selected from the wild trees of A. sinensis in Xinyi City, Maoming City, Guangdong Province, China. This sample was used to prepare fungal inducers. Under aseptic conditions of a clean bench, the agarwood block material from wild A. sinensis was washed once with pure water to remove the decayed xylem. The agarwood block was cut into 5 cm × 5 cm pieces, soaked in 75% ethanol for 1 min, rinsed with sterile water for 1 h, then soaked in 0.1% HgCl2 solution for 15 min, rinsed with sterile water four times, and cut into 1 cm × 1 cm blocks with a sterilized blade. For the enrichment of fungi, the dried materials were cultured at 30 °C for 7 d in a shake flask that was placed in a sterile PDA liquid medium. After 7 d, the enriched fungal liquid and PDA liquid medium were cultured at a ratio of 1:100 (v:v) and 30 °C for 7 d to obtain the fungal inducer. Fungal species were identified by high-throughput sequencing. At the species level, 51 species were obtained. Aspergillus penicillioides was the most dominant species with a relative abundance of 31%. The remaining dominant species were mainly Gongronella butleri (11.38%), Aspergillus sydowii (8.69%), and Cladosporium halotolerans (8.36%), etc. [25]
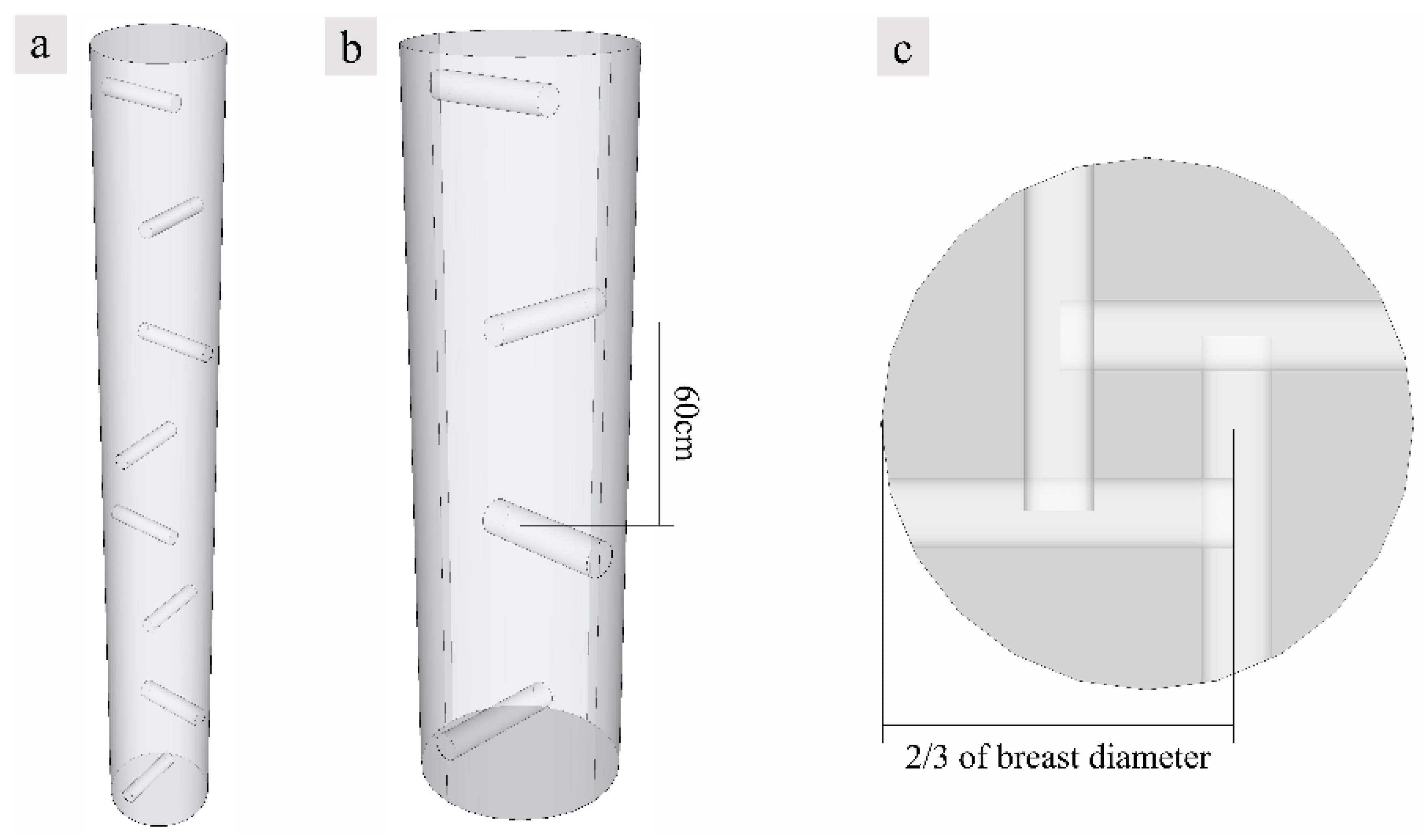
The experiment was performed at Senyuan Precious Tree Planting Base, North Sea city in Guangxi Municipality (21°48′26″ N, 109°10′20″ E) where the climate type is maritime monsoon, with an average altitude of 10~15 m. The forest type is subtropical broad-leaved forest, while the soil type is red soil and limestone soil. All trees were planted on flat ground with 3 m spacing for each planting row. The planting pattern was monoculture, and no other plants were planted in the forest. Fertilizer was applied and weeded every spring and fall, and no pesticides or herbicides were applied. A. sinensis usually blooms there from mid-March to mid-April and bears fruit from June to July. Healthy 10-year-old A. sinensis trees, with a uniform diameter (15.6 ± 0.8 cm) at breast height and uniform tree height (5 ± 0.5 m), were induced with fungus, and holes were drilled in the trunk surface in a spiral arrangement as shown in Figure 2. Spiral holes were drilled to a depth of 2/3 of the tree diameter into the A. sinensis, and 150 mL fungal inoculant was inoculated into each hole using a high-pressure perfusion device. The induced agarwood samples were collected after 12 and 18 months, respectively, and A. sinensis without fungi inoculation as the control blank groups were collected after 18 months. The samples were taken in November 2019 and May 2020. Three trees were selected each time, and the height and diameter at breast height were measured for each tree. After felling the trees, 3-cm thick wood slices were collected at 20 cm above the roots, the diameter at breast height, and the branching point, respectively. Three wood slices were collected from each part and marked, and two of them were placed in Formalin-Aceto-Alcohol (FAA) and dry ice while the other one was air-dried. The samples stored in dry ice were subsequently transferred to an ultra-low temperature refrigerator for cryopreservation.
2.2. Observation of Xylem Tissue Structure
The air-dried wood slices were analyzed from the outside to the inside of the white zone, transition zone, agarwood zone and decay zone, and cut into blocks to expose the transverse and radial surfaces. Macrostructural characteristics of the transverse and radial sections at each zone were observed using a stereo microscope (Nikon, Japan).
The cryopreserved wood slices were categorized into zones and cut into blocks. Then, 20-µm thick sections were sliced from the blocks using a freezing microtome (Leica CM1860 UV, Nussloch, Germany), and the transverse surface and radial surfaces were obtained. The sections preserved by FAA were stained by acetocarmine to detect nuclei. To identify sugars, the sections were stained with I2-KI to detect starch grains, whereas soluble sugars compounds were detected by staining with periodic acid-Schiff (PAS) reagent. The sections preserved in an ultra-low temperature refrigerator were fixed in saturated lead acetate for 36 h and then stained with acid vanillin to detect the sesquiterpenoids. All sections were observed under a light microscope (Nikon E100, Sendai-shi, Japan).
2.3. Scanning Electron Microscope (SEM)
The 60-µm thick sections were sliced from the blocks using a freezing microtome (Leica CM1860, Nussloch, Germany) to expose the transverse and radial surfaces. The blocks were air-dried for approximately 24 h and then observed by SEM after gold coating.
2.4. Data Statistics and Analysis
The number of cell nuclei and starch grains in an arbitrarily selected square with 100 μm sides were determined from micrographs at 40× magnification, and the morphology and the distribution locations were recorded. The soluble sugars and the sesquiterpenoids were counted by relative area, and the areas of dark secondary metabolites were determined. Micrographs at 10× and 40× magnification were selected, and the area of color development was determined using Image-Pro and plotted using Origin software.
Analysis of variance (ANOVA) was performed using SPSS for each indicator of inoculation time, xylem zone, and xylem zone with xylem structure such as the number of nuclei and starch grains, and relative area of soluble sugars and sesquiterpene. Data were first subjected to homogeneity test, and if p > 0.05, homogeneity of variance was satisfied. One-way ANOVA was then performed, and if p < 0.05, the factor was significantly different, then further multiple comparisons were performed by least significant difference (LSD). If p > 0.05, the factor was not significantly different.
3. Results
3.1. Macrostructural Differences at Different Zones of the Xylem
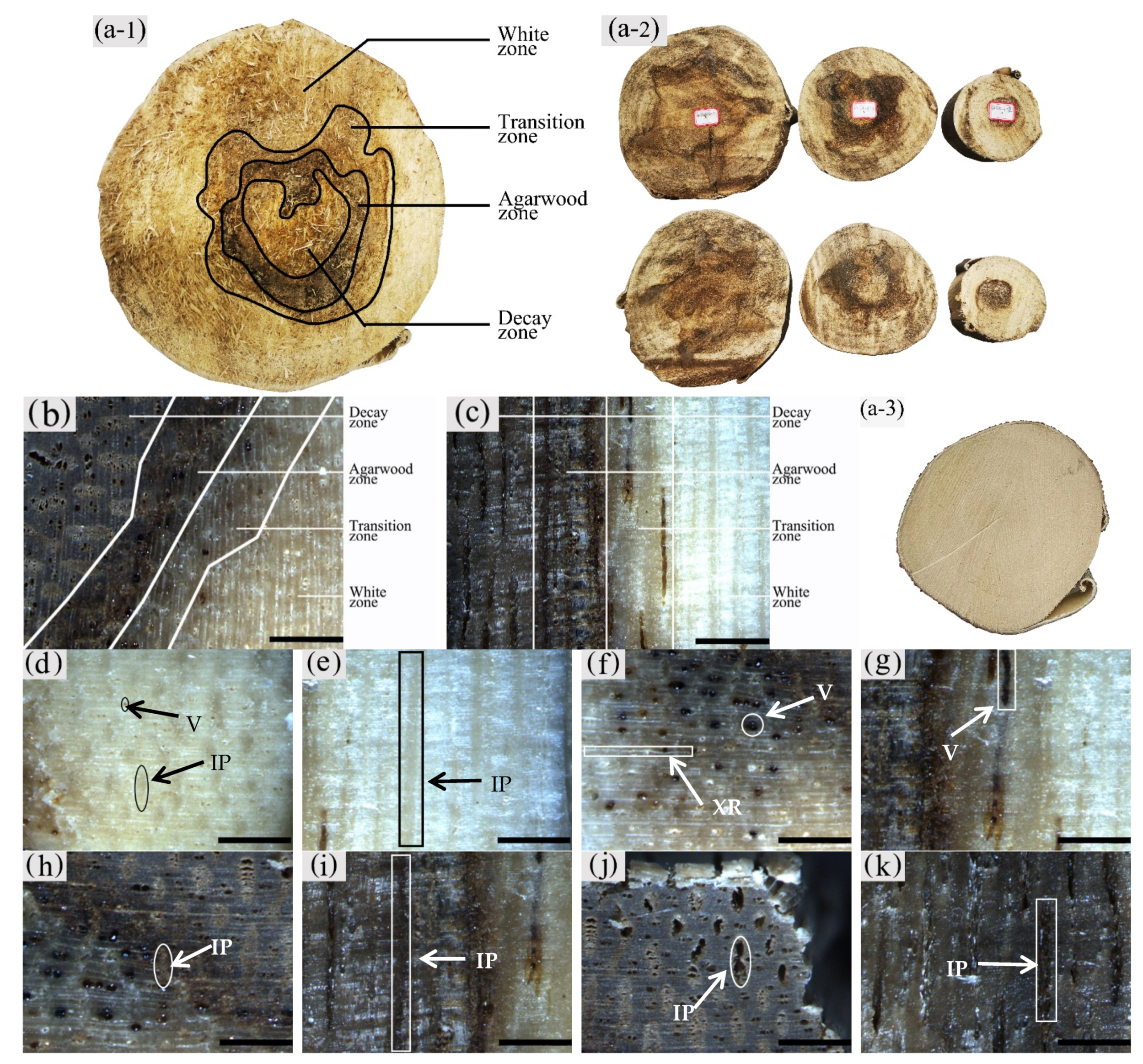
The sapwood of uninoculated A. sinensis was white and without an aromatic smell as shown in Figure 3(a-3). After agarwood formation, the black-brown pathological heartwood was produced, which had a greasy luster and a distinct agarwood odor. The central part of the xylem where agarwood production began could be divided radially into four different zones, from outside to inside: the white zone, transition zone, agarwood zone, and decay zone from outside to inside (Figure 3a–c). The wood slices collected at 20 cm above the roots, the diameter at breast height, and the branching point were as Figure 3(a-2). The main differences between zones were color, odor, and the distribution and morphology of parenchyma cells and vessels.
It was observed that the macrostructural of the white zone was similar to that of the blank control. The white zone was the outside area of the xylem without agarwood formation, where the wood color was white, and it did not have an aroma (Figure 3d,e). The transition zone was the area near the white zone, usually 5–10 mm wide, where an aromatic fragrance was first detected. The interxylary phloem was light brown to dark brown, the xylem ray was white to light brown, and the color of all tissues tended to deepen from the outside to the inside. The vessels with a distinct greasy luster were visible on the transverse surface (Figure 3f). The axial parenchyma was not noticeable on the radial surface; however, vessels filled with grease-like inclusions were observed. The agarwood zone had the main accumulation of agarwood. It was located on the inner side of the transition zone, usually 10–25 mm wide. It had a distinct odor and was dark brown to black in color. On the transverse surface, the yellowish-brown interxylary phloem and its adjacent xylem rays could be observed with the naked eye (Figure 3h), whereas the dark striped vessels and interxylary phloem were seen on the radial surface (Figure 3i). The decay zone was the innermost region of the xylem, without a noticeable odor or greasy sheen. It was typically light to dark brown in color with loose and fragile wood. The cavity-like interxylary phloem was visible to the naked eye on the transverse surfaces, and the xylem rays remained intact (Figure 3j). On the transverse surface, there were banded interxylary phloem and disconnected vessels, both of which were cavity-shaped (Figure 3k).
As shown in Figure 3(a-2) that compares the agarwood formation and stratification of the wood slices at different heights, the wood slices at the branching point just started to form agarwood, while the agarwood zone was located in the middle of the wood slices, and this was a small area of decay zone. At the breast diameter and the ground diameter, the agarwood zone expands outward, and the area of the decay zone increases. It can be seen that the agarwood was first produced in the pith of the tree, combined with the wood of the xylem to form the agarwood zone, and spread outward as the agarwood form.
The macroscopic structure of the xylem of A. sinensis inoculated for 12 and 18 months differed. In terms of zoning characteristics, the decay zone of A. sinensis with 12 months of inoculation was lighter in color, usually light yellow-brown, and the agarwood zone had a similar width with a distinct transitional zone and a wider white zone.
3.2. Observation on the Nucleus of Parenchyma Cells in the Xylem of A. sinensis
The morphology of the nuclei in different zones of the xylem is shown in Figure 4, and the statistics are shown in Figure 5.
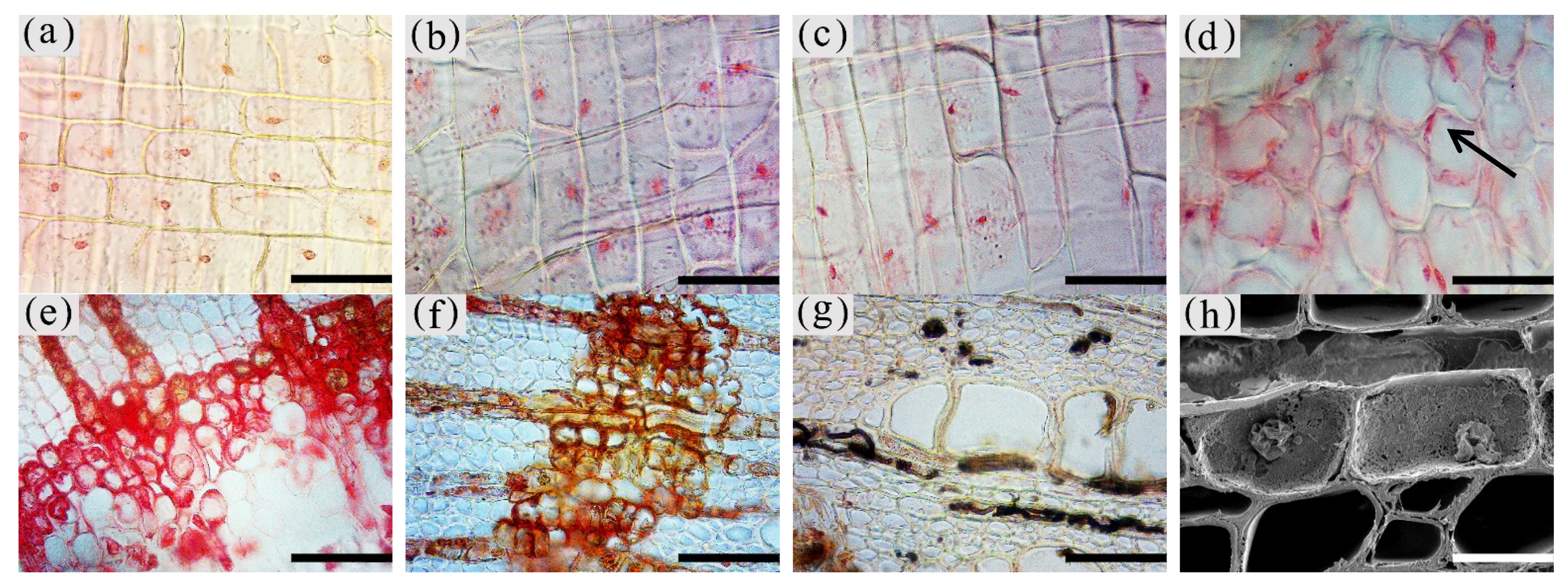
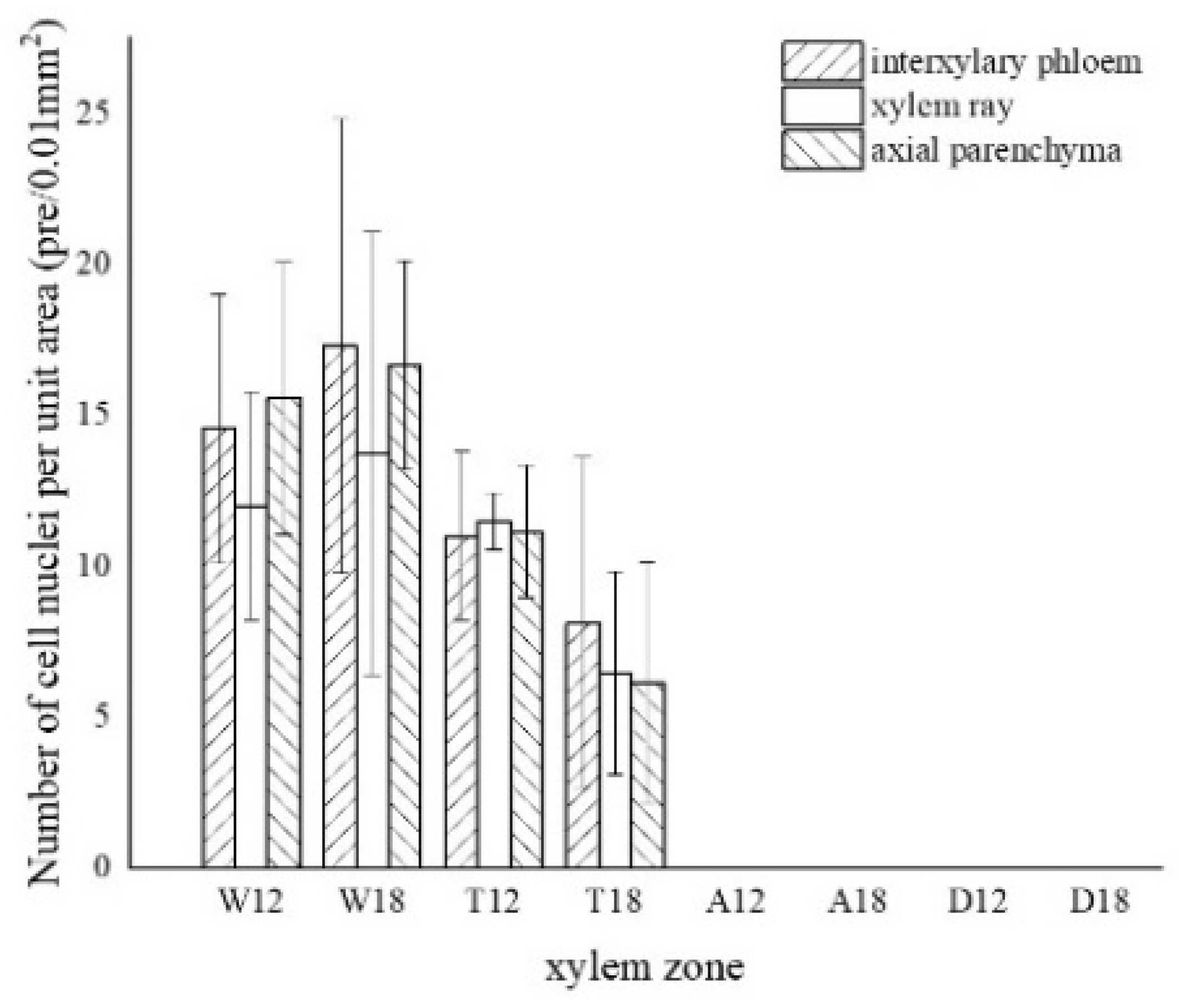
The nuclei in the white zone were full and rounded and were mainly distributed in the parenchyma cells which were similar to the nuclei of the blank control (as in Figure 4a). The average number of nuclei in the white zone was 14.98 nuclei/10−2 mm2. The highest number of nuclei was in the axial parenchyma, approximately 16.13 nuclei/10−2 mm2, followed by the interxylary phloem with approximately 15.95 nuclei/10−2 mm2. The lowest number of nuclei was in the xylem rays, approximately 12.87 nuclei/10−2 mm2. On the radial surface, there were more nuclei in the xylem rays near the cambium, but they gradually decreased closer to the pith. The micrograph of xylem rays on the outside of the white zone shows that nuclei were present in each cell and were more rounded, as shown in Figure 4b. The micrograph of the xylem rays on the inside of the white zone is shown in Figure 4c, with no nuclei in some cells and fewer nuclei overall than the outer white zone, and the nuclei started to crumple and became oval. Nuclei were observed in a few xylem rays and interxylary phloem in the transition zone. They were usually pyknotic and close to the cell wall (Figure 4d at the arrow). They were considerably crinkled, as observed by SEM (Figure 4h). No nuclei were observed in the inner part of the transition zone near the agarwood zone (Figure 4e). The number of nuclei in the transitional zone was less than that in the white zone, with an average of 9.06 nuclei/10−2 mm2. The highest number of nuclei in the interxylary phloem was approximately 9.57 nuclei/10−2 mm2, followed by the xylem rays at about 8.97 nuclei/10−2 mm2. The lowest number of nuclei was in the axial parenchyma, approximately 8.64 nuclei/10−2 mm2. Cells of interxylary phloem and xylem rays in the agarwood zone were stained as a whole, and no nuclei were observed (Figure 4f). There was no discoloration in any cells in the decay area (Figure 4g). Nuclei were found to be more abundant in 18 months than 12 months of inoculation, with the greatest number in the interxylary phloem, whereas 12 months of inoculation was mainly distributed in the axial parenchyma.
3.3. Distribution of Sugars in the Xylem of A. sinensis
The morphology of starch grains in different zones of xylem is shown in Figure 6, and the statistics are shown in Figure 7.
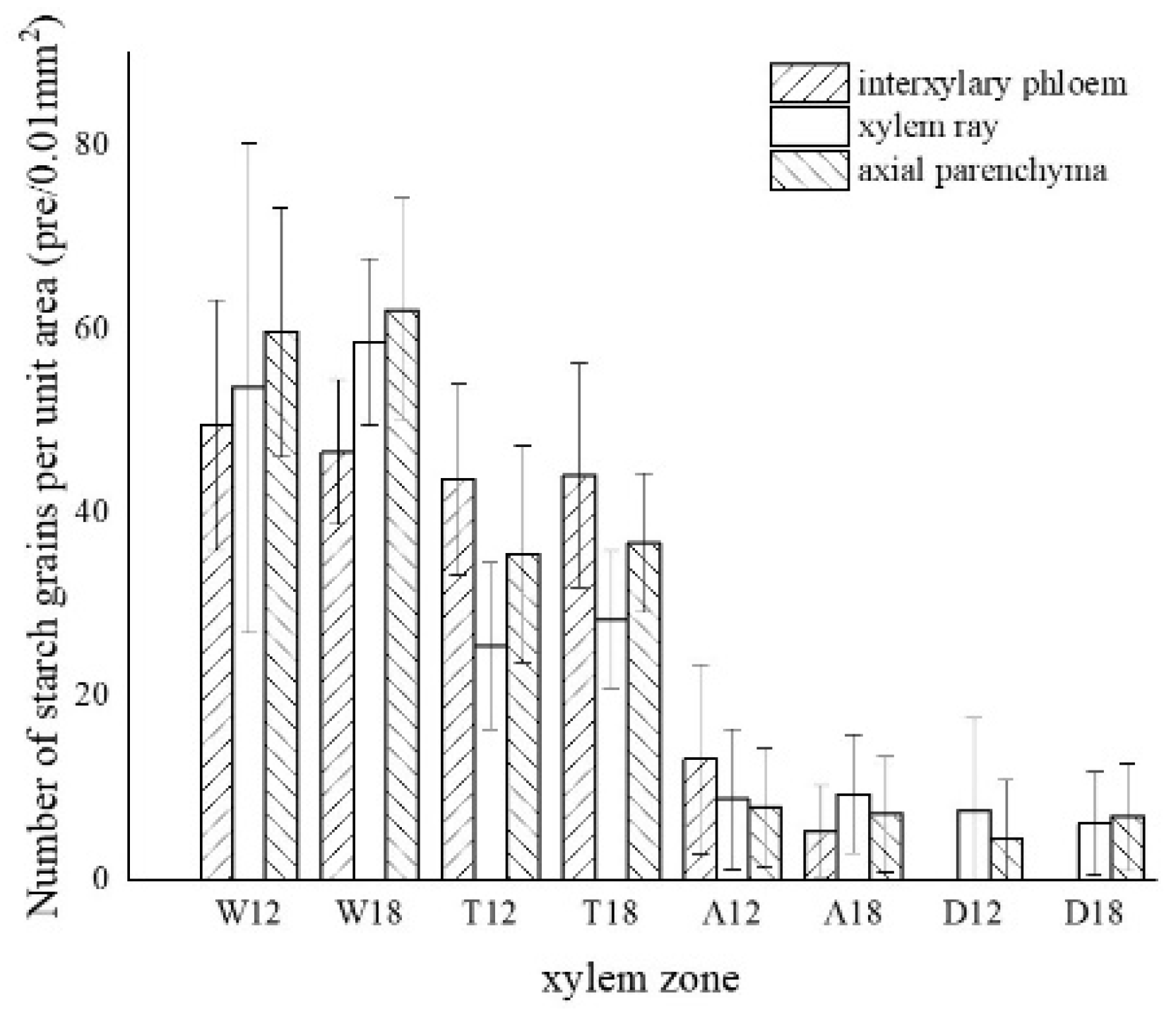
The starch grains in the blank control were mainly distributed in the xylem rays, axial parenchyma, and interxylary phloem, which were rounded and more densely distributed (Figure 6a). In the white zone, starch grains were concentrated in the xylem rays and axial parenchyma, which were similar to the blank control, with an average of approximately 55.04 grains/10−2 mm2. The starch grains in the xylem rays were arranged more closely and a noticeable group-like arrangement was observed, with approximately 56.13 grains/10−2 mm2. The distribution of starch grains in the axial parenchyma and interxylary phloem were more scattered and arranged in a striped pattern (Figure 6b,c), with approximately 60.91 grains/10−2 mm2 and 48.08 grains/10−2 mm2, respectively. The number of starch grains in the xylem rays of the white zone gradually decreased from the outside to the inside, as shown in Figure 6d. The number of starch grains in the transition zone was significantly reduced, with an average number of 35.62 grains/10−2 mm2. The starch grains were mainly distributed in xylem rays, axial parenchyma, and interxylary phloem, where the majority were in the interxylary phloem, approximately 43.85 grains/10−2 mm2 (Figure 6e). The axial parenchyma contained approximately 36.09 grains/10−2 mm2, and the number of wood ray starch grains was the lowest with approximately 26.93 grains/10−2 mm2 (Figure 6f). In this region, the number of starch grains gradually decreased from the outside to the inside, with almost no starch grains observed in the area near the agarwood zone (Figure 6g). The agarwood zone contained an average of 8.62 grains/10−2 mm2; however, most of the parenchyma cells had no starch grains (Figure 6h). The interxylary phloem contained the largest amount of starch grains with approximately 9.21 grains/10−2 mm2, which were smaller in size but more densely distributed (Figure 6i). The xylem rays were more scattered and mainly distributed at the cell edges (Figure 6j), with approximately 9.09 grains/10−2 mm2. The axial parenchyma had the lowest grain concentration with approximately 7.56 grains/10−2 mm2. There were more starch grains in the 18 months of inoculation in the decay zone than in the 12 months of inoculation, and both had a similar distribution pattern in different zones. In the decay zone, very small amounts of starch grains could still be observed, almost entirely in the xylem rays (Figure 6k) and in the axial parenchyma (Figure 6l), with an average number of 6.80 grains/10−2 mm2 and 7.98 grains/10−2 mm2, respectively, and not found in the intrinsic siliques.
The soluble sugars staining in different compartments of the xylem is shown in Figure 8. The statistics are shown in Figure 9.
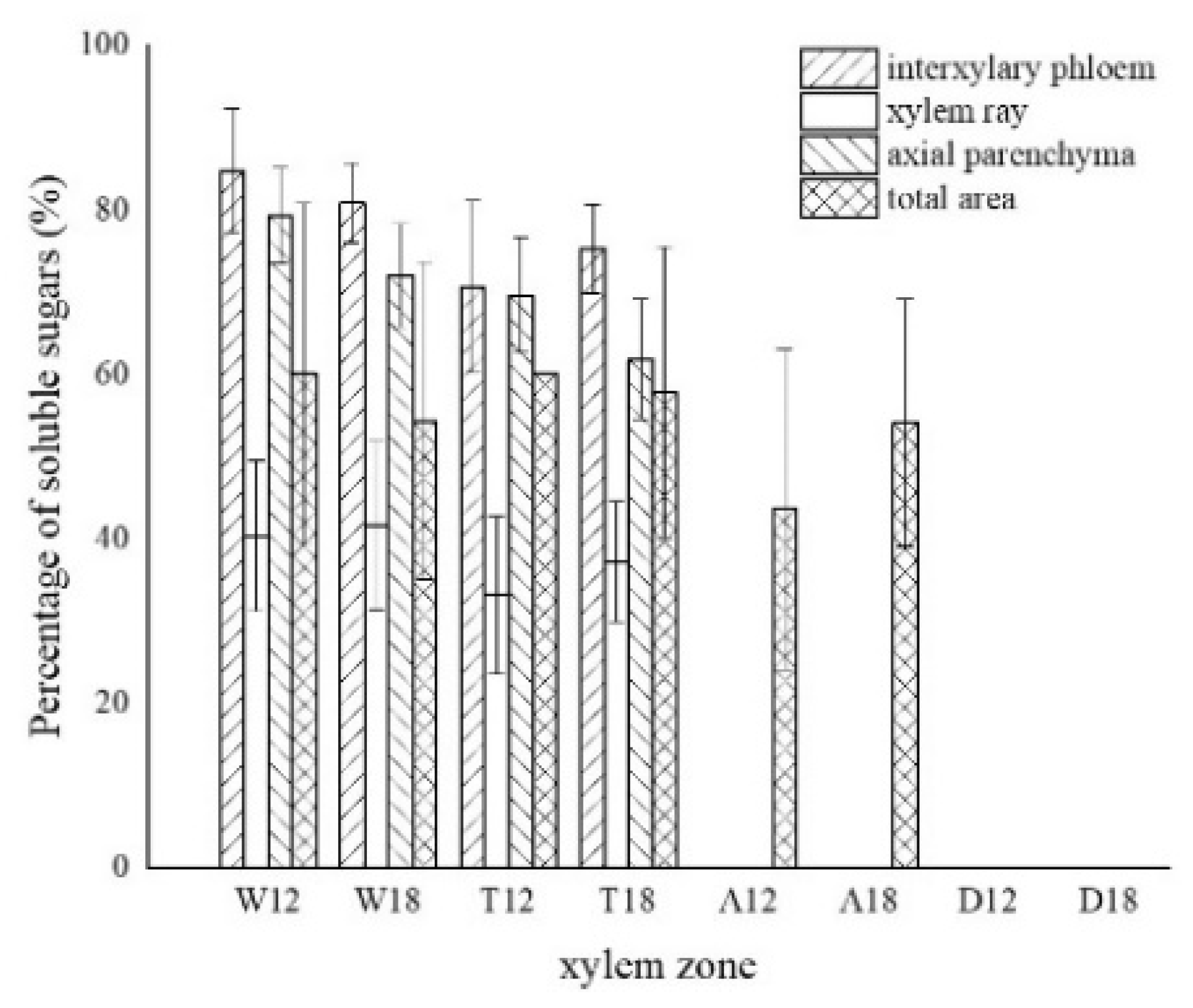
The distribution of soluble sugars in the blank control is shown in Figure 8a. The soluble sugars in the white zone were mainly distributed in the axial parenchyma and the interxylary phloem and less in the xylem rays. It was similar to the distribution in the blank control. Approximately 82.82% of the interxylary phloem was purplish-red in the transverse surface (Figure 8b), and approximately 75.68% of the axial parenchyma was stained purplish-red in the radial surface (Figure 8c). The xylem rays were only purplish-red near the interxylary phloem, with a color development rate of approximately 40.99%. In the transition zone, the colored area of the interxylary phloem and axial parenchyma was larger, whereas a small portion of the xylem rays was purplish-red and lighter in color. The purplish-red interxylary phloem was observed on the transverse surface (Figure 8d) with a colored area of approximately 72.98%. The purplish-red axial parenchyma could be observed in the radial surface (Figure 8e) with a colored area of approximately 65.75%. The colored area in the xylem rays was less than 35.16%. Some cells in the agarwood zone were purplish-red, as shown in Figure 8f,g. No noticeable pattern was observed in the distribution of soluble sugars in this zone. They were present in various cells and not restricted to parenchyma cells. The colored area was approximately 48.85% of the total area. No discoloration of parenchyma cells was observed in the decay areas (Figure 8h). The soluble sugar content of the 18 months of inoculation in A. sinensis was less than that of the 12 months of inoculation.
An insignificant amount of starch grains was observed in the decay zone, which is inconsistent with the results of previous studies [16]. Starch grains were typically located in the outer xylem rays of the decay zone. A likely reason for this is that the companion cells with metabolic functions and the sieve tubes with transport functions in interxylary phloem were blocked by agarwood resin and subsequently died due to insufficient supply of nutrients [21], while the parenchyma cells lost their ability to transport and break down sugar substances such as starch. This process accelerated when there were reduced amounts of unmetabolized starch grains in the parenchyma cells, which could not be metabolized to soluble sugars or transported and had to be temporarily retained in the resin-filled parenchyma cells. As the xylem decayed deeper, this area became the decay zone with the agarwood resin loss. These starch grains gradually decomposed into nutrients for fungi. Thus, the center of the decay zone contained no starch grains. The SEM results for the parenchyma cells of the decay zone supported this hypothesis. The cell walls of the parenchyma cells in the decay zone were covered with inclusions, which blocked the channels for intercellular material transport, such as the starch grain. A negligible amount of incompletely decomposed starch grains was observed in this zone.
3.4. Distribution of Sesquiterpenoids in the Xylem of A. sinensis
The color development of each zone is shown in Figure 10. From the top left to the bottom right are the white zone, transition zone, agarwood zone, and decay zone in Figure 10a. The statistics of color development are shown in Figure 11.
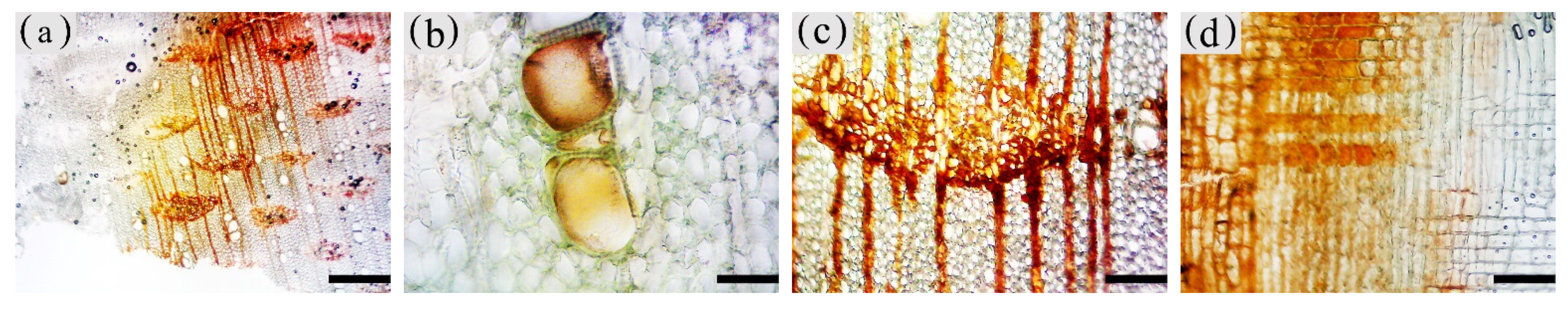
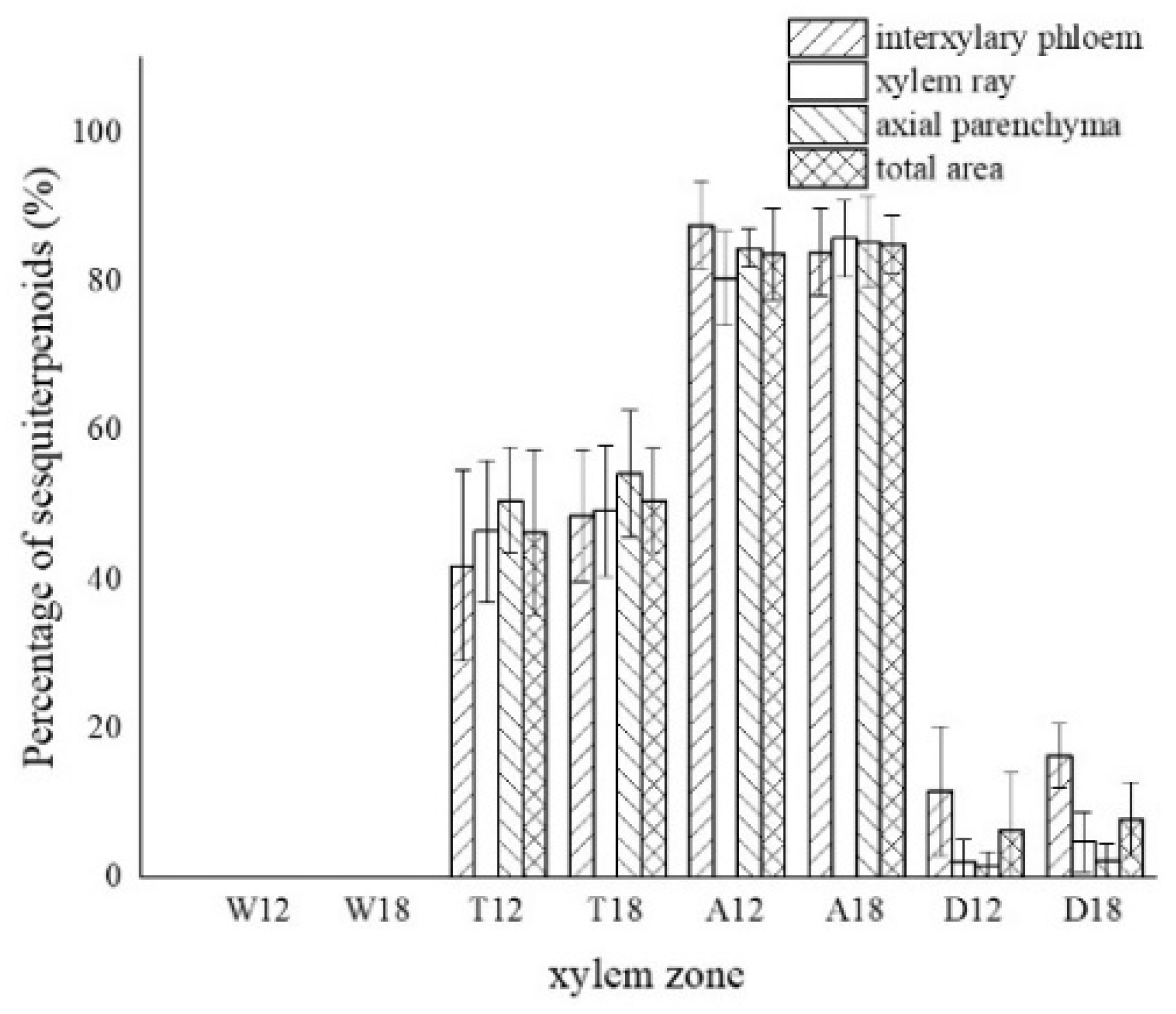
The blank control had no coloration, which was similar to the white zone. No coloration of parenchyma cells was observed in the white zone (Figure 10a). However, some of the vessels were reddish-brown (Figure 10b). In the transition zone, it was observed that some parenchyma cells were stained red, with the largest area of 52.46% for the axial parenchyma, followed by 47.87% for the xylem rays, and 48.50% the interxylary phloem. The total colored area of parenchyma cells was 46.36%. As shown in Figure 10c, the xylem rays and interxylary phloem in the transverse surface were yellowish-brown on the outside and reddish-brown on the inside adjacent to the agarwood zone. This indicated a gradual increase in sesquiterpenes from outside to inside (Figure 10d). As shown in Figure 10e,f, the interxylary phloem, xylem rays, and axial parenchyma cells in the agarwood zone were almost red. The interxylary phloem showed the largest color area of 85.76%, followed by the axial parenchyma. The xylem rays showed the least color with 83.14%, and the total color area of parenchyma cells with 84.61%. In the decayed zones, some parenchyma cells were red. Most of the cells within the decayed area showed no color (Figure 10g), and only some of the incompletely decayed interxylary phloem were red (Figure 10g). The interxylary phloem showed the largest color area (8.96%), followed by the xylem rays (3.43%), and the axial parenchyma (2.09%). The total color area of parenchyma cells was 6.42%. In contrast, the sesquiterpene content of 18 months of inoculation in A. sinensis was higher than that in 12 months of inoculation.
3.5. Production of Inclusions in the Xylem of A. sinensis Parenchyma Cell
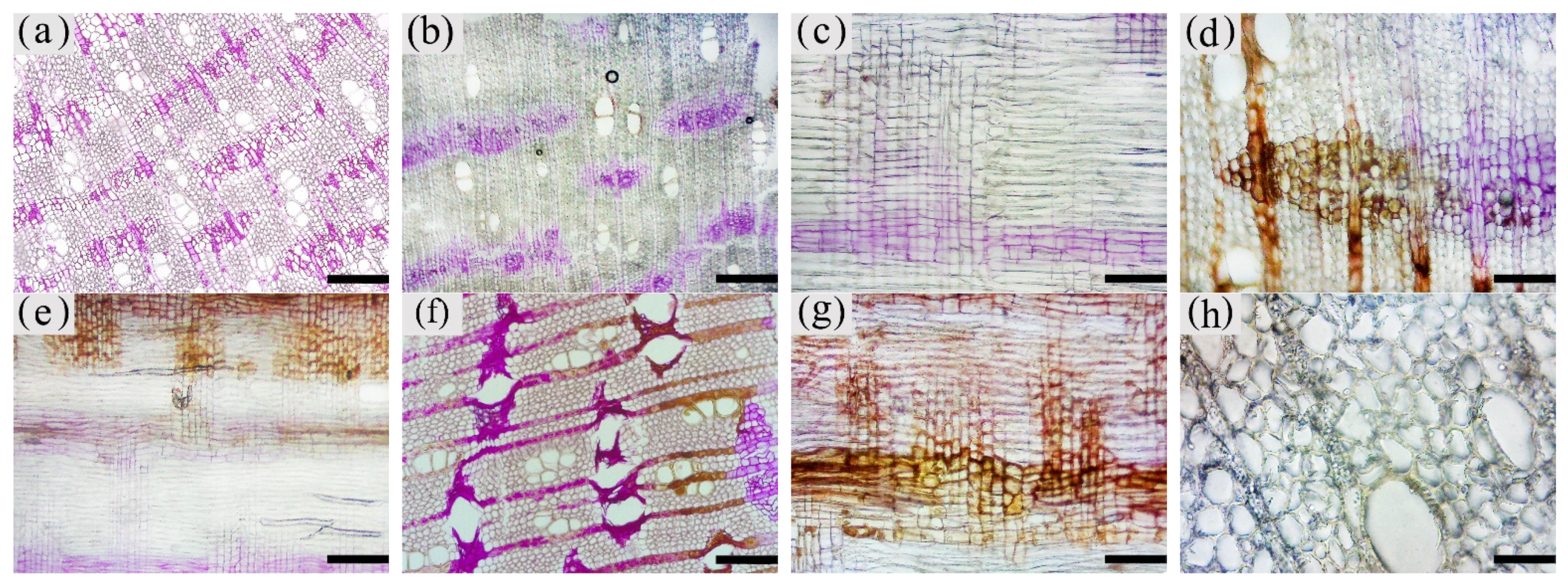
The inclusions in the xylem cells of A. sinensis were divided into two categories: invaginations and gums. Inclusions were often absent in the various cell types of the blank control and the white zone. However, granular starch grains were observed in the parenchyma cells (Figure 12a), and smooth cell walls and clear pores were observed (Figure 12b). In the transition zone, inclusions appeared in the parenchyma cells. The inclusions filled the cells and entered the adjacent cells through the grain (Figure 12c). Precipitates were visible on the cell walls, globular invaginations appeared in the cells (Figure 10e and Figure 12f), and gums that adhered to each other in the shape of spherical droplets began to appear. These greasy substances existed in some xylem rays and vessels (Figure 12d). There was a higher accumulation of inclusions in the decay zone and in the interxylary phloem, than in the xylem rays and axial parenchyma, which made them darker. The parts of the interxylary phloem that were connected to the xylem rays were darker, and the remainder of the interxylary phloem was lighter (Figure 12j). There was also an accumulation of inclusions in some of the fibers (Figure 12k). SEM showed that the starch grains in the parenchyma cells were almost completely dissolved and that the interxylary phloem contained a large accumulation of inclusions (Figure 12g,h). The xylem rays and axial parenchyma in the decay zone were full of inclusions (Figure 12i). The grains were not visible, some of the interxylary phloem was decayed, inclusions were present only in the living cells, and no inclusions remained in the vessels (Figure 12l).
3.6. Distribution and Morphology of Fungal Hyphae and Propagules in the Xylem of A. sinensis
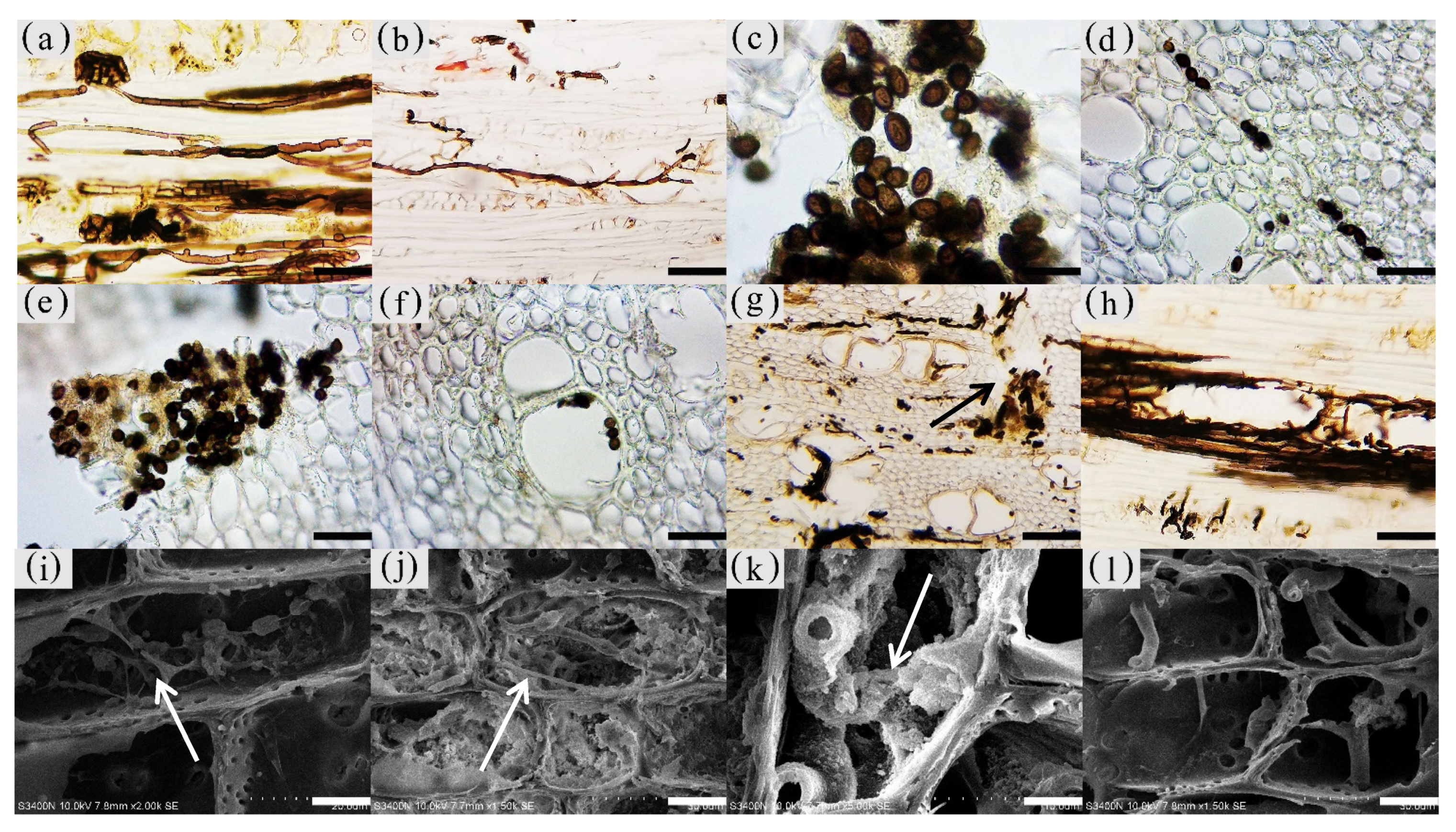
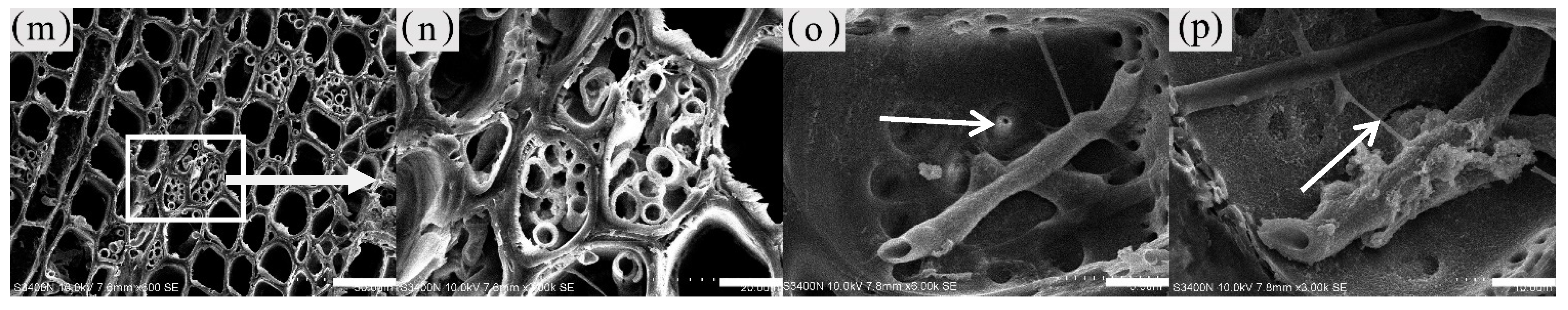
The fungal hyphae in the xylem of A. sinensis were branched. The main hyphae were 3–4 μm wide, and the branched hyphae were approximately 1.5 μm wide. Both had a smooth surface (Figure 13a) and spread axially (Figure 13b) [27]. The propagules were dark brown and ellipsoidal, with a long axis of 3–4 μm and a short axis of 1–3 μm (Figure 13c).
No fungal propagules or mycelia were observed in the blank control and the white zone. Very few branched mycelia were present in the wood rays of the transitional zone (Figure 13i). An insignificant number of fungal propagules or mycelia were distributed in parenchyma cells such as xylem rays (Figure 13j) in the agarwood zone. However, a considerable number of inclusions were attached to the surface of the mycelium (Figure 13k). In the decay zone, a considerable number of oval fungal propagules were observed in the axial parenchyma, xylem rays (Figure 13d), interxylary phloem (Figure 13e), and vessels (Figure 13f). The mycelia of the decay zone were widely distributed in all tissues. Mycelia were present in xylem rays (Figure 13l), axial parenchyma, interxylary phloem, and fibers (Figure 13m,n). The interior of the interxylary phloem in the decay zone contained a significant amount of fungi, and there was no longer an intact cell form (as in Figure 13g). The vessels were filled with mycelia (Figure 13h), and the branched hyphae expanded into adjacent cells through the grains in the cell wall (Figure 13o arrow). Mycelial fusion occurred at the point of contact between the branching mycelium and the main mycelium (as shown in Figure 13p arrow), forming a mycelial network in the xylem [28]. There was no significant difference in the distribution of 12 and 18 months of inoculation in A. sinensis.
The degree of fungal accumulation varied widely among zones and there was a decreasing trend from the inside to the outside [29]. This phenomenon may be attributed to two factors. First, the agarwood resin blocked vessels and various types of parenchyma cells, preventing the outward expansion of hyphae, also called the physical defense of the tree. Second, sesquiterpenes and other characteristic products of agarwood have antibacterial properties and can inhibit the growth of fungi, which is also known as the chemical defense of the tree [30,31].
3.7. Variance Analysis of Factors and Xylem Structure
The homogeneity test was performed on each group of data and the results show that all the homogeneity of the variance was satisfied. One-way ANOVA was then used for significance analysis.
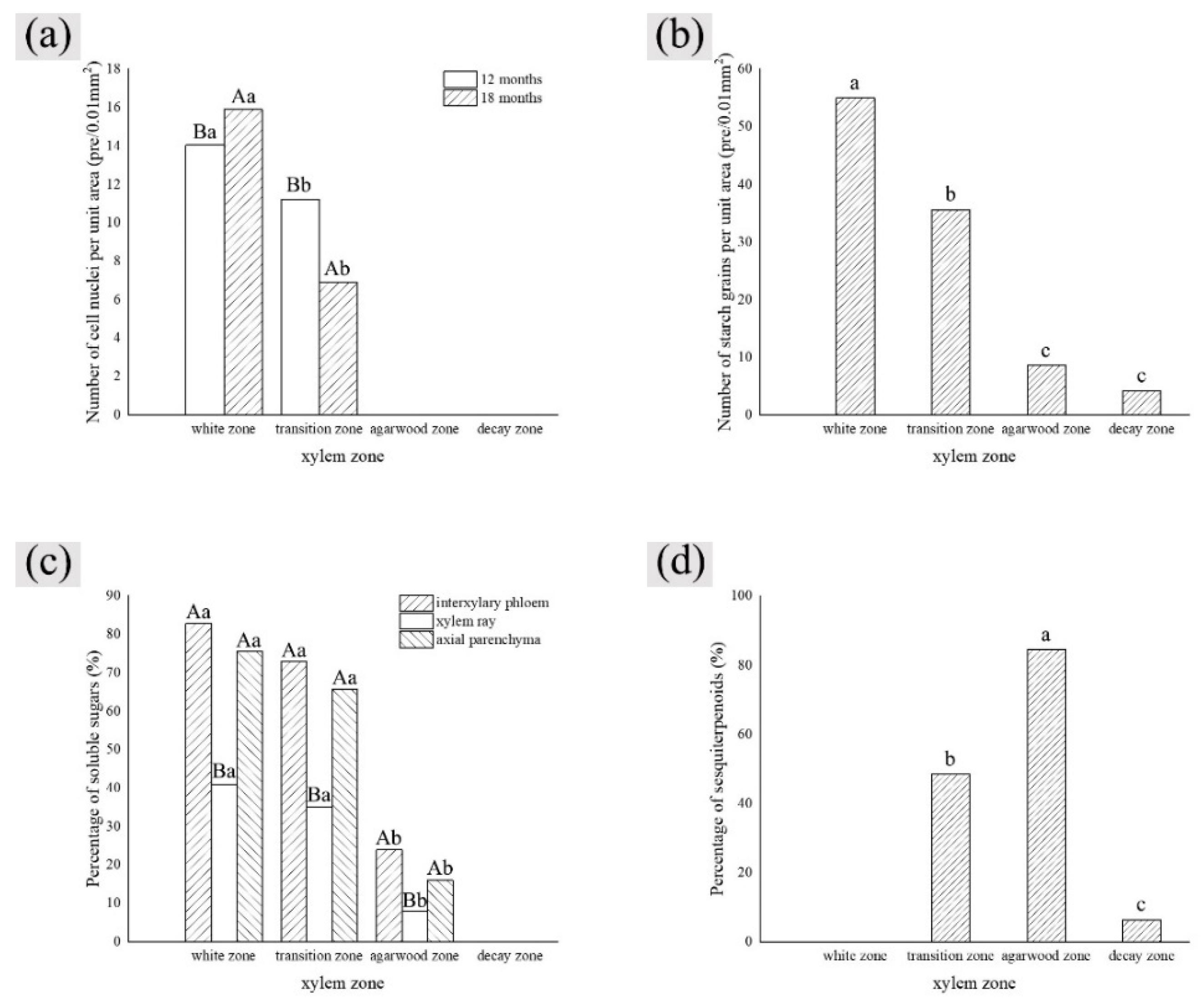
The results of one-way ANOVA of xylem zone, parenchyma cells type and inoculation time with nuclei, starch grains, area of soluble sugar, and sesquiterpene color development in xylem are shown in Table 1. The results show that the xylem zone had an extremely significant variability (p < 0.01) on all of the above parameters. Parenchyma cell type had an extremely significant variability (p < 0.01) on the soluble sugar color area, but not on the number of nuclei, starch grains, nor sesquiterpene color area. Inoculation time had an extremely significant variability (p < 0.01) on the number of nuclei, starch grains, and sesquiterpene color area. The number of nuclei was of significant variability (p < 0.05) by inoculation time, but not on the number of starch grains, area of soluble sugar, nor area of sesquiterpene color development. The results show that xylem zone had the greatest effect on the change of xylem structure among the three factors.
LSD comparisons were performed for factors that showed significant differences, while the average value and differences between each group are shown in Figure 14. The results show that the number of nuclei was influenced by two factors, and in terms of xylem zone, it was statistically more significantly different except for the agarwood and decay zone, which were not significant with respect to each other. In terms of inoculation time, 18 months of inoculation was statistically more significant, higher than 12 months of inoculation. The number of starch grains was affected by xylem zone, with the white zone being statistically more significant and higher than the other zones, the transition zone being statistically more significant and higher than the agarwood and decay zones, and not significant between the agarwood and decay zones. Soluble sugars were only affected by the xylem zone, and there was no significant difference between the white zone and transition zone, while they were statistically more significant and higher than the other zones. In terms of parenchyma cells type, the interxylary phloem and axial parenchyma were statistically more significant and higher than the xylem rays. The sesquiterpenes were influenced by the xylem zone, with statistically more significant relationships between all zones.
4. Discussion
4.1. Structural Changes in the Xylem of A. sinensis during Agarwood Formation
Based on the anatomical structure and histochemical analysis on the xylem of A. sinensis, it was found that after injection with the fungal inducers, agarwood was first generated in the pith of the tree. It combined with the wood of the xylem to form the agarwood zone, which spread outward with the agarwood development process. The outermost part of the agarwood zone was adjacent to the transition zone, where the physiological reactions of the agarwood process were concentrated. The white zone had not started to produce agarwood. As agarwood formation accelerated, the conducting tissue in the central part of the trunk was completely blocked by agarwood resin, resulting in the death of all types of cells in the area due to lack of nutrients and water [32]. The presence of more fungi in this area accelerated cell death and nutrient depletion. The tree gradually rotted and formed a decay zone.
By exploring the role of various tissues in the formation of agarwood, it was observed that agarwood resin was first produced in the interxylary phloem, then spread outward through the xylem rays and production continued in the outer interxylary phloem. As the accumulation of agarwood resin increased it appeared in the vessels and wood fibers adjacent to the parenchyma cells. The resin attached to the vessel walls and diffused through the paired pits to the adjacent vessels. Resin in the vessels diffused axially to the upper and lower ends.
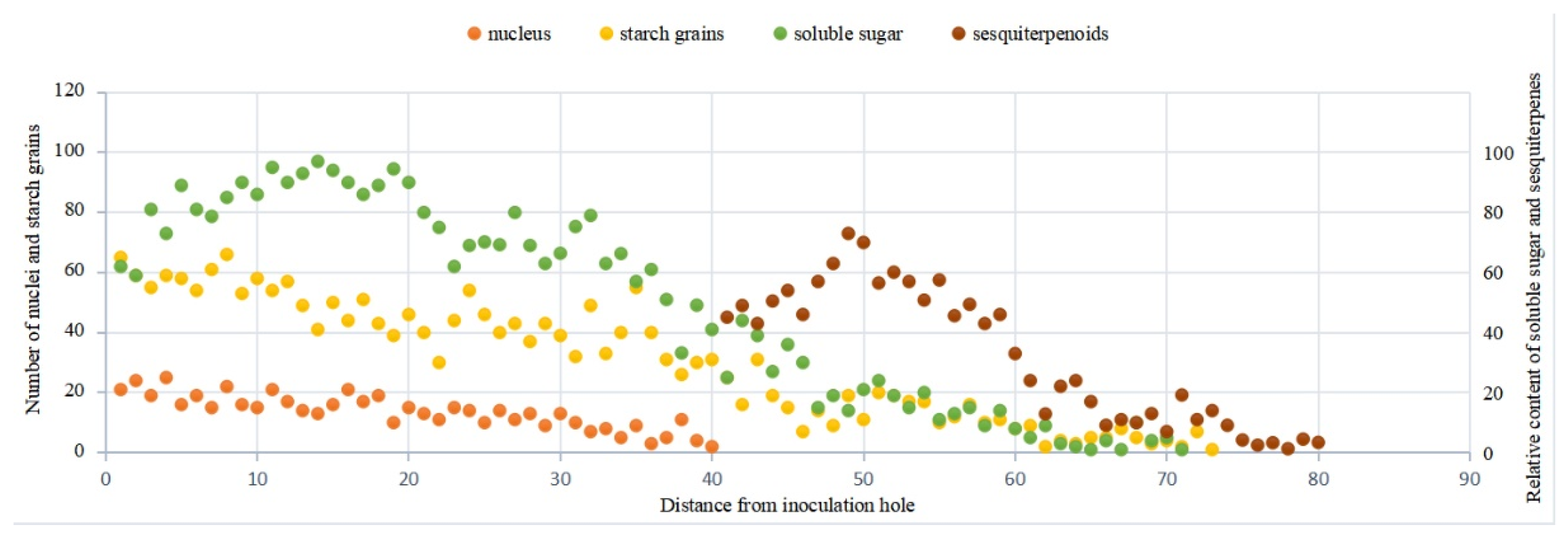
Serial sections were made on the transverse section of the xylem of agarwood-bearing A. sinensis, and the changes of various indexes were counted (Figure 15). It was discovered that the starch content decreased sharply whereas the soluble sugar content increased and then decreased, accompanied by the generation of sesquiterpenoids. The content of secondary metabolites, such as sesquiterpenes, increased gradually with the expansion of the agarwood. In this process, a transformation from starch grains to soluble sugars occurred in parenchyma cells, which also implied a transformation of storage nutrients to nutrients that can participate in physiological reactions [22]. The reduction in starch, variation in soluble sugar content and presence of sesquiterpenoids suggest that the parenchyma cells enhanced physiological functions spontaneously after external stress, thereby converting starch, soluble sugar, and other stores into large amounts of extracts [33]. These extracts combined with the xylem of A. sinensis to form agarwood, and the results were the same as those reported by Xingli Zhang [34].
4.2. Relationship between Fungi and Inclusions Production
Fungal induction caused the production of inclusions such as agarwood resin in A. sinensis [11,35]. The distribution of hyphae, inclusions, and sesquiterpenes in various types of cells was compared. It was found that the fungi entered the tree and moved axially along the vessels and radially along the xylem rays to accumulate and induce agarwood production in the interxylary phloem and xylem rays. Infestation of A. sinensis by fungi induced a local necrotic response in near tissues, which promotes the accumulation of callose in response to plant adversity stress. The deposition of callose can alter the transport function of the sieve duct and also change the osmotic potential of cells and solutions, resulting in the change of water content in the cells, creating an environment where fungal growth is inhibited [36].
SEM of the areas where hyphae and inclusions existed found that the decay zone contained significant amounts of hypha but insignificant amounts of inclusions. This showed that inclusions and fungal hyphae did not coexist. Therefore, the agarwood resin was not the direct product of fungal growth and reproduction in the xylem of A. sinensis but may be the product of a defense mechanism activated by the tree as a result of fungal induction [33]. Some studies have shown that the occurrence of terpenoids, such as sesquiterpenes, monoterpenes, and diterpenes was closely related to fungal diversity, which was a primary determinant of agarwood properties [37].
Some of the dominant species in the fungal inducers we used belonged to endophytic fungi, and experiments showed that unprocessed A. sinensis also had the presence of these fungi [38]. A large number of forestry practices have shown that unprocessed A. sinensis cannot produce agarwood, but inoculation with endophytic fungi enables it to produce agarwood. That means that the mere presence of the original endophytic fungi in the tree without inoculation does not lead to agarwood formation. In a previous study, we found that drilling without the injection of the fungal also did not lead to agarwood formation. Thus, we believe that, in this experiment, the synergistic effect of fungi and drilling is the key to producing agarwood.
The fungal inducers used in this experiment came from wild A. sinensis, which had a high yield of agarwood. These fungal species have been shown in experiments to have better agarwood production effects. In the following experiments, these fungi will be isolated and inoculated, Whereafter, experiments on single fungal specie and mixed fungal species will be conducted, expecting to research fungal species or combinations for better agarwood formation.
4.3. Structural Differences of Different Induction Methods
In terms of macrostructure, the agarwood obtained by the physical trauma is concentrated near the wound, and the agarwood formation area is limited and shaped into thin layers or bundles. The chemical induction can obtain a larger area of agarwood, instead of concentrating only around the injection hole, while the agarwood area is blocky. To some extent, the area of agarwood formation reflected the degree of accumulation of secondary metabolites in the process of exogenous substances-induced agarwood formation and was directly related to the quality of agarwood formation and volatile oil content [39]. In terms of microstructure, the agarwood obtained by the physical trauma is able to fill the entire cell cavity, while the agarwood resin obtained by the chemical induction is usually attached to the cell wall and does not fill the entire cell cavity [20,22].
The fungal induction has obvious advantages over the physical and chemical induction in that the agarwood is not only formed around the injection hole but almost around the whole tree. In addition, the agarwood can fill the entire cell, thus obtaining a large continuous area of agarwood. In agarwood production, these advantages are even more evident. Larger areas of agarwood allow for higher yields, and more continuous agarwood allows for more complete and pure agarwood.
Compared with chemical induction, fungal induction has a higher success rate in producing agarwood. The fungi used in the experiment were mostly endophytes of A. sinensis that did not destroy the original microbial community structure in the tree. Therefore, almost no A. sinensis died during the agarwood formation. In addition, A. sinensis inducted by fungal had no chemical residue after harvesting, which makes it safer when used in medicine and everyday products.
4.4. Differences in Xylem Structure at Different Inoculation Times
By ANOVA, we found that the time of inoculation was only extremely significantly different in terms of the number of nuclei. This extremely significant difference may be explained by the fact that secondary metabolites acted to resist fungal infestation from 12 to 18 months, which allowed the tree to recover some vitality. In the previous study, we found that there were fewer fungal abundances in the xylem of A. sinensis inoculated for 18 months than inoculated for 12 months, also confirming the suspicion. The time of inoculation did not affect the variation tendency of sesquiterpene content, which means there was no difference in the active ingredients of the agarwood produced by 12 and 18 months of inoculation with A. sinensis.
In our previous study, we found that the alcohol-soluble extract of the samples inoculated at 12 months was higher. The total relative content of sesquiterpenes and chromones was the same as at 18 months of treatment [25]. It means that the quality of agarwood was higher at 12 months after inoculation compared to 18 months. In terms of time cost, harvesting A. sinensis at 12 months after inoculation could yield higher economic value and higher production efficiency. Therefore, we believe that in production practice, A. sinensis artificially inducted by fungal inducers have the best agarwood formation effect at 12 months after inoculation.
5. Conclusions
- The interxylary phloem was significantly related to cell extinction, sugar conversion, and generation of characteristic agarwood products during agarwood formation. Among all types of parenchyma cells, the interxylary phloem had the fastest response to the agar-wood process and was the first region of agarwood generation;
- Compared with the 12 months of inoculation in A. sinensis, the xylem of the 18 months of inoculation was more vigorous. There were no significant differences between the 12 and 18 months of inoculation in terms of sugars and agarwood characteristic products. In production, harvesting after 12 months of inoculation can improve harvesting efficiency;
- The fungus entered the A. sinensis, expanded outward along the vessels and xylem rays, aggregated in the interxylary phloem and xylem rays, and induced the production of agarwood. In agarwood-bearing trees, fungal propagules and hyphae were mainly distributed in the decay zone.
Author Contributions
Conceptualization, Y.F. and Y.L. (Yuxin Liu); methodology, Y.F.; software, Y.L. (Yuxin Liu); validation, Y.F., Y.L. (Yuxin Liu) and P.W.; formal analysis, Z.L.; investigation, M.Q.; resources, Y.L. (Yingjian Li); data curation, P.W., Y.L. (Yingjian Li) and M.Q.; writing—original draft preparation, Y.L. (Yuxin Liu); writing—review and editing, Y.F. and Z.L.; visualization, Y.L. (Yuxin Liu); supervision, P.W.; project administration, Y.F.; funding acquisition, Y.F. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by the National Natural Science Foundation of China, grant number 32171702.
Data Availability Statement
Data are contained within the article.
Acknowledgments
We thank Guangxi Yuexiangtian Biotechnology Co., LTD for providing experimental samples and the agarwood-inducing technique.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Wu, Z.; Lu, A.; Tang, Y.; Chen, Z.; Li, D. The Families and Genera of Angiosperms in China—A Comprehensive Analysis; China Science Publishing & Media Ltd.: Beijing, China, 2003. [Google Scholar]
- Li, L.; Shou, H.; Ma, Q. Distribution of Aquilaria sinensis (Thymelaeaceae). J. Anhui Agri. Sci. 2012, 40, 9254–9256. [Google Scholar]
- National Key Protection of Wild Plants List (the First Batch); Communique of the State Council of the People’s Republic of China: Beijing, China, 1999; pp. 39–47.
- Convention on International Trade in Endangered Species of Wild Fauna and Flora. J. Int. Wildl. Law AMP Policy 2005, 8, 229.
- Chen, Y.; Liu, H.; Heinen, J. Challenges in the Conservation of an Over-Harvested Plant Species with High Socioeconomic Values. Sustainability 2019, 11, 4194. [Google Scholar] [CrossRef] [Green Version]
- Liu, Y.; Chen, H.; Yang, Y.; Zhang, Z.; Wei, J.; Meng, H.; Chen, W.; Feng, J.; Gan, B.; Chen, X.; et al. Whole-tree Agarwood-Inducing Technique: An Efficient Novel Technique for Producing High-Quality Agarwood in Cultivated Aquilaria sinensis Trees. Molecules 2013, 18, 3086–3106. [Google Scholar] [CrossRef]
- Wu, Z. Studies on the Agarwood Forming Mechanism and Quality Evaluation of Aquilaria sinensis by Biologically Induced Method; Guangzhou University of Chinese Medicine: Guangzhou, China, 2015. [Google Scholar]
- Ei, W.; Zuo, W.; Yang, D.; Dong, W.; Dai, H. Advances in the Mechanism, Artificial Agarwood-induction Techniques and Chemical Constituents of Artificial Agarwood Production. Chin. J. Trop. Crops 2013, 34, 2513–2520. [Google Scholar]
- Liu, Y. Research on Quality Characteristics of Agarwood Produced by Whole-Tree Agarwood-Inducing Technique; Peking Union Medical College: Beijing, China, 2018. [Google Scholar]
- Yun, Y.; Feng, J.; Zhu, J.; Liu, Y.; Chen, D.; Wei, J. Quality of Agarwood Produced by Whole Tree Agarwood-inducing Technique (Agar-Wit) from China and Southeast Asian Countries. Chin. Pharm. J. 2019, 54, 1988–1994. [Google Scholar]
- Chen, X.; Sui, C.; Liu, Y.; Yang, Y.; Liu, P.; Zhang, Z.; Wei, J. Agarwood Formation Induced by Fermentation Liquid of Lasiodiplodia theobromae, the Dominating Fungus in Wounded Wood of Aquilaria sinensis. Curr. Microbiol. 2017, 74, 460–468. [Google Scholar] [CrossRef]
- Chen, X.; Liu, Y.; Yang, Y.; Yang, Y.; Liu, P.; Zhang, Z.; Wei, J. Trunk surface agarwood-inducing technique with Rigidoporus vinctus: An efficient novel method for agarwood production. PLoS ONE 2018, 13, e198111. [Google Scholar] [CrossRef] [PubMed]
- Mohamed, R.; Jong, P.L.; Kamziah, A.K. Fungal inoculation induces agarwood in young Aquilaria malaccensis trees in the nursery. J. For. Res. 2014, 25, 201–204. [Google Scholar] [CrossRef]
- Faizal, A.; Esyanti, R.R.; Aulianisa, E.N.; Santoso, E.; Turjaman, M. Formation of agarwood from Aquilaria malaccensis in response to inoculation of local strains of Fusarium solani. Trees 2017, 31, 189–197. [Google Scholar] [CrossRef]
- Zhang, Z.; Han, X.; Wei, J.; Xue, J.; Yang, Y.; Liang, L.; Li, X.; Guo, Q.; Xu, Y.; Gao, Z. Compositions and Antifungal Activities of Essential Oils from Agarwood of Aquilaria sinensis (Lour.) Gilg Induced by Lasiodiplodia theobromae (Pat.) Griffon. & Maubl. J. Braz. Chem. Soc. 2014, 25, 20–26. [Google Scholar] [CrossRef]
- Liu, P.; Zhang, X.; Yang, Y.; Xue, J.; Yang, Y.; Liang, L.; Li, X.; Guo, Q.; Xu, Y.; Gao, Z. Interxylary phloem and xylem rays are the structural foundation of agarwood resin formation in the stems of Aquilaria sinensis. Trees 2019, 33, 533–542. [Google Scholar] [CrossRef]
- Tabata, Y.; Widjaja, E.; Mulyaningsih, T.; Parman, I.; Wiriadinata, H.; Mandang, Y.I.; Itoh, T. Structural survey and artificial induction of aloeswood. Wood Res. Bull. Wood Res. Inst. Kyoto Univ. 2003, 90, 11–90. [Google Scholar]
- Fahn, A. The Development of the Secondary Body in Plants with Interxylary Phloem. In Xylorama; Springer: Birkhäuser, Basel, 1985. [Google Scholar]
- Carlquist, S. Interxylary phloem: Diversity and functions. Brittonia 2013, 65, 477–495. [Google Scholar] [CrossRef]
- He, H.; Pan, Z.; Liang, Y.; Luo, P.; Qiu, J. Anatomy comparative study of Aquilaria sinensis in different agarwood induced methods. J. For. Eng. 2019, 4, 54–59. [Google Scholar]
- Li, M.; Shen, H.; He, H.; Qiu, J. Anatomic Structure and Extracted Chemical Component Content of Aquilaria sinensis. J. Southwest For. Univ. 2017, 37, 208–213. [Google Scholar]
- Wang, J.; Wang, Y.; Jinling, Y.; Wei, L.I.; Dong, W.; Wenli, M.; Haofu, D. Comparison of the Anatomy Structure and Chemical Compositions of Agarwoods from Two Kinds of Aquilaria sinensis. SCIENTIA SILVAE SINICAE 2019, 55, 146–154. [Google Scholar]
- Keisuke, O.; Hideo, K.; Hiroo, F. Direct Evidence of Active and Rapid Nuclear Degradation Triggered by Vacuole Rupture during Programmed Cell Death in Zinnia. Plant Physiol. 2001, 125, 615–626. [Google Scholar]
- Janssen, T.A.J.; Hölttä, T.; Fleischer, K.; Naudts, K.; Dolman, H. Wood allocation trade-offs between fiber wall, fiber lumen, and axial parenchyma drive drought resistance in neotropical trees. Plant Cell Environ. 2020, 43, 965–980. [Google Scholar] [CrossRef] [Green Version]
- Sheng, M.; Yunlin, F.; Yingjian, L.; Penglian, W.; Zhigao, L. The formation and quality evaluation of agarwood induced by the fungi in Aquilaria sinensis. Ind. Crops Prod. 2021, 173, 114129. [Google Scholar]
- Ma, S.; Qiao, M.; Fu, Y.; Wei, P.; Li, Y.; Liu, Z. Comparative Analysis of Biological Activity of Artificial and Wild Agarwood. Forests 2021, 12, 1532. [Google Scholar] [CrossRef]
- Kang, Z. Ultrastructure of Plant Pathogenic Fungi; China Science & Technology Press: Beijing, China, 1995. [Google Scholar]
- Sbrana, C.; Fortuna, P.; Giovannetti, M. Plugging into the network: Belowground connections between germlings and extraradical mycelium of arbuscular mycorrhizal fungi. Mycologia 2011, 103, 307–316. [Google Scholar] [CrossRef] [PubMed]
- Xiang, Z.; Jia-Qi, G.; Yuan, Y.; Quan, Y.; Liang-Yun, Z.; Juan, L.; Lu-Qi, H. Analysis of bacteria distribution characteristics in different layers of agarwood based on Hiseq sequencing. China J. Chin. Mater. Med. 2020, 45, 2374–2381. [Google Scholar]
- Kuo, T.; Huang, H.; Hsu, C. Mass spectrometry imaging guided molecular networking to expedite discovery and structural analysis of agarwood natural products. Anal. Chim. Acta 2019, 1080, 95–103. [Google Scholar] [CrossRef]
- Zhidong, L.; Shuang, Z.; Dailin, L.; Xiaoxia, G.; Yanmin, Z.; Yu, C. Evaluation of three different artificial agarwood-inducing methods from Aquilaria sinensis using antimicrobial activity. Pak. J. Pharm. Sci. 2019, 32, 905–910. [Google Scholar]
- Nussinovitch, A. Plant Gum Exudates of the World:Sources, Distribution, Properties, and Applications; CRC Press: Boca Raton, FL, USA, 2009. [Google Scholar]
- Latt, C.R.; Nair, P.K.R.; Kang, B.T. Reserve carbohydrate levels in the boles and structural roots of five multipurpose tree species in a seasonally dry tropical climate. For. Ecol. Manag. 2001, 146, 145–158. [Google Scholar] [CrossRef]
- Zhang, X. Studies on Relationship between Wound-Induced Defense Response and Agarwood Formation in Aquilaria sinensis. Ph.D. Thesis, Beijing Forestry University, Beijing, China, 2013. [Google Scholar]
- Justin, S.; Lihan, S.; ElvisSulang, M.; Chiew, T. Formulated microbial consortium as inoculant for agarwood induction. J. Trop. For. Sci. 2020, 32, 161–169. [Google Scholar] [CrossRef]
- Gu, L.; Zheng, K.; Liu, Y.; Ma, H.; Xiao, Z. Two Kinds of Fungi Effect on the Tissue Structure of Aquilaria sinensis Xylem. J. West China For. Sci. 2018, 47, 141–144. [Google Scholar]
- Juan, L.; Xiang, Z.; Jian, Y.; Junhui, Z.; Yuan, Y.; Chao, J.; Xiulian, C.; Luqi, H. Agarwood wound locations provide insight into the association between fungal diversity and volatile compounds in Aquilaria sinensis. R. Soc. Open Sci. 2019, 6, 190211. [Google Scholar]
- Zhang, M. Apreliminary Study on the of Lsolation and Identification of Endophytic Bacteria in White Fragrant and Distribution. Master’s Thesis, Guangzhou University of Chinese Medicine, Guangzhou, China, 2017. [Google Scholar]
- Song, X.; Huang, G.; Wang, X.; Song, J.; Zhang, Q.; Liang, K.; Zhou, Z. Resistance Capability of Aquilaria sinensis under Combinations of Inorganic Salts, Hormones and Fungus Induction. Bull. Bot. Res. 2019, 39, 505–513. [Google Scholar]
Figure 1.
Distribution of A. sinensis and A. yunnanensis in China.

Figure 2.
Diagram of fungal inducer inoculation sites; (a) Location of injection holes for the whole tree; (b) Enlargement of the diagram with the spacing of the inoculation holes; (c) Schematic of transverse surface. The height of the tree on the schematic was shortened in a ratio of 1:4 in order to allow for a complete presentation of the location of the inducer injection holes.
Figure 2.
Diagram of fungal inducer inoculation sites; (a) Location of injection holes for the whole tree; (b) Enlargement of the diagram with the spacing of the inoculation holes; (c) Schematic of transverse surface. The height of the tree on the schematic was shortened in a ratio of 1:4 in order to allow for a complete presentation of the location of the inducer injection holes.

Figure 3.
Macrostructure of the xylem of A. sinensis. (a-1,a-2) Different zones of air-dried timber in transverse surface. (a-3) blank control without being inoculated. (b) Different zones in transverse surface. (c) Different zones in radial surface. (d) Transverse surface of white zone. (e) Radial surface of white zone. (f) Transverse surface of transition zone. (g) Radial surface of transition zone. (h) The dark part is the transverse surface of agarwood zone. (i) The dark part is the radial surface of agarwood zone. (j) Transverse surface of decay zone. (k) Radial surface of decay zone. V: vessel; IP: interxylary phloem; XR: xylem ray. Scale bars = (a) 100 mm; (b–k) 400 μm.
Figure 3.
Macrostructure of the xylem of A. sinensis. (a-1,a-2) Different zones of air-dried timber in transverse surface. (a-3) blank control without being inoculated. (b) Different zones in transverse surface. (c) Different zones in radial surface. (d) Transverse surface of white zone. (e) Radial surface of white zone. (f) Transverse surface of transition zone. (g) Radial surface of transition zone. (h) The dark part is the transverse surface of agarwood zone. (i) The dark part is the radial surface of agarwood zone. (j) Transverse surface of decay zone. (k) Radial surface of decay zone. V: vessel; IP: interxylary phloem; XR: xylem ray. Scale bars = (a) 100 mm; (b–k) 400 μm.

Figure 4.
Color development of the nuclei of parenchyma cells in the xylem of A. sinensis. (a) Nuclei in the blank control. (b) Nuclei in the medialis xylem rays of the white zone. (c) Nuclei in the xylem rays of the transition zone. (d) Nuclei in the interxylary phloem of the transition zone. (e) Medialis of transitional zone transverse surface without the nucleus. (f) No nuclei were observed in the agarwood zone. (g) No nuclei were observed in the decay zone. (h) Highly crumpled nuclei in the transition zone. Scale bars = (a–d) 50 μm; (e–g) 80 μm; (h) 25 μm.
Figure 4.
Color development of the nuclei of parenchyma cells in the xylem of A. sinensis. (a) Nuclei in the blank control. (b) Nuclei in the medialis xylem rays of the white zone. (c) Nuclei in the xylem rays of the transition zone. (d) Nuclei in the interxylary phloem of the transition zone. (e) Medialis of transitional zone transverse surface without the nucleus. (f) No nuclei were observed in the agarwood zone. (g) No nuclei were observed in the decay zone. (h) Highly crumpled nuclei in the transition zone. Scale bars = (a–d) 50 μm; (e–g) 80 μm; (h) 25 μm.

Figure 5.
Variation in the number of nuclei in a different zone of cells. W—white zone; T—transition zone; A—agarwood zone; D—decay zone; 12—inoculated for 12 months; 18—inoculated for 18 months.
Figure 5.
Variation in the number of nuclei in a different zone of cells. W—white zone; T—transition zone; A—agarwood zone; D—decay zone; 12—inoculated for 12 months; 18—inoculated for 18 months.

Figure 6.
Color development of starch grains in xylem parenchyma cells of A. sinensis. (a) Starch grains in the blank control. (b) Starch grains in axial parenchyma and xylem rays in radial surface of white zone. (c) Starch grains in the interxylary phloem in the transverse surface of the white zone. (d) Starch grains in the medical wood rays in the transverse surface of the white zone. (e) Starch grains in the interxylary phloem in the transverse surface of the transition zone. (f) Some of the xylem rays and axial parenchyma in the radial surface of the transition zone contain starch grains. (g) No starch grains in the medialis of the interxylary phloem in transition zone. (h) Most parenchyma cells without starch grains in the transverse surface of the agarwood zone. (i) Starch grains in some of the interxylary phloem in the transverse surface of the agarwood zone. (j) Starch grains in some of the interxylary phloem in the transverse surface of the agarwood zone. (k) Starch grains in some of the interxylary phloem in the transverse surface of the decay zone. (l) Starch grains in some of the interxylary phloem in the radial surface of the decay zone. Scale bars = (c,e–i,k,l) 50 μm; (b)200 μm; (a,d,j) 25 μm.
Figure 6.
Color development of starch grains in xylem parenchyma cells of A. sinensis. (a) Starch grains in the blank control. (b) Starch grains in axial parenchyma and xylem rays in radial surface of white zone. (c) Starch grains in the interxylary phloem in the transverse surface of the white zone. (d) Starch grains in the medical wood rays in the transverse surface of the white zone. (e) Starch grains in the interxylary phloem in the transverse surface of the transition zone. (f) Some of the xylem rays and axial parenchyma in the radial surface of the transition zone contain starch grains. (g) No starch grains in the medialis of the interxylary phloem in transition zone. (h) Most parenchyma cells without starch grains in the transverse surface of the agarwood zone. (i) Starch grains in some of the interxylary phloem in the transverse surface of the agarwood zone. (j) Starch grains in some of the interxylary phloem in the transverse surface of the agarwood zone. (k) Starch grains in some of the interxylary phloem in the transverse surface of the decay zone. (l) Starch grains in some of the interxylary phloem in the radial surface of the decay zone. Scale bars = (c,e–i,k,l) 50 μm; (b)200 μm; (a,d,j) 25 μm.

Figure 7.
Statistics of color development of starch grains in parenchyma cells. W—white zone; T—transition zone; A—agarwood zone; D—decay zone; 12—inoculated for 12 months; 18—inoculated for 18 months.
Figure 7.
Statistics of color development of starch grains in parenchyma cells. W—white zone; T—transition zone; A—agarwood zone; D—decay zone; 12—inoculated for 12 months; 18—inoculated for 18 months.

Figure 8.
Color development of soluble reducing sugars in xylem parenchyma cells of A. sinensis. (a) Transverse surface of blank control. (b) Transverse surface of white zone. (c) Radial surface of white zone. (d) Transverse surface of transition zone. (e) Radial surface of transition zone. (f) Transverse surface of agarwood zone. (g) Radial surface of agarwood zone. (h) Transverse surface of decay zone. Scale bars= (a,b,e,f) 200 μm; (c,d,g) 100 μm; (h) 50 μm.
Figure 8.
Color development of soluble reducing sugars in xylem parenchyma cells of A. sinensis. (a) Transverse surface of blank control. (b) Transverse surface of white zone. (c) Radial surface of white zone. (d) Transverse surface of transition zone. (e) Radial surface of transition zone. (f) Transverse surface of agarwood zone. (g) Radial surface of agarwood zone. (h) Transverse surface of decay zone. Scale bars= (a,b,e,f) 200 μm; (c,d,g) 100 μm; (h) 50 μm.

Figure 9.
Statistics of color development of soluble sugar in xylem parenchyma cells. W—white zone; T—transition zone; A—agarwood zone; D—decay zone; 12—inoculated for 12 months; 18—inoculated for 18 months.
Figure 9.
Statistics of color development of soluble sugar in xylem parenchyma cells. W—white zone; T—transition zone; A—agarwood zone; D—decay zone; 12—inoculated for 12 months; 18—inoculated for 18 months.

Figure 10.
Color development of sesquiterpenoids in parenchyma cells of xylem of A. sinensis. (a) Color development of each zone in the transverse surface. (b) Vessel containing inclusions in the transverse surface of white zone. (c) Transverse surface of transition zone. (d) Radial surface of transition zone. (e) Transverse surface of agarwood zone. (f) Radial surface of agarwood zone. (g) Transverse surface of decay zone. (h) Transverse surface of decay zone, the edges of the undecayed interxylary phloem at the arrows are showing color. Scale bars = (a,g) 500 μm; (b,e,f) 50 μm;(c,d,h) 200 μm.
Figure 10.
Color development of sesquiterpenoids in parenchyma cells of xylem of A. sinensis. (a) Color development of each zone in the transverse surface. (b) Vessel containing inclusions in the transverse surface of white zone. (c) Transverse surface of transition zone. (d) Radial surface of transition zone. (e) Transverse surface of agarwood zone. (f) Radial surface of agarwood zone. (g) Transverse surface of decay zone. (h) Transverse surface of decay zone, the edges of the undecayed interxylary phloem at the arrows are showing color. Scale bars = (a,g) 500 μm; (b,e,f) 50 μm;(c,d,h) 200 μm.


Figure 11.
Color development statistics of sesquiterpenes in parenchyma cells of xylem of A. sinensis. W—white zone; T—transition zone; A—agarwood zone; D—decay zone; 12—inoculated for 12 months; 18—inoculated for 18 months.
Figure 11.
Color development statistics of sesquiterpenes in parenchyma cells of xylem of A. sinensis. W—white zone; T—transition zone; A—agarwood zone; D—decay zone; 12—inoculated for 12 months; 18—inoculated for 18 months.

Figure 12.
Accumulation of inclusions in parenchyma cells of xylem of A. sinensis. (a) Radial surface of white zone, with starch grains in wood rays and no inclusions. (b) Grain of xylem rays in white zone, with no inclusions. (c) Grain of xylem rays in transition zone, with inclusions. (d) Transverse surface of transition zone, with the resin in the xylem rays and vessels. (e,f) Xylem rays of transition zone, (f) is a partial enlargement of (e), and the arrow in (e) shows the invaginations. (g) Interxylary phloem of transverse surface in the agarwood zone, with large amounts of inclusions. (h,i) Transverse surface of agarwood zone, (g) is a partial enlargement of (h). (i) Transverse surface of decay zone, with large amounts of inclusions in the xylem rays. (j) Interxylary phloem of transverse surface in the agarwood zone: arrow is darker here than in other areas. (k) Fibers containing inclusions in the transverse surface of the agarwood zone. (l) Partially decayed interxylary phloem in the transverse surface of the decay zone. Scale bars= (a) 15 μm; (b,c,f) 2 μm; (d) 40 μm; (e,h,i) 10 μm; (g) 35 μm; (j,l) 200 μm; (k) 50 μm.
Figure 12.
Accumulation of inclusions in parenchyma cells of xylem of A. sinensis. (a) Radial surface of white zone, with starch grains in wood rays and no inclusions. (b) Grain of xylem rays in white zone, with no inclusions. (c) Grain of xylem rays in transition zone, with inclusions. (d) Transverse surface of transition zone, with the resin in the xylem rays and vessels. (e,f) Xylem rays of transition zone, (f) is a partial enlargement of (e), and the arrow in (e) shows the invaginations. (g) Interxylary phloem of transverse surface in the agarwood zone, with large amounts of inclusions. (h,i) Transverse surface of agarwood zone, (g) is a partial enlargement of (h). (i) Transverse surface of decay zone, with large amounts of inclusions in the xylem rays. (j) Interxylary phloem of transverse surface in the agarwood zone: arrow is darker here than in other areas. (k) Fibers containing inclusions in the transverse surface of the agarwood zone. (l) Partially decayed interxylary phloem in the transverse surface of the decay zone. Scale bars= (a) 15 μm; (b,c,f) 2 μm; (d) 40 μm; (e,h,i) 10 μm; (g) 35 μm; (j,l) 200 μm; (k) 50 μm.

Figure 13.
Distribution of fungal propagule and hypha in parenchyma cells and vessel. (a) Mycelial morphology of the xylem. (b)Type of mycelial dispersal. (c) Fungal propagule morphology. (d) Fungal propagules in xylem rays of decay zone. (e) Fungal propagules in the interxylary phloem of decay zone. (f) Fungal propagules in the vessel of decay zone. (g) Transverse surface of decay zone, mycelium distributed in xylem rays and axial parenchyma, with the decayed interxylary phloem at the arrow. (h) Fungal hypha in vessel of radial surface which in decay zone. (i) Fungal hypha in xylem rays of radial surface which in transition zone, with mycelium at arrows. (j,k) Transverse surface of agarwood zone, inclusions and mycelium in xylem rays, with mycelium at arrows. (l) Fungal hypha in xylem rays of radial surface which in decay zone. (m,n) Fungal hypha in fiber of transverse surface which in decay zone, (n) is a partial enlargement of (m). (o) Branching hyphae infect adjacent cells through the pit. (p) Xylem rays of radial surface which in decay zone-the arrow shows the mycelium fusion between the branching mycelium and the main mycelium. Scale bars= (a,d–f,h) 50 μm; (b) 100 μm; (c,i,n) 10 μm; (g) 200 μm; (j,l) 15 μm; (k) 4 μm; (m) 35 μm; (o) 3.5 μm; (p) 7 μm.
Figure 13.
Distribution of fungal propagule and hypha in parenchyma cells and vessel. (a) Mycelial morphology of the xylem. (b)Type of mycelial dispersal. (c) Fungal propagule morphology. (d) Fungal propagules in xylem rays of decay zone. (e) Fungal propagules in the interxylary phloem of decay zone. (f) Fungal propagules in the vessel of decay zone. (g) Transverse surface of decay zone, mycelium distributed in xylem rays and axial parenchyma, with the decayed interxylary phloem at the arrow. (h) Fungal hypha in vessel of radial surface which in decay zone. (i) Fungal hypha in xylem rays of radial surface which in transition zone, with mycelium at arrows. (j,k) Transverse surface of agarwood zone, inclusions and mycelium in xylem rays, with mycelium at arrows. (l) Fungal hypha in xylem rays of radial surface which in decay zone. (m,n) Fungal hypha in fiber of transverse surface which in decay zone, (n) is a partial enlargement of (m). (o) Branching hyphae infect adjacent cells through the pit. (p) Xylem rays of radial surface which in decay zone-the arrow shows the mycelium fusion between the branching mycelium and the main mycelium. Scale bars= (a,d–f,h) 50 μm; (b) 100 μm; (c,i,n) 10 μm; (g) 200 μm; (j,l) 15 μm; (k) 4 μm; (m) 35 μm; (o) 3.5 μm; (p) 7 μm.


Figure 14.
Differences between each indicator and each factor. (a) Nucleus with xylem zone and inoculation time; (b) starch grains with xylem zone; (c) soluble sugar with xylem zone and tissue types; (d) sesquiterpenoids with xylem zone. Different lowercase letters indicate significant differences between zone; different capital letters in (a) indicate significant differences between inoculation times, and different capital letters in (c) indicate significant differences between tissue types.
Figure 14.
Differences between each indicator and each factor. (a) Nucleus with xylem zone and inoculation time; (b) starch grains with xylem zone; (c) soluble sugar with xylem zone and tissue types; (d) sesquiterpenoids with xylem zone. Different lowercase letters indicate significant differences between zone; different capital letters in (a) indicate significant differences between inoculation times, and different capital letters in (c) indicate significant differences between tissue types.

Figure 15.
Trends of various indicators in xylem of agarwood-bearing A. sinensis.


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
One-way ANOVA results (p) of xylem structure with xylem zone, parenchyma cell types, and inoculation times.
Table 1.
One-way ANOVA results (p) of xylem structure with xylem zone, parenchyma cell types, and inoculation times.
| Parameters | Xylem Zone | Parenchyma Cells Type | Inoculation Times |
|---|---|---|---|
| number of nuclei | 0.000 ** | 0.403 | 0.048 * |
| number of starch grains | 0.000 ** | 0.592 | 0.947 |
| soluble sugar color area | 0.000 ** | 0.005 ** | 0.933 |
| sesquiterpene color area | 0.000 ** | 0.429 | 0.385 |
Note: * significant difference (p < 0.05), ** extremely significant differences (p < 0.01).
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Liu, Y.; Qiao, M.; Fu, Y.; Wei, P.; Li, Y.; Liu, Z. Tissue Structure Changes of Aquilaria sinensis Xylem after Fungus Induction. Forests 2022, 13, 43. https://0-doi-org.brum.beds.ac.uk/10.3390/f13010043
AMA Style
Liu Y, Qiao M, Fu Y, Wei P, Li Y, Liu Z. Tissue Structure Changes of Aquilaria sinensis Xylem after Fungus Induction. Forests. 2022; 13(1):43. https://0-doi-org.brum.beds.ac.uk/10.3390/f13010043
Chicago/Turabian StyleLiu, Yuxin, Mengji Qiao, Yunlin Fu, Penglian Wei, Yingjian Li, and Zhigao Liu. 2022. "Tissue Structure Changes of Aquilaria sinensis Xylem after Fungus Induction" Forests 13, no. 1: 43. https://0-doi-org.brum.beds.ac.uk/10.3390/f13010043
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.