
The Interconnected Relationship between Auxin Concentration Gradient Changes in Chinese Fir Radial Stems and Dynamic Cambial Activity

1
Co-Innovation Center for Sustainable Forestry in Southern China, Key Laboratory of Forest Genetics & Biotechnology of Ministry of Education, Nanjing Forestry University, Nanjing 210037, China
2
College of Biology and Environment, Nanjing Forestry University, Nanjing 210037, China
*
Author to whom correspondence should be addressed.
Forests 2022, 13(10), 1698; https://0-doi-org.brum.beds.ac.uk/10.3390/f13101698
Submission received: 19 September 2022
/
Revised: 10 October 2022
/
Accepted: 12 October 2022
/
Published: 15 October 2022
(This article belongs to the Special Issue Forest Tree Breeding and Directed Cultivation Techniques)
Abstract
:Auxin has been shown to exhibit a striking concentration gradient distribution in radial sections of angiosperm and gymnosperm species, in which peak auxin levels are concentrated in dividing cambial cells, while the absolute auxin concentration sharply declines toward developing secondary phloem and xylem regions. The coincidence of auxin concentration gradient across shoot tissues and xylem cell developmental gradient has prompted that auxin could act as “a plant morphogen” to provide a positional signal for cambial cell development. However, the specific location of vascular cambium and the lack of mutants altering auxin distribution in shoots of woody species made further verification experiments difficult to explore. To address this issue, different concentrations of exogenous IAA were applied to decapitated Chinese fir (Cunninghamia lanceolata (Lamb.) Hook.) trunks in this study to induce the change in the auxin concentration gradient in radial stems, and its effects on cambial activities were examined on the physiological, cellular and molecular levels. Our findings manifested that exogenous IAA treatments resulted in vast changes in endogenous hormone concentrations (including IAA, ZR, GA3 and ABA), cambial cell developmental behaviors and transcriptional activities of genes related to polar auxin transport (PAT), auxin signaling, the biosynthesis and signal transduction of other plant hormones and the genetic control of cambial activity. Based on above findings, we postulated a model of auxin concentration gradient involved in the control of cambial activity and secondary growth in tree trunks. In this model, the contrasting expression of AUX1/LAX and PIN family carriers in distinct Chinese fir wood-forming tissues dynamically modulates PAT into the cambial zone adjacent to the secondary phloem side and secondary xylem tissues, resulting in a sharp and wide auxin spatial gradient distribution across shoots in different stages of secondary growth, respectively. This change in auxin concentration gradient distribution in radial sections in turn acts on cambial developmental behaviors by modulating the expression of auxin signaling genes and key transcription factors and the production of other plant hormones in distinct woody tissues. Findings in this study provide important insights for understanding the biological significance of auxin concentration gradient existing in the radial stems of woody species.
1. Introduction
Secondary growth in the tree trunks comes from the seasonal activity of vascular cambium, whose initial cells can undergo division and differentiation to continuously produce secondary phloem cells outwards and secondary xylem cells inwards, respectively [1]. The xylem precursors undergo further cell division and secondary cell wall modifications, ultimately giving rise to the most abundant renewable cellulose resources for humans but helping to relieve the greenhouse effect caused by excessive carbon dioxide emissions. Although the mechanism of secondary growth in woody species was not well understood, auxin had been proven to play a key role in this process [2,3].
Removal of the shoot apex (a major source of auxin in plants) of woody plants results in sharply reduced the capacity of cambial cell proliferation and secondary xylem cell formation in shoots [4], while application of exogenous IAA to the decapitated stumps restores secondary growth in shoots in a concentration-dependent manner [5], indicating that the PAT from shoot apexes into radial stems is essential for cambial cell development. More interestingly, auxin has been reported to show a striking concentration gradient distribution across shoots of woody plants, in which peak auxin levels occurs in the cambial zone, while the absolute auxin content sharply declines toward the developing secondary phloem and xylem [2]. The coincidence of auxin spatial gradient distribution and xylem cell developmental gradient has prompted that auxin could act as “a plant morphogen” to provide a positional cue for cambial cell development [2,6]. However, the constant existence of auxin peak levels in active and dormant cambiums and its limited correlation with auxin-responsive gene expression make this possibility unlikely [3,7].
At present, most of our knowledge about the action of auxin in cambial activity control comes from the model plant Arabidopsis thaliana (L.) Heynh, which has radial organization structures in the inflorescence stem and hypocotyl similar to those of trees (especially in hypocotyls) [8]. One pathway whereby auxin controls cambial activity is by its asymmetric distribution in shoot tissues [9,10]. AUX1/LAXs and PINs are the major carriers that mediate active auxin transportation into and out of plant cells, respectively [11,12,13]. Of them, PINs, are the key trigger of auxin asymmetric distribution in plant tissues via their polar localization on basal parenchyma cell plasma membranes [14,15]. pin1 mutants severely deform the formation and pattern of vascular tissues in Arabidopsis inflorescence stems and hypocotyls [16,17]. The AUX1/LAX and PIN family orthologous genes in poplar exhibit dynamic expression profiles in specific wood-forming tissues according to changes in seasonal cues [18], indicating that changes in PAT in radial stems of woody plants provides a basic signal for cambial seasonal activity. Response of cambial cells to auxin concentration changes in local shoots is another key aspect of the control of cambial activity mediated by auxin [7]. AUX/IAAs are the major inhibitors of auxin signal transduction in plants by forming heterodimers with ARF proteins under low cellular auxin levels [19]. Elevated cellular auxin acts as ‘a molecular clue’ to facilitate the interaction of AUX/IAAs with TIR1/AFB, resulting in the degradation of AUX/IAAs by a SCFTIR1/AFB E3 ligase complex and therefore the release of ARFs to activate the expression of downstream genes [19]. Overexpression of an artificially modified PttIAA3 gene in transgenic poplar lines to interrupt auxin signal transduction in shoots results in reduced cambial activity and secondary growth relative to the wild type [7]. Recently, ARF-tracing experiments of Brackmann, et al. [20] showed that ARF3 and ARF4 were universally expressed in vascular-forming tissues and acted as general activators of cambial cell proliferation, while the MP/ARF5 protein was specifically expressed in PXY-positive cells and antagonized the role of above ARFs in cambial activity. Moreover, all these ARFs act on cambial activity by modulating the expression of WOX4, a key mediator of cambial cell population maintenance [21,22,23,24]. Meanwhile, MP/ARF5 was found to mediate the expression of HD-ZIP genes encoding transcription factors functioning on cambial cell amplification and xylem cell differentiation [25,26,27]. Because the orthologous WOX4, ARF and HD-ZIP members were found to perform similar roles in secondary growth in poplar [28,29,30], conserved interaction relationships between these transcription factors and the auxin concentration gradient in radial sections could exist in trees.
In addition to abovementioned pathways, auxin shows complicated interaction relationships with other plant hormones, which also contribute to auxin-mediated secondary growth in the tree trunks [31]. Cytokinin (CK) is a master regulator of the identity and population of cambial cells in shoot tissues [32]. Consistent with this role, the highest CK levels were found to be concentrated in the secondary phloem tissue of poplar, distinct from the peak IAA levels observed in dividing cambial cells [33,34]. Interestingly, inducing CK overproduction in transgenic poplar lines increases auxin levels in the distinct cambial zone, indicating that CK acting on cambial activity requires the participation of auxin in the cambial zone [33]. CK mediates auxin concentrations in the distinct cambial zone majorly by stimulating the localization of PINs toward cambial cells [35]. Conversely, auxin, in turn, decreases CK content in shoots by repressing the CK biosynthesis gene expression, and therefore antagonizes the inhibition of CK on xylem cell differentiation [36]. GA is a stimulator of xylem cell development [37]. Application of both IAA and GA to decapitated poplar tree trunks was observed to more strongly stimulate cambial cell proliferation and secondary growth than either hormone treatment alone [38], indicating that GA synergistically interacts with auxin to mediate secondary growth in shoots. Auxin elevates GA content in shoots by upregulating the GA synthetic gene expression, and GA, in turn, promotes PAT into cambial cells by controlling PINs’ expression and localizations [38]. Additionally, IAA and GA were found to share a common signaling pathway in poplar woody tissues [38]. In Scots pine, ethylene levels increase in association with cambial activation in spring and peak during maximal tracheid formation [39], indicating that ethylene is also a stimulator of secondary growth in woody plants. Auxin promotes ethylene biosynthesis in shoots by stimulating the expression of ACS genes, and ethylene, in turn, mediates PAT in shoots [40,41,42]. ABA is a key signal factor controlling cambial activity under unfavorable conditions [43,44,45]. Recent evidence has shown that ABA mediates cambial activity by acting on auxin signaling factors [46,47], while auxin promotes ABA production via the ethylene-mediated pathway [48].
Despite these breakthroughs in unveiling the action of auxin in cambial activity control, the biological significance of auxin spatial gradient distribution in radial stems of woody plants was barely understood, largely because of the specific location of vascular cambium and the lack of PAT mutants in woody species. To address this issue, this study applied different concentrations of exogenous IAA to decapitated Chinese fir stumps to induce a change in auxin concentration gradient across shoot tissues and investigated its influence on cambial activity and related physiological and molecular environments. Chinese fir is one of the most widely cultivated gymnosperm species in China and plays a pivotal role in satisfying the increasing requirement of commercial timbers in China. This study has important significance for understanding the mechanism of wood formation in Chinese fir.
2. Materials and Methods
2.1. Decapitation and Exogenous IAA Treatments
In mid-February, when the vascular cambium in Chinese fir shoots is undergoing the dormancy–activation transition in Nanjing, China, a typical subtropical climate, 5-cm-long shoot apexes on the main trunk and all lateral branches and leaves were removed from two-year-old seedlings of ‘Yangkou’ 020 fir, an elite Chinese fir clone. After these plants were left untreated for 7 days, different concentrations of exogenous IAA and lanolin mixtures (including 0, 1.0, and 3.0 mg.iaa/g.lanolin) were applied to the decapitated stumps. The exogenous IAA treatments were repeated every 3 days and continued for 30 days. Each treatment involved fifty biological replicates, and the plants with intact shoot apexes served as the positive control (PC). All these seedlings were cultivated under natural conditions.
2.2. Measurement of Endogenous Hormones
Two-centimeter-long shoot segments adjacent to the cutting wounds of samples treated with exogenous IAA and at the parallel positions in PC were collected every 2 days. Ultimately, 0.5 g fresh weight of shoot segments from different samples were selected and used as the materials of endogenous hormone isolation. The methods of endogenous hormone extraction and measurement were by methanol and an indirect enzyme-linked immunosorbent assay (ELISA), respectively. Each hormone had triple biological replicates. Two-way ANOVA was executed to analyze the difference in each hormone concentrations in differently treated stages and samples.
2.3. Anatomical Observation and Analysis of Secondary Growth Changes
The sampling method of materials for anatomical analysis was similar to that in the “measurement of endogenous hormones” section. FAA and glycerol–alcohol solutions were used to fix and soften shoot samples, respectively. After treatments of dehydration, clearing and embedding by paraffin, 12.0-μm-thick cross sections were prepared. A 0.1% toluidine blue solution was used to dye sections and the observation and photography of anatomical structures were under an optical microscope. To evaluate the change in secondary growth in differently treated shoot samples, radial thickening growth in cross sections was measured from three directions by ImageJ software and one-way ANOVA was executed to generate statics on their differences.
2.4. Gene Location and Expression Profile Analysis
The bare tree trunks of four-year-old ‘Yangkou’ 020 fir were collected in mid-March when cambial cells were undergoing vigorous secondary growth. Developing secondary phloem and xylem tissues and cambium were specifically isolated by a frozen laser microsection method and used as the materials for gene location analysis. Shoot segments collected from differently treated stages of samples (the sampling method was similar to that in above sections) were used as the materials for gene expression profile analysis.
Total RNA isolation was performed according to the procedure of MiniBEST Universal RNA Extraction Kit (Cat No.9767, Takara, Kusatsu, Japan) and the first-strand cDNA synthesis was by a PrimeScript™ II 1st Strand cDNA Synthesis Kit (Cat No.6210A, Takara, Japan). The original sequences of genes for qPCR was downregulated from a Chinese fir transcriptome submitted by Huang, et al. [49] (http://0-www-ncbi-nlm-nih-gov.brum.beds.ac.uk/Traces/sra, accession number: SRR475258) (accessed on 20 September 2019). cDNA amplification was performed by using Taq Pro Universal SYBR qPCR Master Mix (Cat No. Q712, Vazyme, Nanjing, China) and was run on an ABI 7500 machine (ABI, Foster City, CA, USA). 60S rRNA was the reference gene. One-way ANOVA was executed to analyze the difference in gene RQ values in differently treated samples and the RQ value in PC acted as the control.
3. Results
3.1. Changes in Endogenous Hormones
To reveal the influence of decapitation and subsequent exogenous IAA application on the production of endogenous hormones in Chinese fir shoots, we analyzed the changes in IAA, CK, GA and ABA concentrations in samples from different stages of treatment. The results of variance analysis showed that the difference in each hormone concentration in differently treated samples, stages and their interactions reached extremely significant levels (Table 1), indicating that interfering with PAT from shoot apexes via exogenous IAA application upon decapitated stumps substantially altered the homeostasis of these endogenous hormones in Chinese fir wood-forming tissues.
To further understand how exogenous IAA treatments influenced the content and distribution of IAA, ZR, GA3 and ABA in developing Chinese fir shoots, we compared the fluctuation curves of these endogenous hormones in differently treated samples (Figure 1).
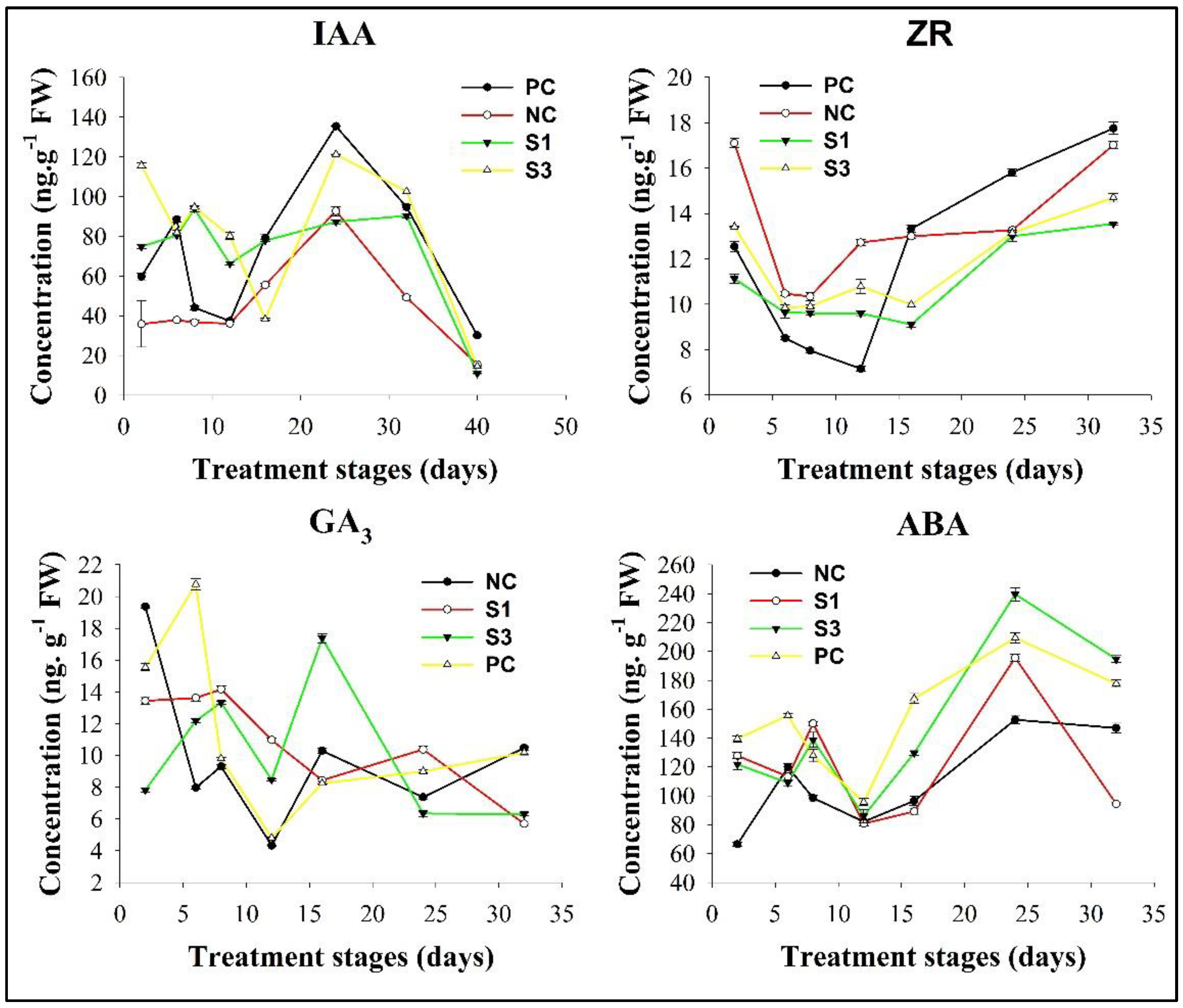
3.1.1. IAA
IAA presented two striking level peaks in PC shoots, which occurred on the 6th and 24th days of treatment, respectively (Figure 1). In addition, the absolute IAA concentration was lower in the initial stage of PC shoots than in the lateral stage, indicating that IAA exhibited a regular concentration change during secondary growth in shoots of woody plants. In contrast to PC, both S3 and NC exhibited a decreased pattern of IAA fluctuation in shoots in the initial stage, while S1 presented an IAA fluctuation pattern similar to that in PC. Despite these distinct IAA fluctuation patterns at the start of treatment, both S1 and S3 reached the 1st IAA level peak on the 8th day of treatment, which was 2 days later than the corresponding one in PC, while IAA concentrations in NC continually decreased until the 12th day of treatment. Interestingly, all decapitated samples presented the 2nd IAA level peak on the 24th day of treatment, overlapping with the 2nd one in PC, although their peak IAA levels decreased in an exogenous IAA concentration-dependent manner.
3.1.2. ZR
ZR is an important intermediate for the tZ type of CK biosynthesis [50] and presented a decreased pattern in the initial stage of PC shoots, in which its two peaks occurred on the 8th and 16th days of treatment, respectively, opposite to the IAA fluctuation patterns in the counterpart stage of PC shoots (Figure 1). Similar correlations of ZR and IAA concentration fluctuation were also observed in the initial stage of S1 and S3 shoots, in which the first ZR peak, which occurred on the 12th day of treatment, although the absolute ZR concentration in S3 shoots was much higher than that in S1. In contrast, NC presented an overlapping ZR peak with PC in the initial stage, but its peak ZR levels were higher than those in PC. Regardless of these changes in the initial stage, all decapitated samples presented another ZR peak on the 24th day of treatment, overlapping with the second IAA peak. Moreover, the absolute ZR concentration in this stage of S3 shoots was much higher than that in the other three samples. These findings indicated that the decapitation and exogenous IAA treatments intensively altered the content and distribution of CK in Chinese fir woody tissues.
3.1.3. GA3
GA3 is a major bioactive form of GAs in plants. Fluctuation analysis showed that this hormone exhibited a concentration change positively correlated with that of IAA in PC shoots, but its absolute levels were much higher in the initial stage of shoots than in the lateral stage (Figure 1), opposite to the performance of endogenous IAA. In contrast to PC, although similar fluctuation patterns of GA3 were observed in S1 and S3 shoots in the initial stage, S3 rapidly developed another GA3 peak on the 16th day of treatment, while S1 presented a GA3 peak overlapping with that in PC on the 24th day. Differing from S1 and S3, NC exhibited a GA3 fluctuation pattern correlated with that of IAA.
3.1.4. ABA
ABA is a key mediator of cambial activity under various stressful conditions [43,44]. Interestingly, ABA exhibited a fluctuation pattern tightly correlated with that of IAA in PC shoots (Figure 1). Similar phenomena were also observed in the samples treated by exogenous IAA, although the absolute ABA concentration was much higher in S3 shoots than in other samples in the later stage. In view of the indirect role of auxin in ABA production in plant tissues by ethylene-mediated pathways [48], we concluded that the correlation of ABA and IAA concentration fluctuation in differently treated samples could be related to ethylene.
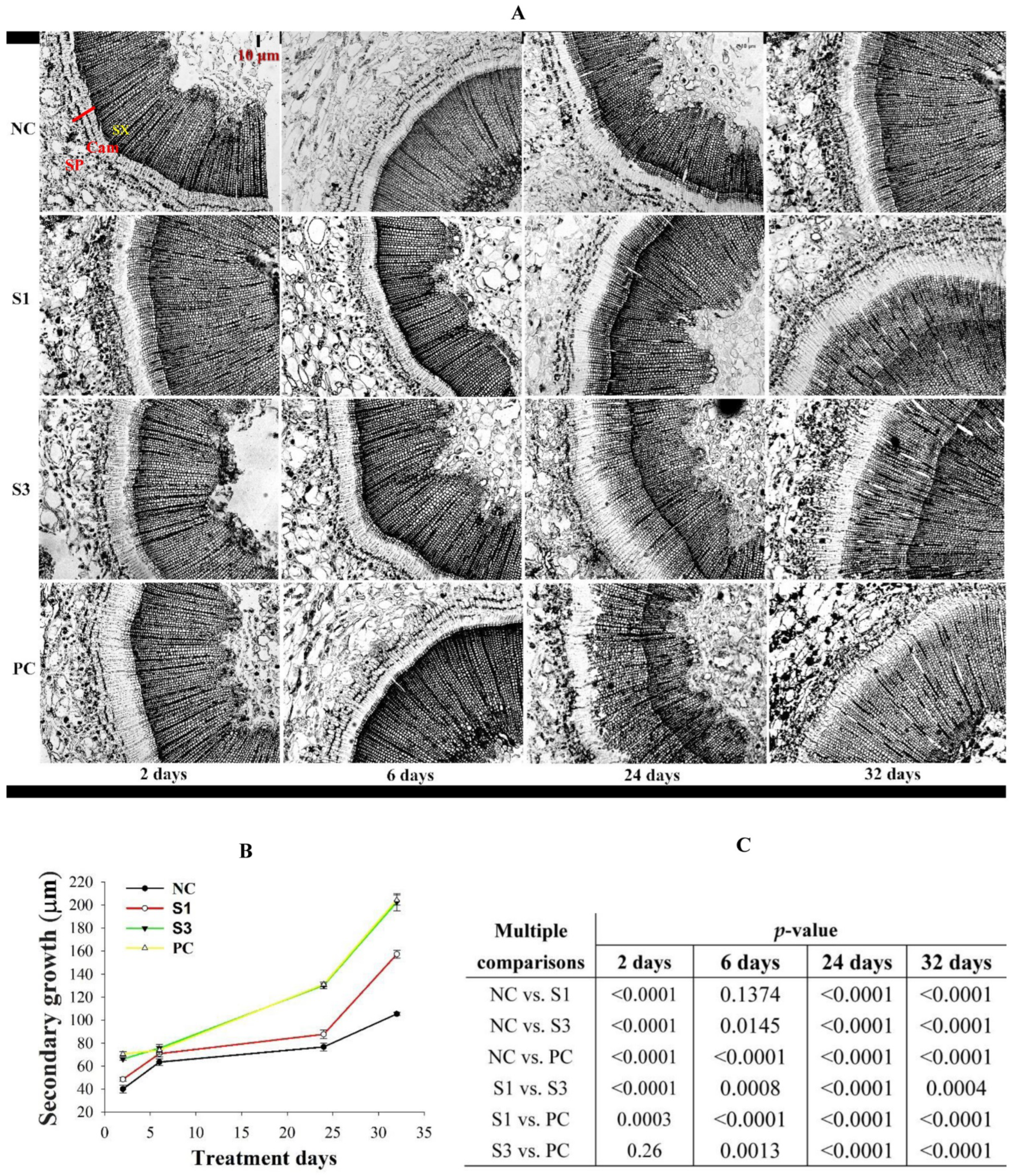
3.2. Changes in Cambial Activity
To unveil the effects of decapitation and exogenous IAA treatments on vascular formation and patterns in Chinese fir shoots, we analyzed the change in anatomical structures across differently treated shoot samples (Figure 2A). In the initial stage of treatment, a large number of cambial dividing cells but limited tracheid cells were observed in PC shoots, indicating that the cambial cells in PC were undergoing vigorous mitosis in this stage. In contrast to PC, decapitated samples exhibited a gradient reduction in cambial activity according to the exogenous IAA concentration applied to decapitated trunks. Interestingly, more tracheid cells were observed in this stage of S3 shoots, while the production of tracheid cells in NC and S1 was comparable to that in PC. Corresponding to the overlapped IAA peaks in all samples in the lateral stage, we found that cambial derivative cells in PC shoots sharply reduced, which was associated with tracheid cells being produced in the xylem side on a large scale, indicating that PC was undergoing vigorous xylem cell differentiation in this stage. In contrast to PC, the samples treated with exogenous IAA retained a strong capacity for cambial cell amplification, including NC, while their production of tracheid cells was reduced in a concentration-dependent manner (especially in NC). As a consequence of differential activities of cambial cell amplification and xylem cell differentiation, S3 ultimately produced secondary growth comparable to that in PC, while the radial thickening growth in NC and S1 shoots was reduced in an IAA concentration-dependent manner (Figure 2B,C).
3.3. Changes in Gene Expression
To reveal the molecular basis underlying the distinct cambial activities and endogenous hormone changes in differently treated samples, we analyzed the expression profiles of related genes (see Table S1 Functional annotation and classification of selected genes in Supplementary Data) in the developing shoots according to their localizations in Chinese fir wood-forming tissues (see Figure S1 Tissue-specific expression of genes in Chinese fir wood-forming tissues in Supplementary Data).
3.3.1. PAT Carrier Genes
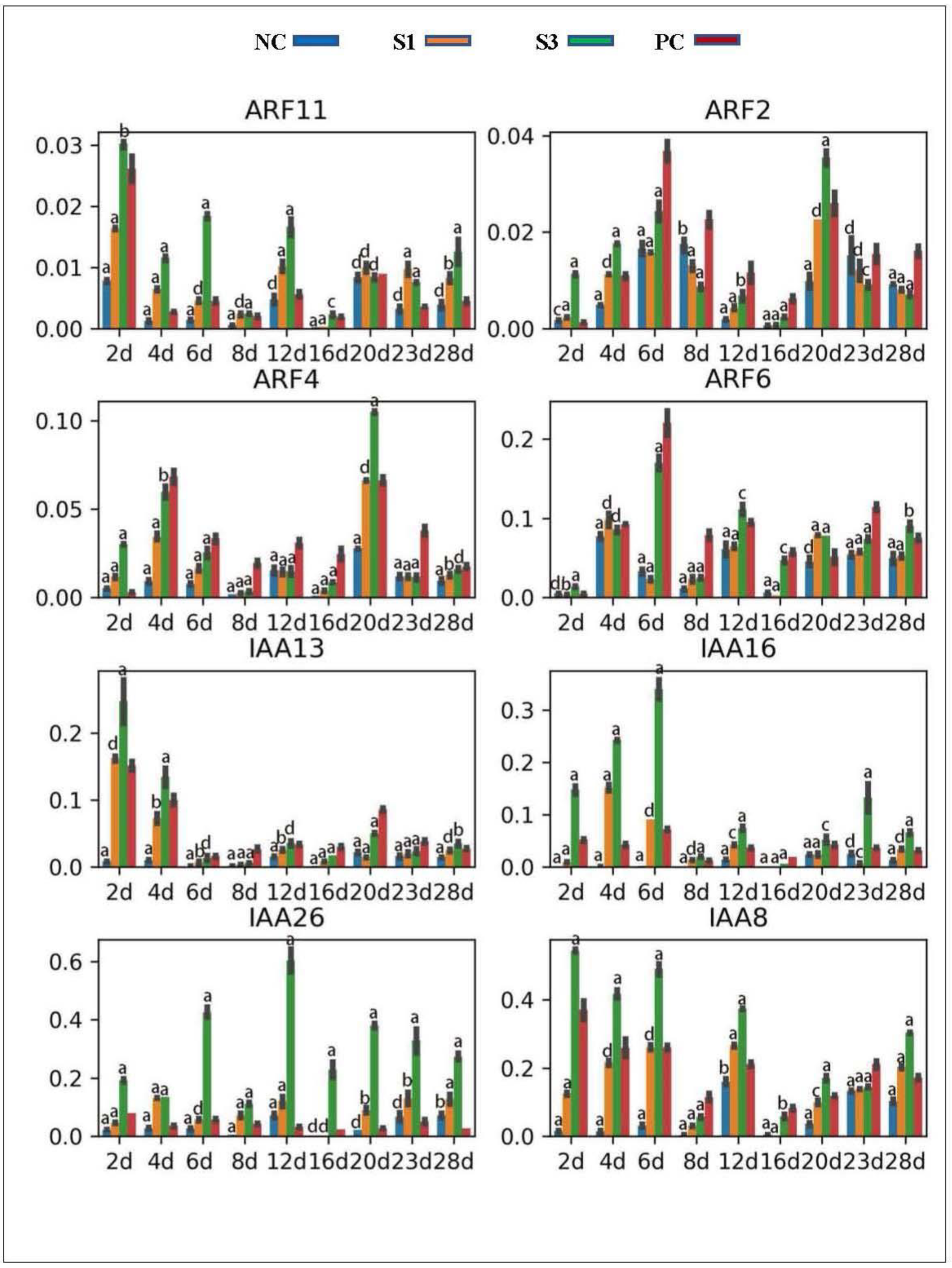
AUX1/LAXs and PINs are the major carriers that mediate auxin active transport into and out of plant cells [11,12,51]. Location analysis showed that the Chinese fir AUX1 gene was specifically expressed in the secondary phloem cells, while the other AUX1/LAX family homolog, LAX3, concentrated its expression in both secondary phloem and cambial cells, indicating that both were mediators of the IAA peak distribution in Chinese fir cambial zone. qPCR analysis revealed that both AUX1 and LAX3 exhibited dynamic expression profiles according to the IAA concentration change in PC shoots, but more of their transcriptional abundance was observed in the initial stage than in the lateral stage (especially for the AUX1 gene) (Figure 3), indicating that the expression of AUX1/LAXs in Chinese fir cambial zone was feedback-regulated by IAA. In contrast to PC, exogenous IAA treatments strongly stimulated the expression of AUX1 in S1 and S3 shoots (especially in the lateral stage) but kept LAX3 at a low transcriptional level in all decapitated samples, indicating that interfering with PAT in shoots resulted in differential expression of AUX1/LAXs according to their locations in wood-forming tissues. PINs are the key triggers of asymmetric auxin distribution in local plant tissues [14,15]. Location analysis found that the Chinese fir PIN1 and PIN3 genes concentrated their transcriptional abundance in cambial cells and secondary xylem tissues, while PIN2 and PIN4 were mostly expressed in the abaxial zone of shoot tissues. Correspondingly, we found that PIN1, 3 and 4 exhibited stronger expression in PC shoots in the initial stage than in the lateral stage, which was opposite to the expression profile of PIN2 in PC shoots (Figure 3), indicating that the expression of PINs was dynamically modulated according to their locations in woody tissues during secondary growth. In contrast to PC, exogenous IAA treatments stimulated the expression of PIN1, 3 and 4 in different stages of S1 and S3 shoots (especially for PIN3) to different extents, but downregulated the transcription of PIN2 in lateral stages. In contrast to S1 and S3, all these PINs merely presented limited transcriptional activities in NC shoots in lateral stages.
3.3.2. Auxin Signaling Genes
AUX/IAAs and ARFs are the key factors the mediate plant response to local auxin concentration changes [19]. Location analysis revealed that the IAA8 and IAA26 genes in Chinese fir were most abundantly expressed in cambial cells, while IAA13 and IAA16 concentrated their transcripts in the secondary phloem tissue, indicating that they were the mediators of the auxin response in the Chinese fir cambial zone. Consistent with this conclusion, all these AUX/IAAs were observed to exhibit expression profiles positively correlated with PAT changes in the cambial zone of differently treated samples (Figure 4). In contrast to AUX/IAAs, the selected ARF genes exhibited differential expression patterns in Chinese fir wood-forming tissues. Both ARF6 and ARF11 focused most of their expression in cambial cells, while ARF2 and ARF4 were expressed on both sides of the cambium and in various wood-forming tissues, respectively. Correspondingly, the expression of ARF6 and ARF11 was more strongly stimulated in PC shoots in the initial stage than in the lateral stage (especially ARF11), while ARF2 and ARF4 presented expression profiles tightly correlated with IAA concentration change in the PC shoots (Figure 4), indicating a differential response of these ARFs to PAT changes in Chinese fir cambial zone. In contrast to PC, exogenous IAA treatments strongly stimulated the expression of ARF11 but downregulated the transcriptional activity of ARF2 and ARF4 in different stages of S1 and S3 shoots (Figure 4).
3.3.3. Genes Participating in CK Production and Signal Transduction
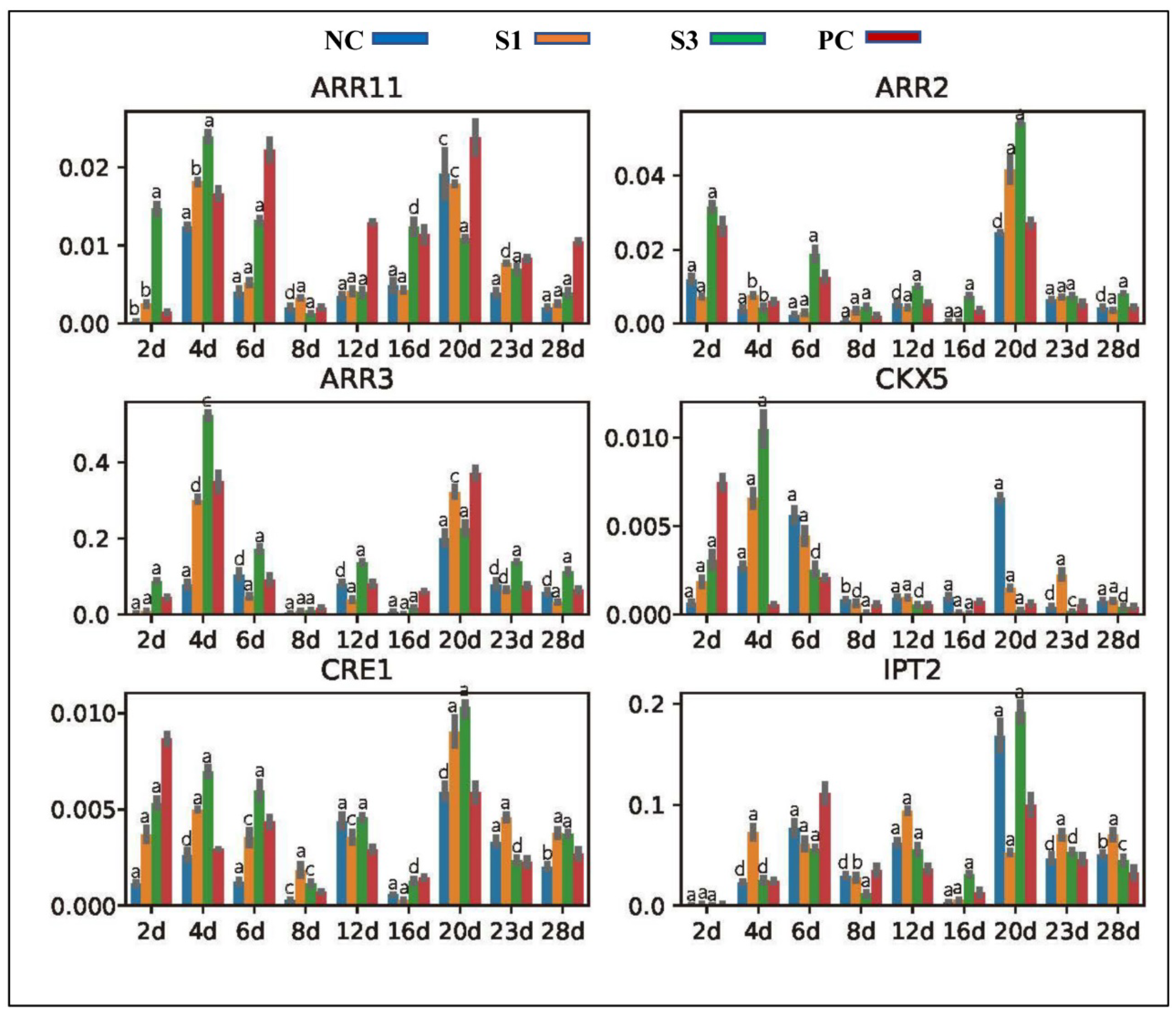
IPT and CKX family proteins are the key mediators of CK homeostasis in plant tissues by their involvement in the processes of CK biosynthesis and catabolism, respectively [52]. Location analysis revealed that the Chinese fir IPT2 gene concentrated most of its transcripts in the secondary phloem tissue and cambial cells, while CKX5 was universally expressed in woody tissues, indicating that they together mediate the CK homeostasis in Chinese fir shoots. qPCR analysis showed that IPT2 presented an expression profile positively correlated with the IAA concentration change in PC shoots, while CKX5 exhibited a downregulated expression profile (Figure 5). Connecting the self-regulatory mechanism of CK production in plant tissues, we concluded that the expression profiles of IPT2 and CKX5 in PC shoots could be the result of overall regulation by CK and auxin signals. Consistent with this conclusion, similar expression profiles of CKX5 and IPT2 were observed in samples treated by exogenous IAA (Figure 5). CK triggers plant response by a series of phosphorylation and dephosphorylation relays, which require the participation of the receptor kinases CRE1/AHK4 and AHK3, as well as ARRs [53,54]. In accordance with the change in CK production in different treatment samples, we found that the CRE1 and ARR2 genes both focus most of their transcripts in Chinese fir cambial cells, as well as ARR3 and ARR11, which were abundantly expressed in the abaxial zone of shoots and exhibited correlated expression profiles with ZR concentration changes (Figure 5).
3.3.4. Genes Participating in GA Production and Signal Transduction
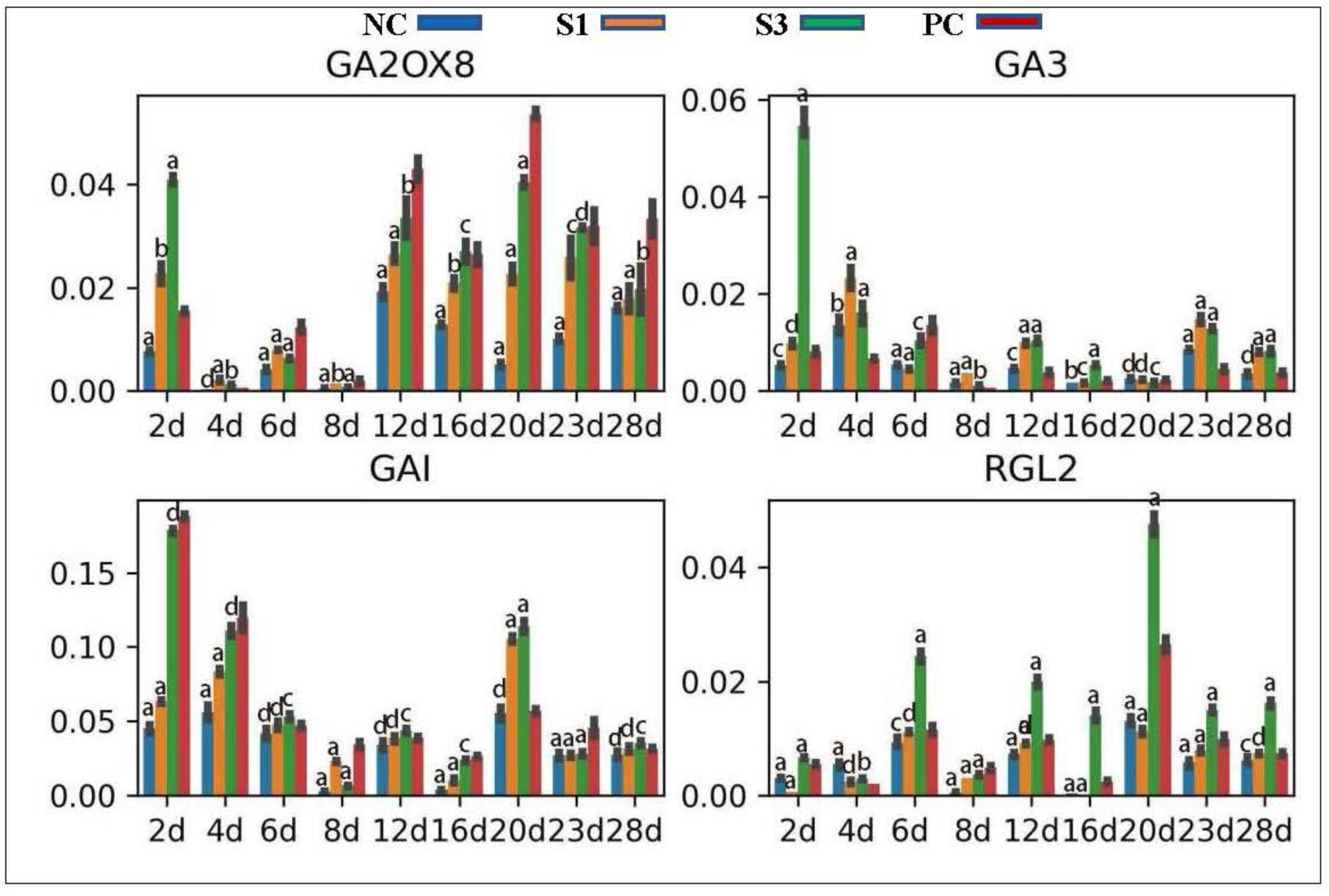
GA3 is a key enzyme in the GA biosynthetic pathway [55], while GA2ox family proteins are essential enzymes for GA degradation [56]. Location analysis showed that both the GA3 and GA2ox8 genes in Chinese fir concentrated most of their transcripts in the secondary phloem tissue, indicating that they together mediated GA homeostasis in this woody tissue. Profiling analysis found that GA3 exhibited a stronger upregulation of expression in PC shoots in the initial stage than in the lateral stage, while the GA2ox8 gene presented an inverse expression profile (Figure 6), consistent with the change in GA3 concentrations in PC. In contrast to PC, exogenous IAA treatments stimulated GA3 expression but decreased the transcripts of GA2ox8 in Chinese fir shoots in a concentration-dependent manner (Figure 6). DELLAs are the inhibitors of GA signaling and GA, in turn, triggers the degradation of DELLAs in plant tissues [57]. Our analysis showed that the Chinese fir GAI gene was abundantly expressed in the secondary phloem and cambial cells, while the RGL2 transcripts was concentrated in the secondary phloem tissue, indicating that both of them mediate GA response in the Chinese fir cambial zone. qPCR analysis revealed that these two DELLA genes exhibited opposite expression profiles in differently treated samples, reflecting a differential change in GA production in Chinese fir woody tissues (Figure 6).
3.3.5. Genes Participating in ABA Production and Signal Transduction
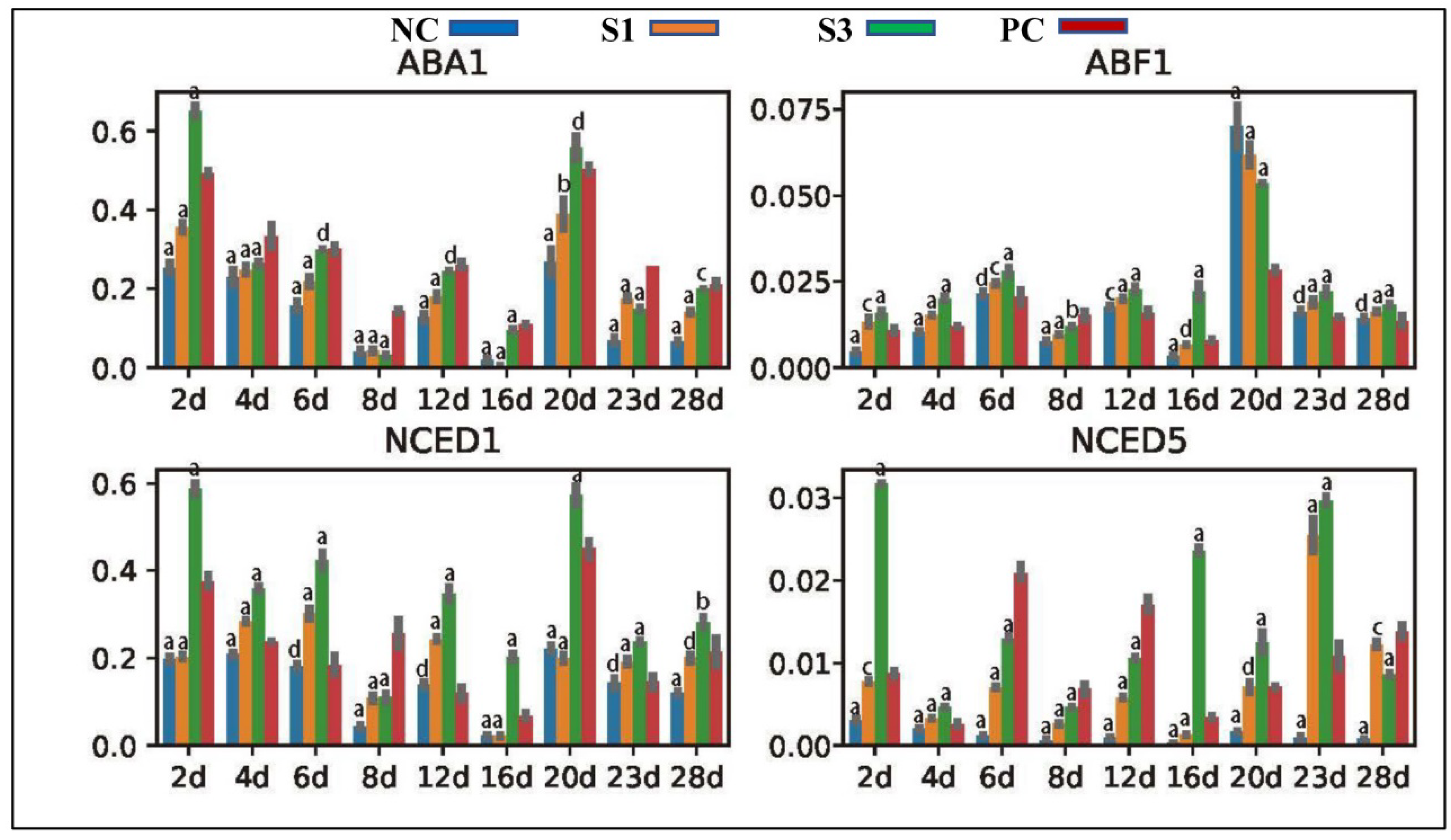
ABA1 and NCEDs are enzymes essential for ABA production in plants [58]. Location analysis found that the ABA1, NECED1 and NCED5 genes in Chinese fir were most abundantly expressed in the adaxial zone of shoots, indicating that all of them modulate ABA production in the Chinese fir cambial zone. qPCR analysis revealed that ABA1 and NCED1 presented a totally downregulated expression profile in PC shoots, which was opposite to the pattern of NCED5 expression in PC (Figure 7), indicating that the expression of these ABA biosynthetic genes in the Chinese fir cambial zone was mediated by different factors, and NCED5 would be an intermediate of IAA indirectly mediating ABA production in Chinese fir shoots via the ethylene pathway. In contrast to PC, exogenous IAA treatments strongly stimulated the transcription of NCED1 and NCED5 in S1 and S3 shoots in the initial stage and lateral stage, respectively, indicating a differential response of these two genes to IAA concentration changes in Chinese fir shoots. ABF1 is a key factor triggering the expression of ABA responsive genes [59]. The ABF1 gene in Chinese fir was universally expressed in wood-forming tissues and exhibited an expression profile positively correlated with IAA concentration changes in differently treated samples (Figure 7), further confirming an indirect role of IAA in ABA production in Chinese fir shoots.
To identify the synchronicity of ABA and ethylene production in Chinese fir shoots, we also analyzed the expression profiles of genes related to ethylene biosynthesis and signal transduction in differently treated samples. qPCR results showed that the ethylene biosynthesis gene ACS12 and ethylene signaling genes (including EIN3, EIL1 and EBF1), all of which focused their abundant expression in the adaxial zone of Chinese fir shoots, exhibited tightly correlated expression profiles with IAA concentration changes in differently treated samples (see Figure S2 Expression profiles of genes participating in ethylene production and signal transduction in Supplementary Data), confirming our conclusion of ABA production in different samples being mediated by ethylene.
3.3.6. Genetic Regulators
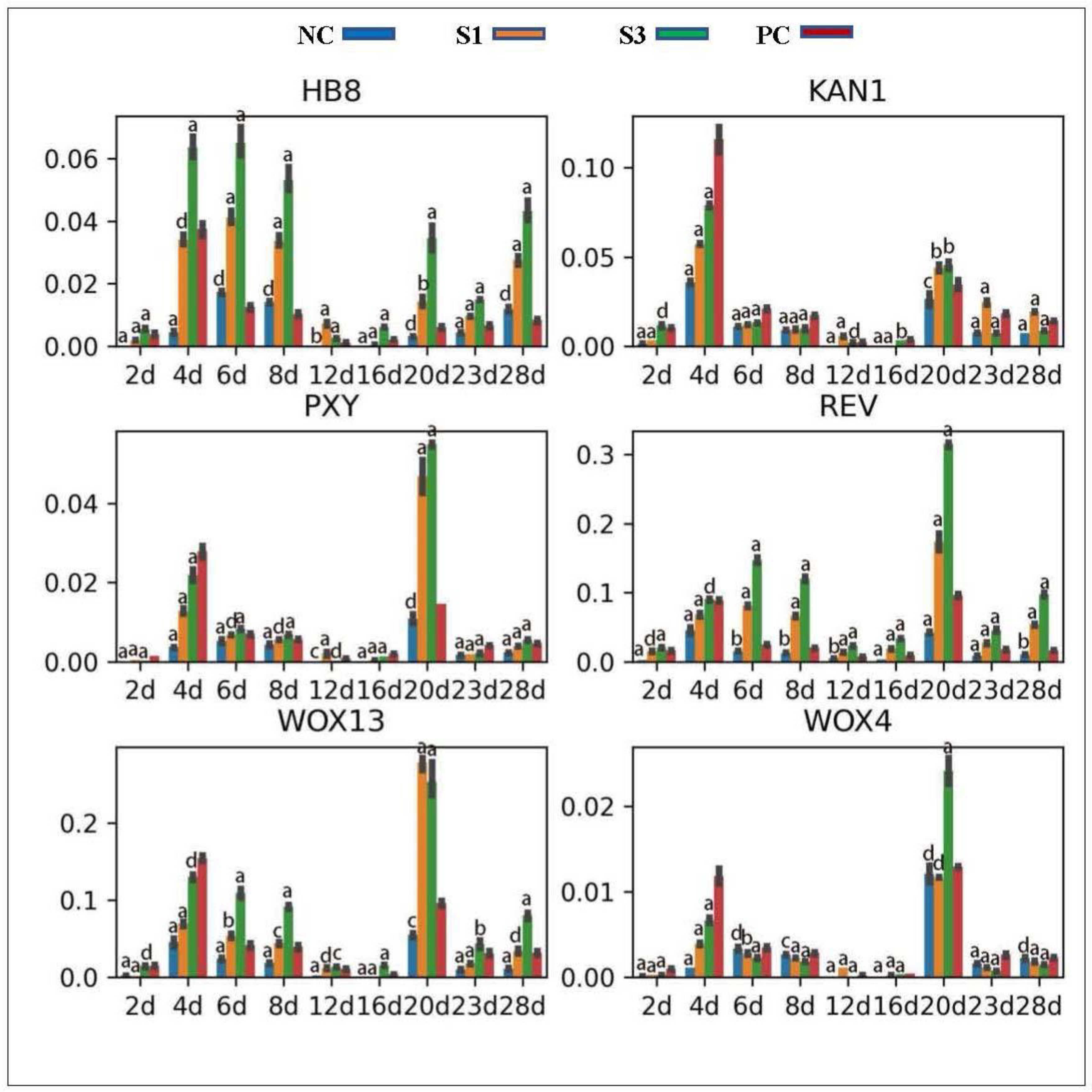
To reveal the response of key cambial regulators to IAA concentration changes in differently treated samples, we analyzed the expression profiles of PXY, WOX4 and WOX13, both of which are key components of the ligand–receptor system involved in the control of cambial cell niche in plant shoot tissues [24,60,61], as well as some HD-ZIP and KAN family genes. Location analysis showed that the PXY and WOX4 genes in Chinese fir were most highly expressed in the secondary phloem and cambial cells, while the WOX13 gene, a redundant allele of WOX4 [62], focused its abundant expression in the secondary phloem tissue. qPCR analysis found that PXY, WOX4 and WOX13 exhibited expression profiles tightly correlated with IAA concentration changes in PC shoots (especially for the WOX13 gene) (Figure 8), indicating that their expression in distinct Chinese fir woody tissues was positively mediated by IAA. In contrast to PC, exogenous IAA treatments strongly stimulated the expression of WOX13 in S3 shoots but downregulated the expression levels of WOX4 in all decapitated samples, indicating a differential response of these genes to auxin concentration gradient changes in shoot tissues. HD-ZIPs and KANs antagonistically interact to mediate vascular formation and patterns in shoots [63,64,65]. In Chinese fir, the HD-ZIP genes HB8 and REV were abundantly expressed in the adaxial and abaxial zones of shoots, respectively, while the KAN1 gene was specifically expressed in the secondary phloem tissue, indicating that they would perform conserved roles in the regulation of secondary growth in Chinese fir shoots. Consistent with this conclusion, we found that these HD-ZIPs and KANs presented expression profiles positively and negatively correlated with IAA concentration changes in differently treated samples, respectively (Figure 8).
4. Discussion
4.1. Auxin Concentration Gradient Changes Provide a Basic Cue for Cambial Activity
Based on the accurate measurement of endogenous IAA in different wood-forming tissues of Scots pine, Uggla, Moritz, Sandberg and Sundberg [2] concluded an IAA concentration gradient distribution pattern across shoot tissues of woody plants, in which the IAA peak levels are concentrated in dividing cambial cells, while the absolute IAA concentration sharply declines toward the secondary phloem and xylem sides. The high coincidence of auxin spatial distribution and xylem cell developmental gradients in radial sections of woody plants had prompted that auxin could act as ‘a plant morphogen’ to provide a positional cue for cambial activity [2,6].
Based on variance analysis, we found that the differences in endogenous IAA concentrations among treatments, treatment stages and their interaction all reached extremely significant levels. Further analysis revealed that exogenous IAA exhibited a regular fluctuation pattern in PC shoots, in which two IAA peaks occurred on the 6th and 24th days of treatment, respectively, moreover, the 1st IAA peak levels were much lower than the 2nd ones. In contrast to PC, exogenous IAA treatments not only interfered with the first IAA peak initiation in Chinese fir shoots, but also induced a dose-dependent change in IAA concentrations in the lateral stage of shoots. Molecular studies revealed that AUX1 and LAX3, two auxin influx genes in Chinese fir and both abundantly expressed in the abaxial zone of Chinese fir shoots, exhibited an expression profile opposite to the IAA fluctuation pattern in PC shoots, indicating that the expression of these two genes is feedback-regulated by auxin and therefore maintains IAA peak levels in the cambial zone adjacent to the secondary phloem side. In contrast to AUX1/LAXs, the PIN1 and PIN3 genes were both abundantly expressed in the adaxial zone of Chinese fir shoots and exhibited a greater upregulation of expression in PC shoots in the initial stage than in the lateral stage, while the PIN2 gene, which focused most of its expression in the secondary phloem cells presented an inverse expression profile. Thus, a sharp and wide IAA concentration gradient distribution pattern is established in different developmental stages of PC shoots, respectively. In contrast to PC, exogenous IAA treatments stimulated the expression of AUX1, PIN1 and PIN3 but reversed the expression profile of PIN2 in different treatment stages of Chinese fir shoots, therefore resulted in distinct IAA gradient distribution patterns in Chinese fir radial sections during secondary growth.
Corresponding to the change in PAT in differently treated samples, we found that PC exhibited a stronger capacity for cambial cell amplification than samples with exogenous IAA treatment in initial stages, while S3 presented a greater ability for tracheid cell differentiation than PC and other two decapitated samples. These observations indicated that the change in IAA concentrations in the cambial zone adjacent to the secondary phloem side is tightly correlated with cambial developmental behaviors. When the IAA concentration in this region exceeds a threshold, it would promote the differentiation of cambial cells toward xylem cells, while a low IAA concentration would facilitate cambial cell proliferation. Differing from the performances of differently treated samples in the initial stage, PC sharply reduced its cambial activity in the lateral stage of shoots, which was replaced by tracheid cell formation at a large scale. In contrast to PC, samples treated by exogenous IAA presented a certain capacity for cambial cell amplification in this stage of shoots, but their production of tracheid cells was reduced in a concentration-dependent manner. These contrasting results implied that the elevated IAA distribution in the adaxial zone of shoots facilitates the cambial cell differentiation into secondary xylem cells.
Altogether, our physiological, cellular and molecular data strongly indicated that changes in auxin concentration gradient across the radial section of woody plants provide a basic cue for cambial activity and secondary growth.
4.2. Pathways of Auxin Concentration Gradient Triggering Cambial Activity
4.2.1. Auxin Signaling Pathways
Cellular response to the PAT change in local tissues is a key aspect of auxin triggering plant growth [19]. Under low auxin levels, AUX/IAAs form heterodimers with ARFs and prevent the activation of ARFs on auxin-responsive gene transcription. When cellular auxin levels are high, auxin binds to and induces the configuration change of AUX/IAA proteins and facilitates AUX/IAAs interacting with TIR1/AFB, resulting in the degradation of AUX/IAAs and therefore releasing ARFs to activate the expression of downstream target genes. Our qPCR data revealed that the ARF and AUX/IAA family genes displayed a differential response to the PAT change in differently treated samples according to their locations in Chinese fir wood-forming tissues. Thus, the expression profiles of ARFs and AUX/IAAs in distinct Chinese fir woody tissues would, in turn, interpret the signals of auxin concentration gradient across shoots into detailed cambial developmental behaviors.
4.2.2. Differential Response of Other Plant Hormones to PAT Changes in Chinese Fir Shoots
The fulfillment of actions of auxin on cambial activities requires the participation of other plant hormones in distinct woody tissues [31]. Consistent with this conclusion, our variance analysis found that exogenous IAA treatments on the decapitated Chinese fir trunk induced an extremely significant change in ZR, GA3 and ABA concentrations in Chinese fir shoots, in contrast to that in PC. ZR is an important intermediate in the tZ and cZ biosynthesis pathways and curve analysis found that this substance exhibited a fluctuation pattern complementary to that of IAA in PC shoots. In contrast to PC, exogenous IAA treatments did not alter the ZR fluctuation pattern in the initial stage of Chinese fir shoots, but resulted in overlapping ZR and IAA peaks in the lateral stage. Correspondingly, genes related to CK synthesis, catabolism and signaling exhibited regular expression profiles according to the change in ZR concentration in differently treated samples. According to the self-regulatory mechanism of CK biosynthesis, we concluded that auxin could negatively mediate CK production in Chinese fir shoots by conserved pathways [36] and therefore relieves the role of CK in cambial cell niche to promote tracheid cell differentiation [34]. GA is a stimulator of wood formation and has been indicated to synergistically interact with auxin to mediate cambial activity [38]. In line with this conclusion, GA3 was observed to exhibit a fluctuation pattern positively correlated with that of IAA in differently treated samples (especially in the initial stage of shoots), which was further confirmed by expression profiles of genes related to GA production and signaling. ABA is a key mediator of cambial activity under unfavorable conditions [43,44,45]. Interestingly, our analysis found that either the ABA levels or the expression of genes related to ABA production and signaling exhibited positively correlated changes with the IAA concentration fluctuation in differently treated samples. Based on these findings and the interconnected relationship of ethylene and ABA production [48], we concluded that an intensive change in ethylene production could occur in differently treated samples, moreover, which was confirmed by expression profiles of genes participating in ethylene production and signal transduction in these samples.
4.2.3. Response of Key Genetic Regulators to PAT Changes in Chinese Fir Shoots
Auxin mediates cambial activity in plant shoots partly by interacting with key genetic regulators, including the ligan–receptor system, HD-ZIPs and KANs [20,21,22,23,24,66]. Profiling analysis revealed that WOX13, which focused most of its expression in the secondary phloem tissue of Chinese fir shoots, exhibited positively correlated expression profiles with PAT changes in differently treated samples, while the PXY and WOX4 genes both concentrated their abundant expression in cambial cells and secondary xylem tissues presented a limited response to IAA concentration changes in various samples. HD-ZIPs and KANs antagonistically interact to mediate vasculature formation and patterns in plants. Profiling analysis revealed that HB8 and REV presented a positive response to IAA concentration changes in differently treated samples, distinct from the expression profiles of KAN1. These findings implied that auxin concentration gradient participates in cambial activity control partly by influencing the expression of key genetic factors in distinct woody tissues.
5. Conclusions
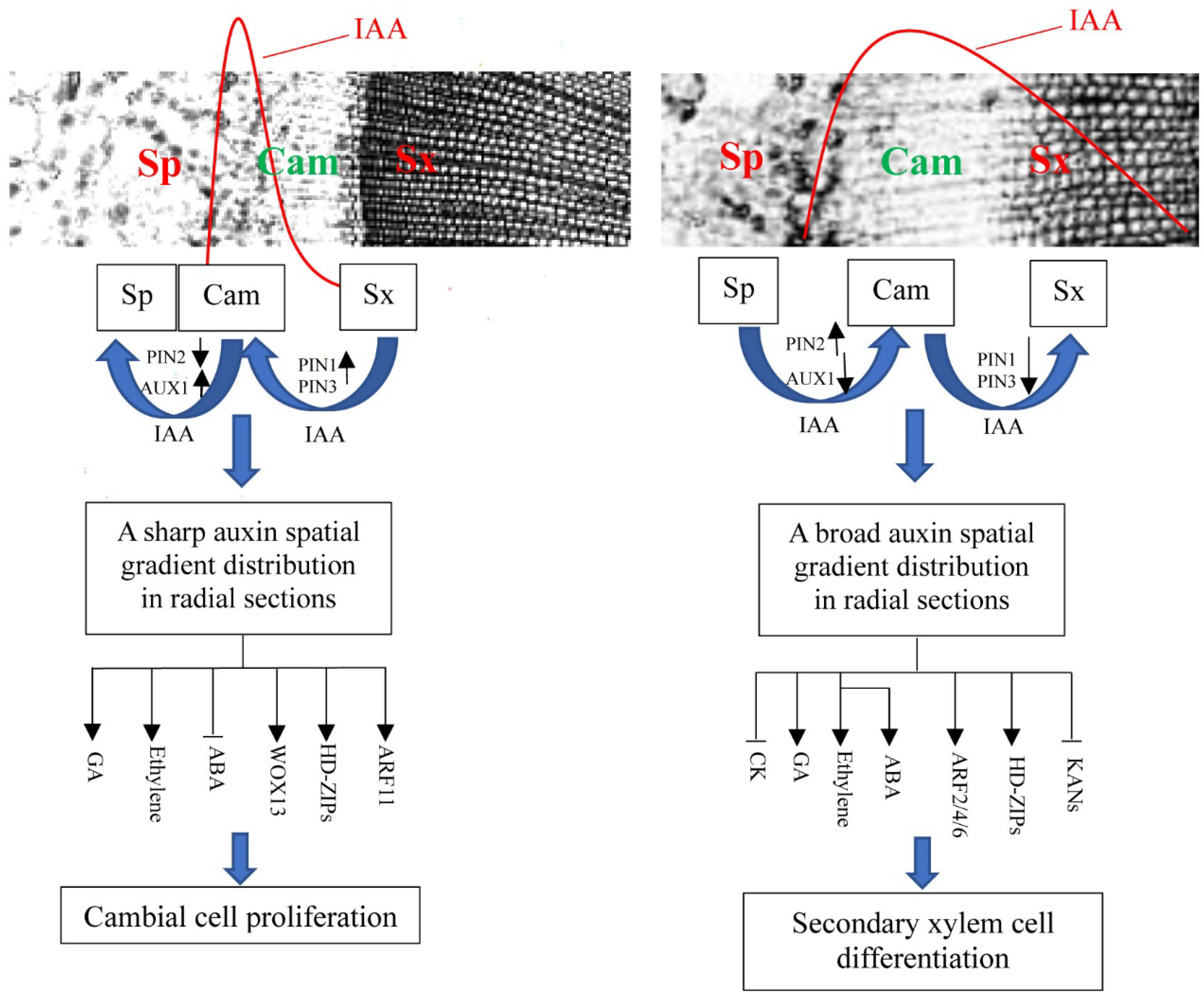
Based on the dynamic effects of decapitation and exogenous IAA treatments on endogenous hormone production, cambial activity and related gene expression in Chinese fir shoots, we proposed a model of auxin spatial gradient distribution across tree trunks acting on cambial activity control (Figure 9). In initially growing shoots, the differential expression profiles of AUX1/LAXs and PINs in distinct wood-forming tissues drives auxin directional flow into the cambial zone adjacent to the secondary phloem side but decreases the polar auxin distribution in the secondary xylem region, thereby resulting in a sharp IAA gradient distribution across shoots. During the secondary xylem cells being largely produced, the opposite expression profiles of AUX1/LAXs and PINs in distinct wood-forming tissues reverse the IAA distribution patterns in cambium and the secondary xylem side and therefore result in a wide IAA concentration gradient in radial sections. The pattern change in IAA spatial gradient distribution in tree trunks in turn provides a basic cue for the expression of genes related to auxin signaling, the genetic control of cambial activity and the biosynthesis and catabolism of other plant hormones, according to their locations in wood-forming tissues and their response to auxin signals, thereby forming a physiological and molecular microenvironment that facilitates cambial cell amplification and differentiation toward the xylem cell type.
Supplementary Materials
The following supporting information can be downloaded at: https://0-www-mdpi-com.brum.beds.ac.uk/article/10.3390/f13101698/s1, Figure S1: Tissue-specific expression of genes in Chinese fir wood-forming tissues; Figure S2: Expression profiles of genes related to ethylene biosynthesis and signaling in differently treated samples; Table S1: Functional annotation and classification of selected genes.
Author Contributions
L.Y. was responsible for the design of this research, exogenous IAA treatments, experimental material collection, anatomical analysis, gene expression analysis et al. S.Z. was responsible for the measurement of endogenous hormones and the statical analysis of hormone concentration changes among differently treated samples. All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
All data are included in the article.
Acknowledgments
We appreciated directions and help from Jisen Shi, Jin Xu, Mengli Xi, Nanjing Forestry University, China.
Conflicts of Interest
The authors declare no conflict of interest.
Abbreviations
AUX1/LAX, AUXIN-RESISTANT1 (AUX1)/LIKE AUX1; ARF, AUXIN RESPONSE FACTOR; AFB, AUXIN SIGNALING F-BOX protein; ACS, 1-AMINOCYCLOPROPANE-1-CARBOXYLATE SYNTHASE; AHK4, ARABIDOPSIS HISTONE KINASE 4; AHK3, ARABIDOPSIS HISTONE KINASE 3; ARR, ARABIDOPSIS RESPONSE REGULATOR; ABA1, ABA-DEFICIENT 1; ABF1, ABSCISIC ACID-RESPONSIVE ELEMENT-BINDING FACOR 1; CKX, CYTOKININ OXIDASE; CRE1, CYTOKININ RESPONSE 1; CLE/TDIF, CLAVATA3/ESR-RELATED/TRACHEARY ELEMENT DIFFERENTIATION INHIBITORY FACTOR; EIN3, ETHYLENE-INSENSITIVE 3; EIL1, EIN3-LIKE 1; EBF1, EIN3-BINDING F BOX PROTEIN 1; GA3, GA REQUIRING 3; GA2ox, GA2-oxidase; GAI, GA INSENSITIVE I; HB8, HOMEOBOX 8; HD-ZIP, HOMEODOMAIN LEUCINE ZIPPER; IPT, ISOPENTENYL TRANSFERASE; KAN, KANAD I; MP, MONOPTEROS; NCED, NIEN-CIS-EPOXYCAROTENOID DIOXYGENASE; PIN, PIN-FORMED; PXY/TDR, PHLOEM INTERCALATED WITH XYLEM/TDIF RECEPTOR; RGL2, REPRESSOR OF GA (RGA) LIKE 2; REV, REVOLUTA; TIR1, TRANSPORT INHIBITOR RESPONSE 1; WOX4, WUSCHEL-related HOMEOBOX 4; WOX13, WUSCHEL-related HOMEOBOX 13; ZR, zeatin ribosome; cZ, cis-zeatin; tZ, trans-zeatin.
References
- Plomion, C.; Leprovost, G.; Stokes, A. Wood formation in trees. Plant Physiol. 2001, 127, 1513–1523. [Google Scholar] [CrossRef]
- Uggla, C.; Moritz, T.; Sandberg, G.; Sundberg, B. Auxin as a positional signal in pattern formation in plants. Proc. Natl. Acad. Sci. USA 1996, 93, 9282–9286. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sundberg, B.; Uggla, C.; Tuominen, H. Cambial Growth and Auxin Gradients. In Cell and Molecular Biology of Wood Formation; Savidge, R.A., Barnett, J.R., Napier, R., Eds.; BIOS Scientific: Oxford, UK, 2000; pp. 169–188. [Google Scholar]
- Sundberg, B.; Uggla, C. Origin and dynamics of indoleacetic acid under polar transport in Pinus sylvestris. Physiol. Plant. 1998, 104, 22–29. [Google Scholar] [CrossRef]
- Little, C.; Savidge, R. The role of plant growth regulators in forest tree cambial growth. In Hormonal Control of Tree growth; Springer: Berlin/Heidelberg, Germany, 1987; pp. 137–169. [Google Scholar]
- Tuominen, H.; Puech, L.; Fink, S.; Sundberg, B. A Radial Concentration Gradient of Indole-3-Acetic Acid Is Related to Secondary Xylem Development in Hybrid Aspen. Plant Physiol. 1997, 115, 577–585. [Google Scholar] [CrossRef] [Green Version]
- Nilsson, J.; Karlberg, A.; Antti, H.; Lopez-Vernaza, M.; Mellerowicz, E. Dissecting the molecular basis of the regulation of wood formation by auxin in hybrid aspen. Plant Cell 2008, 20, 843–855. [Google Scholar] [CrossRef] [Green Version]
- Zhang, J.; Nieminen, K.; Serra, J.A.; Helariutta, Y. The formation of wood and its control. Curr. Opin. Plant Biol. 2014, 17, 56–63. [Google Scholar] [CrossRef]
- Tanaka, H.; Dhonukshe, P.; Brewer, P.B.; Friml, J. Spatiotemporal asymmetric auxin distribution: A means to coordinate plant development. Cell. Mol. Life Sci. 2006, 63, 2738–2754. [Google Scholar] [CrossRef]
- Michniewicz, M.; Brewer, P.B.; Friml, J. Polar Auxin Transport and Asymmetric Auxin Distribution. Arabidopsis Book 2007, 5, e0108. [Google Scholar]
- Friml, J.; Palme, K. Polar auxin transport—Old questions and new concepts? Plant Mol. Biol. 2002, 49, 273–284. [Google Scholar] [CrossRef]
- Kramer, E.M. PIN and AUX/LAX proteins: Their role in auxin accumulation. Trends Plant Sci. 2004, 9, 578–582. [Google Scholar] [CrossRef]
- Goldsmith, M. The Polar Transport of Auxin. Annu. Rev. Plant Physiol. 1977, 28, 439–478. [Google Scholar] [CrossRef]
- Zaímalová, E.; Keek, P.; Skpa, P.; Hoyerová, K.; Petráek, J. Polar transport of the plant hormone auxin–the role of PIN-FORMED (PIN) proteins. Cell. Mol. Life Sci. Cmls 2007, 64, 1621–1637. [Google Scholar] [CrossRef]
- Mazur, E.; Gallei, M.; Adamowski, M.; Han, H.; Robert, H.S.; Friml, J. Clathrin-mediated trafficking and PIN trafficking are required for auxin canalization and vascular tissue formation in Arabidopsis. Plant Sci. 2020, 293, 110414. [Google Scholar] [CrossRef]
- Galweiler, L.; Guan, C.; Muller, A.; Wisman, E.; Mendgen, K.; Yephremov, A.; Palme, K. Regulation of Polar Auxin Transport by AtPIN1 in Arabidopsis Vascular Tissue. Science 1998, 282, 2226–2230. [Google Scholar] [CrossRef] [Green Version]
- Schrader, J.; Bhalerao, R.P.; May, S.T.; Palme, K.; Bennett, M.J.; Sandberg, G. “Polar Auxin Transport in Hybrid Aspen”. Aspen Bibliography. Paper 969. 1999. Available online: https://digitalcommons.usu.edu/aspen_bib/969 (accessed on 18 September 2022).
- Hellgren, J.M.; Olofsson, K.; Sundberg, B. Patterns of auxin distribution during gravitational induction of reaction wood in poplar and pine. Plant Physiol. 2004, 135, 212–220. [Google Scholar] [CrossRef] [Green Version]
- Quint, M.; Gray, W.M. Auxin signaling. Curr. Opin. Plant Biol. 2006, 9, 448–453. [Google Scholar] [CrossRef]
- Brackmann, K.; Qi, J.; Gebert, M.; Jouannet, V.; Schlamp, T.; Grünwald, K.; Wallner, E.S.; Novikova, D.D.; Levitsky, V.G.; Agustí, J.; et al. Spatial specificity of auxin responses coordinates wood formation. Nat. Commun. 2018, 9, 875. [Google Scholar] [CrossRef] [Green Version]
- Hirakawa, Y.; Kondo, Y.; Fukuda, H. TDIF peptide signaling regulates vascular stem cell proliferation via the WOX4 homeobox gene in Arabidopsis. Plant Cell 2010, 22, 2618–2629. [Google Scholar] [CrossRef] [Green Version]
- Ji, J.; Strable, J.; Shimizu, R.; Koenig, D.; Sinha, N.; Scanlon, M.J. WOX4 promotes procambial development. Plant Physiol. 2010, 152, 1346–1356. [Google Scholar] [CrossRef] [Green Version]
- Suer, S.; Agusti, J.; Sanchez, P.; Schwarz, M.; Greb, T. WOX4 imparts auxin responsiveness to cambium cells in Arabidopsis. Plant Cell 2011, 23, 3247–3259. [Google Scholar] [CrossRef] [Green Version]
- Etchells, J.P.; Provost, C.M.; Mishra, L.; Turner, S.R. WOX4 and WOX14 act downstream of the PXY receptor kinase to regulate plant vascular proliferation independently of any role in vascular organisation. Development 2013, 140, 2224–2234. [Google Scholar] [CrossRef]
- Donner, T.J.; Sherr, I.; Scarpella, E. Regulation of preprocambial cell state acquisition by auxin signaling in Arabidopsis leaves. Development 2009, 136, 3235–3246. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jouannet, V.; Brackmann, K.; Greb, T. (Pro) cambium formation and proliferation: Two sides of the same coin? Curr. Opin. Plant Biol. 2015, 23, 54–60. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Müller, C.J.; Valdés, A.E.; Wang, G.; Ramachandran, P.; Beste, L.; Uddenberg, D.; Carlsbecker, A. PHABULOSA mediates an auxin signaling loop to regulate vascular patterning in Arabidopsis. Plant Physiol. 2016, 170, 956–970. [Google Scholar] [CrossRef] [Green Version]
- Mishra, L.S. The role of the PXY-CLE signalling pathway in regulating cell division during wood formation in Poplar. Ph.D. Dissertation, The University of Manchester, Manchester, UK, 2016. [Google Scholar]
- Li, J.; Jia, H.; Zhang, J.; Liu, B.; Lu, M. Effect of Overexpression of Populus tomentosa WUSCHEL-related homeobox 4 (PtoWOX4a) on the Secondary Growth of Poplar. Sci. Silvae Sin. 2018, 54, 52–59. [Google Scholar]
- Johnson, L.A. An Investigation into the Molecular Basis of Secondary Vascular Tissue Formation in Poplar and Arabidopsis with an Emphasis on the Role of Auxin and the Auxin Response Factor MONOPTEROS. Ph.D. Dissertation, University of British Columbia, Vancouver, BC, Canada, 2006. [Google Scholar]
- Björklund, S. Plant Hormones in wood formation: A novel insight into the roles of ethylene and gibberellins. Ph.D. Thesis, Swedish University of Agricultural Sciences, Umea, Sweden, 2007; pp. 19–22. [Google Scholar]
- Buttò, V.; Deslauriers, A.; Rossi, S.; Rozenberg, P.; Morin, H. The role of plant hormones in tree-ring formation. Trees 2020, 34, 315–335. [Google Scholar] [CrossRef]
- Fu, X.; Su, H.; Liu, S.; Du, X.; Xu, C.; Luo, K. Cytokinin signaling localized in phloem noncell-autonomously regulates cambial activity during secondary growth of Populus stems. New Phytol. 2021, 230, 1476–1488. [Google Scholar] [CrossRef]
- Immanen, J.; Nieminen, K.; Smolander, O.P.; Kojima, M.; Serra, J.A.; Koskinen, P.; Zhang, J.; Elo, A.; Mähönen, A.P.; Street, N.; et al. Cytokinin and auxin display distinct but interconnected distribution and signaling profiles to stimulate cambial activity. Curr. Biol. 2016, 26, 1990–1997. [Google Scholar] [CrossRef] [Green Version]
- Nieminen, K.; Immanen, J.; Laxell, M.; Kauppinen, L.; Tarkowski, P.; Dolezal, K.; Tähtiharju, S.; Elo, A.; Decourteix, M.; Ljung, K.; et al. Cytokinin signaling regulates cambial development in poplar. Proc. Natl. Acad. Sci. USA 2008, 105, 20032–20037. [Google Scholar] [CrossRef] [Green Version]
- Ioio, R.D.; Nakamura, K.; Moubayidin, L.; Perilli, S.; Taniguchi, M.; Morita, M.T.; Aoyama, T.; Costantino, P.; Sabatini, S. A genetic framework for the control of cell division and differentiation in the root meristem. Science 2008, 322, 1380–1384. [Google Scholar] [CrossRef] [Green Version]
- Müller, B.; Sheen, J. Cytokinin and auxin interaction in root stem-cell specification during early embryogenesis. Nature 2008, 453, 1094–1097. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Eriksson, M.E.; Israelsson, M.; Olsson, O.; Moritz, T. Increased gibberellin biosynthesis in transgenic trees promotes growth, biomass production and xylem fiber length. Nat. Biotechnol. 2000, 18, 784–788. [Google Scholar] [CrossRef] [PubMed]
- Björklund, S.; Antti, H.; Uddestrand, I.; Moritz, T.; Sundberg, B. Cross-talk between gibberellin and auxin in development of Populus wood: Gibberellin stimulates polar auxin transport and has a common transcriptome with auxin. Plant J. 2007, 52, 499–511. [Google Scholar] [CrossRef] [PubMed]
- Klintborg, A.; Eklund, L.; Little, C.A. Ethylene metabolism in Scots pine (Pinus sylvestris) shoots during the year. Tree Physiol. 2002, 22, 59–66. [Google Scholar] [CrossRef] [Green Version]
- Muday, G.K.; Rahman, A.; Binder, B.M. Auxin and ethylene: Collaborators or competitors? Trends Plant Sci. 2012, 17, 181–195. [Google Scholar] [CrossRef]
- Hellgren, J.M. Ethylene and auxin in the control of wood formation. Ph.D. Thesis, Department of Forest, Genetics and Plant Physiology, Swedish University of Agricultural Sciences, Umea, Sweden, 2003. [Google Scholar]
- Junghans, U.; Langenfeld-Heyser, R.; Polle, A.; Teichmann, T. Effect of auxin transport inhibitors and ethylene on the wood anatomy of poplar. Plant Biol. 2004, 6, 22–29. [Google Scholar]
- Thomashow, M.F. Plant cold acclimation: Freezing tolerance genes and regulatory mechanisms. Annu. Rev. Plant Biol. 1999, 50, 571–599. [Google Scholar] [CrossRef] [Green Version]
- Suzuki, N. Hormone signaling pathways under stress combinations. Plant Signal. Behav. 2016, 11, e1247139. [Google Scholar] [CrossRef] [Green Version]
- Suzuki, N.; Bassil, E.; Hamilton, J.S.; Inupakutika, M.A.; Zandalinas, S.I.; Tripathy, D.; Luo, Y.; Dion, E.; Fukui, G.; Kumazaki, A.; et al. ABA is required for plant acclimation to a combination of salt and heat stress. PLoS ONE 2016, 11, e0147625. [Google Scholar] [CrossRef] [Green Version]
- Baba, K.; Karlberg, A.; Schmidt, J.; Schrader, J.; Hvidsten, T.R.; Bako, L.; Bhalerao, R.P. Activity-dormancy transition in the cambial meristem involves stage-specific modulation of auxin response in hybrid aspen. Proc. Natl. Acad. Sci. USA 2011, 108, 3418–3423. [Google Scholar] [CrossRef] [Green Version]
- Hou, H.W.; Zhou, Y.T.; Mwange, K.N.; Li, W.F.; He, X.Q.; Cui, K.M. ABP1 expression regulated by IAA and ABA is associated with the cambium periodicity in Eucommia ulmoides Oliv. J. Exp. Bot. 2006, 57, 3857–3867. [Google Scholar] [CrossRef] [Green Version]
- Hansen, H.; Grossmann, K. Auxin-induced ethylene triggers abscisic acid biosynthesis and growth inhibition. Plant Physiol. 2000, 124, 1437–1448. [Google Scholar] [CrossRef] [Green Version]
- Huang, H.H.; Xu, L.L.; Tong, Z.K.; Lin, E.P.; Liu, Q.P.; Cheng, L.J.; Zhu, M.Y. De novo characterization of the Chinese fir (Cunninghamia lanceolata) transcriptome and analysis of candidate genes involved in cellulose and lignin biosynthesis. BMC Genom. 2012, 13, 648. [Google Scholar] [CrossRef]
- Takei, K.; Yamaya, T.; Sakakibara, H. Arabidopsis CYP735A1 and CYP735A2 Encode Cytokinin Hydroxylases That Catalyze the Biosynthesis of trans-Zeatin. J. Biol. Chem. 2004, 279, 41866–41872. [Google Scholar] [CrossRef] [Green Version]
- Frebort, I.; Kowalska, M.; Hluska, T.; Frebortova, J.; Galuszka, P. Evolution of cytokinin biosynthesis and degradation. J. Exp. Bot. 2011, 62, 2431–2452. [Google Scholar] [CrossRef] [PubMed]
- Hwang, I.; Sheen, J.; Muller, B. Cytokinin signaling networks. Annu. Rev. Plant Biol. 2012, 63, 353–380. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Romanov, G.A.; Lomin, S.N.; Schmulling, T. Cytokinin signaling: From the ER or from the PM? That is the question! New Phytol. 2018, 218, 41–53. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Helliwell, C.A.; Sheldon, C.C.; Olive, M.R.; Walker, A.R.; Zeevaart, J.A.; Peacock, W.J.; Dennis, E.S. Cloning of the Arabidopsis ent-kaurene oxidase gene GA3. Proc. Natl. Acad. Sci. USA 1998, 95, 9019–9024. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yamaguchi, S. Gibberellin metabolism and its regulation. Annu. Rev. Plant Biol. 2008, 59, 225–251. [Google Scholar] [CrossRef]
- Daviere, J.M.; Achard, P. Gibberellin signaling in plants. Development 2013, 140, 1147–1151. [Google Scholar] [CrossRef] [Green Version]
- Dong, T.; Park, Y.; Hwang, I. Abscisic acid: Biosynthesis, inactivation, homoeostasis and signalling. Essays Biochem. 2015, 58, 29–48. [Google Scholar] [PubMed]
- Raghavendra, A.S.; Gonugunta, V.K.; Christmann, A.; Grill, E. ABA perception and signalling. Trends Plant Sci. 2010, 15, 395–401. [Google Scholar] [CrossRef] [PubMed]
- Etchells, J.P.; Turner, S.R. The PXY-CLE41 receptor ligand pair defines a multifunctional pathway that controls the rate and orientation of vascular cell division. Development 2010, 137, 767–774. [Google Scholar] [CrossRef]
- Qiang, Y.; Wu, J.; Han, H.; Wang, G. CLE peptides in vascular development. J Integr Plant Biol. 2013, 55, 389–394. [Google Scholar] [CrossRef] [PubMed]
- Dolzblasz, A.; Nardmann, J.; Clerici, E.; Causier, B.; van der Graaff, E.; Chen, J.; Davies, B.; Werr, W.; Laux, T. Stem Cell Regulation by Arabidopsis WOX Genes. Mol. Plant 2016, 9, 1028–1039. [Google Scholar] [CrossRef]
- Sieburth, L.E.; Deyholos, M.K. Vascular development: The long and winding road. Curr. Opin. Plant Biol. 2006, 9, 48–54. [Google Scholar] [CrossRef]
- Baima, S.; Possenti, M.; Matteucci, A.; Wisman, E.; Altamura, M.M.; Ruberti, I.; Morelli, G. The arabidopsis ATHB-8 HD-zip protein acts as a differentiation-promoting transcription factor of the vascular meristems. Plant Physiol. 2001, 126, 643–655. [Google Scholar] [CrossRef] [Green Version]
- Zhao, C.; Craig, J.C.; Petzold, H.E.; Dickerman, A.W.; Beers, E.P. The xylem and phloem transcriptomes from secondary tissues of the Arabidopsis root-hypocotyl. Plant Physiol. 2005, 138, 803–818. [Google Scholar] [CrossRef] [Green Version]
- Ilegems, M.; Douet, V.; Meylan-Bettex, M.; Uyttewaal, M.; Brand, L.; Bowman, J.L.; Stieger, P.A. Interplay of auxin, KANADI and Class III HD-ZIP transcription factors in vascular tissue formation. Development 2010, 137, 975–984. [Google Scholar] [CrossRef]
Figure 1.
Fluctuation patterns of endogenous hormones in differently treated shoot samples. These curves were drawn according to the mean value of endogenous hormone concentrations in three biological replicates and error bars represented ± SD.
Figure 1.
Fluctuation patterns of endogenous hormones in differently treated shoot samples. These curves were drawn according to the mean value of endogenous hormone concentrations in three biological replicates and error bars represented ± SD.

Figure 2.
Exogenous IAA-induced changes in secondary growth in Chinese fir shoots. (A), Radial anatomical structures in shoots of different samples, in which the red arrow represented the radial thickening growth at specific treatment stages. The scale bar was 1:10 μm. SP, secondary phloem; Cam, cambium; SX, secondary xylem. (B), Patterns of secondary growth fluctuation in different samples, which was drawn according to the mean value ±SD of radial thickening growth measured from three different directions in the radial section. (C), Multiple comparisons of secondary growth among different treatment stages of shoot samples.
Figure 2.
Exogenous IAA-induced changes in secondary growth in Chinese fir shoots. (A), Radial anatomical structures in shoots of different samples, in which the red arrow represented the radial thickening growth at specific treatment stages. The scale bar was 1:10 μm. SP, secondary phloem; Cam, cambium; SX, secondary xylem. (B), Patterns of secondary growth fluctuation in different samples, which was drawn according to the mean value ±SD of radial thickening growth measured from three different directions in the radial section. (C), Multiple comparisons of secondary growth among different treatment stages of shoot samples.

Figure 3.
Expression profiles of AUX1/LAX and PIN family genes in differently treated samples. Three biological replications were included. The reference gene was ubiquitin 10 (UBQ10). a, b, c and d represented the difference in gene expression levels in the samples treated by exogenous IAA compared to those in PC. a, p-value < 0.001; b, p-value < 0.01; c, p-value < 0.05; d, p-value > 0.05. Similar parameter settings were performed in the following figures.
Figure 3.
Expression profiles of AUX1/LAX and PIN family genes in differently treated samples. Three biological replications were included. The reference gene was ubiquitin 10 (UBQ10). a, b, c and d represented the difference in gene expression levels in the samples treated by exogenous IAA compared to those in PC. a, p-value < 0.001; b, p-value < 0.01; c, p-value < 0.05; d, p-value > 0.05. Similar parameter settings were performed in the following figures.

Figure 4.
Expression profiles of AUX/IAAs and ARFs in differently treated samples. a, p-value < 0.001; b, p-value < 0.01; c, p-value < 0.05; d, p-value > 0.05.
Figure 4.
Expression profiles of AUX/IAAs and ARFs in differently treated samples. a, p-value < 0.001; b, p-value < 0.01; c, p-value < 0.05; d, p-value > 0.05.

Figure 5.
Expression patterns of CK synthesis—and signaling-related genes in differently treated samples. a, p-value < 0.001; b, p-value < 0.01; c, p-value < 0.05; d, p-value > 0.05.
Figure 5.
Expression patterns of CK synthesis—and signaling-related genes in differently treated samples. a, p-value < 0.001; b, p-value < 0.01; c, p-value < 0.05; d, p-value > 0.05.

Figure 6.
Expression patterns of genes participating in GA production and signaling transduction in differently treated samples. a, p-value < 0.001; b, p-value < 0.01; c, p-value < 0.05; d, p-value > 0.05.
Figure 6.
Expression patterns of genes participating in GA production and signaling transduction in differently treated samples. a, p-value < 0.001; b, p-value < 0.01; c, p-value < 0.05; d, p-value > 0.05.

Figure 7.
Expression profiles of genes participating in ABA production and signal transduction in differently treated samples. a, p-value < 0.001; b, p-value < 0.01; c, p-value < 0.05; d, p-value > 0.05.
Figure 7.
Expression profiles of genes participating in ABA production and signal transduction in differently treated samples. a, p-value < 0.001; b, p-value < 0.01; c, p-value < 0.05; d, p-value > 0.05.

Figure 8.
Expression profiles of genetic regulators in differently treated samples. a, p-value < 0.001; b, p-value < 0.01; c, p-value < 0.05; d, p-value > 0.05.
Figure 8.
Expression profiles of genetic regulators in differently treated samples. a, p-value < 0.001; b, p-value < 0.01; c, p-value < 0.05; d, p-value > 0.05.

Figure 9.
A proposed model whereby the auxin concentration gradient across tree trunks is involved in the control of cambial activity. Sp, secondary phloem; Cam, cambium; Sx, secondary xylem.
Figure 9.
A proposed model whereby the auxin concentration gradient across tree trunks is involved in the control of cambial activity. Sp, secondary phloem; Cam, cambium; Sx, secondary xylem.


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
Variance analysis of hormone concentrations in differently treated stages of samples.
| Hormone | Variance Source | Sum of Squares | df | Mean Square | F-Value | p-Value |
|---|---|---|---|---|---|---|
| IAA | Treatments | 7425.00 | 3 | 2475.00 | 1934.00 | <0.001 |
| Treatment stages | 22,568.00 | 6 | 3761.00 | 2940.00 | <0.001 | |
| Interaction | 25,613.00 | 18 | 1423.00 | 1112.00 | <0.001 | |
| Error | 72 | 56 | 1.29 | |||
| Total | ||||||
| ZR | Treatments | 659.00 | 3 | 219.80 | 5261.00 | <0.001 |
| Treatment stages | 2375.00 | 6 | 395.90 | 9474.00 | <0.001 | |
| Interaction | 4638.00 | 18 | 257.70 | 6167.00 | <0.001 | |
| Error | 2.00 | 56 | 0.04 | |||
| Total | ||||||
| GA3 | Treatments | 23.50 | 3 | 7.84 | 319.60 | <0.001 |
| Treatment stages | 548.10 | 6 | 91.35 | 3722.00 | <0.001 | |
| Interaction | 836.50 | 18 | 46.47 | 1893.60 | <0.001 | |
| Error | 1.40 | 56 | 0.02 | |||
| Total | ||||||
| ABA | Treatments | 53,660.00 | 3 | 17,887.00 | 2192.20 | <0.001 |
| Treatment stages | 67,108.00 | 6 | 11,185.00 | 1370.80 | <0.001 | |
| Interaction | 53,403.00 | 18 | 2967.00 | 363.60 | <0.001 | |
| Error | 457.00 | 56 | 8.00 | |||
| Total |
Treatments included application of 0, 1.0 or 3.0 mg.iaa/g.lanolin to the decapitated Chinese fir trunks (these samples were designated NC, S1 and S3, respectively) and plants with intact shoot apexes act as the positive control (PC). Treatment stages included the 2nd, 6th, 8th, 12th, 16th, 24th and 32nd days of treatment by exogenous IAA. Variance analysis was executed with a repeatable two-way ANOVA model.
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Yang, L.; Zhu, S. The Interconnected Relationship between Auxin Concentration Gradient Changes in Chinese Fir Radial Stems and Dynamic Cambial Activity. Forests 2022, 13, 1698. https://0-doi-org.brum.beds.ac.uk/10.3390/f13101698
AMA Style
Yang L, Zhu S. The Interconnected Relationship between Auxin Concentration Gradient Changes in Chinese Fir Radial Stems and Dynamic Cambial Activity. Forests. 2022; 13(10):1698. https://0-doi-org.brum.beds.ac.uk/10.3390/f13101698
Chicago/Turabian StyleYang, Liwei, and Sheng Zhu. 2022. "The Interconnected Relationship between Auxin Concentration Gradient Changes in Chinese Fir Radial Stems and Dynamic Cambial Activity" Forests 13, no. 10: 1698. https://0-doi-org.brum.beds.ac.uk/10.3390/f13101698
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.