

Improvement of Rooting Performance in Stem Cuttings of Savin Juniper (Juniperus sabina L.) as a Function of IBA Pretreatment, Substrate, and Season

 ,
,
Abstract
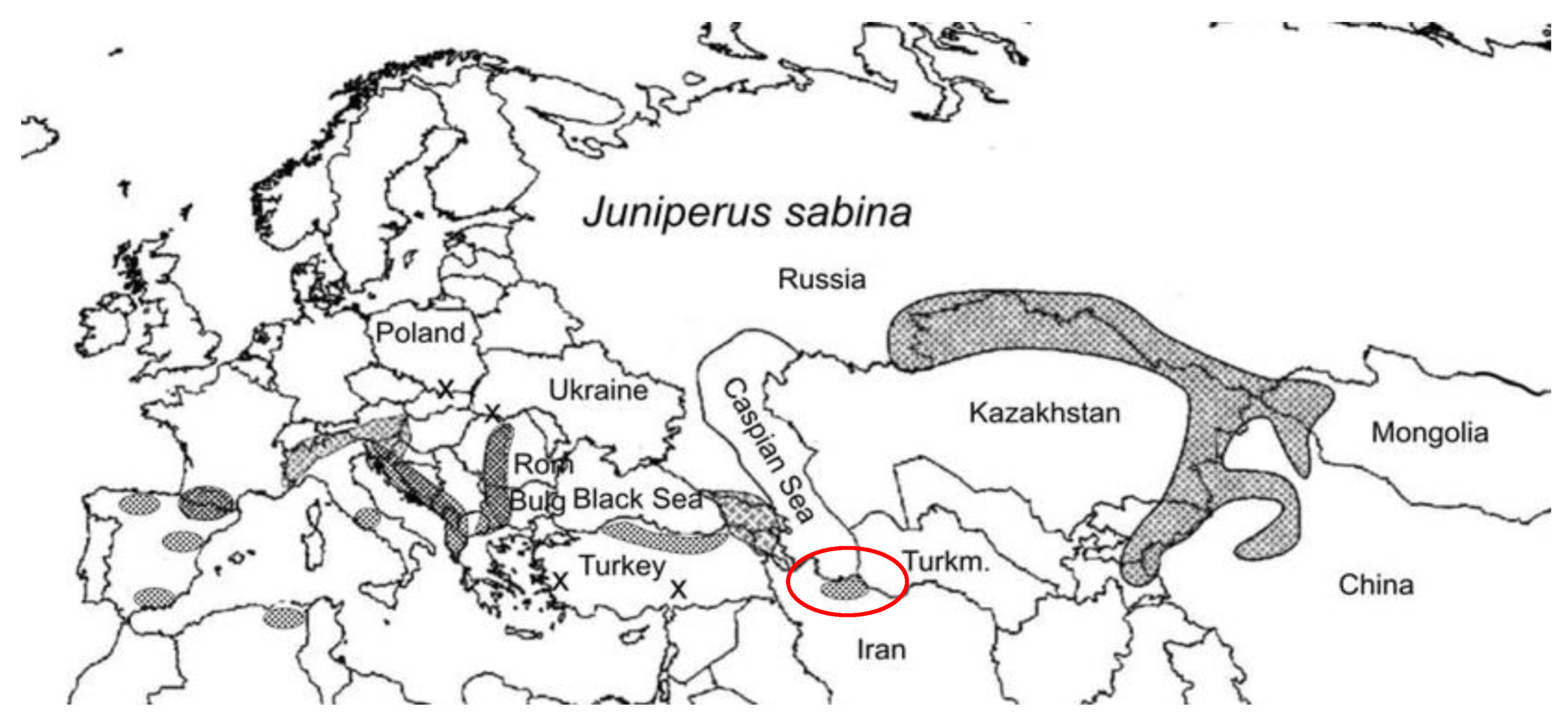
:1. Introduction
2. Materials and Methods
2.1. Cutting Preparation, Pretreatment with Indole Butyric Acid (IBA), and Substrate Composition
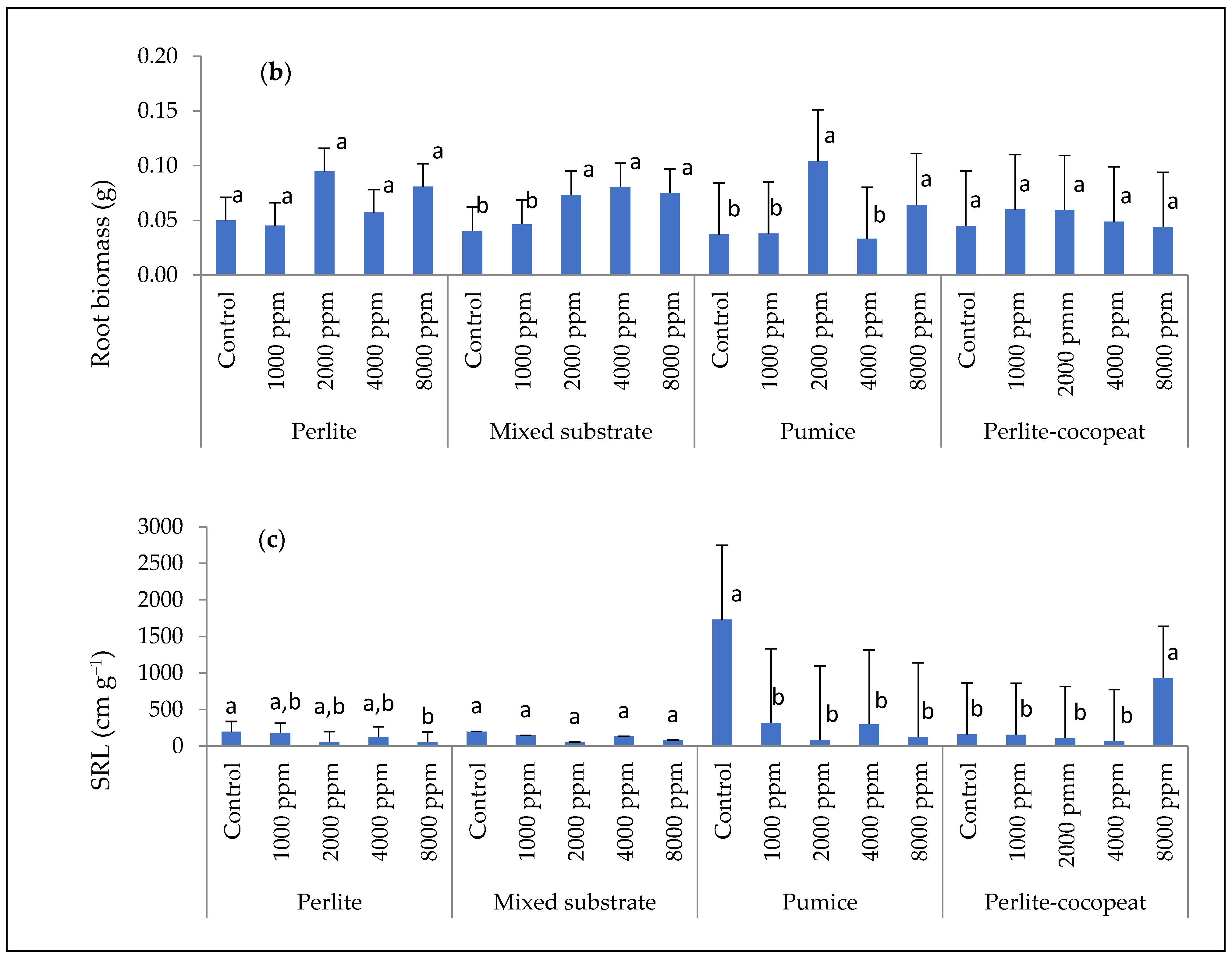
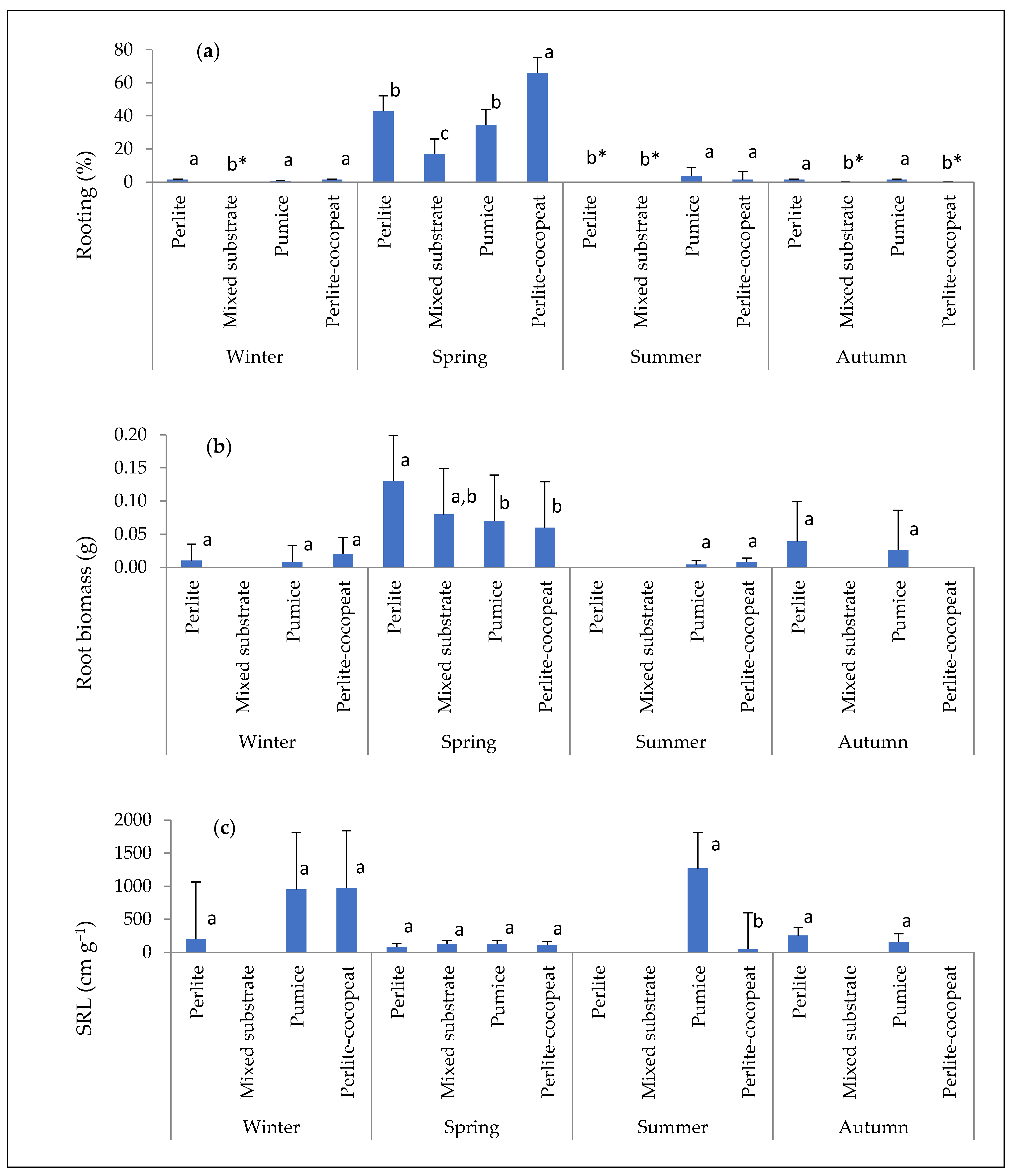
2.2. Rooting Performance: Rooting (%), Root Biomass, and Specific Root Length (SRL)
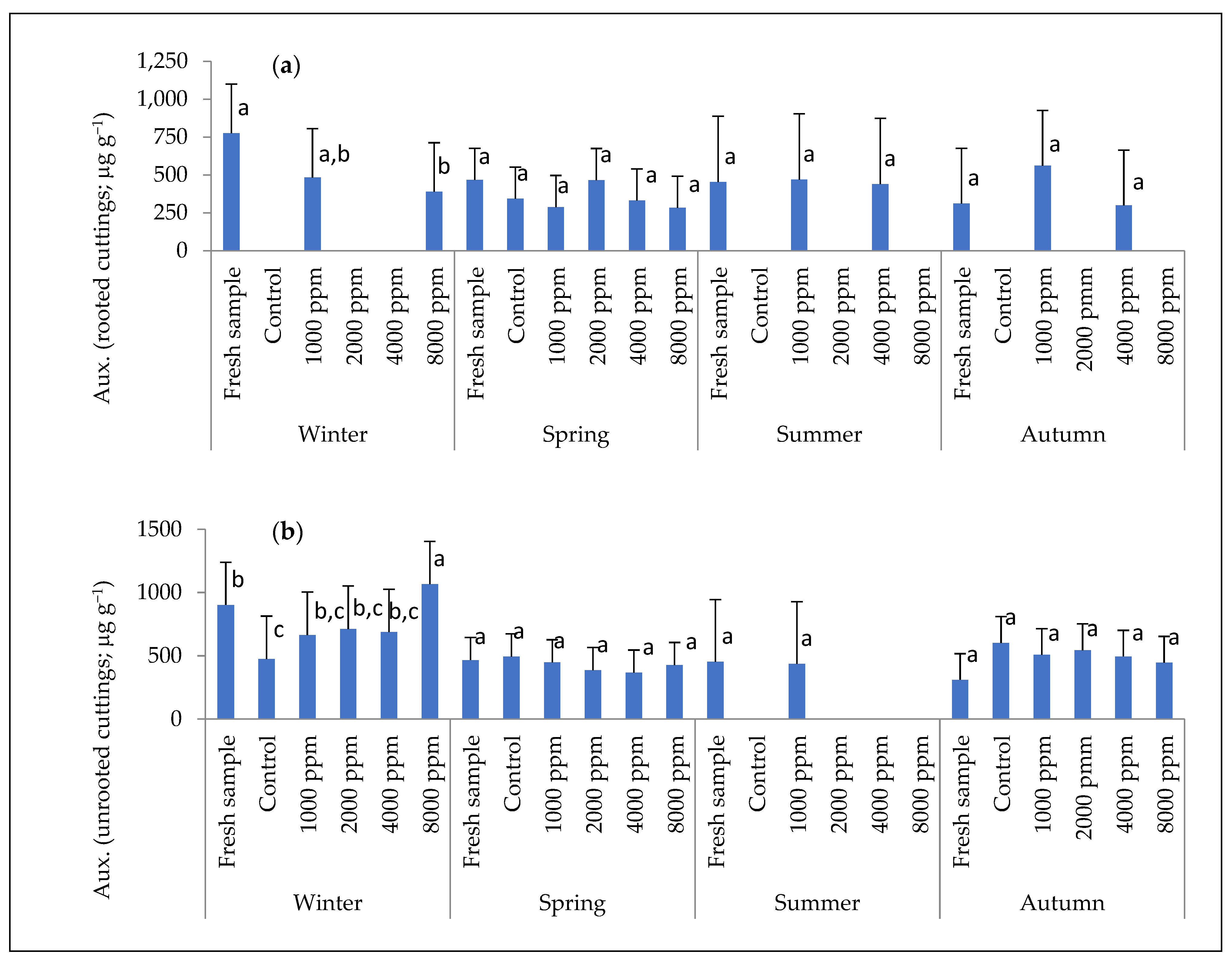
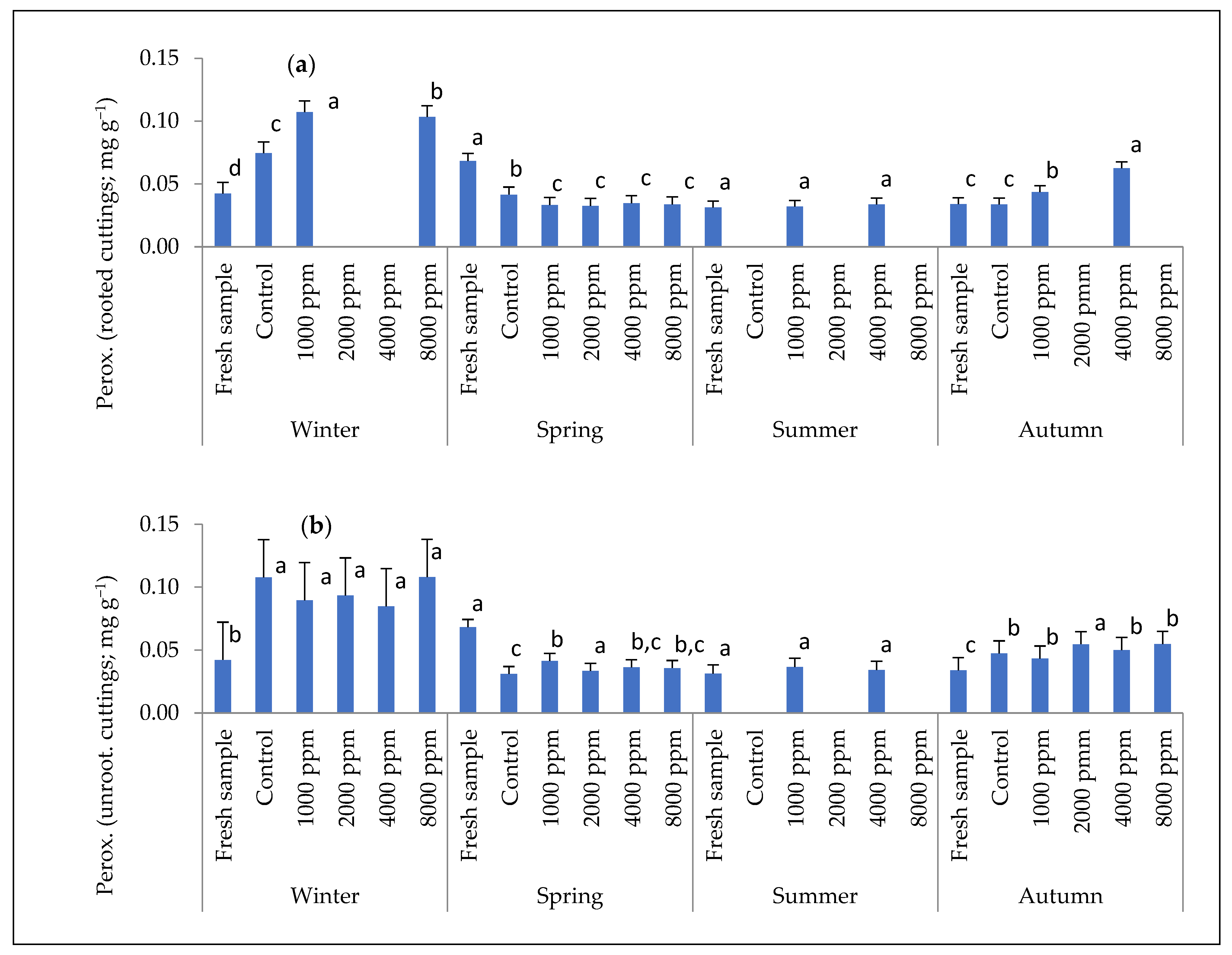
2.3. Internal Chemical Compounds of Stem Cuttings: Auxin and Peroxidase Enzyme Concentration
2.4. Statistical Analysis
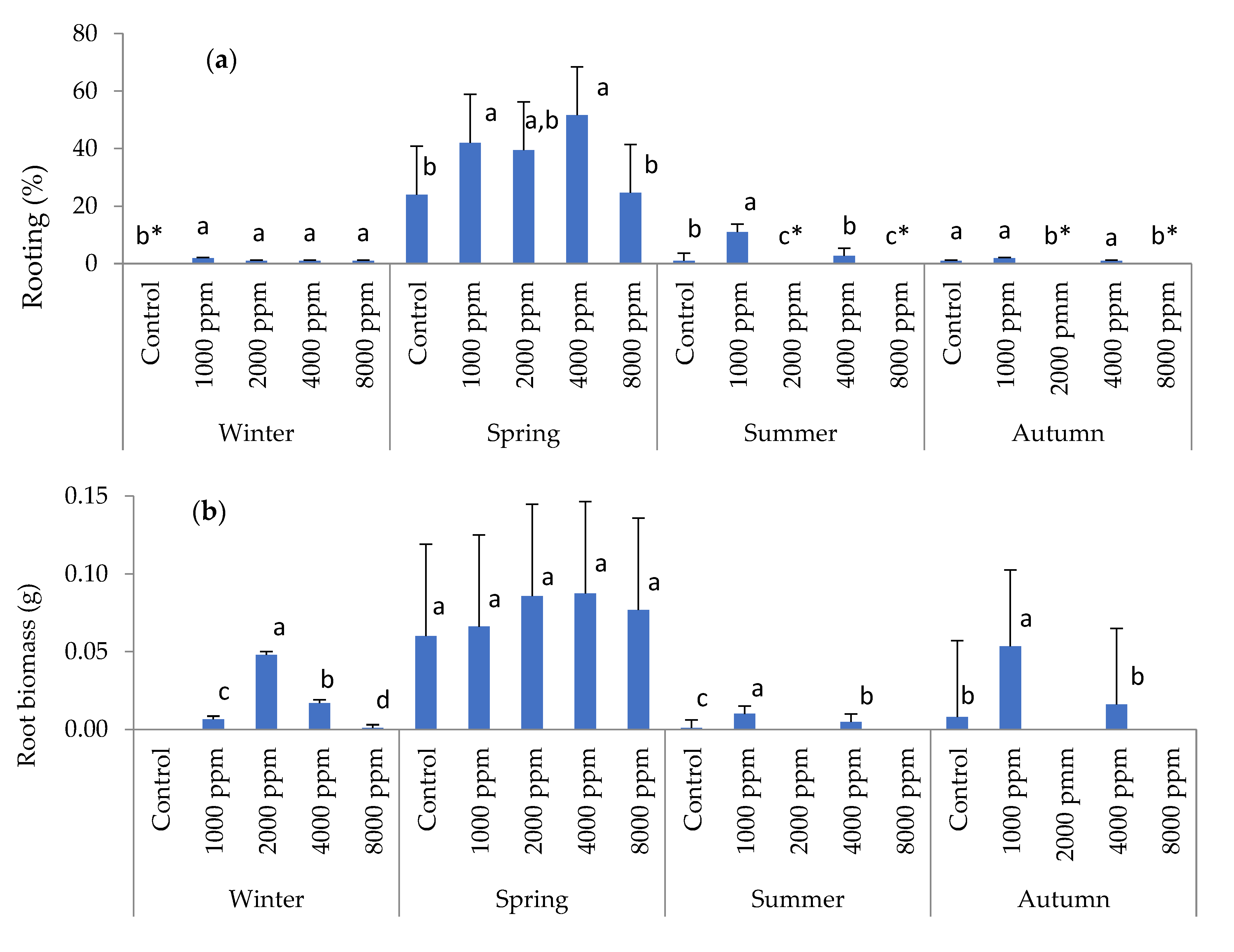
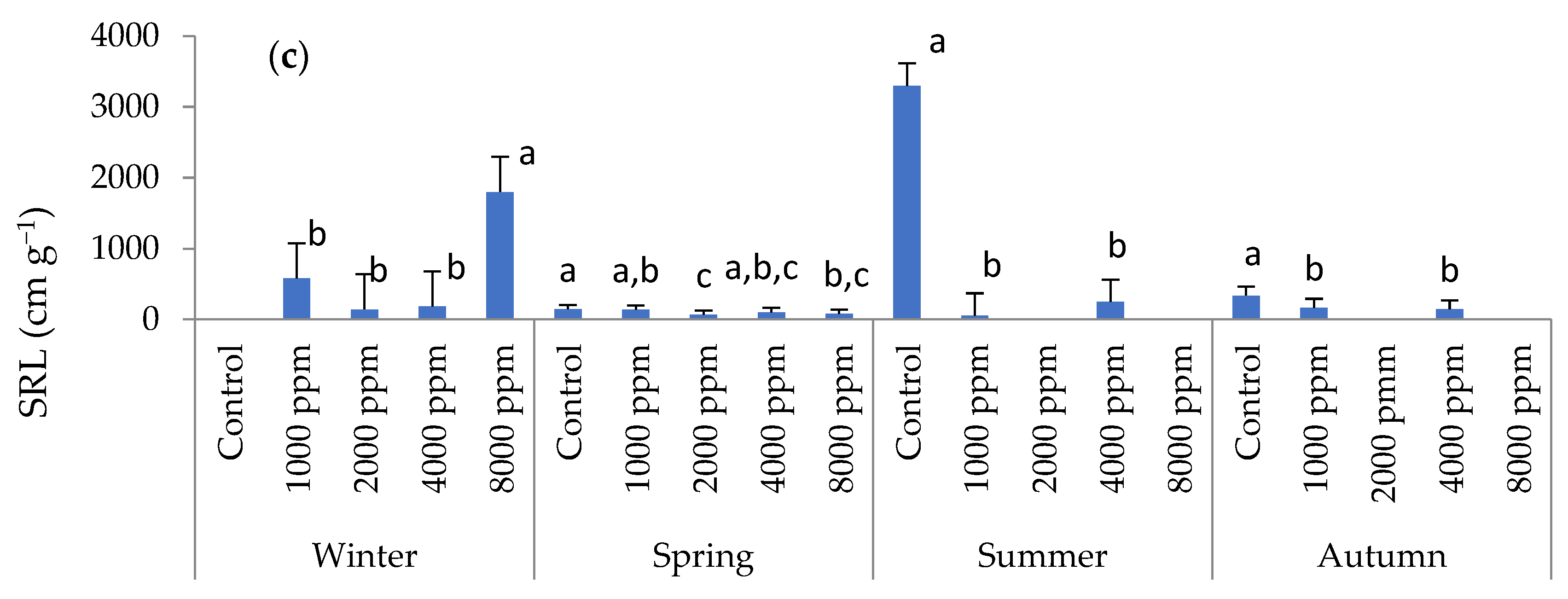
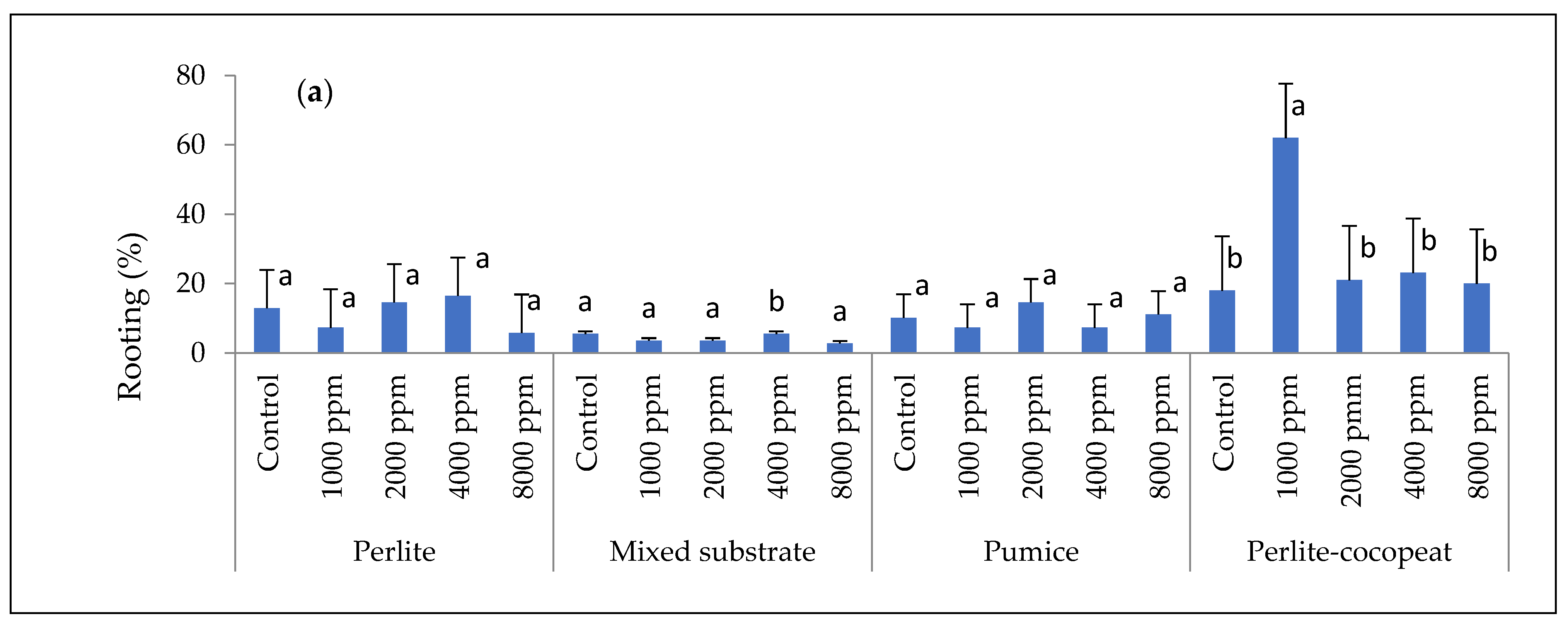
3. Results
3.1. Rooting Performance
3.2. Auxin and Peroxidase Concentration in Cuttings
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Farjon, A. A Monograph of Cupressaceae and Sciadopitys; Royal Botanic Gardens, Kew: Richmond, Surrey, UK, 2005; p. 648. [Google Scholar]
- Adams, R.P. The Junipers of the World: The Genus Juniperus, 4th ed.; Trafford Publishing: Victoria, BC, Canada, 2014; p. 422. [Google Scholar]
- García Morote, F.A.; Andrés Abellán, M.; Rubio, E.; Pérez Anta, I.; García Saucedo, F.; López Serrano, F.R. Stem CO2 Efflux as an Indicator of Forests’ Productivity in Relict Juniper Woodlands (Juniperus thurifera L.) of Southern Spain. Forests 2021, 12, 1340. [Google Scholar] [CrossRef]
- Comino, E.; Marengo, P. Root tensile strength of three shrub species: Rosa canina, Cotoneaster dammeri and Juniperus horizontalis Soil reinforcement estimation by laboratory tests. Catena 2010, 82, 227–235. [Google Scholar] [CrossRef]
- Piotto, B.; Di Noi, A. Seed Propagation of Mediterranean Trees and Shrubs; Agency for the Protection of the Environment and for Technical Services: Roma, Italy, 2003; p. 108.
- Lu, D.; Huang, H.; Wang, A.; Zhang, G. Genetic Evaluation of Juniperus sabina L. (Cupressaceae) in Arid and Semi-Arid Regions of China Based on SSR Markers. Forests 2022, 13, 231. [Google Scholar] [CrossRef]
- Esmaeilnia, M.; Jalali, S.G.; Tabari, M.; Hosseini, S.M. Influence of plant growth regulator IBA on vegetative propagation of Juniperus excelsa. Iran. J. For. Poplar Res. 2006, 14, 221–227. [Google Scholar]
- Momeni, M.; Ganji-Moghadam, E.; Kazemzadeh-Beneh, H.; Asgharzadeh, A. Direct organogenesis from shoot tip explants of Juniperus polycarpos L.: Optimizing basal media and plant growth regulators on proliferation and root formation. Plant Cell Biotechnol. Mol. Biol. 2018, 19, 40–50. [Google Scholar]
- Hazubska-Przybył, T. Propagation of Juniper Species by Plant Tissue Culture: A Mini-Review. Forests 2019, 10, 1028. [Google Scholar] [CrossRef] [Green Version]
- Amri, E.; Lyaruu, H.; Nyomora, A.; Kanyeka, Z. Vegetative propagation of African Blackwood (Dalbergia melanoxylon Guill. & Perr.): Effects of age of donor plant, IBA treatment and cutting position on rooting ability of stem cuttings. New For. 2010, 39, 183–194. [Google Scholar] [CrossRef]
- Nordstrom, A.C.; Jacobs, F.A.; Eliasson, L. Effect of exogenous Indole-3-Acetic-acid and Indole-3-Butyric acid on internal levels of the respective auxins and their conjugation with aspartic-acid during adventitious root-formation in Pea cuttings. Plant Physiol. 1991, 96, 856–861. [Google Scholar] [CrossRef] [Green Version]
- Rifaki, N.; Economou, A.; Hatzilazarou, S. Factors affecting vegetative propagation of Juniperus excelsa bieb. by stem cuttings. Propag. Ornam. Plants 2002, 2, 9–13. [Google Scholar]
- Henry, P.H.; Blazich, F.A.; Hinesley, L.E. Vegetative propagation of Eastern redcedar by stem cuttings. Hortscience 1992, 27, 1272–1274. [Google Scholar] [CrossRef] [Green Version]
- Pacurar, D.; Perrone, I.; Bellini, C. Auxin is a central player in the hormone cross-talks that control adventitious rooting. Physiol. Plant. 2014, 151, 83–96. [Google Scholar] [CrossRef] [PubMed]
- Ludwig-Muller, J. Indole-3-butyric acid in plant growth and development. Plant Growth Regul. 2000, 32, 219–230. [Google Scholar] [CrossRef]
- Daveis, P.J. Plant Hormones: Biosynthesis, Signal Transduction, Action! Kluwer Academic Publishers: Dordrecht, The Netherlands, 2004; p. 802. [Google Scholar]
- Gao, J.; Xiao, Q.; Ding, L.; Chen, M.; Yin, L.; Li, J.; Zhou, S.; He, G. Differential responses of lipid peroxidation and antioxidants in Alternanthera philoxeroides and Oryza sativa subjected to drought stress. Plant Growth Regul. 2008, 56, 89–95. [Google Scholar] [CrossRef]
- Aliahmad Koruri, S.; Khoushnevis, M.; Matinizadeh, M. Comprehensive Studies of Juniper Species in Iran; Publication of Pooneh, Forest & Ranges and Watershed Organization of Iran: Tehran, Iran, 2011; p. 554.
- Kentelky, E. The Analysis of Rooting and Growth Peculiarities of Juniperus Species Propagated by Cuttings. Horticulture 2011, 68, 380–385. [Google Scholar]
- Hong-wei, Y.; Yong-sheng, G.; Hai-jun, S.; Yan-he, R. Experiment on Cutting Propagation of Sabina procumbens. J. Inn. Mong. For. Sci. Technol. 2011, 4, 6. [Google Scholar]
- Sabina, P.; Cornelia, H. Researches concerning the production of planting material using vegetative propagation on Juniperus horizontalis mnch. J. Hortic. For. Biotechnol. 2009, 13, 462–464. [Google Scholar]
- Wendling, I.; Warburton, P.; Trueman, S. Maturation in Corymbia torelliana x C. citriodora Stock Plants: Effects of Pruning Height on Shoot Production, Adventitious Rooting Capacity, Stem Anatomy, and Auxin and Abscisic Acid Concentrations. Forests 2015, 6, 3763–3778. [Google Scholar] [CrossRef] [Green Version]
- Bryant, P.; Trueman, S. Stem Anatomy and Adventitious Root Formation in Cuttings of Angophora, Corymbia and Eucalyptus. Forests 2015, 6, 1227–1238. [Google Scholar] [CrossRef]
- Garcia Morote, F.A.; Lopez Serrano, F.R.; Andres, M.; Rubio, E.; Gonzalez Jimenez, J.L.; de las Heras, J. Allometries, biomass stocks and biomass allocation in the thermophilic Spanish juniper woodlands of Southern Spain. For. Ecol. Manag. 2012, 270, 85–93. [Google Scholar] [CrossRef]
- Paula, S.; Pausas, J. Root traits explain different foraging strategies between resprouting life histories. Oecologia 2011, 165, 321–331. [Google Scholar] [CrossRef] [Green Version]
- Hernandez, E.; Vilagrosa, A.; Pausas, J.; Bellot, J. Morphological traits and water use strategies in seedlings of Mediterranean coexisting species. Plant Ecol. 2010, 207, 233–244. [Google Scholar] [CrossRef] [Green Version]
- Adams, R.P.; Schwarzbach, A.E. Chloroplast capture by a new variety, Juniperus sabina var. balkanensis R. P. Adams and A. N. Tashev, from the Balkan peninsula: A putative stabilized relictual hybrid between J. sabina and ancestral J. thurifera. Phytologia 2016, 98, 100–111. [Google Scholar]
- Kottek, M.; Grieser, J.; Beck, C.; Rudolf, B.; Rubel, F. World Map of the Köppen-Geiger climate classification. Meteorol. Z. 2006, 15, 259–263. [Google Scholar] [CrossRef]
- Bohlenius, H.; Fransson, T.; Holmstrom, E.; Salk, C. Influence of Cutting Type and Fertilization in Production of Containerized Poplar Plants. Forests 2017, 8, 164. [Google Scholar] [CrossRef] [Green Version]
- Judd, L.A.; Jackson, B.E.; Fonteno, W.C. Advancements in Root Growth Measurement Technologies and Observation Capabilities for Container-Grown Plants. Plants 2015, 4, 369–392. [Google Scholar] [CrossRef] [Green Version]
- Sridhar, R.; Mohanty, S.; Anjaneyulu, A. Physiology of rice tungro virus-disease-increased cytokinin activity in tungro-infected rice cultivars. Physiol. Plant. 1978, 43, 363–366. [Google Scholar] [CrossRef]
- Hoshmand, H.R. Design of Experiments for Agriculture and the Natural Sciences, 2nd ed.; Champan & Hall/CRC: New York, NY, USA, 2006; p. 437. [Google Scholar]
- Neter, J.; Kutner, M.; Wasserman, W.; Nachtsheim, C. Applied Linear Statistical Models, 4th ed.; McGraw-Hill-Irwin: Chicago, IL, USA, 1996; p. 720. [Google Scholar]
- Davies, F.T.; Geneve, R.L.; Kester, D.E.; Hartmann, H.T. Techniques of Propagation by Cuttings. In Plant-Propagation-Principles and Practices, 8th ed.; Hartmann, K., Davies, G., Eds.; Pearson Education Limited: Harlow, Essex, UK, 2014; p. 927. [Google Scholar]
- Khoushnevis, M.; Teimouri, M.; Matinizadeh, M.; Shirvany, A. Effects of hormones, light and media treatments on rooting of Juniperus oblonga cuttings. Iran. J. For. 2012, 4, 135–142. [Google Scholar]
- Moncousin, C.; Gaspar, T. Peroxidase as a marker for rooting improvement of Cynara scolymus L. cultured in vitro. Biochem. Physiol. Pflanz. 1983, 178, 263–271. [Google Scholar] [CrossRef]
- Kocer, Z.A.; Gozen, A.G.; Onde, S.; Kaya, Z. Indirect organogenesis from bud explants of Juniperus communis L.: Effects of genotype, gender, sampling time and growth regulator combinations. Dendrobiology 2011, 66, 33–40. [Google Scholar]
- Negash, L. Successful vegetative propagation techniques for the threatened African pencil cedar (Juniperus procera Hoechst. ex Endl.). For. Ecol. Manag. 2002, 161, 53–64. [Google Scholar] [CrossRef]
- Epstein, E.; Ludwig-Muller, J. Indole-3-butyric acid in plants- ocurrence, synthesis, metabolism and transport. Physiol. Plant. 1993, 88, 382–389. [Google Scholar] [CrossRef]
- Poupart, J.; Waddell, C. The rib1 mutant is resistant to indole-3-butyric acid, an endogenous auxin in arabidopsis. Plant Physiol. 2000, 124, 1739–1751. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cope, K.; Rupp, L. Vegetative propagation of Juniperus osteosperma (Utah Juniper) by cuttings. Nativ. Plants J. 2013, 14, 76–84. [Google Scholar] [CrossRef]
- Bielenin, M. Rooting and gas exchange of conifer cuttings treated with indole butyric acid. J. Fruit Ornam. Plant Res. 2003, 11, 99–105. [Google Scholar]
- Tektas, I.; Türkoğlu, N.; Cavusoğlu, S. Effects of auxin doses on rooting of Juniperus L. Prog. Nutr. 2017, 19, 130–136. [Google Scholar]
- Ma, W.; Zhang, S.; Wang, J.; Sun, X.; Zhao, H.; Ning, Y. Endogenous hormones, nutritive material and phenolic acid variation in cuttings of japanese larch during rooting. Acta Bot. Boreali-Occident. Sin. 2013, 1, 19. [Google Scholar]
- Chowdhuri, T.K. Performance evaluation of different growth regulators on propagation of Chinese juniper (Juniperus chinensis Var. pyramidalis) in subtropical zone. J. Pharmacogn. Phytochem. 2017, 6, 2190–2193. [Google Scholar]
- Stuepp, C.A.; Ruffellato-Ribas, K.C.; Macanhão, G.; Fragoso, R.; Rickli, H.C. Rooting of Juniperus chinensis var. kaizuka for different concentrations of IBA and heights collection. Rev. Agrar. 2014, 7, 496–503. [Google Scholar]
- Fragoso, R.; Zuffellato-Ribas, K.; Macanhão, G.; Stuepp, C.A.; Koehler, H. Vegetative propagation of Juniperus chinensis. Comun. Sci. 2015, 6, 307–316. [Google Scholar] [CrossRef] [Green Version]
- Chong, C. Influence of Bottom Heat and Mulch on Rooting of Evergreen Cuttings. Comb. Proc. Int. Plant Propagators Soc. 2003, 53, 496–500. [Google Scholar]
- Cuevas-Cruz, J.; Jimenez-Casas, M.; Jasso-Mata, J.; Perez-Rodriguez, P.; Lopez-Upton, J.; Villegas-Monter, A. Asexual propagation of Pinus leiophylla Schiede ex Schltdl. et Cham. Rev. Chapingo Ser. Cienc. For. Ambiente 2015, 21, 81–95. [Google Scholar] [CrossRef]
- Ayan, S.; Küçük, M.; Ulu, F.; Gerçek, V.; Sahin, A.; Sivacioğlu, A. Vegetative propagation possibilities of some natural Juniper (Juniperus L.) species. J. For. Fac. Gazi Uni. Kast. 2004, 4, 1–12. [Google Scholar]
- Manios, V.I.; Papadimitriou, M.D.; Kefakis, M.D. Hydroponic culture of tomato and gerbera at different substrates. Acta Hortic. 1995, 408, 11–15. [Google Scholar] [CrossRef]
- Maloupa, E.; Samartzidis, P.; Couloumbis, P.; Komnin, A. Yield quality and photosynthetic activity of greenhouse-grown “Madelom” roses on perlite-zeolite substrate mixtures. Acta Hortic. 1999, 481, 97–99. [Google Scholar] [CrossRef]
- Pratt, R.; North, G.; Jacobsen, A.; Ewers, F.; Davis, S. Xylem root and shoot hydraulics is linked to life history type in chaparral seedlings. Funct. Ecol. 2010, 24, 70–81. [Google Scholar] [CrossRef]
- Guerrero-Campo, J.; Palacio, S.; Perez-Rontome, C.; Montserrat-Marti, G. Effect of root system morphology on root-sprouting and shoot-rooting abilities in 123 plant species from eroded lands in north-east Spain. Ann. Bot. 2006, 98, 439–447. [Google Scholar] [CrossRef]
- Luo, J.; Zhou, J.; Zhang, J. Aux/IAA Gene Family in Plants: Molecular Structure, Regulation, and Function. Int. J. Mol. Sci. 2018, 19, 259. [Google Scholar] [CrossRef] [Green Version]
- Blakesley, D.; Weston, G.; Hall, J. The role of endogenous auxin in root initiation. 1. Evidence from studies on auxin application, and analysis of endogenous levels. Plant Growth Regul. 1991, 10, 341–353. [Google Scholar] [CrossRef]
- Hausman, J. Changes in Peroxidase-activity, auxin and ethylene production during root-formation by Poplar shoots raised in-vitro. Plant Growth Regul. 1993, 13, 263–268. [Google Scholar] [CrossRef]
- Rival, A.; Bernard, F.; Mathieu, Y. Changes in peroxidase activity during in vitro rooting of oil palm (Elaeis guineensis Jacq.). Sci. Hortic. 1997, 71, 103–112. [Google Scholar] [CrossRef]
- Yan, S.; Yang, R.; Wang, F.; Sun, L.; Song, X. Effect of Auxins and Associated Metabolic Changes on Cuttings of Hybrid Aspen. Forests 2017, 8, 117. [Google Scholar] [CrossRef]
- Torabian, Y.; Korori, S.; Adeli, A.; Falahchai, M. Quantitative and qualitative study of peroxidase and amylase enzymes of seeds and shoots of Juniper trees (Juniperus excelsa) depending on the quality of the bases in the habitat of Razavi Khorasan. J. Biol. Sci. 2009, 3, 19–27. [Google Scholar]
- Moncousin, C. Rooting of in vitro cuttings. In High-Tech and Micropropagation I; Springer: Berlin/Heidelberg, Germany, 1991; pp. 231–261. [Google Scholar]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Growing Season | Annual Values | |||||
|---|---|---|---|---|---|---|
| Variables | Effects | Winter | Spring | Summer | Autumn | |
| Rooting | Pretreatment | 0.10 | <0.001 | 0.0005 | 0.11 | <0.0001 |
| (log-transformed units) | Substrate | 0.91 | 0.03 | 0.22 | 0.08 | <0.0001 |
| Season | - | - | - | - | <0.0001 | |
| Pretreatment × Substrate | 0.40 | 0.24 | * | * | <0.0001 | |
| Pretreatment × Season | - | - | - | - | <0.0001 | |
| Substrate × Season | - | - | - | - | <0.0001 | |
| Root biomass (g) | Pretreatment | <0.0001 | 0.63 | <0.0001 | 0.04 | 0.39 |
| Substrate | 0.30 | 0.01 | 0.11 | 0.29 | 0.08 | |
| Season | - | - | - | - | <0.0001 | |
| Pretreatment × Substrate | 0.64 | 0.04 | * | * | 0.76 | |
| Pretreatment × Season | - | - | - | - | 0.01 | |
| Substrate × Season | - | - | - | - | 0.87 | |
| SRL (cm g−1) | Pretreatment | <0.0001 | 0.04 | <0.0001 | 0.01 | <0.0001 |
| Substrate | 0.08 | 0.29 | 0.02 | 0.95 | <0.0001 | |
| Season | - | - | - | - | <0.0001 | |
| Pretreatment × Substrate | 0.32 | 0.86 | * | * | 0.93 | |
| Pretreatment × Season | - | - | - | - | <0.0001 | |
| Substrate × Season | - | - | - | - | <0.0001 | |
| Growing Season | Annual Values | |||||
|---|---|---|---|---|---|---|
| Chemical Compound | Effects | Winter | Spring | Summer | Autumn | |
| Auxin (μg g−1) (Rooted cuttings) | Pretreatment | 0.02 | 0.33 | 1.00 | 0.22 | 0.95 |
| Substrate | 0.69 | 0.05 | 0.99 | 0.94 | 0.97 | |
| Season | - | - | - | - | 0.39 | |
| Pretreatment × Substrate | 0.37 | 0.53 | * | 0.98 | 0.99 | |
| Pretreatment × Season | - | - | - | - | 0.69 | |
| Substrate × Season | - | - | - | - | 0.99 | |
| Auxin (μg g−1) (Unrooted cuttings) | Pretreatment | 0.01 | 0.57 | 0.94 | 0.11 | 0.70 |
| Substrate | 0.83 | 0.07 | 0.99 | 0.36 | 0.66 | |
| Season | - | - | - | - | <0.0001 | |
| Pretreatment × Substrate | <0.0001 | 0.38 | * | 0.89 | 0.92 | |
| Pretreatment × Season | - | - | - | - | 0.01 | |
| Substrate × Season | - | - | - | - | 0.99 | |
| Peroxidase (mg g−1) (Rooted cuttings) | Pretreatment | <0.0001 | <0.0001 | 0.73 | <0.0001 | <0.0001 |
| Substrate | 0.67 | 0.43 | 0.71 | 0.99 | 0.91 | |
| Season | - | - | - | - | <0.0001 | |
| Pretreatment × Substrate | 0.62 | 0.86 | * | 0.80 | 0.81 | |
| Pretreatment × Season | - | - | - | - | <0.0001 | |
| Substrate × Season | - | - | - | - | 0.99 | |
| Peroxidase (mg g−1) (Unrooted cuttings) | Pretreatment | 0.0001 | <0.0001 | 0.13 | 0.0003 | 0.009 |
| Substrate | 0.47 | 0.14 | 0.99 | 0.77 | 0.86 | |
| Season | - | - | - | - | <0.0001 | |
| Pretreatment × Substrate | 0.96 | 0.0005 | * | 0.43 | 0.68 | |
| Pretreatment × Season | - | - | - | - | <0.0001 | |
| Substrate × Season | - | - | - | - | 0.77 | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Abshahi, M.; García-Morote, F.A.; Zarei, H.; Zahedi, B.; Nejad, A.R. Improvement of Rooting Performance in Stem Cuttings of Savin Juniper (Juniperus sabina L.) as a Function of IBA Pretreatment, Substrate, and Season. Forests 2022, 13, 1705. https://0-doi-org.brum.beds.ac.uk/10.3390/f13101705
Abshahi M, García-Morote FA, Zarei H, Zahedi B, Nejad AR. Improvement of Rooting Performance in Stem Cuttings of Savin Juniper (Juniperus sabina L.) as a Function of IBA Pretreatment, Substrate, and Season. Forests. 2022; 13(10):1705. https://0-doi-org.brum.beds.ac.uk/10.3390/f13101705
Chicago/Turabian StyleAbshahi, Maliheh, Francisco Antonio García-Morote, Hossein Zarei, Bahman Zahedi, and Abdolhossein Rezaei Nejad. 2022. "Improvement of Rooting Performance in Stem Cuttings of Savin Juniper (Juniperus sabina L.) as a Function of IBA Pretreatment, Substrate, and Season" Forests 13, no. 10: 1705. https://0-doi-org.brum.beds.ac.uk/10.3390/f13101705