
Leaf Stoichiometry of Halophyte Shrubs and Its Relationship with Soil Factors in the Xinjiang Desert

1
College of Ecology and Environment, Xinjiang University, Urumqi 830017, China
2
Key Laboratory of Oasis Ecology of Education Ministry, Urumqi 830017, China
3
Xinjiang Jinghe Observation and Research Station of Temperate Desert Ecosystem, Ministry of Education, Xinjiang University, Urumqi 830017, China
4
Ecological Postdoctoral Research Station, Xinjiang University, Urumqi 830017, China
*
Author to whom correspondence should be addressed.
†
These authors contributed equally to this work.
Forests 2022, 13(12), 2121; https://0-doi-org.brum.beds.ac.uk/10.3390/f13122121
Submission received: 13 October 2022
/
Revised: 7 December 2022
/
Accepted: 9 December 2022
/
Published: 11 December 2022
(This article belongs to the Special Issue Plant Adaptation to Extreme Environments in Drylands)
Abstract
:Desert halophytes are a special plant group widely distributed in desert ecosystems. Studying their ecological stoichiometric characteristics is helpful for understanding their nutrient utilization characteristics and survival strategies. In this study, three functional groups of halophyte shrubs (euhalophytes, pseudohalophytes, and secretohalophytes) were studied in the Xinjiang desert, and the ecological stoichiometric characteristics of their leaves and their relationships with soil factors were evaluated. The results showed that the C content in secretohalophytes (442.27 ± 3.08 mg g−1) was significantly higher than that in the other functional groups (p < 0.05). The N and P contents in euhalophytes (22.17 ± 0.49 mg g−1 and 1.35 ± 0.04 mg g−1, respectively) were significantly higher than those in halophytes (p < 0.05). The N/P results showed that the growth rates of euhalophytes and pseudohalophytes were more susceptible to P limitation, whereas that of secretohalophytes was more susceptible to both N and P limitations, indicating that there were differences in nutrient characteristics among different functional groups. The results of the redundancy analysis showed that the leaf C, N, and P contents of euhalophytes were most affected by electrical conductivity (EC), whereas those of pseudohalophytes and secretohalophytes were most affected by the soil C content, indicating that different functional groups of halophyte shrubs had different responses to soil factors. The results of this study revealed the nutrient utilization characteristics of different functional groups of halophyte shrubs in the Xinjiang desert and their response and adaptation mechanisms to soil factors, thereby providing a basis for desert ecosystem management.
1. Introduction
Ecological stoichiometry studies the balance between C, N, P, and other nutrients among plants during their adaptation to and interactions with their environment [1]. Studying the ecological stoichiometric characteristics of desert plants can effectively evaluate and analyze the nutrient supply potential of desert soils and the productivity of desert ecosystems [2]. As a special desert plant group widely distributed in desert ecosystems, desert halophytes play an important role in maintaining nutrients and biodiversity and improving the regional climate in desert ecosystems [3]. Studying the ecological stoichiometric characteristics of desert halophytes is helpful for understanding their nutrient utilization characteristics and survival strategies [4,5]. With the aggravation of global soil salinization and N and P nutrient imbalances, the nutrient characteristics of desert halophytes have also changed [6]. Therefore, it is of great significance for the management and restoration of desert ecosystems to study the stoichiometric characteristics of desert halophyte leaves and their relationship with soil factors and to elucidate the nutrient limitations of desert halophytes and their response mechanisms to soil factors.
In recent years, many scholars have conducted extensive research on desert plants, mainly focusing on the relationship between desert plant nutrient characteristics, the climate, and the physical and chemical factors of soil [7,8,9,10]. Some studies believed that most soil factors directly affect the N and P stoichiometry, while climate factors indirectly affect the N and P stoichiometry of desert plants. Therefore, soil factors are still key factors affecting the nutrient characteristics of desert plants [11]. Relevant studies have shown that EC has direct effects on the leaf N and P contents of desert plants, and that pH and STN (soil total nitrogen) have direct effects on leaf N:P ratios [11]. Soil water content, total carbon (TC), total nitrogen (TN), and available phosphorus (AP) content are positively correlated with leaf C and N content [12], and soil water content is positively correlated with N and P in plant leaves and negatively correlated with other stoichiometric ratios. Soil salinity is positively correlated with leaf C/P and negatively correlated with leaf P [13]. However, according to some studies, the leaf stoichiometric characteristics of desert plants are not directly determined by soil nutrient content but are more affected by the genetic characteristics of plants [14,15]. The C, N, and P stoichiometries of plant leaves in different functional groups vary to different degrees; for example, succulent woody plants have higher N contents and lower C contents than herbaceous plants and non-succulent woody plants, reflecting the unique adaptability of desert plants to extreme habitats [16].
Xinjiang is an extremely arid area with scarce precipitation, strong evaporation, and a fragile ecological environment [17]. It is an area with the largest distribution area of saline alkali land in China; under such natural environmental conditions, the widespread distribution of desert halophytes has been promoted [18]. Under the long-term adaptation to soil salt, the halophytes in Xinjiang mainly evolved into three categories (euhalophytes, species that accumulate and isolate salt within succulent leaf or stem tissues; secretohalophytes, species with salt-secreting glands; pseudohalophytes, species that restrict the entry of saline ions into the transpiration stream) [19]. However, up to now, the stoichiometric characteristics of nutrients in different halophyte species have still been unclear, and there have been few studies on the relationship between the ecological stoichiometries of desert halophytes and environmental factors.
With the intensification of drought and salinization, the ecological stoichiometric characteristics of desert halophytes have changed [20,21]. Desert halophyte shrubs play an irreplaceable role in maintaining the stability of biodiversity in desert ecosystems [19]. The stoichiometry of C, N, and P in the leaves of three different functional groups of halophyte shrubs and their relationship with soil factors were studied in this paper; we clarify their response characteristics to soil factors in extremely arid areas. We assume that (1) under arid and poor nutrient conditions, halophytes have higher C content and lower N and P content; (2) owing to the different physiological functions of desert halophytes, the stoichiometric ratios of C, N, and P of different functional groups may have different response characteristics to soil factors. This study reveals the adaptation and feedback mechanisms of different functional groups of desert halophytes shrubs, enriches the research on stoichiometry in the desert ecosystem, and provides a scientific basis for the management of desert plants.
2. Materials and Methods
2.1. Study Area
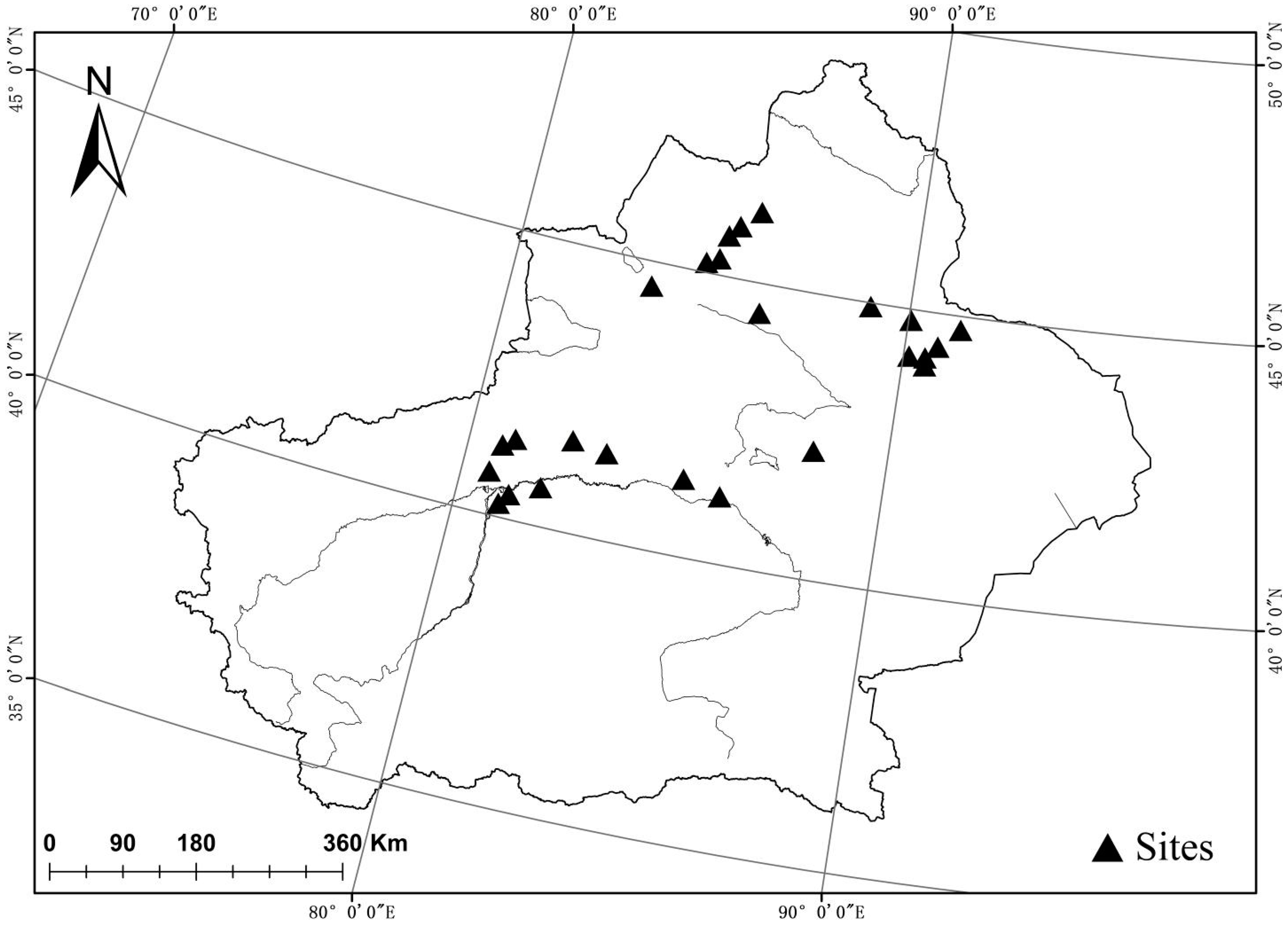
In this study, 25 sample plots were selected in the desert areas of the Junggar Basin and Tarim Basin in Xinjiang (Figure 1), spanning 80°39′–91°19′ E, 40°14′–46°14′ N, and 270–1025 m above sea level (Table S1). The study area has a typical temperate continental arid climate, with little rainfall and strong evaporation, and a high soil salt content. The mean annual precipitation (MAP) ranges from 45 mm to 159 mm, and the mean annual temperature (MAT) ranges from 5.85 to 11.87 °C. Our database consisted of 10 species from 6 families, including 225 samples (euhalophytes, 100 samples; pseudohalophytes, 40 samples; secretohalophytes, 85 samples). The dominant euhalophyte species include Haloxylon ammodendron and Halostachys caspica, the dominant pseudohalophyte species include Calligonum leucocladum and Halimodendron halodendron, and the dominant secretohalophyte species include Tamarix ramosissima and Tamarix arceuthoides (Table S2).
2.2. Field Sampling
Based on the data from the previous field survey and literature such as Xinjiang Saline Flora, typical sampling sites in Xinjiang desert were identified. In accordance with the dominance, the constructive species or common plant species within the sample area of the sampling site were selected so as to adequately represent the composition and structure of the plant community there. At the same sampling site (plant community), individuals of the same species were selected at a uniform distance for each species, so that the selected individuals could adequately represent the growth of the species in the community.
In July 2018, plant and soil surveys and sample collections were conducted at sites far from human disturbance. Three 10 × 10 m quadrats were randomly set up at each site and species composition was investigated, and the dominant species and soil samples were collected. At the species level, five individual plants of the same species were selected according to their growth status, and 50 g of fresh, healthy, and mature (non-destructive, sunny, petioles removed) plant leaves were collected from each individual plant. The collected plant leaves were loaded into an envelope with an appropriate amount of desiccant and labeled. In order to analyze soil nutrients, we randomly selected two sampling points from the four corners of each sample, took five 0–50 cm soil samples at each sampling point (remove impurities such as plants and rocks), and then fully mixed them to create a composite sample for each sample. Collected 100 g of soil samples from the composite samples, and put them into the sealed bag to take back to the laboratory for determination of soil nutrient content. In addition, we used GPS to record the altitude and geographical coordinates of each plot.
2.3. Chemical Analysis
The leaf samples were rinsed with deionized water to remove salt and dust from the leaf surface, then stored in a drying oven at 105 °C and dried for 30 min to reduce nutrient loss due to the respiration and decomposition of the leaves, and then dried to constant weight in an oven at 65 °C. The leaves were ground using a grinder and passed through an 80-mesh sieve, weighed, bagged, and sealed for the determination of C, N, and P contents. The collected soil samples were naturally dried, any remaining plant tissues were removed, and the soil samples were ground through a 60-mesh sieve. The sieved soil samples were divided into five equal parts and stored in numbered sample bags for the determination of soil nutrients and other indicators. An elemental analyzer (Elementar Inc., Hanau, Hessen, Germany) was used to determine the plant and soil carbon and nitrogen content (soil C and soil TN). The samples were digested with H2SO4-H2O2-HF, and the plant and soil phosphorus contents (soil TP) were analyzed by colorimetry. The ratio of soil to water in the test solution of soil pH value and electrical conductivity (EC) was 1:2.5 and 1:5, respectively, and then measured with a pH meter and electrical conductivity meter (Mettler-Toledo International Inc., Zurich, Greifensee, Switzerland).
2.4. Data Analysis
SPSS (Version 26.0, Armonk, NY, USA) was used for test data processing and statistical analysis. Levene’s test was used to test the homogeneity of variance. Duncan’s method was used when there was homogeneity of variance, and Tamhane’s T2 method was used when their variance was not homogeneous (α = 0.05). Canono (Version 5.0, Ithaca, NY, USA) was used for redundancy analysis of the leaf stoichiometric characteristics and soil factors. Blue solid arrows represent the ecological stoichiometric characteristics of halophyte shrub leaves, and red hollow arrows represent soil factors. The arrow length of each environmental factor represents the length of its eigenvector; this can also be seen as the effect of soil factors on leaf ecological stoichiometry. The longer the line, the greater the absolute cosine value, indicating a greater impact, and vice versa. The angle between the environmental factor arrow and sorting axis indicates the size of the correlation. The smaller the angle, the higher the correlation; that is, the sorting axis reflects the gradient of the environmental factor. The data in the chart are presented as the mean ± standard error (p < 0.05). MAP and MAT data were obtained from WorldClim Version 2.0 (http://worldclim.org/version2 (accessed on 7 March 2020)). In this study, the data were interpolated with a precision of 30 s, and the longitude, latitude, and elevation data were measured with GPS during the sampling process.
3. Results
3.1. Ecological Stoichiometric Characteristics of C, N, and P in Desert Saline Shrub Leaves
The mean C, N, and P contents in the leaves of Xinjiang desert halophyte shrubs were 409.31, 19.92, and 1.25 mg g−1, respectively (Table 1). The mean C/N, C/P, and N/P ratios were 22.25, 352.20, and 16.53, respectively. The coefficients of variation (CV) of C, N, and P content were 11.96, 29.42, and 28.18, respectively; the order from large to small was N > P > C. The coefficients of variation of C/N, C/P, and N/P ratios were 30.11, 29.01, and 28.67, respectively; the order from large to small was C/N > C/P > N/P.
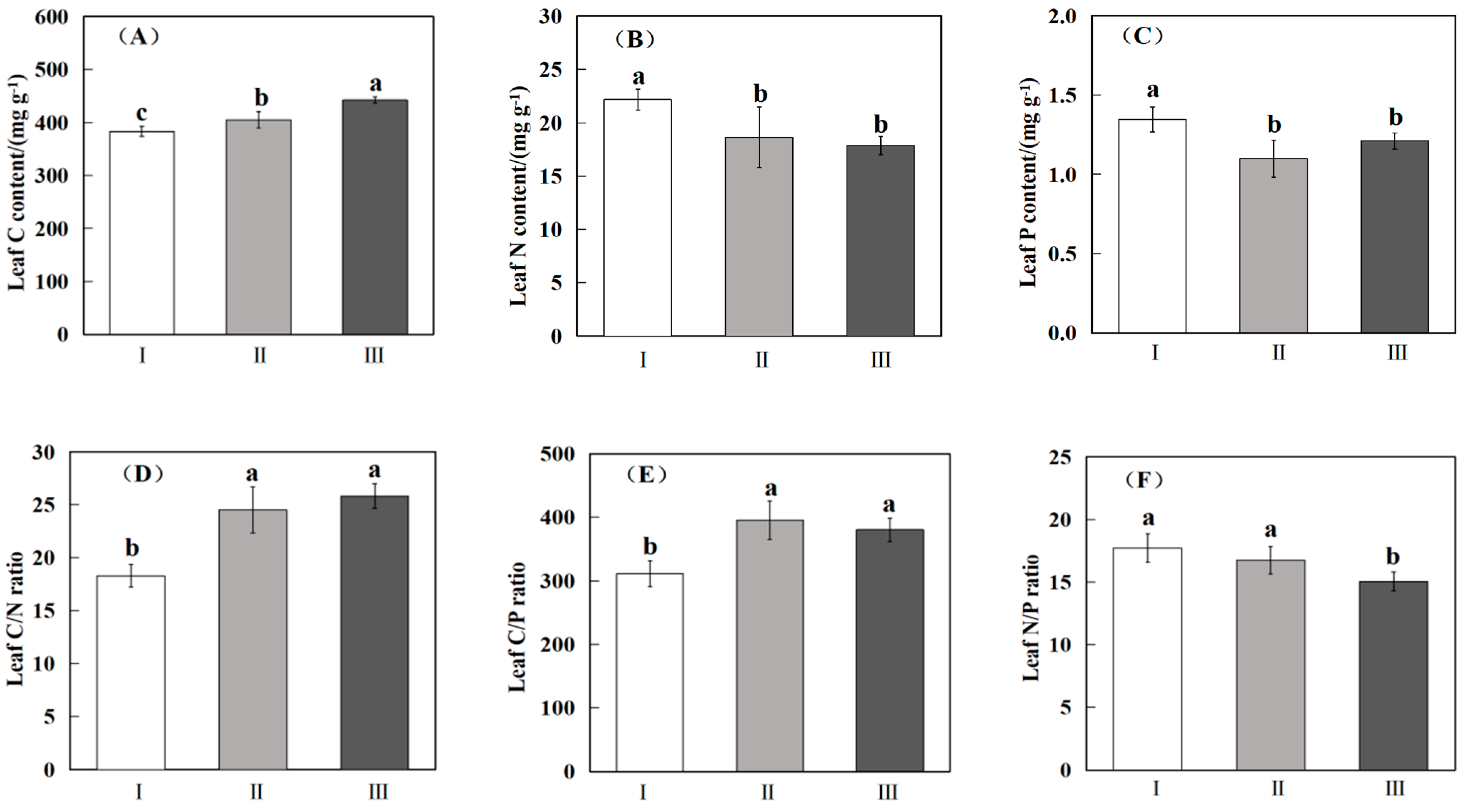
There were significant differences in the C, N, and P contents and stoichiometric characteristics of desert halophyte shrubs in different functional groups (Figure 2). The leaf C content of secretohalophytes (442.27 ± 3.08 mg g−1) was significantly higher than that of both pseudohalophytes (405.12 ± 7.49 mg g−1) and euhalophytes (382.96 ± 4.70 mg g−1) (p < 0.05), the N content of euhalophytes (22.17 ± 0.49 mg g−1) was significantly higher than that of pseudohalophytes (18.64 ± 1.41 mg g−1) and secretohalophytes (17.88 ± 0.43 mg g−1) (p < 0.05), and the P content of euhalophytes (1.35 ± 0.04 mg g−1) was significantly higher than that of both of pseudohalophytes (1.10 ± 0.06 mg g−1) and secretohalophytes (1.21 ± 0.03 mg g−1) (p < 0.05).
The stoichiometric ratios of C/N, C/P, and N/P in the leaves of different functional groups were also different (Figure 2). The C/N ratios of pseudohalophytes (24.53 ± 1.08) and secretohalophytes (25.83 ± 0.59) were significantly higher than that of euhalophytes (18.29 ± 0.54) (p < 0.05), the C/P of euhalophytes (310.98 ± 10.31) was significantly lower than that of both pseudohalophytes (395.41 ± 14.87) and secretohalophytes (380.36 ± 9.37) (p < 0.05), and the N/P of euhalophytes (17.71 ± 0.57) and pseudohalophytes (16.74 ± 0.55) were significantly higher than that of secretohalophytes (15.08 ± 0.38) (p < 0.05).
3.2. Characteristics of Soil Factors of Desert Halophyte Shrubs
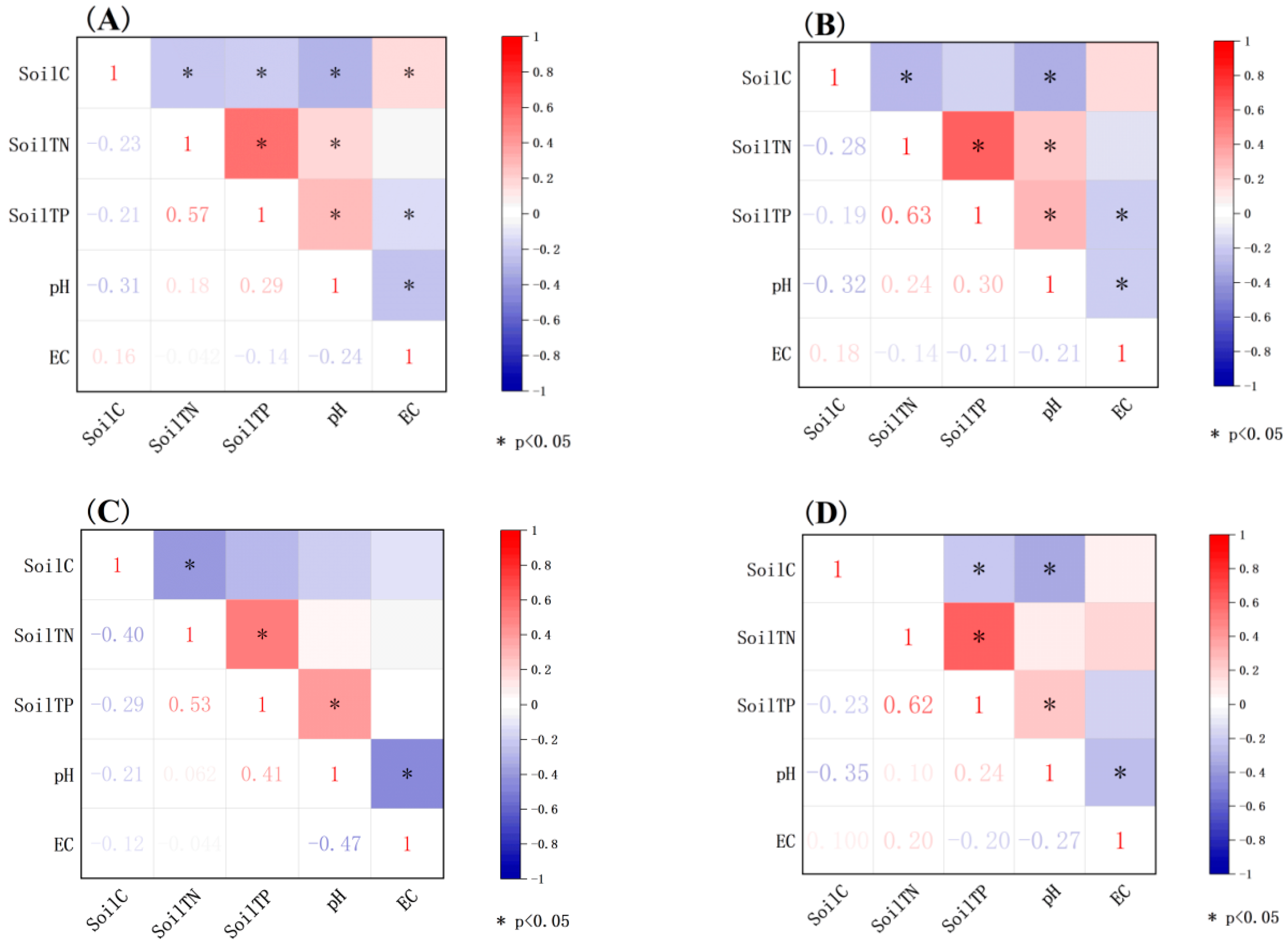
Except for the soil C content (8.75 mg g−1) and EC (1.58) of pseudohalophytes that were significantly lower than those of other functional groups, other factors had no significant differences between the soils of different desert halophyte functional groups (Table 2). The correlation results showed that except for soil TN and EC, other soil indicators showed significant correlations at the overall level (Figure 3A). For euhalophyte soil, soil C showed a significant negative correlation with soil TN and pH, soil TN was positively correlated with pH and soil TP, and soil TP was positively correlated with pH and negatively correlated with EC (Figure 3B). For pseudohalophyte soil, soil C and soil TN and pH and EC were negatively correlated, and soil TN and soil TP and soil TP and pH were significantly positively correlated (Figure 3C). For secretohalophyte soil, soil C indicated a significant negative correlation with soil TP and pH, and soil TN was positively correlated with soil TP, whereas soil TP was significantly positively correlated with pH, and there was a significant negative correlation between pH and EC (Figure 3D).
3.3. Relationship between Ecological Stoichiometric Characteristics of Desert Halophyte Shrub Leaves and Soil Factors
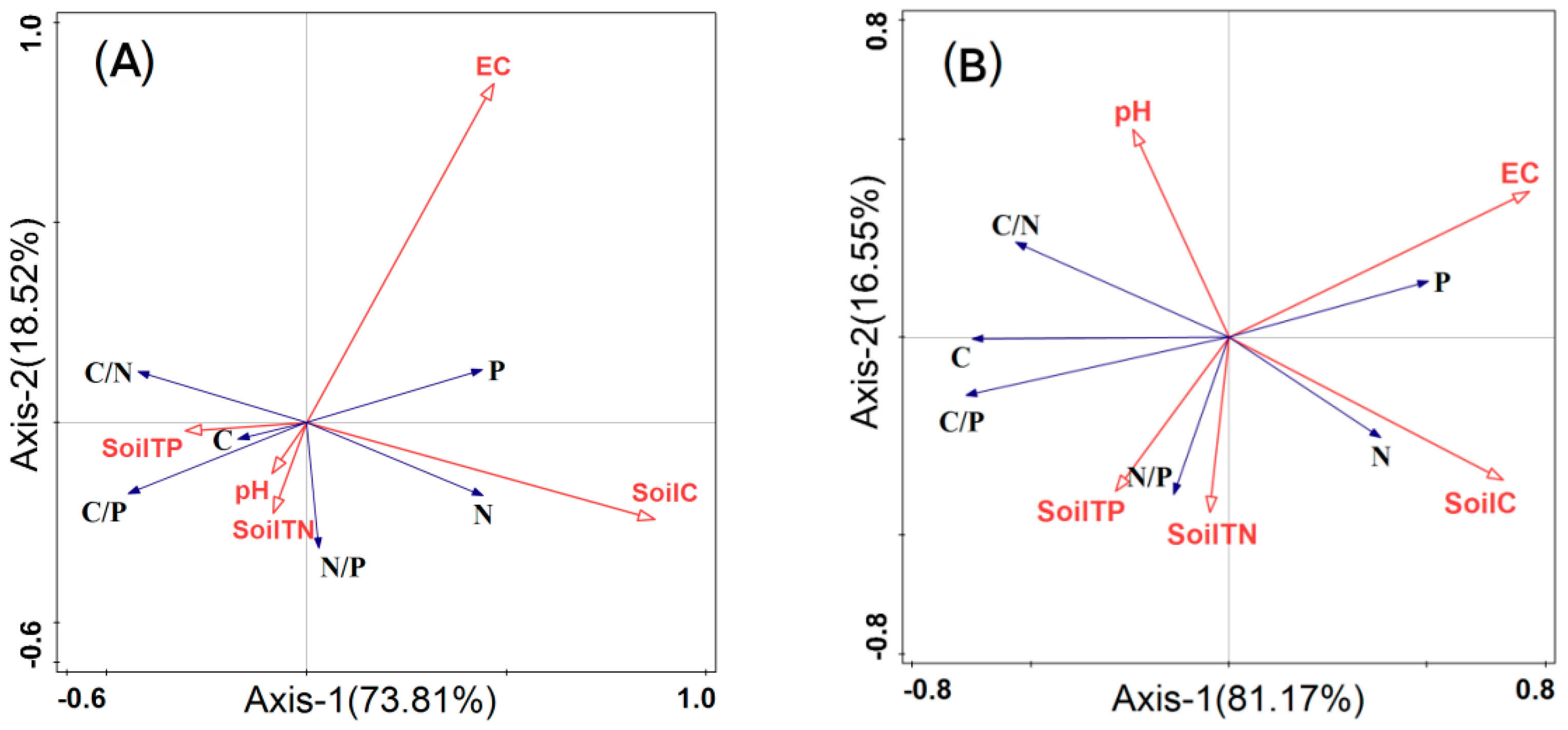
To explore the correlation between the ecological stoichiometric characteristics of the leaves of different functional groups of halophyte shrubs and their soil factors, a redundancy analysis was performed (Figure 4). Data of all plants in the study area are shown in Figure 4A. The total explanation rates of the C, N, and P ecological stoichiometric characteristics of whole halophyte shrub leaves on the first and second ordination axes were 73.81% and 18.52%, respectively, and the cumulative interpretation amount reached 92.33%. Among all soil factors, the largest absolute value of the soil C cosine value shows that soil C has the greatest impact on plants. Soil C and EC had the longest arrow connection. Therefore, the changes in the ecological stoichiometric characteristics of plant leaves can be well explained by soil C content and EC. Soil C was proportional to leaf N, P content, and N/P, and inversely proportional to C/N, C/P, and C, among which soil C had the greatest correlation with N; in addition, EC and P were positively correlated. Soil TN and pH were proportional to C/P and N/P, and inversely proportional to P; soil TP was inversely proportional to N and P and proportional to C, and the correlation with P was significantly greater than that with the other leaf ecological stoichiometric characteristics. There were some differences in the effects of soil factors on the ecological stoichiometry characteristics of plant leaves. The order of importance of the influence of each soil factor from the ecological stoichiometry characteristics of plant leaves was soil C (57.8%) > EC (23.2%) > soil TP (7.4%) > pH (5.9%) > soil TN (5.8%) (Table 3). Among all the factors, soil C and EC had very significant effects on the ecological stoichiometry of halophyte leaves (p < 0.01), and their interpretation rates for the ecological stoichiometry characteristics of halophyte leaves were 57.8% and 23.2%, respectively. The effect of soil TP and pH on the ecological stoichiometry characteristics of plant leaves was significant (p < 0.05), while the effect of soil TN on the ecological stoichiometry characteristics of plant leaves was not significant (p = 0.05) (Table 3).
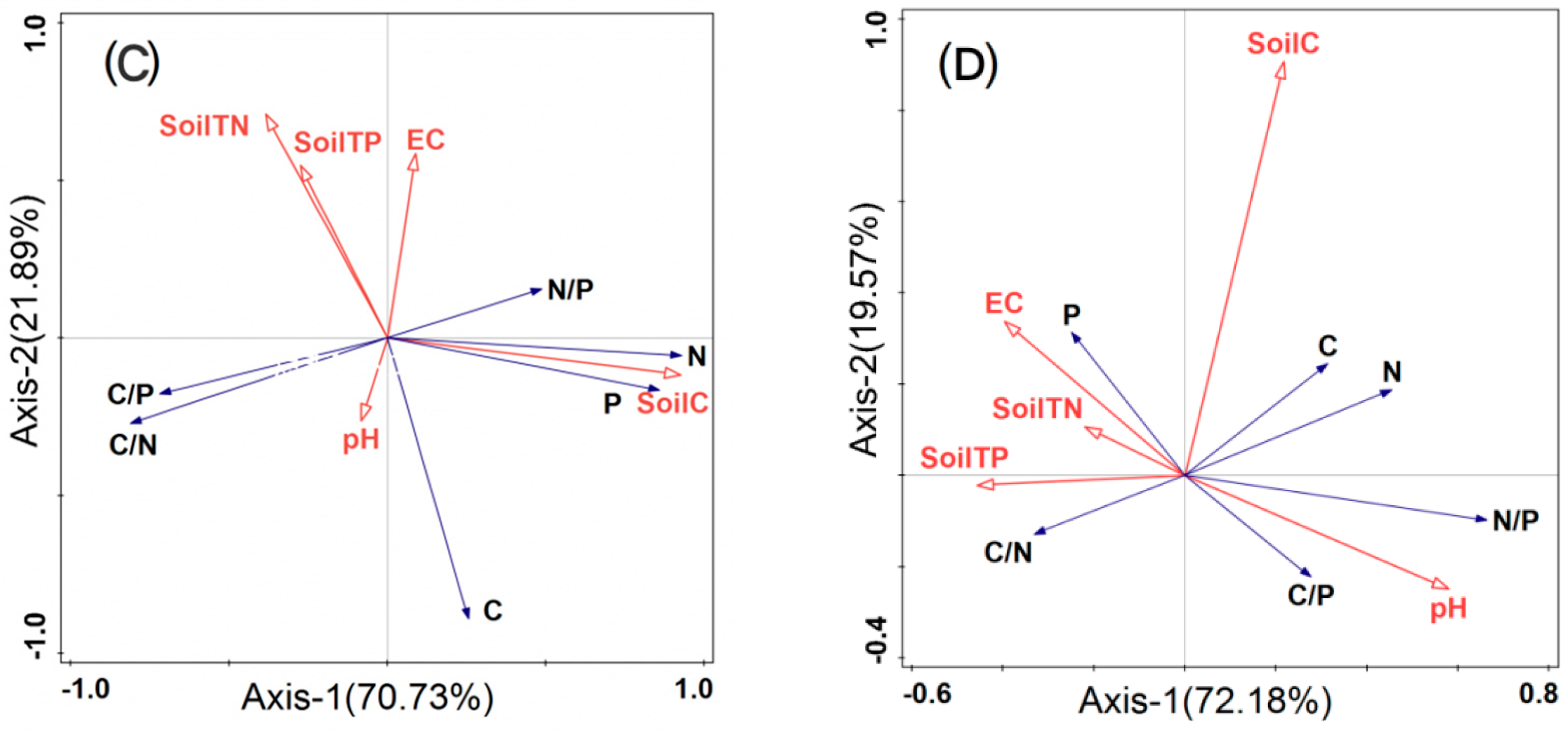
The leaf C, N, and P stoichiometry of euhalophytes (Figure 4B), pseudohalophytes (Figure 4C), and secretohalophytes (Figure 4D) were explained by 81.17%, 71.73%, and 72.18%, respectively, in the first ordination axis, but the soil factors had different effects on the leaf C, N, and P stoichiometric characteristics. Meanwhile, EC had the greatest impact on euhalophytes, and soil C had the greatest impact on pseudohalophytes and secretohalophytes. According to the angle in the diagram (Figure 4B–D), it can be observed that soil C was significantly negatively correlated with the C/N, C/P, and C content of euhalophytes, and positively correlated with the N content. Soil C was positively correlated with the C, N, and P of pseudohalophytes and secretohalophytes, and negatively correlated with the C/N and C/P of secretohalophytes. The soil TN was significantly negatively correlated with the P content in euhalophytes, N/P and C/P in secretohalophytes, and C, N, and P in pseudohalophytes. The soil TN was positively correlated with the N/P of euhalophytes, and the P content and C/N of secretohalophytes. The soil TP was significantly positively correlated with the C content, C/P, N/P, and C/N of euhalophytes. However, it was significantly negatively correlated with the P content of euhalophytes, C and P of pseudohalophytes, and N and N/P of secretohalophytes. There was a positive correlation between the pH, C, and C/N of euhalophytes, C content of pseudohalophytes, and N, C/P, and N/P of halophytes, but pH was only significantly negatively correlated with P of secretohalophytes. EC was significantly negatively correlated with the C content and C/P of euhalophytes and pseudohalophytes, and the C/P and N/P of secretohalophytes. EC was positively correlated with the phosphorus content in both euhalophytes and secretohalophytes.
4. Discussion
4.1. Stoichiometric Characteristic of C, N, and P Elements in Leaves of Desert Halophyte Shrubs
The characteristics of plants’ C, N, and P not only reflect the characteristics of plants, but also reflect the long-term adaptation and response to their environment [22,23]. The C content (409.31 ± 3.26 mg g−1) of the whole halophyte shrub leaves was lower than that of the global terrestrial plants (464.00 ± 32.10 mg g−1) [23] and the Sonoran Desert plants (434.8 ± 1.8 mg g−1) [2], indicating that the C reserve capacity was weak in this study, which was related to the strong diurnal temperature difference and low annual precipitation in the study area. The contents of N (19.9 mg g−1) and P (1.2 mg g−1) in the leaves of halophyte shrubs in this study were lower than in the China desert plants (24.4 and 1.7 mg g−1) [24], China desert halophytes (28.1 and 1.9 mg g−1) [25], China sand plants (34.1, (2.5 mg g−1) [26], and Sonoran Desert plants (25.9 and 1.5 mg g−1) [2] (Table S3). Scarce rainfall and intense evaporation result in extremely low soil moisture and nutrient contents in the study area, which ultimately leads to inadequate nutrient availability to desert halophyte shrubs [13]. Under conditions of water and nutrient stress, desert halophyte shrubs may be limited in their ability to take up nitrogen and phosphorus nutrients, which reduces nutrient cycling and metabolic activity, resulting in reduced nutrient feedback mechanisms between plants and the soil. As a result, plant nutrient contents are also expected to decrease. Desert halophyte shrubs adapt to their environment through a number of physiological regulators, such as slowing plant growth to reduce metabolic rates and reducing resource requirements to cope with the barren environment [27].
There were significant differences in the C, N, and P stoichiometric characteristics among the leaves of desert halophyte shrub groups in the study area, indicating that there were some differences in the resource utilization efficiency and adaptation strategies in response to adverse environments [19,28]. As the main element of plant dry matter, C is also the substrate and energy source of various physiological metabolic processes of plants [25]. The C content in the leaves of halophyte shrubs was significantly different among different halophyte species. The leaves of some halophytes are scaly, forming a special stem leaf callus similar to the stem. The tissue is composed of large epidermis, porous cells, and water storage and vascular tissues. Salinity also improves the function of these tissues, making the saline plants better able to adapt to extreme drought conditions, which helps plants to enhance photosynthesis and improve the ability of plant leaves to synthesize carbohydrates [28]. The low C content in the leaves of euhalophytes may be related to their salt tolerance. The fleshy development of euhalophyte leaves can dilute excess salt in the body to alleviate physiological water shortages [3]. N and P are two important limiting elements in plant growth and development. Their effectiveness is one of the main factors regulating the plant litter decomposition rate and ecosystem nutrient balance, affecting the entire growth process of plants [29]. Euhalophytes have strong and unique salt tolerance mechanisms. Under salt stress conditions, the N and P contents in the leaves are significantly increased. In addition, non-protein N accumulation in euhalophytes is higher under salt stress conditions, which plays a crucial role in osmotic regulation [25,30]. Pseudohalophytes and secretohalophytes have different salt tolerance mechanisms to euhalophytes. The low N content in secretohalophytes is mainly due to the formation of salt glands or salt vesicles on their leaves, which can secrete excessive salt ions in plants [30]. The low P content in pseudohalophytes is due to the suberization of the root cortex. The main component of suberization is water-insoluble fatty substances, which are impermeable, making it difficult for salts dissolved in soil water to enter plants [31].
C/N and C/P ratios are important physiological indicators of plants. Generally, plants with higher C/N and C/P ratios have relatively lower growth rates [24] and higher carbon sequestration efficiencies [32]. Secretohalophytes and pseudohalophytes had the highest C/N and C/P ratios, and they had higher carbon sequestration advantages in nutrient-poor habitats than euhalophytes [13]. Overall, C/N and C/P were high, which can be attributed to the extreme drought, high temperature, and strong evaporation in the study area [16,30]. To resist harsh environments, plants reduce their growth rate, resulting in relatively high C/N and C/P ratios in the leaves [32]. Leaf N/P is considered an important indicator of plant and soil nutrient limitation [33]. The N/P of euhalophytes and pseudohalophytes was greater than 16, and that of secretohalophytes was less than 16. This indicates that the growth of euhalophytes and pseudohalophytes is more susceptible to P limitation, whereas the growth of secretohalophytes is more susceptible to both N and P limitations. Overall, the N/P ratio was much higher than in other regions [34,35], and greater than 16, indicating that P is the main limiting element for desert plant growth in the study area. Nutrient deficiency and water shortage lead to insufficient nutrient supply for desert halophytes [25]. Under limited water and nutrient conditions, the ability of desert halophytes to assimilate P may be limited, thereby reducing nutrient cycling and metabolic activity, and ultimately leading to a decrease in nutrient feedback mechanisms between plants and soil [30]. In addition, less precipitation and greater evaporation makes it difficult for desert halophytes to obtain phosphorus from the soil, thereby making it more vulnerable to P limitation in this area.
4.2. Characteristics and Relationship of Soil Factors
Soil C, N, and P contents can indicate soil nutrient storage, nutrient cycling, and balance and are important indicators of soil organic matter composition and quality [36]. The soil C content (8.75 mg g−1) of pseudohalophytes was the lowest among the halophyte groups, and the overall C content was low (9.71 mg g−1), which was far lower than the national level (11.12 mg g−1) [37]. It is possible that the soil productivity of desert halophyte shrubs in Xinjiang is low, which is related to the nutrient conditions of the soil itself and the lower feedback of plant litter to the soil in the study area [30]. There was no significant difference in soil N and P contents among different halophyte functional groups, but the overall N and P contents were high (2.47 and 2.48 mg g−1); they were significantly higher than the average global level (1.06 and 0.65 mg g−1) [37] because the N and P contents increased under salt stress [4], which is consistent with the results of Wang et al. (2017) [25]. EC can represent some physical information about soil, such as salt state, nutrient content, and water content [38,39]. The pH was significantly negatively correlated with EC in the soil of all three halophytes, and EC was related to the salt content. The larger the EC, the higher the content of soluble salt ions [38]. The soil EC (1.58) of pseudohalophytes was the lowest among the halophytes, indicating that the soil soluble salt ion content was the lowest.
4.3. Response of Desert Halophyte Shrubs’ Stoichiometry to Soil Factors
Plant growth and development are closely related to their environment. Changes in the element content and the stoichiometric ratio in plants reflect the response and adaptation of plants to the environment [40,41,42]. Many studies have found that soil is the main nutrient source for plant growth and development, and its nutrient characteristics have an important impact on plant leaf N and P contents [43,44]. Soil nutrients significantly affect the ecological stoichiometric characteristics of plant leaves [45]. Based on the redundancy analysis of the relationship between the stoichiometric characteristics of different functional groups of salinized shrubs and soil factors (Figure 4), there was a significant correlation between the stoichiometric characteristics of the different functional groups of the halophyte shrub and soil factors. Overall, the growth and development of desert saline shrubs were limited by soil C and EC. However, for the different functional groups of halophyte shrubs, the responses of leaf C, N, and P ecological stoichiometric characteristics to soil factors were different. EC represents soil salinity, and among them, it had the greatest impact on euhalophytes. It was found that the protein content of euhalophytes was very high under drought and salt stress, indicating that the plant has a higher nitrogen uptake or metabolism capacity [3]. In addition, it was shown that euhalophytes also absorb large amounts of salts and store them in their bodies during growth, and in addition to accumulating salts, they grow certain contents whose growth is positively correlated with external salinity [3,6]. For pseudohalophytes and secretohalophytes, soil C is one of the greater factors affecting their nutrient characteristics. Soil C is mainly derived from the content of soil organic matter and the decomposition of apoplastic matter, and is strongly influenced by plants, water and heat, and parent material. Due to drought and salinity stress, plants need to increase the proportion of C-rich tissues (e.g., lignin) in their own bodies to protect the plant body from damage to adapt to the drought environment [3,46].
Only leaf N had a significant positive correlation with soil TN in euhalophytes (Figure 4B), indicating that soil TN has a limiting effect on euhalophyte growth. The soil TN level determines the absorption of N by euhalophyte leaves to a certain extent [47,48]. The C and P contents in the leaves of secretohalophytes were positively correlated with soil C and soil TP, respectively (Figure 4C,D), indicating that secretohalophytes were limited by soil C and P. However, leaf C was only positively correlated with soil C in pseudohalophytes, indicating that the growth and development of pseudohalophytes were limited by soil C [49]. This may be because pseudohalophytes increase their salt tolerance through the compensatory growth of roots [3], and reduced C content is transported up leaves by pseudohalophytes; thus, leaf C was positively correlated with soil C. There was no significant difference in soil N and P contents among the different functional groups of halophyte shrubs, but the utilization and consumption strategies of nutrient elements in different functional groups of halophyte shrubs were different [30]. This indicates that the ecological stoichiometric characteristics of desert saline shrub leaves are not directly determined by the characteristics of soil nutrient contents, but are related to their own genetic characteristics, which reflects the unique adaptation mechanism of different desert halophyte shrubs to habitats [50], consistent with the results of Luo et al. (2017) [51] and Song et al. (2020) [52]. Under soil moisture- and nutrient-scarce conditions, Xinjiang desert halophyte shrubs have formed their own unique stoichiometric characteristics and physiological ecology, reflecting the relatively stable adaptability of desert saline shrubs to extreme environments [16]. The environmental indicators selected in this study are limited, and more environmental indicators will be combined in future studies to conduct a comprehensive analysis of the relationship between halophytes and environmental factors.
5. Conclusions
It is of great importance to reveal the nutritional characteristics and ecological adaptation mechanisms of desert halophytes to study the element accumulation and utilization characteristics of different functional groups and their adaptability to harsh soil from the perspective of plant nutrition. In this study, C content of the whole halophyte shrub leaves was lower than that of the global terrestrial plants and the Sonoran desert plants. The contents of N and P in the leaves of halophyte shrubs were lower than in the China desert plants, China desert halophytes, China sand plants, and Sonoran desert plants. Through the analysis of the stoichiometric characteristics of C, N, and P in the leaves of euhalophytes, secretohalophytes, and pseudohalophytes, and their correlations with soil factors, it was found that there were significant differences in the leaf contents and ratios. This indicates that the N and P nutrient limitations and nutrient resource utilization efficiencies of halophytes in different functional groups were different. The redundancy analysis results showed that the C, N, and P contents in the leaves of euhalophytes were most affected by EC, whereas the leaves of pseudohalophytes and secretohalophytes were most affected by soil C content, indicating that different functional groups of halophyte shrubs had different responses to soil factors. In this study, euhalophytes and pseudohalophytes can be applied with phosphate fertilizer due to P limitation, and salt-secreting halophytes can be appropriately applied with nitrogen fertilizer and phosphate fertilizer due to the common limitation of N and P. Desert halophytes show unique advantages, especially in saline alkali habitats. Desert halophytes can not only prevent salt accumulation in the tilth, but also improve soil structure and fertility, and promote good plant soil nutrient cycling in desert ecosystems. Therefore, in terms of desert halophytes’ management, we need to pay attention to their adaptation to salt and differences in nutrient utilization so as to better use halophytes to improve the soil conditions in desert areas.
Supplementary Materials
The following supporting information can be downloaded at: https://0-www-mdpi-com.brum.beds.ac.uk/article/10.3390/f13122121/s1, Table S1: Geography information and climate characteristics of the sampling sites in the deserts of Xinjiang, China; Table S2: List of species sampled in the deserts of Xinjiang, China; Table S3: Leaf C, N, P content and C/N, C/P, N/P ratio of plants in various regions.
Author Contributions
Y.L. carried out the fieldwork and wrote the first draft of the manuscript. C.L. wrote the first draft of the manuscript. L.G. assisted with revising the draft manuscript. C.M. carried out the fieldwork. All authors contributed to the article and approved the submitted version. All authors have read and agreed to the published version of the manuscript.
Funding
Third Xinjiang Scientific Expedition Program (No. 2021xjkk0903), China Postdoctoral Science Foundation (No. 2022M722667), and Department of Education of Xinjiang Uygur Autonomous Region, Dr. Tianchi Program Project (No. TCBS202123).
Data Availability Statement
The available data can be obtained in the supplementary files, but anyone who uses the data must get the consent of the authors.
Acknowledgments
We thank Yanming Gong for their assistance with field sampling and laboratory work.
Conflicts of Interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
References
- Sterner, R.W.; Elser, J.J. Ecological Stoichiometry: Biology of Elements from Molecules to the Biosphere; Princeton University Press: Princeton, NJ, USA, 2002. [Google Scholar]
- Castellanos, A.E.; Llano-Sotelo, J.M.; Machado-Encinas, L.I.; Lopez-Pina, J.E.; Romo-Leon, J.R.; Sardans, J.; Penuelas, J. Foliar C, N, and P stoichiometry characterize successful plant ecological strategies in the Sonoran Desert. Plant Ecol. 2018, 219, 775–788. [Google Scholar] [CrossRef] [Green Version]
- Zhao, K.F.; Li, F.Z.; Zhang, F.S. Halophytes in China, 2nd ed.; Science Press: Beijing, China, 2013. [Google Scholar]
- He, M.Z.; Dijkstra, F.A.; Zhang, K.; Li, X.R.; Tan, H.J.; Gao, Y.H.; Li, G. Leaf nitrogen and phosphorus of temperate desert plants in response to climate and soil nutrient availability. Sci. Rep. 2014, 4, 6932. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chatterton, N.J.; Goodin, J.R.; Duncan, C. Nitrogen metabolism in Atriplex polycarpa as affected by substrate nitrogen and NaCl salinity. Agron. J. 1971, 63, 271–274. [Google Scholar] [CrossRef]
- Zhang, K.; Li, C.J.; Li, Z.S.; Zhang, F.H.; Zhao, Z.Y.; Tian, C.Y. Characteristics of mineral elements in shoots of three annual halophytes in a saline desert, Northern Xinjiang. J. Arid. Land 2013, 5, 244–254. [Google Scholar] [CrossRef] [Green Version]
- He, M.Z.; Dijkstra, F.A.; Zhang, K.; Tan, H.J.; Zhao, Y.; Li, X.R. Influence of life form, taxonomy, climate, and soil properties on shoot and root concentrations of 11 elements in herbaceous plants in a temperate desert. Plant Soil 2016, 398, 339–350. [Google Scholar] [CrossRef]
- Zhang, X.M.; Wang, Y.D.; Zhao, Y.; Xu, X.W.; Lei, J.Q.; Hill, R.L. Litter decomposition and nutrient dynamics of three woody halophytes in the Taklimakan desert highway shelterbelt. Arid. Land Res. Manag. 2017, 31, 335–351. [Google Scholar] [CrossRef]
- Liu, R.; Cieraad, E.; Li, Y. Summer rain pulses may stimulate a CO2 release rather than absorption in desert halophyte communities. Plant Soil 2013, 373, 799–811. [Google Scholar] [CrossRef]
- Amatangelo, K.L.; Vitousek, P.M. Stoichiometry of ferns in Hawaii: Implications for nutrient cycling. Oecologia 2008, 157, 619–627. [Google Scholar] [CrossRef]
- Luo, Y.; Peng, Q.W.; Li, K.H.; Gong, Y.M.; Liu, Y.Y.; Han, W.X. Patterns of nitrogen and phosphorus stoichiometry among leaf, stem and root of desert plants and responses to climate and soil factors in Xinjiang, China. Catena 2021, 199, 105100. [Google Scholar] [CrossRef]
- Zhang, X.L.; Zhou, J.H.; Guan, T.Y.; Cai, W.T.; Jiang, L.H.; Lai, L.M.; Gao, N.N.; Zheng, Y.R. Spatial variation in leaf nutrient traits of dominant desert riparian plant species in an arid inland river basin of China. Ecol. Evol. 2019, 9, 1523–1531. [Google Scholar] [CrossRef]
- Wang, L.; Zhao, G.; Li, M.; Zhang, M.; Zhang, L.; Zhang, X.; Xu, S. C:N:P stoichiometry and leaf traits of halophytes in an arid saline environment, northwest China. PLoS ONE 2015, 10, e0119935. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yuan, K.Y.; Xu, H.L.; Zhang, G.P.; Yan, J.J. Aridity and high salinity, rather than soil nutrients, regulate nitrogen and phosphorus stoichiometry in desert plants from the individual to the community level. Forests 2022, 13, 890. [Google Scholar] [CrossRef]
- Luo, Y.; Gong, L.; Zhu, M.L.; An, S.Q. Leaf and soil ecological stoichiometric characteristics of four shrub species in desert area of upper reaches of Tarim River. Acta Ecol. Sin. 2017, 37, 8326–8335. (In Chinese) [Google Scholar] [CrossRef]
- Sun, M.M.; Zhai, B.C.; Chen, Q.W.; Li, G.Q.; Du, S. Response of density-related fine root production to soil and leaf traits in coniferous and broad-leaved plantations in the semiarid loess hilly region of China. J. For. Res. 2022, 33, 1071–1082. [Google Scholar] [CrossRef]
- Zheng, Z.Y.; Ma, Z.G.; Li, M.X.; Xia, J.J. Regional water budgets and hydroclimatic trend variations in Xinjiang from 1951 to 2000. Clim. Chang. 2017, 144, 447–460. [Google Scholar] [CrossRef] [Green Version]
- Zhang, W.T.; Wu, H.Q.; Gu, H.B.; Feng, G.L.; Wang, Z.; Sheng, J.D. Variability of soil salinity at multiple spatio-temporal scales and the related driving factors in the oasis areas of Xinjiang, China. Pedoshere 2015, 24, 753–762. [Google Scholar] [CrossRef]
- Xi, J.B.; Zhang, F.S.; Tian, C.Y. Halophytes in Xinjiang; Science Press: Beijing, China, 2006. [Google Scholar]
- Zhang, X.; Guan, T.; Zhou, J.; Cai, W.; Gao, N.; Du, H.; Zheng, Y. Community characteristics and leaf stoichiometric traits of desert ecosystems regulated by precipitation and soil in an arid area of China. Int. J. Environ. Res. Public Health 2018, 15, 109. [Google Scholar] [CrossRef] [Green Version]
- Sardans, J.; Grau, O.; Chen, H.Y.H.; Janssens, I.A.; Ciais, P.; Piao, S.L.; Penuelas, J. Changes in nutrient concentrations of leaves and roots in response to global change factors. Glob. Chang. Biol. 2017, 23, 3849–3856. [Google Scholar] [CrossRef] [Green Version]
- Hou, X.Y. Vegetation Geography and Chemical Constituents of Dominant Plants in China; Science Press: Beijing, China, 1982. [Google Scholar]
- Elser, J.J.; Fagan, W.F.; Denno, R.F.; Dobberfuhl, D.R.; Folarin, A.; Huberty, A.A.; Interlandi, S.; Kilham, S.S.; McCauley, E.; Schulz, K.L.; et al. Nutritional constraints in terrestrial and freshwater food webs. Nature 2000, 408, 578–580. [Google Scholar] [CrossRef]
- Li, Y.L.; Mao, W.; Zhao, X.Y.; Zhang, T.H. Leaf nitrogen and phosphorus stoichiometry in typical desert and desertified regions, north China. Environ. Sci. 2010, 31, 1716–1725. [Google Scholar] [CrossRef]
- Wang, L.L.; Wang, L.; He, W.L.; An, L.Z.; Xu, S.J. Nutrient resorption or accumulation of desert plants with contrasting sodium regulation strategies. Sci. Rep. 2017, 7, 17035. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, L.F.; Wang, L.L.; He, W.L.; Zhang, X.F.; An, L.Z.; Xu, S.J. Patterns of leaf N:P stoichiometry along climatic gradients in sandy region, north of China. J. Plant Ecol. 2018, 11, 218–225. [Google Scholar] [CrossRef] [Green Version]
- Li, J.L.; Sun, X.; Zheng, Y.; Lü, P.P.; Wang, Y.L.; Guo, L.D. Diversity and community of culturable endophytic fungi from stems and roots of desert halophytes in northwest China. MycoKeys 2020, 62, 75–95. [Google Scholar] [CrossRef] [PubMed]
- Meng, X.Q.; Zhou, J.; Sui, N. Mechanisms of salt tolerance in halophytes: Current understanding and recent advances. Open Life Sci. 2018, 13, 149–154. [Google Scholar] [CrossRef]
- Yang, Y.; Liu, B.R.; An, S.S. Ecological stoichiometry in leaves, roots, litters and soil among different plant communities in a desertified region of Northern China. Catena 2018, 166, 328–338. [Google Scholar] [CrossRef]
- Luo, Y.; Chen, Y.; Peng, Q.W.; Li, K.H.; Mohammat, A.; Han, W.X. Nitrogen and phosphorus resorption of desert plants with various degree of propensity to salt in response to drought and saline stress. Ecol. Indic. 2021, 125, 107488. [Google Scholar] [CrossRef]
- Li, A.; Guo, D.; Wang, Z.Q.; Liu, H.Y. Nitrogen and phosphorus allocation in leaves, twigs, and fine roots across 49 temperate, subtropical and tropical tree species: A hierarchical pattern. Funct. Ecol. 2010, 24, 224–232. [Google Scholar] [CrossRef]
- Gong, Y.M.; Lv, G.H.; Guo, Z.J.; Chen, Y.; Cao, J. Influence of aridity and salinity on plant nutrients scales up from species to community level in a desert ecosystem. Sci. Rep. 2017, 7, 6811. [Google Scholar] [CrossRef] [Green Version]
- Güsewell, S. N:P ratios in terrestrial plants: Variation and functional significance. New Phytol. 2004, 164, 243–266. [Google Scholar] [CrossRef]
- Tong, R.; Zhou, B.Z.; Jiang, L.N.; Ge, X.G.; Cao, Y.H.; Shi, J.X. Leaf litter carbon, nitrogen and phosphorus stoichiometry of Chinese fir (Cunninghamia lanceolata) across China. Glob. Ecol. Conserv. 2021, 27, e01542. [Google Scholar] [CrossRef]
- Han, W.X.; Fang, J.Y.; Guo, D.L.; Zhang, Y. Leaf nitrogen and phosphorus stoichiometry across 753 terrestrial plant species in China. New Phytol. 2005, 168, 377–385. [Google Scholar] [CrossRef] [PubMed]
- Gao, R.; Ai, N.; Liu, G.Q.; Liu, C.H.; Zhang, Z.Y. Soil C:N:P stoichiometric characteristics and soil quality evaluation under different restoration modes in the loess region of Northern Shanxi Province. Forests 2022, 13, 913. [Google Scholar] [CrossRef]
- Li, Y.Q.; Zhao, X.Y.; Zhang, F.X.; Awada, T.; Wang, S.K.; Zhao, H.L.; Zhang, T.H.; Li, Y.L. Accumulation of soil organic carbon during natural restoration of desertified grassland in China’s Horqin Sandy Land. J. Arid. Land 2015, 7, 328–340. [Google Scholar] [CrossRef]
- Yang, Y.H.; Ji, C.J.; Ma, W.H.; Wang, S.F.; Wang, S.P.; Han, W.X.; Mohammat, A.; Robinson, D.; Smith, P. Significant soil acidification across northern China’s grasslands during 1980s–2000s. Glob. Chang. Biol. 2012, 18, 2293–2300. [Google Scholar] [CrossRef]
- Liu, W.; Li, R.J.; Han, T.T.; Cai, W.; Fu, Z.W.; Lu, Y.T. Salt stress reduces root meristem size by nitric oxide-mediated modulation of auxin accumulation and signaling in arabidopsis. Plant Physiol. 2015, 168, 343–U607. [Google Scholar] [CrossRef] [Green Version]
- Yang, X.; Tang, Z.; Ji, C.; Liu, H.; Ma, W.; Mohhamot, A.; Zheng, C. Scaling of nitrogen and phosphorus across plant organs in shrubland biomes across Northern China. Sci. Rep. 2014, 4, 5448. [Google Scholar] [CrossRef] [Green Version]
- Zheng, S.X.; Shangguan, Z.P. Spatial patterns of leaf nutrient traits of the plants in the Loess Plateau of China. Trees 2007, 21, 357–370. [Google Scholar] [CrossRef]
- He, J.S.; Fang, J.Y.; Wang, Z.H.; Guo, D.L.; Flynn, D.F.B.; Geng, Z. Stoichiometry and large-scale patterns of leaf carbon and nitrogen in the grassland biomes of China. Oecologia 2006, 149, 115–122. [Google Scholar] [CrossRef]
- Zhang, L.; Guo, H.; Bao, A.K. Unique salt-secreting structure of halophytes-salt vesicles. Plant Physiol. 2019, 55, 232–240. [Google Scholar] [CrossRef]
- Cho, Y.; Sudduth, K.A.; Chung, S.O. Soil physical property estimation from soil strength and apparent electrical conductivity sensor data. Biosyst. Eng. 2016, 152, 68–78. [Google Scholar] [CrossRef]
- An, H.; Tang, Z.S.; Keesstra, S.; Shangguan, Z.P. Impact of desertification on soil and plant nutrient stoichiometry in a desert grassland. Sci. Rep. 2019, 9, 9422. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rong, Q.Q.; Liu, J.T.; Cai, Y.P.; Lu, Z.H.; Zhao, Z.Z.; Yue, W.C.; Xia, J.B. Leaf carbon, nitrogen and phosphorus stoichiometry of Tamarix chinensis Lour. in the Laizhou Bay coastal wetland, China. Ecol. Eng. 2015, 76, 57–65. [Google Scholar] [CrossRef]
- Flowers, T.J.; Torke, P.F.; Yeo, A.R. The mechanism of salt tolerance in halophytes. Annu. Rev. Plant Physiol. 1977, 28, 89–121. [Google Scholar] [CrossRef]
- Flowers, T.J.; Colmer, T.D. Salinity tolerance in halophytes. New Phytol. 2008, 179, 945–963. [Google Scholar] [CrossRef] [PubMed]
- Yi, L.P.; Ma, J.; Li, Y. Soil salt and nutrient concentration in the rhizosphere of desert halophytes. Acta Ecol. Sin. 2007, 27, 3565–3571. [Google Scholar] [CrossRef]
- Didi, D.A.; Su, S.P.; Sam, F.E.; Tiika, R.J.; Zhang, X. Effect of plant growth regulators on osmotic regulatory substances and antioxidant enzyme activity of Nitraria tangutorum. Plants 2022, 11, 2559. [Google Scholar] [CrossRef]
- Sardans, J.; Peñuelas, J. The role of plants in the effects of global change on nutrient availability and stoichiometry in the plant-soil system. Plant Physiol. 2012, 160, 1741–1761. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Song, Z.P.; Liu, Y.H.; Su, H.X.; Hou, J.H. N-P utilization of Acer mono leaves at different life history stages across altitudinal gradients. Ecol. Evol. 2020, 10, 851–862. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Locations of the 29 sampling sites in Xinjiang, China.

Figure 2.
Comparison of leaf C, N, P content (A–C) and ratio (D–F) of different functional groups of desert saline shrubs. Note: I: euhalophyte; II: pseudohalophytes; III: secretohalophyte. Lowercase letters indicate significant differences between plants (p < 0.05).
Figure 2.
Comparison of leaf C, N, P content (A–C) and ratio (D–F) of different functional groups of desert saline shrubs. Note: I: euhalophyte; II: pseudohalophytes; III: secretohalophyte. Lowercase letters indicate significant differences between plants (p < 0.05).

Figure 3.
Correlation heat map of soil factors of desert halophytes in different functional groups. Note: (A) whole soil; (B) euhalophyte soil; (C) pseudohalophyte soil; (D) secretohalophyte soil.
Figure 3.
Correlation heat map of soil factors of desert halophytes in different functional groups. Note: (A) whole soil; (B) euhalophyte soil; (C) pseudohalophyte soil; (D) secretohalophyte soil.

Figure 4.
Redundancy analysis two−dimensional ordination diagram of the relationship between leaf C, N, P stoichiometric characteristics and soil factors. Note: (A) whole plants; (B) euhalophytes; (C) pseudohalophytes; (D) secretohalophytes.
Figure 4.
Redundancy analysis two−dimensional ordination diagram of the relationship between leaf C, N, P stoichiometric characteristics and soil factors. Note: (A) whole plants; (B) euhalophytes; (C) pseudohalophytes; (D) secretohalophytes.



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
The overall descriptive statistical characteristics of C, N, P stoichiometric characteristics of desert saline shrub leaves.
Table 1.
The overall descriptive statistical characteristics of C, N, P stoichiometric characteristics of desert saline shrub leaves.
| Variable | n | Mean | SE | Minimum | Maximum | CV (%) |
|---|---|---|---|---|---|---|
| C (mg g−1) | 225 | 409.31 | 3.26 | 303.64 | 512.29 | 11.96 |
| N (mg g−1) | 225 | 19.92 | 0.39 | 10.21 | 43.97 | 29.42 |
| P (mg g−1) | 225 | 1.25 | 0.24 | 0.59 | 2.39 | 28.18 |
| C/N | 225 | 22.25 | 0.45 | 9.40 | 44.38 | 30.11 |
| C/P | 225 | 352.20 | 6.81 | 145.12 | 654.44 | 29.01 |
| N/P | 225 | 16.53 | 0.32 | 7.02 | 37.39 | 28.67 |
Note: n, sample size.
Table 2.
Multiple comparisons of soil factors of different functional groups of saline shrubs.
| Plant Group | Soil C (mg g−1) | Soil TN (mg g−1) | Soil TP (mg g−1) | pH | EC (mS cm−1) |
|---|---|---|---|---|---|
| Euhalophyte | 9.99 ± 0.22 a | 2.54 ± 0.17 a | 2.34 ± 0.19 a | 8.90 ± 0.34 a | 10.04 ± 1.30 a |
| Pseudohalophyte | 8.75 ± 0.31 b | 2.74 ± 0.32 a | 2.28 ± 0.33 a | 8.90 ± 0.50 a | 1.58 ± 0.40 b |
| Secretohalophyte | 9.82 ± 0.23 a | 2.24 ± 0.11 a | 2.71 ± 0.19 a | 8.80 ± 0.30 a | 14.29 ± 1.86 a |
| Mean | 9.71 ± 0.14 a | 2.47 ± 0.10 a | 2.48 ± 0.13 a | 8.86 ± 0.21 a | 10.14 ± 0.96 a |
Note: Lowercase letters indicate significant differences in the same soil factors between different functional groups of saline shrubs (p < 0.05).
Table 3.
Results of the importance ranking and significance tests of the overall soil factors’ variables interpretation.
Table 3.
Results of the importance ranking and significance tests of the overall soil factors’ variables interpretation.
| Name | Importance Rank | Contribution % | Pseudo-F | p |
|---|---|---|---|---|
| Soil C | 1 | 57.8 | 25.6 | 0.002 |
| Soil EC | 2 | 23.2 | 10.7 | 0.002 |
| Soil TP | 3 | 7.4 | 3.5 | 0.02 |
| pH | 4 | 5.9 | 2.8 | 0.048 |
| Soil TN | 5 | 5.8 | 2.7 | 0.05 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Luo, Y.; Lian, C.; Gong, L.; Mo, C. Leaf Stoichiometry of Halophyte Shrubs and Its Relationship with Soil Factors in the Xinjiang Desert. Forests 2022, 13, 2121. https://0-doi-org.brum.beds.ac.uk/10.3390/f13122121
AMA Style
Luo Y, Lian C, Gong L, Mo C. Leaf Stoichiometry of Halophyte Shrubs and Its Relationship with Soil Factors in the Xinjiang Desert. Forests. 2022; 13(12):2121. https://0-doi-org.brum.beds.ac.uk/10.3390/f13122121
Chicago/Turabian StyleLuo, Yan, Cuimeng Lian, Lu Gong, and Chunnan Mo. 2022. "Leaf Stoichiometry of Halophyte Shrubs and Its Relationship with Soil Factors in the Xinjiang Desert" Forests 13, no. 12: 2121. https://0-doi-org.brum.beds.ac.uk/10.3390/f13122121
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.