
Impact of Ice-Storms and Subsequent Salvage Logging on the Productivity of Cunninghamia lanceolata (Chinese Fir) Forests

 , and
, and
Abstract
:1. Introduction
2. Materials and Methods
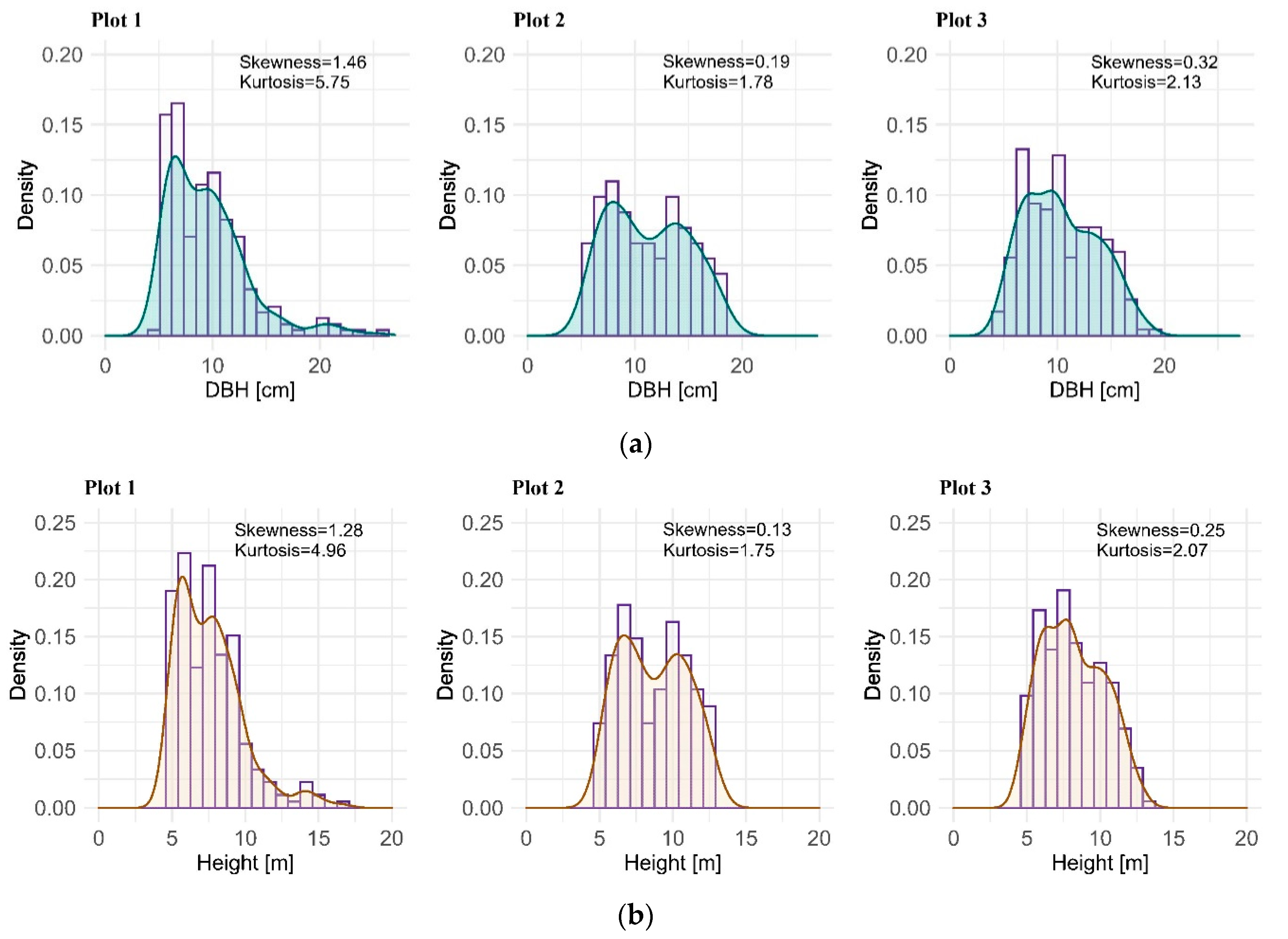
2.1. Data Collection
2.2. Model Configuration and Simulation Setup
2.2.1. Model Overview
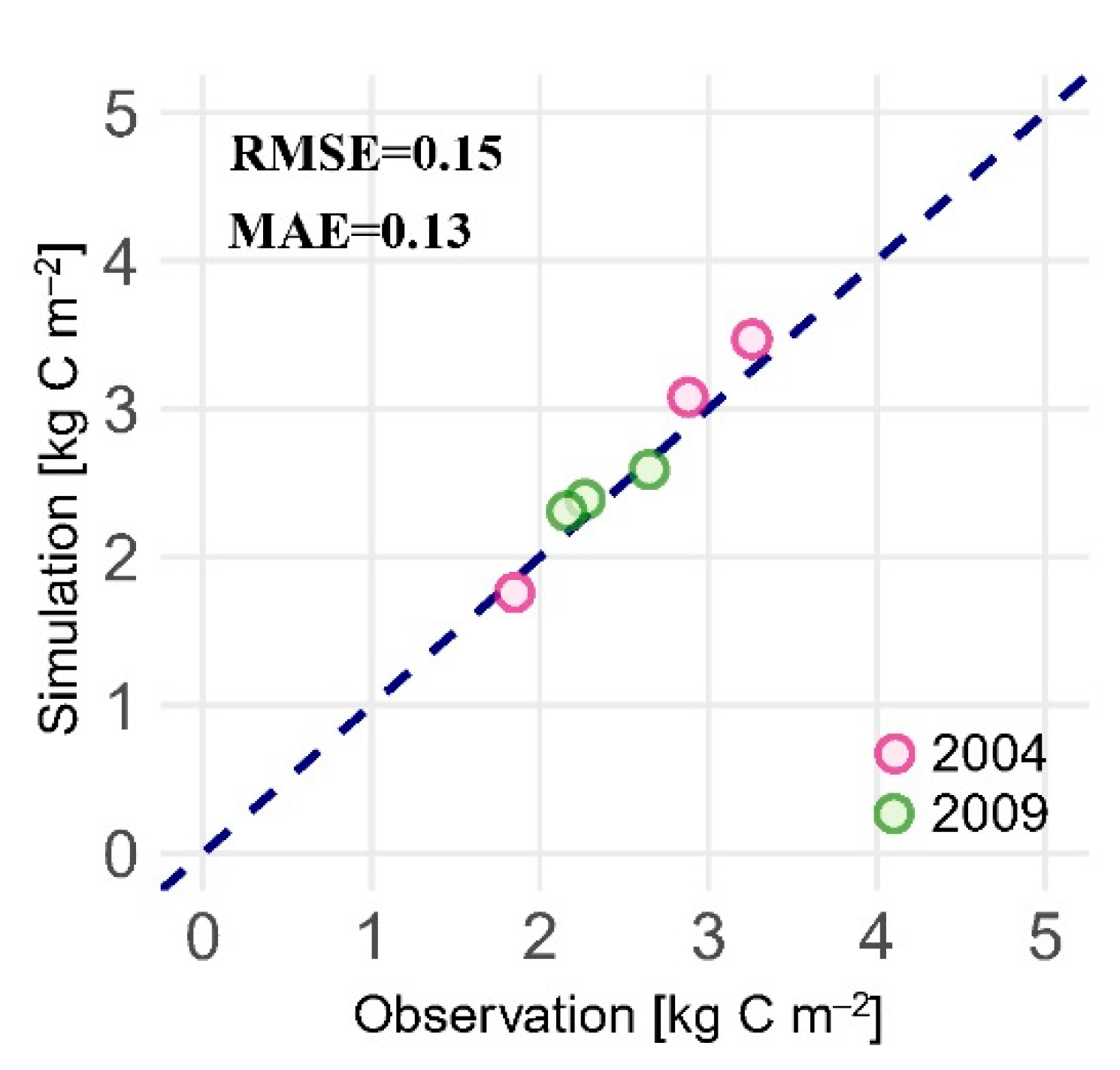
2.2.2. Model Calibration, Verification, and Validation
2.2.3. Scenarios Setup and Simulation
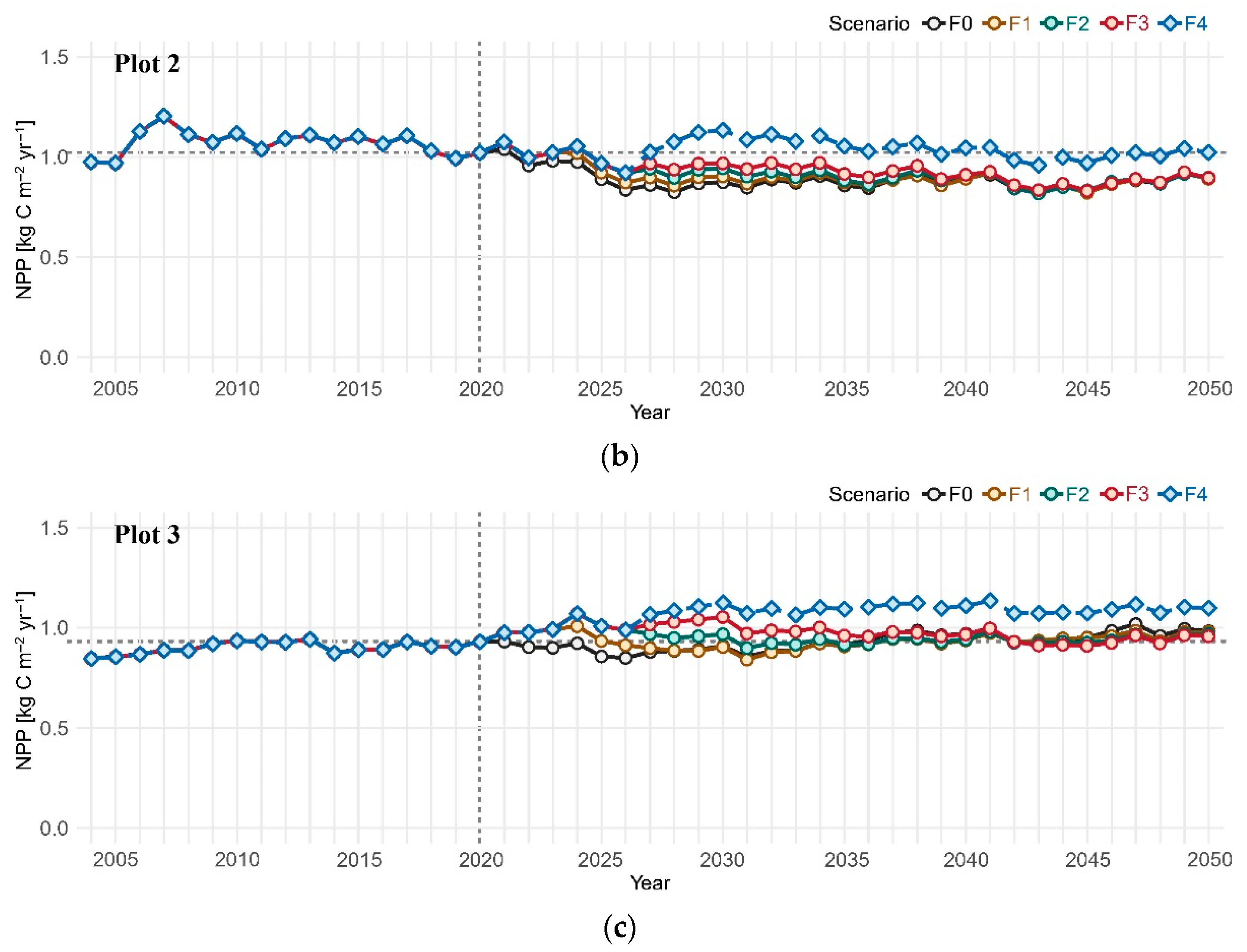
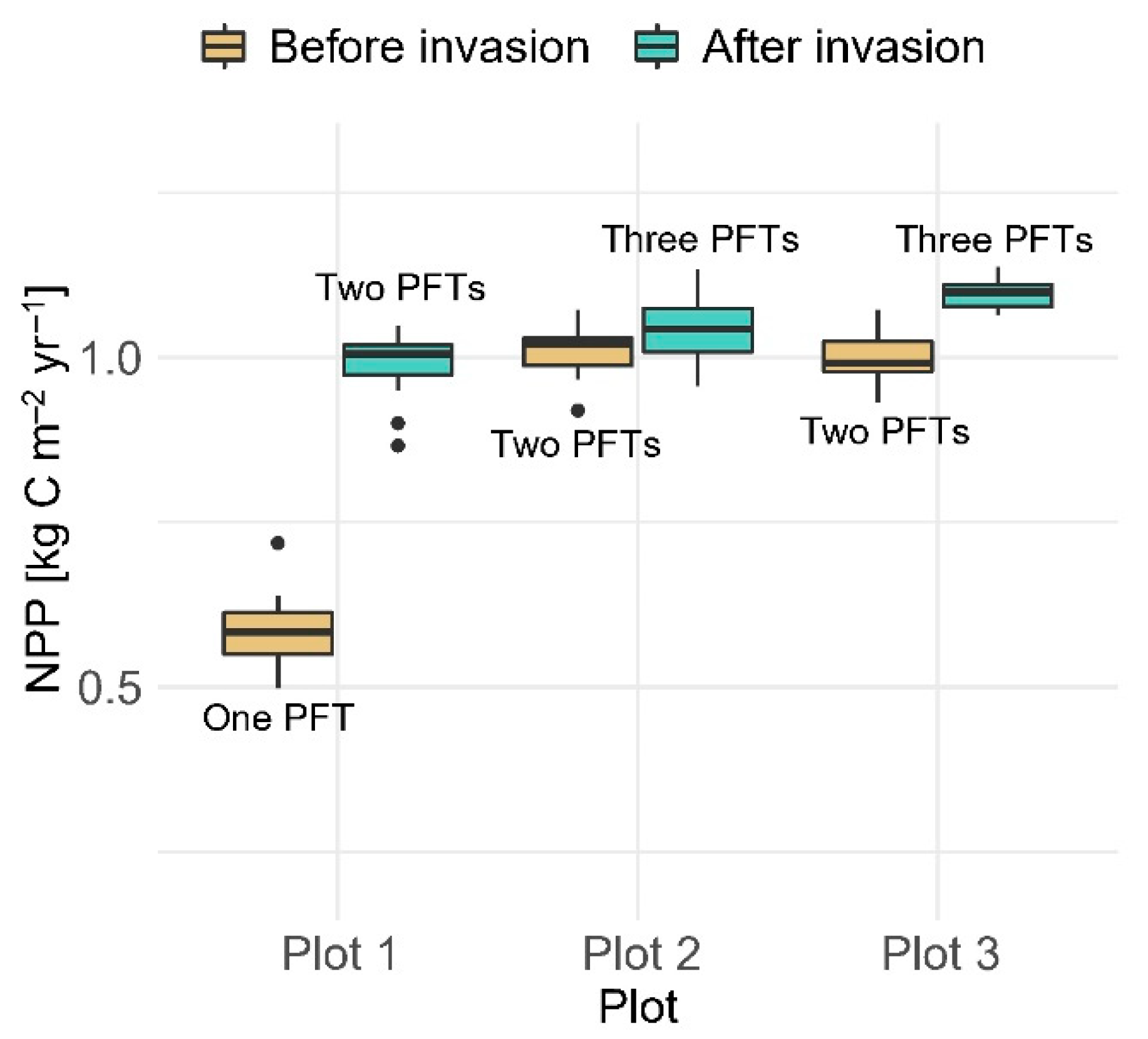
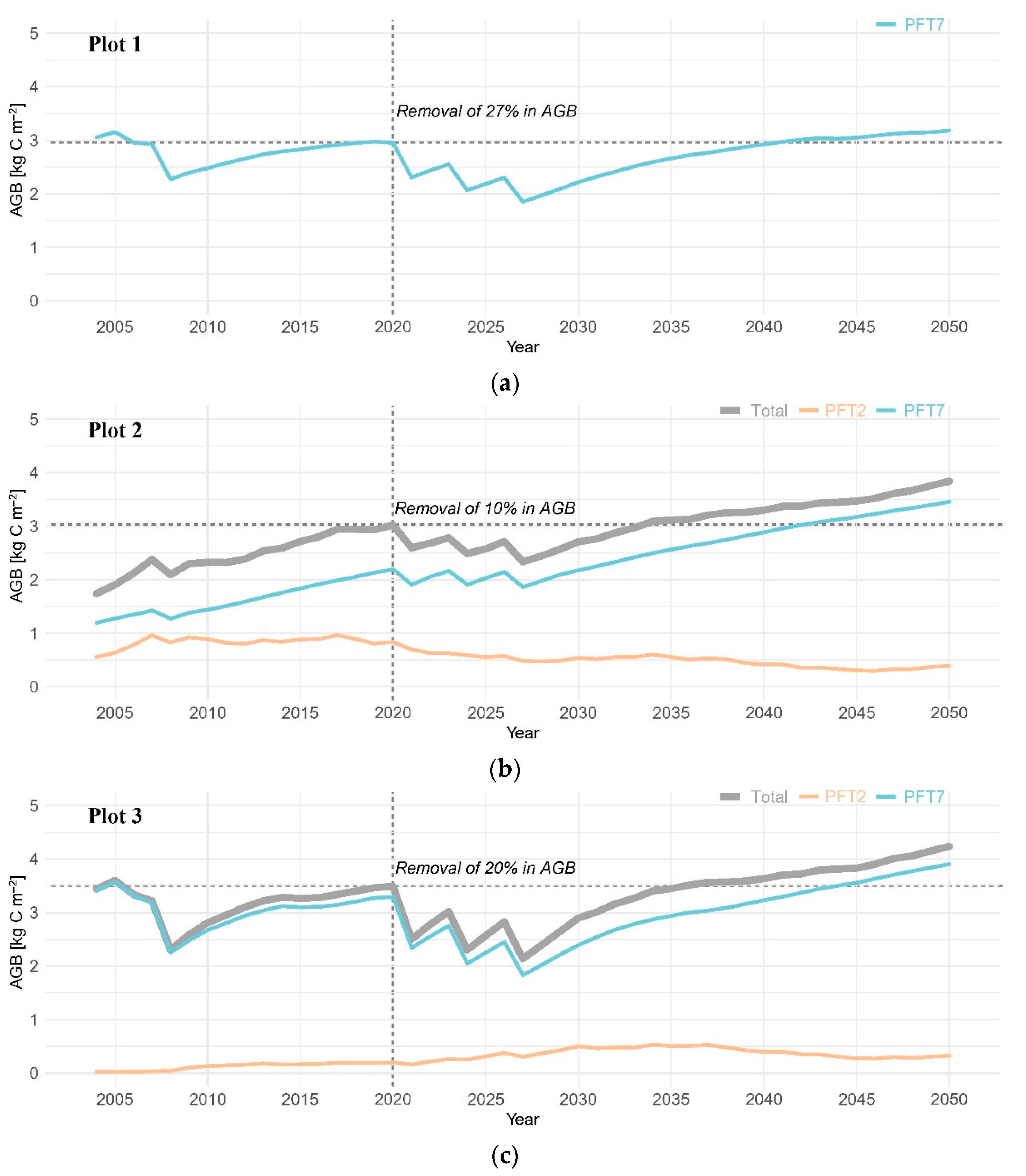
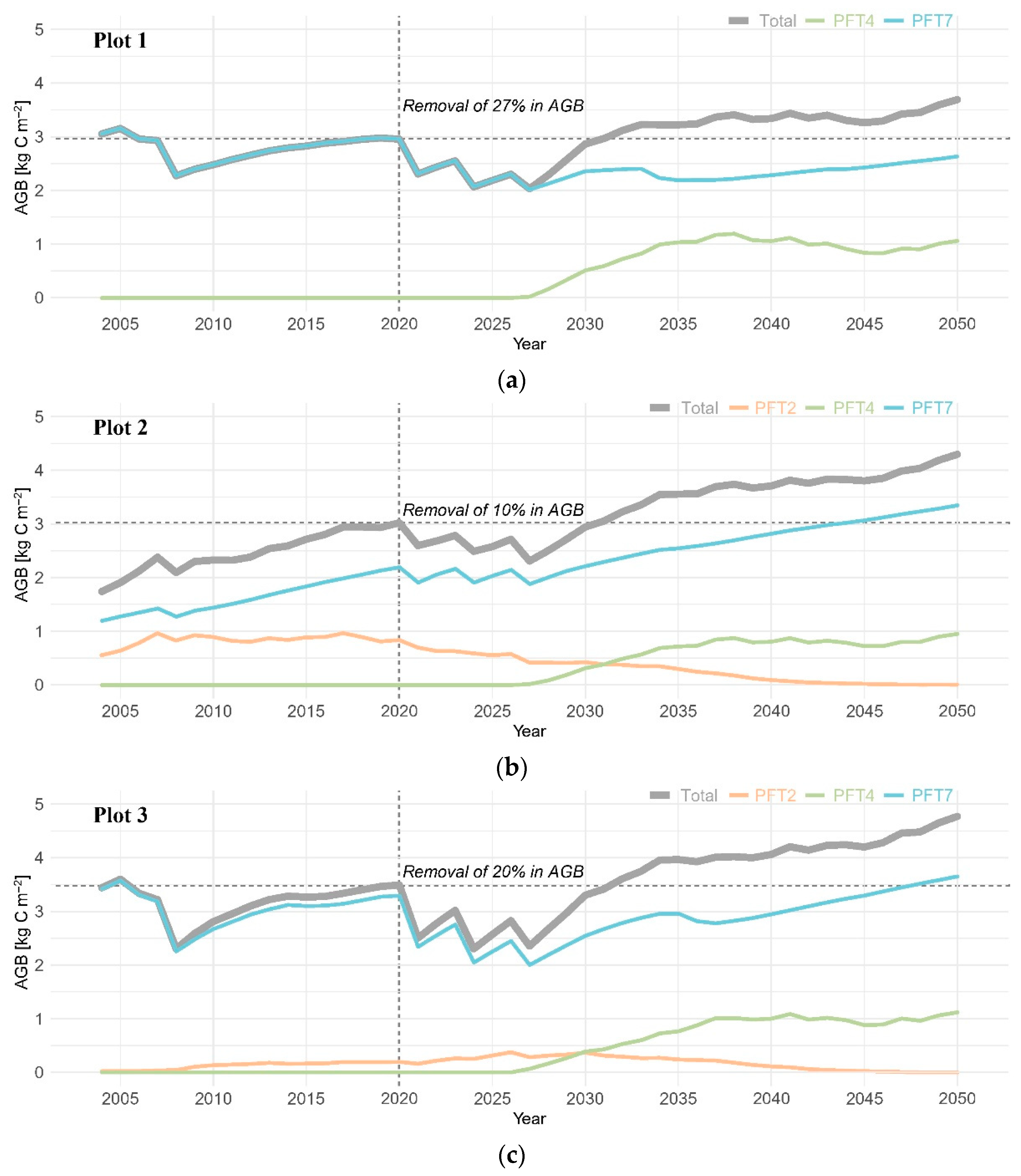
3. Results
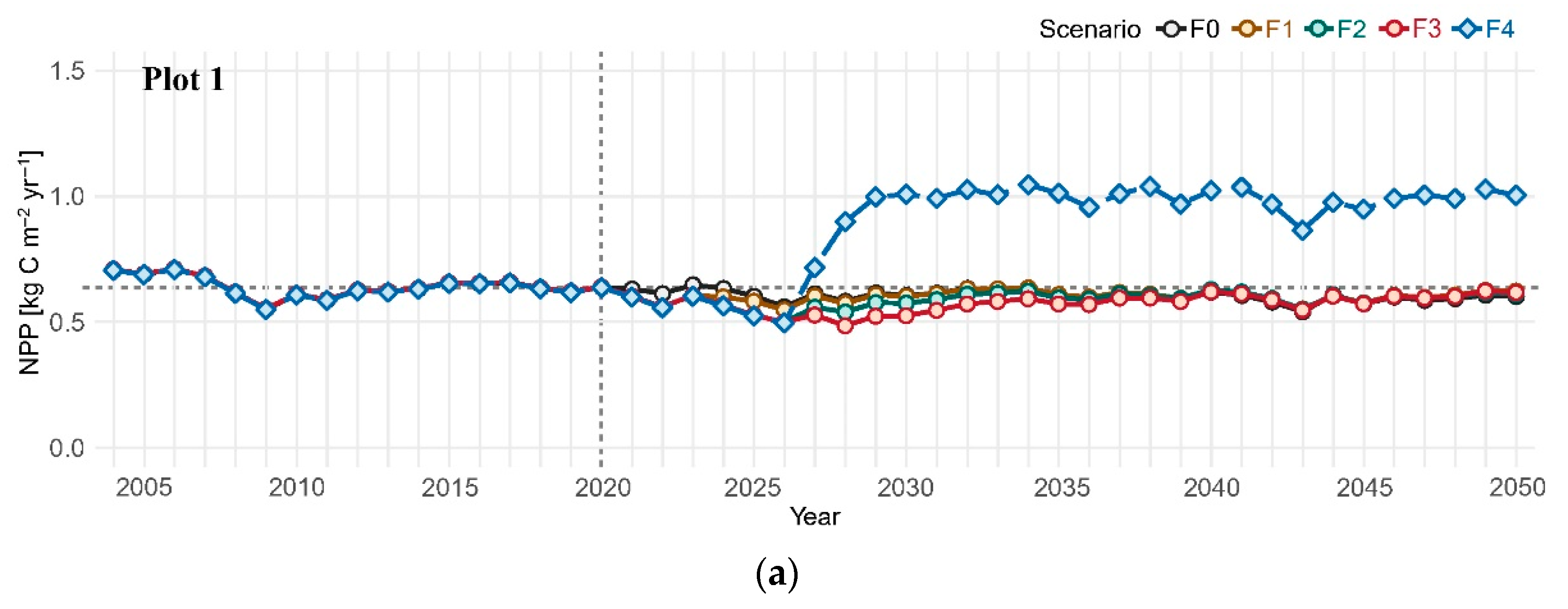
3.1. Net Primary Productivity
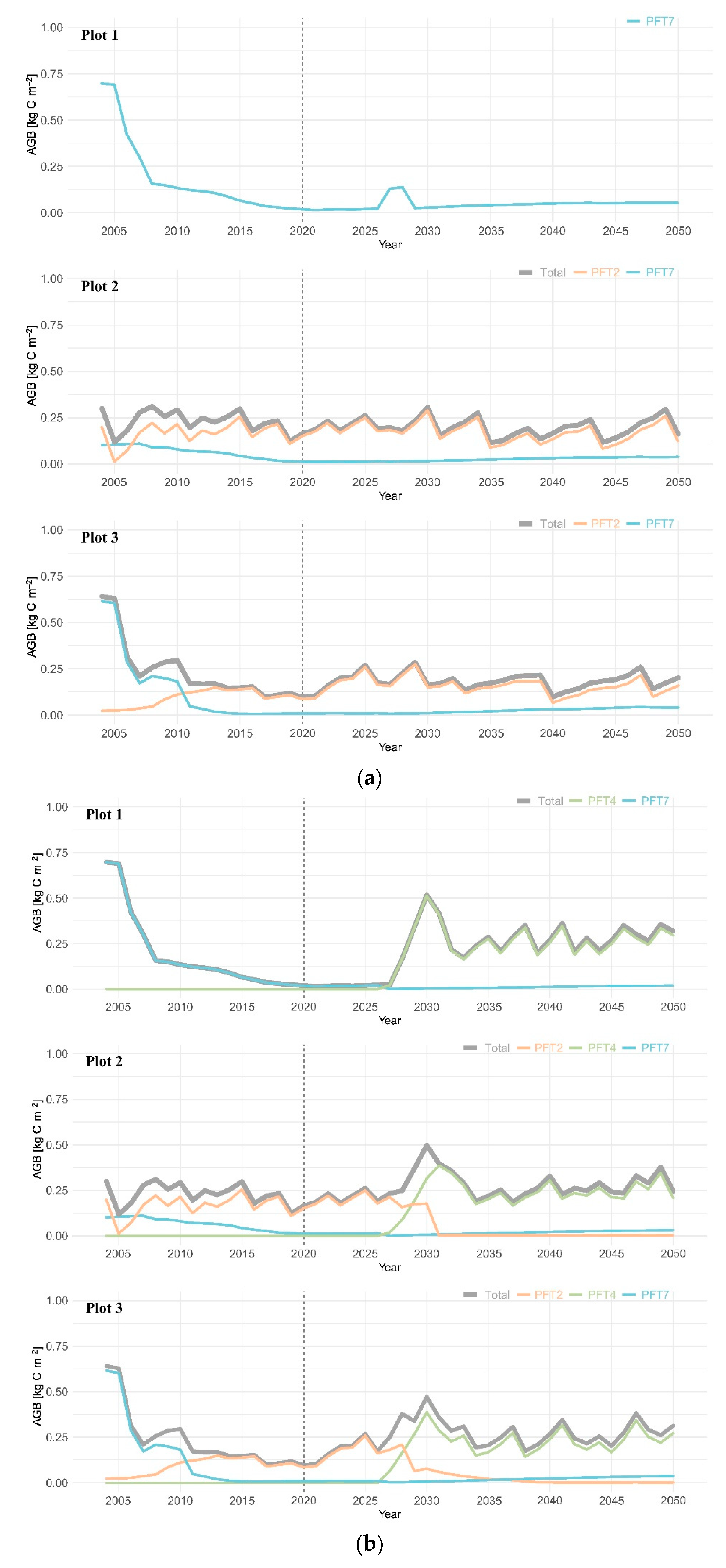
3.2. Aboveground Biomass
4. Discussion
4.1. Post-Disturbance Forest Productivity: Coniferous Forest vs. Mixed Coniferous and Broadleaved Forests
4.2. Coniferous Trees Continue to Dominate the Canopy
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A

References
- Changnon, S.A. Characteristics of ice storms in the United States. J. Appl. Meteorol. 2003, 42, 630–639. [Google Scholar] [CrossRef]
- Irland, L.C. Ice storms and forest impacts. Sci. Total Environ. 2000, 262, 231–242. [Google Scholar] [CrossRef]
- Sperry, J.S.; June, E.M.S. Xylem embolism in response to freeze-thaw cycles and water stress in ring-porous, diffuse-porous, and conifer species. Plant Physiol. 1992, 100, 605–613. [Google Scholar] [CrossRef]
- Lindenmayer, D.B.; Foster, D.R.; Franklin, J.F.; Hunter, M.L.; Noss, R.F.; Schmiegelow, F.A.; Perry, D. Ecology. Salvage harvesting policies after natural disturbance. Science 2004, 303, 1303. [Google Scholar] [CrossRef]
- Donato, D.C.; Fontaine, J.B.; Campbell, J.L.; Robinson, W.D.; Kauffman, J.B.; Law, B.E. Post-wildfire logging hinders regeneration and increases fire risk. Science 2006, 311, 352. [Google Scholar] [CrossRef] [Green Version]
- Müller, J.; Noss, R.F.; Thorn, S.; Bässler, C.; Leverkus, A.B.; Lindenmayer, D. Increasing disturbance demands new policies to conserve intact forest. Conserv. Lett. 2019, 12, e12449. [Google Scholar] [CrossRef] [Green Version]
- Thorn, S.; Bassler, C.; Brandl, R.; Burton, P.J.; Cahall, R.; Campbell, J.L.; Castro, J.; Choi, C.Y.; Cobb, T.; Donato, D.C.; et al. Impacts of salvage logging on biodiversity: A meta-analysis. J. Appl. Ecol. 2018, 55, 279–289. [Google Scholar] [CrossRef] [PubMed]
- Fahey, R.T.; Stuart-Haëntjens, E.J.; Gough, C.M.; De La Cruz, A.; Stockton, E.; Vogel, C.S.; Curtis, P.S. Evaluating forest subcanopy response to moderate severity disturbance and contribution to ecosystem-level productivity and resilience. For. Ecol. Manag. 2016, 376, 135–147. [Google Scholar] [CrossRef] [Green Version]
- Stuart-Haëntjens, E.; Curtis, P.S.; Fahey, R.T.; Vogel, C.S.; Gough, C.M. Net primary production of a temperate deciduous forest exhibits a threshold response to increasing disturbance severity. Ecology 2015, 96, 2478–2487. [Google Scholar] [CrossRef]
- Sun, Y.; Gu, L.; Dickinson, R.E.; Zhou, B. Forest greenness after the massive 2008 Chinese ice storm: Integrated effects of natural processes and human intervention. Environ. Res. Lett. 2012, 7, 035702. [Google Scholar] [CrossRef] [Green Version]
- Zhou, B.; Wang, X.; Cao, Y.; Ge, X.; Gu, L.; Meng, J. Damage assessment to subtropical forests following the 2008 Chinese ice storm. Iforest-Biogeosciences For. 2017, 10, 406–415. [Google Scholar] [CrossRef] [Green Version]
- Cao, Y.; Zhou, B.; Wang, X.; Gu, L. Resprouting responses dynamics of schima superba following a severe ice storm in early 2008 in southern China: A six-year study. Forests 2020, 11, 184. [Google Scholar] [CrossRef] [Green Version]
- Holzmueller, E.J.; Gibson, D.J.; Suchecki, P.F. Accelerated succession following an intense wind storm in an oak-dominated forest. For. Ecol. Manag. 2012, 279, 141–146. [Google Scholar] [CrossRef]
- Covey, K.R.; Barrett, A.L.; Ashton, M.S. Ice storms as a successional pathway for Fagus grandifolia advancement in Quercus rubra dominated forests of southern New England. Can. J. For. Res. 2015, 45, 1628–1635. [Google Scholar] [CrossRef]
- Lu, D.; Pile, L.S.; Yu, D.; Zhu, J.; Bragg, D.C.; Wang, G.G. Differential responses of tree species to a severe ice storm and their implications to forest composition in the southeast United States. For. Ecol. Manag. 2020, 468, 118177. [Google Scholar] [CrossRef]
- Zhao, H.; Li, Z.; Zhou, G.; Qiu, Z.; Wu, Z. Aboveground biomass allometric models for evergreen broad-leaved forest damaged by a serious ice storm in southern China. Forests 2020, 11, 320. [Google Scholar] [CrossRef] [Green Version]
- Plotkin, A.B.; Foster, D.; Carlson, J.; Magill, A. Survivors, not invaders, control forest development following simulated hurricane. Ecology 2013, 94, 414–423. [Google Scholar] [CrossRef] [Green Version]
- Rustad, L.E.; Campbell, J.L.; Driscoll, C.T.; Fahey, T.J.; Groffman, P.M.; Schaberg, P.G.; Hawley, G.J.; Halm, I.; Bowles, F.; Leuenberger, W.; et al. Experimental approach and initial forest response to a simulated ice storm experiment in a northern hardwood forest. PLoS ONE 2020, 15, e0239619. [Google Scholar] [CrossRef]
- Cheng, C.S.; Li, G.; Auld, H. Possible impacts of climate change on freezing rain using downscaled future climate scenarios: Updated for eastern Canada. Atmos.-Ocean 2011, 49, 8–21. [Google Scholar] [CrossRef]
- Lemon, P.C. Forest ecology of ice storms. Bull. Torrey Bot. Club 1961, 88, 21–29. [Google Scholar]
- Everham, E.M.; Brokaw, N.V.L. Forest damage and recovery from catastrophic wind. Bot. Rev. 1996, 62, 113–185. [Google Scholar]
- Proulx, O.J.; Greene, D.F. The relationship between ice thickness and northern hardwood tree damage during ice storms. Can. J. For. Res. 2001, 31, 1758–1767. [Google Scholar] [CrossRef]
- Frelich, L.E. Forest Dynamics and Disturbance Regimes: Studies from Temperate Evergreen-Deciduous Forests, Cambridge Studies in Ecology; Cambridge University Press: New York, NY, USA, 2002. [Google Scholar]
- Rhoads, A.G.; Hamburg, S.P.; Fahey, T.J.; Siccama, T.G.; Hane, E.N.; Battles, J.; Cogbill, C.; Randall, J.; Wilson, G. Effects of an intense ice storm on the structure of a northern hardwood forest. Can. J. For. Res. 2002, 32, 1763–1775. [Google Scholar] [CrossRef] [Green Version]
- Xi, W.; Peet, R.K.; Urban, D.L. Changes in forest structure, species diversity and spatial pattern following hurricane disturbance in a piedmont North Carolina forest, USA. J. Plant Ecol. 2008, 1, 43–57. [Google Scholar] [CrossRef]
- Allen, M.S.; Thapa, V.; Arévalo, J.R.; Palmer, M.W. Windstorm damage and forest recovery: Accelerated succession, stand structure, and spatial pattern over 25 years in two Minnesota forests. Plant Ecol. 2012, 213, 1833–1842. [Google Scholar] [CrossRef]
- Boerner, R.E.J.; Runge, S.D.; Cho, D.; Kooser, J.G. Localized ice storm damage in an Appalachian Plateau watershed. Am. Midl. Nat. 1988, 119, 199–208. [Google Scholar] [CrossRef]
- Lindenmayer, D.B.; Noss, R.F. Salvage logging, ecosystem processes, and biodiversity conservation. Conserv. Biol. 2006, 20, 949–958. [Google Scholar] [CrossRef]
- Thorn, S.; Chao, A.; Georgiev, K.B.; Muller, J.; Bassler, C.; Campbell, J.L.; Castro, J.; Chen, Y.H.; Choi, C.Y.; Cobb, T.P.; et al. Estimating retention benchmarks for salvage logging to protect biodiversity. Nat. Commun. 2020, 11, 4762. [Google Scholar]
- Zhou, B.; Gu, L.; Ding, Y.; Shao, L.; Wu, Z.; Yang, X.; Li, C.; Li, Z.; Wang, X.; Cao, Y.; et al. The great 2008 Chinese ice storm: Its socioeconomic–ecological impact and sustainability lessons learned. Bull. Am. Meteorol. Soc. 2011, 92, 47–60. [Google Scholar] [CrossRef] [Green Version]
- Piao, S.; Fang, J.; Ciais, P.; Peylin, P.; Huang, Y.; Sitch, S.; Wang, T. The carbon balance of terrestrial ecosystems in China. Nature 2009, 458, 1009–1013. [Google Scholar] [CrossRef]
- Wang, S.; Qi, C.; Li, Z. Hunan Forests; China Forestry Press: Beijing, China, 1991. [Google Scholar]
- Yang, F.; Lu, S.; Wang, B. Value estimation of service function of forest ecosystem damaged by frozen rain and snow in the south China. Sci. Silvae Sin. 2008, 44, 101–110. [Google Scholar]
- Bazzaz, F.A. The physiological ecology of plant succession. Annu. Rev. Ecol. Syst. 1979, 10, 351–371. [Google Scholar] [CrossRef] [Green Version]
- Gelaro, R.; McCarty, W.; Suarez, M.J.; Todling, R.; Molod, A.; Takacs, L.; Randles, C.; Darmenov, A.; Bosilovich, M.G.; Reichle, R.; et al. The Modern-Era Retrospective analysis for Research and Applications, version 2 (MERRA-2). J. Clim. 2017, 30, 5419–5454. [Google Scholar] [CrossRef]
- GMAO. MERRA-2 tavg1_2d_slv_nx: 2d, 1-Hourly, Time-Averaged, Single-Level, Assimilation, Single-Level Diagnostics v5.12.4; 2015. Available online: https://disc.gsfc.nasa.gov/datasets/M2T1NXSLV_5.12.4/summary (accessed on 7 January 2022).
- GMAO. MERRA-2 tavg1_2d_rad_nx: 2d, 1-Hourly, Time-Averaged, Single-Level, Assimilation, Radiation Diagnostics v5.12.4; 2015. Available online: https://disc.gsfc.nasa.gov/datasets/M2T1NXRAD_5.12.4/summary (accessed on 7 January 2022).
- GMAO. MERRA-2 tavg1_2d_lnd_nx: 2d, 1-Hourly, Time-Averaged, Single-Level, Assimilation, Land Surface Diagnostics v5.12.4; 2015. Available online: https://disc.gsfc.nasa.gov/datasets/M2T1NXLND_5.12.4/summary (accessed on 7 January 2022).
- GMAO. MERRA-2 tavg1_2d_flx_nx: 2d, 1-Hourly, Time-Averaged, Single-Level, Assimilation, Surface Flux Diagnostics v5.12.4; 2015. Available online: https://disc.gsfc.nasa.gov/datasets/M2T1NXFLX_5.12.4/summary (accessed on 7 January 2022).
- Poggio, L.; de Sousa, L.M.; Batjes, N.H.; Heuvelink, G.B.M.; Kempen, B.; Ribeiro, E.; Rossiter, D. Soilgrids 2.0: Producing soil information for the globe with quantified spatial uncertainty. Soil 2021, 7, 217–240. [Google Scholar] [CrossRef]
- Pelletier, J.D.; Broxton, P.D.; Hazenberg, P.; Zeng, X.; Troch, P.A.; Niu, G.Y.; Williams, Z.; Brunke, M.A.; Gochis, D. A gridded global data set of soil, intact regolith, and sedimentary deposit thicknesses for regional and global land surface modeling. J. Adv. Modeling Earth Syst. 2016, 8, 41–65. [Google Scholar] [CrossRef]
- Longo, M.; Knox, R.G.; Medvigy, D.M.; Levine, N.M.; Dietze, M.C.; Kim, Y.; Swann, A.L.S.; Zhang, K.; Rollinson, C.R.; Bras, R.L.; et al. The biophysics, ecology, and biogeochemistry of functionally diverse, vertically and horizontally heterogeneous ecosystems: The Ecosystem Demography model, version 2.2—Part 1: Model description. Geosci. Model Dev. 2019, 12, 4309–4346. [Google Scholar] [CrossRef] [Green Version]
- Medvigy, D.; Moorcroft, P.R. Predicting ecosystem dynamics at regional scales: An evaluation of a terrestrial biosphere model for the forests of northeastern North America. Philos. Trans. R. Soc. B Biol. Sci. 2012, 367, 222–235. [Google Scholar] [CrossRef] [Green Version]
- Medvigy, D.; Wofsy, S.C.; Munger, J.W.; Hollinger, D.Y.; Moorcroft, P.R. Mechanistic scaling of ecosystem function and dynamics in space and time: Ecosystem Demography model version 2. J. Geophys. Res. 2009, 114, G01002. [Google Scholar] [CrossRef] [Green Version]
- Moorcroft, P.R.; Hurtt, G.C.; Pacala, S.W. A method for scaling vegetation dynamics: The Ecosystem Demography model (ed). Ecol. Monogr. 2001, 71, 557–586. [Google Scholar] [CrossRef]
- Powell, T.L.; Galbraith, D.R.; Christoffersen, B.O.; Harper, A.; Imbuzeiro, H.M.A.; Rowland, L.; Almeida, S.; Brando, P.M.; da Costa, A.C.L.; Costa, M.H.; et al. Confronting model predictions of carbon fluxes with measurements of Amazon forests subjected to experimental drought. New Phytol. 2013, 200, 350–365. [Google Scholar] [PubMed] [Green Version]
- Zhang, K.; de Almeida Castanho, A.D.; Galbraith, D.R.; Moghim, S.; Levine, N.M.; Bras, R.L.; Coe, M.T.; Costa, M.H.; Malhi, Y.; Longo, M.; et al. The fate of Amazonian ecosystems over the coming century arising from changes in climate, atmospheric CO2, and land use. Glob. Change Biol. 2015, 21, 2569–2587. [Google Scholar] [CrossRef] [PubMed]
- Levine, N.M.; Zhang, K.; Longo, M.; Baccini, A.; Phillips, O.L.; Lewis, S.L.; Alvarez-Davila, E.; Segalin de Andrade, A.C.; Brienen, R.J.; Erwin, T.L.; et al. Ecosystem heterogeneity determines the ecological resilience of the Amazon to climate change. Proc. Natl. Acad. Sci. USA 2016, 113, 793–797. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Longo, M.; Saatchi, S.; Keller, M.; Bowman, K.; Ferraz, A.; Moorcroft, P.R.; Morton, D.C.; Bonal, D.; Brando, P.; Burban, B.; et al. Impacts of degradation on water, energy, and carbon cycling of the Amazon tropical forests. J. Geophys. Res. Biogeosci. 2020, 125, e2020JG005677. [Google Scholar] [CrossRef] [PubMed]
- Fer, I.; Kelly, R.; Moorcroft, P.R.; Richardson, A.D.; Cowdery, E.M.; Dietze, M.C. Linking big models to big data: Efficient ecosystem model calibration through Bayesian model emulation. Biogeosciences 2018, 15, 5801–5830. [Google Scholar] [CrossRef] [Green Version]
- The Forestry Department of Hunan Province. Hunan Provincial Protocol of National Forest Inventory of China; The Forestry Department of Hunan Province: Changsha, China, 2014. [Google Scholar]
- Kumar, U.; Singh, P.; Boote, K.J. Chapter two-Effect of climate change factors on processes of crop growth and development and yield of groundnut (Arachis hypogaea L.). Advances in Agronomy 2012, 116, 41–69. [Google Scholar]
- Zhao, M.; Xiang, W.; Peng, C.; Tian, D. Simulating age-related changes in carbon storage and allocation in a Chinese fir plantation growing in southern China using the 3-PG model. For. Ecol. Manag. 2009, 257, 1520–1531. [Google Scholar] [CrossRef]
- Kattge, J.; DÍAz, S.; Lavorel, S.; Prentice, I.C.; Leadley, P.; BÖNisch, G.; Garnier, E.; Westoby, M.; Reich, P.B.; Wright, I.J.; et al. Try—A global database of plant traits. Glob. Change Biol. 2011, 17, 2905–2935. [Google Scholar] [CrossRef]
- Tian, D. Chinese Ecosystem Positioning Observation and Research Data-Forest Ecosystem Volume: Huitong National Station for Scientific Observation and Research of Chinese Fir Plantation Ecosystems (1982~2009); China Agricultural Press: Beijing, China, 2011. [Google Scholar]
- Feng, Z.; Chen, C.; Zhang, J.; Zeng, S. A coniferous broad-leaved mixed forest with higher productivity and ecological harmony in subtropics-study on mixed forests of Cunninghamia lanceolata and Michelia macclurei. Acta Phytoecol. Et Geobot. Sin. 1988, 12, 165–180. [Google Scholar]
- Tian, D.; Kang, W.; Wen, S. Chinese Fir Ecosystems; Science Press: Beijing, China, 2003. [Google Scholar]
- Chen, C.; Zhang, J.; Zhou, C.; Zheng, H. Researches on improving the quality of forest land and the productivity of artificial cunninghamia lanceolata stands. Chin. J. Appl. Ecol. 1990, 1, 97–106. [Google Scholar]
- Reichstein, M.; Bahn, M.; Ciais, P.; Frank, D.; Mahecha, M.D.; Seneviratne, S.I.; Zscheischler, J.; Beer, C.; Buchmann, N.; Frank, D.C.; et al. Climate extremes and the carbon cycle. Nature 2013, 500, 287–295. [Google Scholar] [CrossRef]
- Campbell, J.L.; Rustad, L.E.; Driscoll, C.T.; Halm, I.; Fahey, T.J.; Fakhraei, H.; Groffman, P.M.; Hawley, G.J.; Leuenberger, W.; Schaberg, P.G. Simulating impacts of ice storms on forest ecosystems. J. Vis. Exp. 2020, 160, e61492. [Google Scholar] [CrossRef]
- Ouyang, S.; Xiang, W.; Wang, X.; Xiao, W.; Chen, L.; Li, S.; Sun, H.; Deng, X.; Forrester, D.I.; Zeng, L.; et al. Effects of stand age, richness and density on productivity in subtropical forests in china. J. Ecol. 2019. [Google Scholar] [CrossRef]
- Janzen, D.H. Herbivores and the number of tree species in tropical forests. Am. Nat. 1970, 104, 501–528. [Google Scholar] [CrossRef]
- Connell, J.H. On the Role of Natural Enemies in Preventing Competitive Exclusion in Some Marine Animals and in Rain Forest Trees; Centre for Agricultural Publication and Documentation: Wageningen, The Netherlands, 1971. [Google Scholar]
- Paine, C.E.; Norden, N.; Chave, J.; Forget, P.M.; Fortunel, C.; Dexter, K.G.; Baraloto, C. Phylogenetic density dependence and environmental filtering predict seedling mortality in a tropical forest. Ecol. Lett. 2012, 15, 34–41. [Google Scholar] [CrossRef] [PubMed]
- Lebrija-Trejos, E.; Wright, S.J.; Hernandez, A.; Reich, P.B. Does relatedness matter? Phylogenetic density-dependent survival of seedlings in a tropical forest. Ecology 2014, 95, 940–951. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Liang, M.; Etienne, R.S.; Wang, Y.; Staehelin, C.; Yu, S. Experimental evidence for a phylogenetic Janzen-Connell effect in a subtropical forest. Ecol. Lett. 2012, 15, 111–118. [Google Scholar] [CrossRef] [PubMed]
- Forrester, D.I.; Albrecht, A.T. Light absorption and light-use efficiency in mixtures of Abies alba and Picea abies along a productivity gradient. For. Ecol. Manag. 2014, 328, 94–102. [Google Scholar] [CrossRef]
- Amiro, B.D.; Barr, A.G.; Barr, J.G.; Black, T.A.; Bracho, R.; Brown, M.; Chen, J.; Clark, K.L.; Davis, K.J.; Desai, A.R.; et al. Ecosystem carbon dioxide fluxes after disturbance in forests of North America. J. Geophys. Res. 2010, 115, G00K02. [Google Scholar] [CrossRef]
- Hanson, J.J.; Lorimer, C.G. Forest structure and light regimes following moderate wind storms: Implications for multi-cohort management. Ecol. Appl. 2007, 17, 1325–1340. [Google Scholar] [CrossRef]
- Cooper-Ellis, S.; Foster, D.R.; Carlton, G.; Lezberg, A. Forest response to catastrophic wind: Results from an experimental hurricane. Ecology 1999, 80, 2683–2696. [Google Scholar] [CrossRef]
- Woods, K.D. Intermediate disturbance in a late-successional hemlock-northern hardwood forest. J. Ecol. 2004, 92, 464–476. [Google Scholar] [CrossRef]
- Nave, L.E.; Gough, C.M.; Maurer, K.D.; Bohrer, G.; Hardiman, B.S.; Le Moine, J.; Munoz, A.B.; Nadelhoffer, K.J.; Sparks, J.P.; Strahm, B.D.; et al. Disturbance and the resilience of coupled carbon and nitrogen cycling in a north temperate forest. J. Geophys. Res. 2011, 116, G04016. [Google Scholar] [CrossRef] [Green Version]
- Flower, C.E.; Gonzalez-Meler, M.A. Responses of temperate forest productivity to insect and pathogen disturbances. Annu. Rev. Plant Biol. 2015, 66, 547–569. [Google Scholar] [CrossRef] [PubMed]
- Gough, C.M.; Curtis, P.S.; Hardiman, B.S.; Scheuermann, C.M.; Bond-Lamberty, B. Disturbance, complexity, and succession of net ecosystem production in North America’s temperate deciduous forests. Ecosphere 2016, 7, e01375. [Google Scholar] [CrossRef]
- Halpin, C.R.; Lorimer, C.G. Trajectories and resilience of stand structure in response to variable disturbance severities in northern hardwoods. For. Ecol. Manag. 2016, 365, 69–82. [Google Scholar] [CrossRef]
- Gough, C.M.; Hardiman, B.S.; Nave, L.; Bohrer, G.; Maurer, K.D.; Vogel, C.S. Sustained carbon uptake and storage following moderate disturbance in a Great Lakes forest. Ecol. Appl. 2013, 23, 1202–1215. [Google Scholar] [CrossRef] [Green Version]
- Connell, J.H. Diversity in tropical rain forests and coral reefs. Science 1978, 199, 1302–1310. [Google Scholar] [CrossRef] [Green Version]
- Roxburgh, S.H.; Shea, K.; Wilson, J.B. The intermediate disturbance hypothesis: Patch dynamics and mechanisms of species coexistence. Ecology 2004, 85, 359–371. [Google Scholar] [CrossRef]
- Loreau, M.; Hector, A. Partitioning selection and complementarity in biodiversity experiments. Nature 2001, 412, 72–76. [Google Scholar] [CrossRef]
- Ratcliffe, S.; Wirth, C.; Jucker, T.; van der Plas, F.; Scherer-Lorenzen, M.; Verheyen, K.; Allan, E.; Benavides, R.; Bruelheide, H.; Ohse, B.; et al. Biodiversity and ecosystem functioning relations in European forests depend on environmental context. Ecol. Lett. 2017, 20, 1414–1426. [Google Scholar] [CrossRef]
- Dudley, N.; Schlaepfer, R.; Jackson, W. Forest Quality: Assessing Forests at a Landscape Scale; Earthscan: London, UK, 2006. [Google Scholar]
- Nilsson, M.-C.; Wardle, D.A. Understory vegetation as a forest ecosystem driver: Evidence from the northern Swedish boreal forest. Front. Ecol. Environ. 2005, 3, 421–428. [Google Scholar] [CrossRef]
- Zackrisson, O.; Nilsson, L.O.; Wardle, D.A. Key ecological function of charcoal from wildfire in the boreal forest. Oikos 1996, 77, 10–19. [Google Scholar] [CrossRef]
- Hart, S.A.; Chen, H.Y.H. Understory vegetation dynamics of North American boreal forests. Crit. Rev. Plant Sci. 2006, 25, 381–397. [Google Scholar] [CrossRef]
- Xiang, W.; Li, L.; Ouyang, S.; Xiao, W.; Zeng, L.; Chen, L.; Lei, P.; Deng, X.; Zeng, Y.; Fang, J.; et al. Effects of stand age on tree biomass partitioning and allometric equations in Chinese fir (Cunninghamia lanceolata) plantations. Eur. J. For. Res. 2020, 140, 317–332. [Google Scholar] [CrossRef]
- Zhou, X.; Yang, M.; Liu, Z.; Li, P.; Xie, B.; Peng, C. Dynamic allometric scaling of tree biomass and size. Nat. Plants 2021, 7, 42–49. [Google Scholar] [CrossRef]
- Zhao, M.; Tian, S.; Zhu, Y.; Li, Z.; Zeng, S.; Liu, S. Allometric relationships, functional differentiations, and scaling of growth rates across 151 tree species in China. Ecosphere 2021, 12, e03522. [Google Scholar] [CrossRef]
- LeBauer, D.S.; Wang, D.; Richter, K.T.; Davidson, C.C.; Dietze, M.C. Facilitating feedbacks between field measurements and ecosystem models. Ecol. Monogr. 2013, 83, 133–154. [Google Scholar] [CrossRef]
- Dietze, M.C.; Serbin, S.P.; Davidson, C.; Desai, A.R.; Feng, X.; Kelly, R.; Kooper, R.; LeBauer, D.; Mantooth, J.; McHenry, K.; et al. A quantitative assessment of a terrestrial biosphere model’s data needs across North American biomes. J. Geophys. Res. Biogeosciences 2014, 119, 286–300. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Plot No. | Plot 1 | Plot 2 | Plot 3 |
|---|---|---|---|
| Elevation [m] | 450 | 110 | 440 |
| Mean age [a] | 15 | 29 | 17 |
| Mean DBH [cm] | 10 | 12 | 13 |
| Canopy height [m] | 16.6 | 12.9 | 13.1 |
| Species composition (Whole community) | CULA; PIMA | CULA; PIMA; CINN; SATZ; OBLD | CULA; PIMA; OBLD |
| Species composition (Understory) | CULA | CULA; CINN; SATZ; OBLD | CULA |
| Species composition (Recruitment 2004~2009) | Null | CULA; OBLD | CULA; CINN |
| Basal area [m2 ha−1] | CULA (24.2); PIMA (1.9) | CULA (10.7); PIMA (0.3); CINN (1.1); SATZ (1.1); OBLD (0.1) | CULA (28.1); PIMA (1.1); OBLD (0.1) |
| Stand density [ha−1] (Whole community) | 3209 | 1209 | 3104 |
| Stand density [ha−1] (Understory) | 1851 | 373 | 1343 |
| Stand density [ha−1] (Recruitment 2004~2009) | 0 | 45 | 90 |
| Salvage logging [AGB%] | 27 | 10 | 20 |
| Salvage logging [ha−1] | CULA (791); PIMA (45) | CULA (75); CINN (15); SATZ (30) | CULA (716); PIMA (15); OBLD (30) |
| Plant Functional Type | SLA [m2 kgC−1] | b1Ht [m] | b2Ht [cm−1] | b1Bs [kgC] | b2Bs |
|---|---|---|---|---|---|
| PFT2 | 24.5 | 0.035 | 0.695 | 0.166 | 2.432 |
| PFT4 | 36.6 | 0.042 | 0.522 | 0.282 | 2.432 |
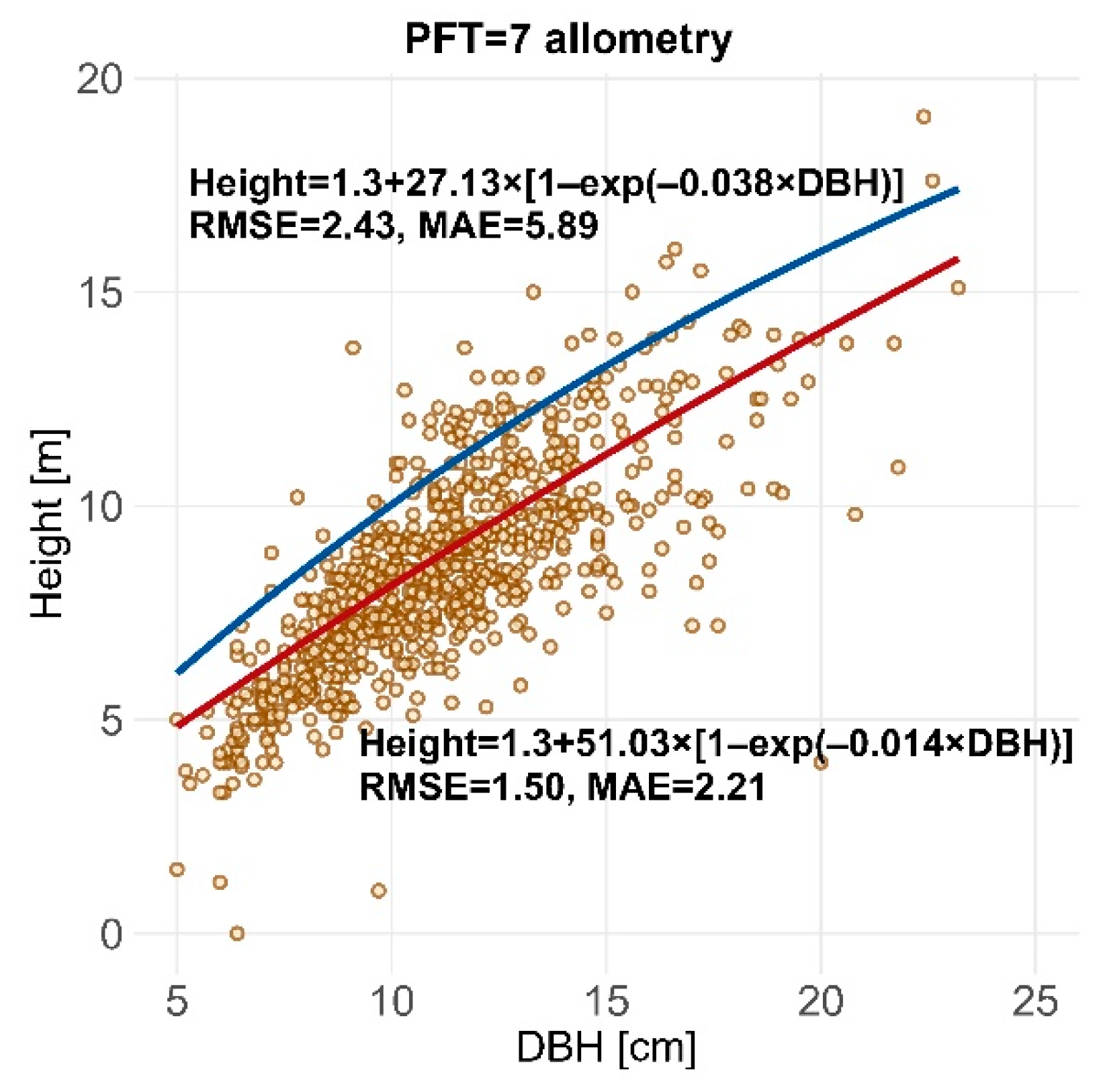
| PFT7 | 7.4 | 51.037 | −0.014 | 0.117 | 2.240 |
| Plot No. | ED-2.2 Simulation | Previous Studies | Reference | ||
|---|---|---|---|---|---|
| Min. | Mean | Max. | Range | ||
| Plot 1 | 0.54 | 0.61 | 0.71 | [0.45, 0.77] | [57] |
| Plot 2 | 0.82 | 0.95 | 1.20 | [0.69, 1.17] | [56] |
| Plot 3 | 0.84 | 0.92 | 1.02 | [0.69, 1.17] | [56] |
| Scenario | Ice-Storm and Subsequent Salvage Logging | ||
|---|---|---|---|
| 2021 | 2024 | 2027 | |
| Frequency 0 (F0, baseline) | Not occurred | Not occurred | Not occurred |
| Frequency 1 (F1) | Occurred | Not occurred | Not occurred |
| Frequency 2 (F2) | Occurred | Occurred | Not occurred |
| Frequency 3 (F3) | Occurred | Occurred | Occurred |
| Frequency 4 (F4) | Occurred | Occurred | Occurred * |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhu, Y.; Liu, S.; Yan, W.; Deng, D.; Zhou, G.; Zhao, M.; Gao, F.; Zhu, L.; Wang, Z.; Xie, M. Impact of Ice-Storms and Subsequent Salvage Logging on the Productivity of Cunninghamia lanceolata (Chinese Fir) Forests. Forests 2022, 13, 296. https://0-doi-org.brum.beds.ac.uk/10.3390/f13020296
Zhu Y, Liu S, Yan W, Deng D, Zhou G, Zhao M, Gao F, Zhu L, Wang Z, Xie M. Impact of Ice-Storms and Subsequent Salvage Logging on the Productivity of Cunninghamia lanceolata (Chinese Fir) Forests. Forests. 2022; 13(2):296. https://0-doi-org.brum.beds.ac.uk/10.3390/f13020296
Chicago/Turabian StyleZhu, Yu, Shuguang Liu, Wende Yan, Deming Deng, Guangyi Zhou, Meifang Zhao, Fei Gao, Liangjun Zhu, Zhao Wang, and Menglu Xie. 2022. "Impact of Ice-Storms and Subsequent Salvage Logging on the Productivity of Cunninghamia lanceolata (Chinese Fir) Forests" Forests 13, no. 2: 296. https://0-doi-org.brum.beds.ac.uk/10.3390/f13020296