
Biochar and/or Compost to Enhance Nursery-Produced Seedling Performance: A Potential Tool for Forest Restoration Programs

 ,
,  , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Experimental Design and Growth Characteristics
2.2. Soil, Biochar, and Compost Characteristics
2.3. Growing Substrate Analysis
2.4. Plant Analysis
2.4.1. Morphological Traits
2.4.2. Phenology
2.4.3. Lignin and Chlorophyll Content
2.5. Statistical Analysis
3. Results
3.1. Growing Substrate Characteristics
3.2. Plant Characteristics
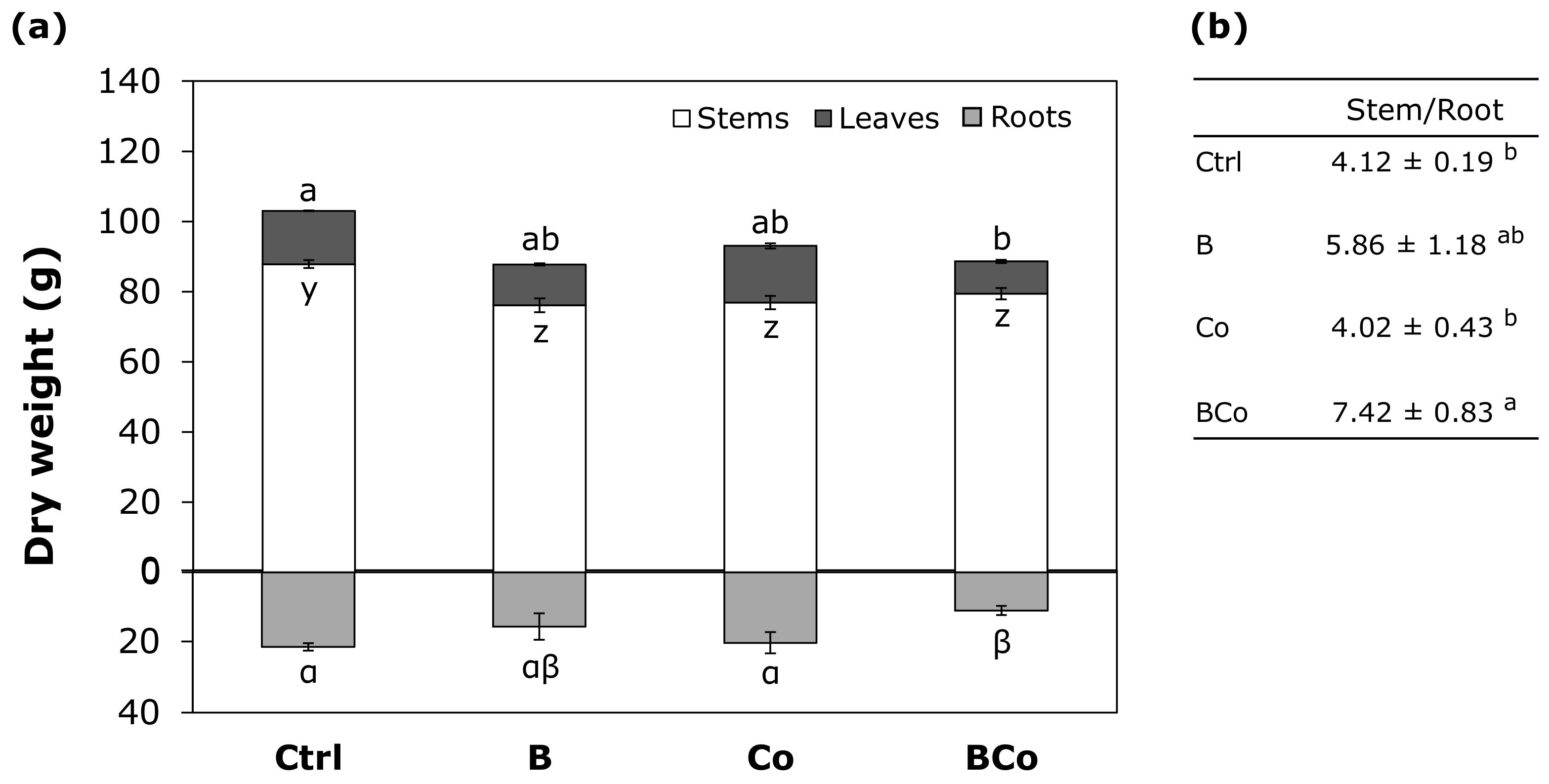
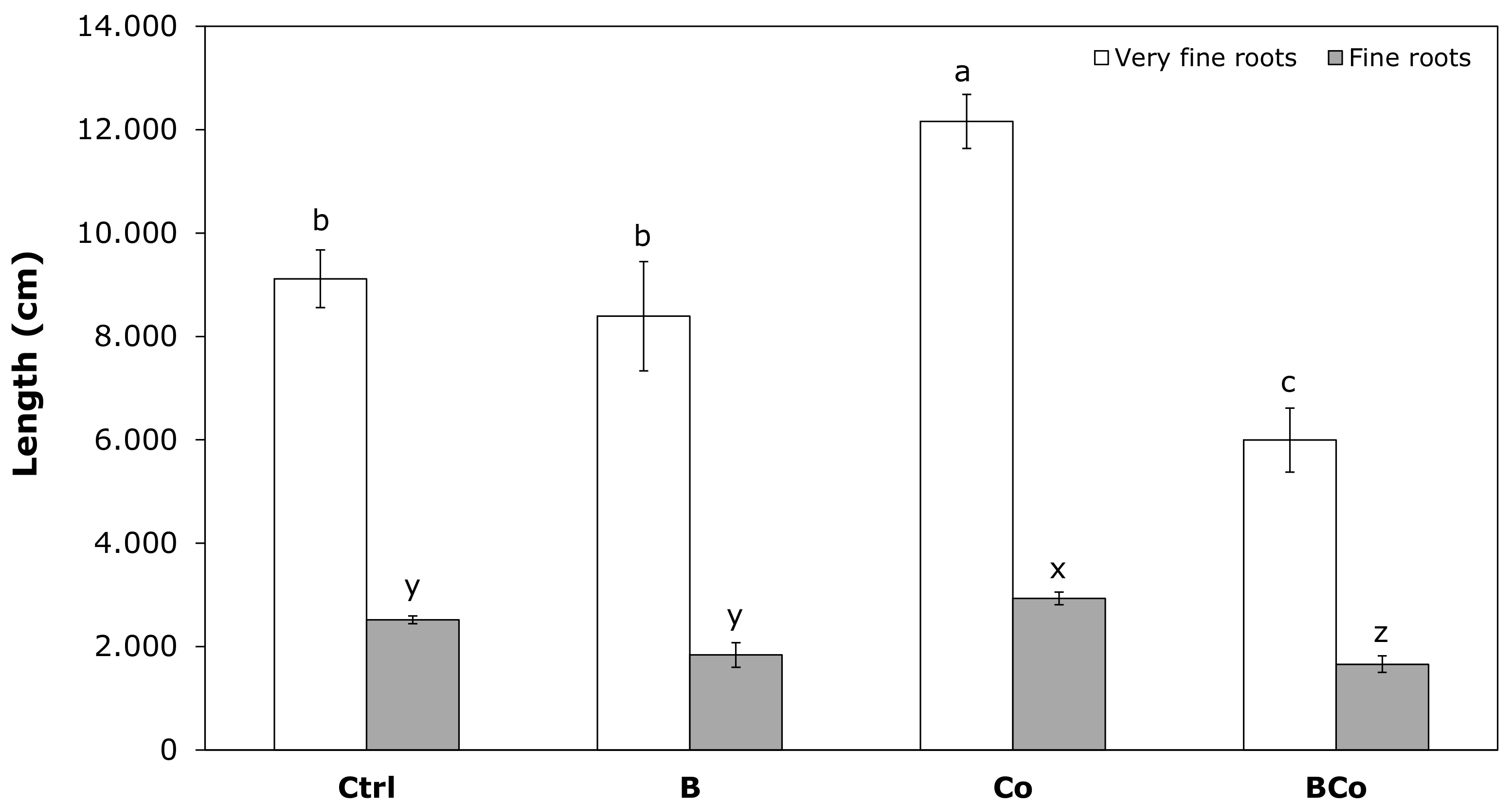
3.2.1. Morphological Traits
3.2.2. Phenology
3.2.3. Lignin and Chlorophyll Content
4. Discussion
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Li, Y.; Ye, Y.; Fang, X.; Liu, Y. Reconstruction of agriculture-driven deforestation in western Hunan province of China during the 18th century. Land 2022, 11, 181. [Google Scholar] [CrossRef]
- Skole, D.L.; Mbow, C.; Mugabowindekwe, M.; Brandt, M.S.; Samek, J.H. Trees outside of forests as natural climate solutions. Nat. Clim. Chang. 2021, 11, 1013–1016. [Google Scholar] [CrossRef]
- Hoekman, D.; Kooij, B.; Quiñones, M.; Vellekoop, S.; Carolita, I.; Budhiman, S.; Arief, R.; Roswintiarti, O. Wide-area near-real-time monitoring of tropical forest degradation and deforestation using Sentinel-1. Remote Sens. 2020, 12, 3263. [Google Scholar] [CrossRef]
- Curtis, P.G.; Slay, C.M.; Harris, N.L.; Tyukavina, A.; Hansen, M.C. Classifying drivers of global forest loss. Science 2018, 361, 1108–1111. [Google Scholar] [CrossRef]
- Ritchie, H.; Roser, M. Forests and Deforestation. 2021. Available online: https://ourworldindata.org/forests-and-deforestation (accessed on 20 January 2022).
- Breshears, D.D.; Fontaine, J.B.; Ruthrof, K.X.; Field, J.P.; Feng, X.; Burger, J.R.; Law, D.J.; Kala, J.; Hardy, G.E.S.J. Underappreciated plant vulnerabilities to heat waves. New Phytol. 2021, 231, 32–39. [Google Scholar] [CrossRef]
- Matusick, G.; Ruthrof, K.X.; Brouwers, N.C.; Dell, B.; Hardy, G.S. Sudden forest canopy collapse corresponding with extreme drought and heat in a Mediterranean-type eucalypt forest in south western Australia. Eur. J. For. Res. 2013, 132, 497–510. [Google Scholar] [CrossRef] [Green Version]
- Payne, R.J.; Anderson, A.R.; Sloan, T.; Gilbert, P.; Newton, A.; Ratcliffe, J.; Mauquoy, D.; Jessop, W.; Andersen, R. The future of peatland forestry in Scotland: Balancing economics, carbon and biodiversity. Scott. For. 2018, 100, 34–40. [Google Scholar]
- Brown, I. Challenges in delivering climate change policy through land use targets for afforestation and peatland restoration. Environ. Sci. Policy 2020, 107, 36–45. [Google Scholar] [CrossRef]
- Dumroese, R.K.; Landis, T.D.; Pinto, J.R.; Haase, D.L.; Wilkinson, K.W.; Davis, A.S. Meeting forest restoration challenges: Using the target plant concept. Reforesta 2016, 1, 37–52. [Google Scholar] [CrossRef] [Green Version]
- Mansourian, S. In the eye of the beholder: Reconciling interpretations of forest landscape restoration. Land Degrad. Dev. 2018, 29, 2888–2898. [Google Scholar] [CrossRef]
- Le, H.D.; Smith, C.; Herbohn, J. Identifying interactions among reforestation success drivers: A case study from the Philippines. Ecol. Modell. 2015, 316, 62–77. [Google Scholar] [CrossRef]
- Fargione, J.; Haase, D.L.; Burney, O.T.; Kildisheva, O.A.; Edge, G.; Cook-Patton, S.C.; Chapman, T.; Rempel, A.; Hurteau, M.D.; Davis, K.T.; et al. Challenges to the reforestation pipeline in the United States. Front. For. Glob. Chang. 2021, 4, 8. [Google Scholar] [CrossRef]
- North, M.P.; Stevens, J.T.; Greene, D.F.; Coppoletta, M.; Knapp, E.E.; Latimer, A.M.; Restaino, C.M.; Tompkins, R.E.; Welch, K.R.; York, R.A.; et al. Tamm Review: Reforestation for resilience in dry western US forests. For. Ecol. Manag. 2019, 432, 209–224. [Google Scholar] [CrossRef] [Green Version]
- Puettmann, M.; Sahoo, K.; Wilson, K.; Oneil, E. Life cycle assessment of biochar produced from forest residues using portable systems. J. Clean. Prod. 2020, 250, 119564. [Google Scholar] [CrossRef]
- Davis, A.S.; Pinto, J.R. The scientific basis of the Target Plant Concept: An overview. Forests 2021, 12, 1293. [Google Scholar] [CrossRef]
- Andivia, E.; Villar-Salvador, P.; Oliet, J.A.; Puértolas, J.; Dumroese, R.K. How can my research paper be useful for future meta-analyses on forest restoration plantations? New For. 2019, 50, 255–266. [Google Scholar] [CrossRef]
- Meli, P.; Isernhagen, I.; Brancalion, P.H.; Isernhagen, E.C.; Behling, M.; Rodrigues, R.R. Optimizing seeding density of fast-growing native trees for restoring the Brazilian Atlantic Forest. Restor. Ecol. 2018, 26, 212–219. [Google Scholar] [CrossRef]
- Brancalion, P.H.; Holl, K.D. Guidance for successful tree planting initiatives. J. Appl. Ecol. 2020, 57, 2349–2361. [Google Scholar] [CrossRef]
- Drake, J.A.; Carrucan, A.; Jackson, W.R.; Cavagnaro, T.R.; Patti, A.F. Biochar application during reforestation alters species present and soil chemistry. Sci. Total Environ. 2015, 514, 359–365. [Google Scholar] [CrossRef] [Green Version]
- Montagnoli, A.; Dumroese, R.K.; Terzaghi, M.; Pinto, J.R.; Fulgaro, N.; Scippa, G.S.; Chiatante, D. Tree seedling response to LED spectra: Implications for forest restoration. Plant Biosyst.-Int. J. Deal. All Asp. Plant Biol. 2018, 152, 515–523. [Google Scholar] [CrossRef]
- Barrow, C. Biochar: Potential for countering land degradation and for improving agriculture. Appl. Geogr. 2012, 34, 21–28. [Google Scholar] [CrossRef]
- Lefebvre, D.; Román-Dañobeytia, F.; Soete, J.; Cabanillas, F.; Corvera, R.; Ascorra, C.; Fernandez, L.E.; Silman, M. Biochar effects on two tropical tree species and its potential as a tool for reforestation. Forests 2019, 10, 678. [Google Scholar] [CrossRef] [Green Version]
- Lehmann, J.; Rillig, M.; Thies, J.; Masiello, C.; Hockaday, W.; Crowley, D. Biochar effects on soil biota—A review. Soil Biol. Biochem. 2011, 43, 1812–1836. [Google Scholar] [CrossRef]
- Woolf, D.; Amonette, J.E.; Street-Perrott, F.A.; Lehmann, J.; Joseph, S. Sustainable biochar to mitigate global climate change. Nat. Commun. 2010, 1, 56. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kameyama, K.; Miyamoto, T.; Iwata, Y.; Shiono, T. Influences of feedstock and pyrolysis temperature on the nitrate adsorption of biochar. Soil Sci. Plant Nutr. 2016, 62, 180–184. [Google Scholar] [CrossRef]
- Kavitha, B.; Reddy, P.V.L.; Kim, B.; Lee, S.S.; Pandey, S.K.; Kim, K.H. Benefits and limitations of biochar amendment in agricultural soils: A review. J. Environ. Manag. 2018, 227, 146–154. [Google Scholar] [CrossRef]
- Thomas, S.C.; Gale, N. Biochar and forest restoration: A review and meta-analysis of tree growth responses. New For. 2015, 46, 931–946. [Google Scholar] [CrossRef]
- Karim, M.; Halim, M.A.; Gale, N.V.; Thomas, S.C. Biochar effects on soil physiochemical properties in degraded managed ecosystems in northeastern Bangladesh. Soil Syst. 2020, 4, 69. [Google Scholar] [CrossRef]
- Wu, S.; He, H.; Inthapanya, X.; Yang, C.; Lu, L.; Zeng, G.; Han, Z. Role of biochar on composting of organic wastes and remediation of contaminated soils—A review. Environ. Sci. Pollut. Res. 2017, 24, 16560–16577. [Google Scholar] [CrossRef]
- Lal, R. Restoring soil quality to mitigate soil degradation. Sustainability 2015, 7, 5875–5895. [Google Scholar] [CrossRef] [Green Version]
- Saer, A.; Lansing, S.; Davitt, N.H.; Graves, R.E. Life cycle assessment of a food waste composting system: Environmental impact hotspots. J. Clean. Prod. 2013, 52, 234–244. [Google Scholar] [CrossRef]
- Post, W.M.; Kwon, K.C. Soil carbon sequestration and land-use change: Processes and potential. Glob. Chang. Biol. 2000, 6, 317–327. [Google Scholar] [CrossRef] [Green Version]
- Diacono, M.; Montemurro, F. Long-term effects of organic amendments on soil fertility: A review. Agron. Sustain. Dev. 2010, 30, 401–422. [Google Scholar] [CrossRef] [Green Version]
- Persiani, A.; Diacono, M.; Monteforte, A.; Montemurro, F. Agronomic performance, energy analysis and carbon balance comparing different fertilization strategies in horticulture under Mediterranean conditions. Environ. Sci. Pollut. Res. 2019, 26, 19250–19260. [Google Scholar] [CrossRef] [PubMed]
- Pane, C.; Palese, A.M.; Spaccini, R.; Piccolo, A.; Celano, G.; Zaccardelli, M. Enhancing sustainability of a processing tomato cultivation system by using bioactive compost teas. Sci. Hort. 2016, 202, 117–124. [Google Scholar] [CrossRef]
- Martínez-Blanco, J.; Muñoz, P.; Anton, A.; Rieradevall, J. Life cycle assessment of the use of compost from municipal organic waste for fertilization of tomato crops. Resour. Conserv. Recycl. 2009, 53, 340–351. [Google Scholar] [CrossRef]
- Steiner, C.; Das, K.C.; Melear, N.; Lakly, D. Reducing nitrogen loss during poultry litter composting using biochar. J. Environ. Qual. 2010, 39, 1236–1242. [Google Scholar] [CrossRef] [Green Version]
- Fischer, D.; Glaser, B. Synergisms between compost and biochar for sustainable soil amelioration. In Management of Organic Waste; Kumar, S., Ed.; InTech: Rijeka, Croatia, 2012; pp. 167–198. [Google Scholar]
- Liu, J.; Schulz, H.; Brandl, S.; Miehtke, H.; Huwe, B.; Glaser, B. Short-Term effect of biochar and compost on soil fertility and water status of a Dystric Cambisol in NE Germany under field conditions. J. Plant Nutr. Soil Sci. 2012, 175, 698–707. [Google Scholar] [CrossRef]
- Grossnickle, S.C.; MacDonald, J.E. Seedling quality: History, application, and plant attributes. Forests 2018, 9, 283. [Google Scholar] [CrossRef] [Green Version]
- Grossnickle, S.C. Why seedlings survive: Importance of plant attributes. New For. 2012, 43, 711–738. [Google Scholar] [CrossRef]
- Grossnickle, S.C.; South, D.B. Seedling quality of southern pines: Influence of plant attributes. Tree Plant. Notes 2017, 60, 29–40. [Google Scholar]
- Pinto, J.R. Morphology targets: What do seedling morphological attributes tell us. In National Proceedings: Forest and Conservation Nursery Associations-2010; RMRS-P-65; U.S. Department of Agriculture, Forest Service: Fort Collins, CO, USA, 2011; pp. 74–79. [Google Scholar]
- Jha, K.K. Root carbon sequestration and its efficacy in forestry and agroforestry systems: A case of Populus euramericana I-214 cultivated in Mediterranean condition. Not. Sci. Biol. 2018, 10, 68–78. [Google Scholar] [CrossRef] [Green Version]
- Fuertes, A.; Oliveira, N.; Cañellas, I.; Sixto, H.; Rodríguez-Soalleiro, R. An economic overview of Populus spp. in Short Rotation Coppice systems under Mediterranean conditions: An assessment tool for decision-making. Renew. Sustain. Energy Rev. 2021, 151, 111577. [Google Scholar] [CrossRef]
- Pallardy, S.G.; Gibbins, D.E.; Rhoads, J.L. Biomass production by two-year-old poplar clones on floodplain sites in the Lower Midwest, USA. Agrofor. Syst. 2003, 59, 21–26. [Google Scholar] [CrossRef]
- Nielsen, U.B.; Madsen, P.; Hansen, J.K.; Nord-Larsen, T.; Nielsen, A.T. Production potential of 36 poplar clones grown at medium length rotation in Denmark. Biomass Bioenergy 2014, 64, 99–109. [Google Scholar] [CrossRef]
- Giri, A.; Kumar, G.; Arya, R.; Mishra, S.; Mishra, A.K. Carbon sequestration in Populus deltoides based agroforestry system in northern India. Int. J. Chem. Stud. 2019, 7, 2184–2188. [Google Scholar]
- VanWallendael, A.; Lowry, D.B.; Hamilton, J.A. One hundred years into the study of ecotypes, new advances are being made through large-scale field experiments in perennial plant systems. Curr. Plant Biol. 2022, 66, 102152. [Google Scholar] [CrossRef] [PubMed]
- U.S Department of Agriculture, Natural Resources Conservation Service. National Soil Survey Handbook. Available online: http://www.nrcs.usda.gov/wps/portal/nrcs/detail/soils/ref/?cid=nrcs142p2_054242 (accessed on 27 January 2022).
- Alfano, G.; Lustrato, G.; Lima, G.; Vitullo, D.; Ranalli, G. Characterization of composted olive mill wastes to predict potential plant disease suppressiveness. Biol. Control 2011, 58, 199–207. [Google Scholar] [CrossRef]
- Trupiano, D.; Cocozza, C.; Baronti, S.; Amendola, C.; Vaccari, F.P.; Lustrato, G.; Di Lonardo, S.; Fantasma, F.; Tognetti, R.; Scippa, G.S. The effects of biochar and its combination with compost on lettuce (Lactuca sativa L.) growth, soil properties, and soil microbial activity and abundance. Int. J. Agron. 2017, 12, 3158207. [Google Scholar] [CrossRef] [Green Version]
- Black, C.A. Methods of Soil Analysis: Part I Physical and Mineralogical Properties; American Society of Agronomy: Madison, WI, USA, 1965. [Google Scholar]
- Rayment, G.E.; Higginson, F.R. Australian Laboratory Handbook of Soil and Water Chemical Methods; Inkata Press Pty Ltd.: Melbourne, Australia, 1992. [Google Scholar]
- Jones, J.B., Jr.; Kalra, Y.P. Soil testing and plant analysis activities-The United States and Canada. Commun. Soil Sci. Plant Anal. 1992, 23, 2015–2027. [Google Scholar] [CrossRef]
- Mehlich, A. Use of triethanolamine acetate-barium hydroxide buffer for the determination of some base exchange properties and lime requirement of soil. Soil Sci. Soc. Am. Proc. 1938, 29, 374–378. [Google Scholar] [CrossRef]
- Dumas, J.B.A. Procédés de l’analyse organique. Ann. Chim. Phys. 1831, 247, 198–213. [Google Scholar]
- Bowman, R.A. A rapid method to determine total phosphorus in soils. Soil Sci. Soc. Am. J. 1988, 52, 1301–1304. [Google Scholar] [CrossRef]
- Olsen, S.R.; Cole, C.V.; Watanabe, F.S.; Dean, L.A. Estimation of Available Phosphorus in Soils by Extraction with Sodium Bicarbonate. USDA Circular 939; U.S. Government Publishing Office: Washington, DC, USA, 1954. [Google Scholar]
- Bouyoucos, G.J. Hydrometer method improved for making particle size analysis of soils. Agron. J. 1962, 54, 464–465. [Google Scholar] [CrossRef]
- Beretta, N.; Silbermann, A.V.; Paladino, L.; Torres, D.; Bassahun, D.; Musselli, R.; Lamohte, A.G. Soil texture analyses using a hydrometer modification of the Bouyoucos method. Cienc. Investig. Agrar. 2014, 41, 263–271. [Google Scholar] [CrossRef] [Green Version]
- Kemper, W.D.; Koch, E.J. Aggregate Stability of Soils from Western USA and Canada USDA Technical Bulletin No.1355; US Government Printing Office: Washington, DC, USA, 1966. [Google Scholar]
- Montagnoli, A.; Di Iorio, A.; Terzaghi, M.; Trupiano, D.; Scippa, G.S.; Chiatante, D. Influence of soil temperature and water content on fine-root seasonal growth of European beech natural forest in Southern Alps, Italy. Eur. J. For. Res. 2014, 133, 957–968. [Google Scholar] [CrossRef] [Green Version]
- Trupiano, D.; Rocco, M.; Renzone, G.; Scaloni, A.; Montagnoli, A.; Terzaghi, M.; Di Iorio, A.; Chiatante, D.; Scippa, G.S. Poplar woody root proteome during the transition dormancy-active growth. Plant Biosyst.-Int. J. Deal. All Asp. Plant Biol. 2013, 147, 1095–1100. [Google Scholar] [CrossRef] [Green Version]
- Doster, M.A.; Bostock, R.M. Quantification of lignin formation in almond bark in response to wounding and infection by Phytophthora species. Phytopathology 1988, 784, 73–477. [Google Scholar]
- Trupiano, D.; Di Iorio, A.; Montagnoli, A.; Lasserre, B.; Rocco, M.; Grosso, A.; Scaloni, A.; Marra, M.; Chiatante, D.; Scippa, G.S. Involvement of lignin and hormones in the response of woody poplar taproots to mechanical stress. Physiol. Plant. 2012, 146, 39–52. [Google Scholar] [CrossRef] [Green Version]
- Inskeep, W.P.; Bloom, P.R. Extinction coefficients of chlorophyll a and b in N,N Dimethylformamide and 80% acetone. Plant Physiol. 1985, 77, 483–485. [Google Scholar] [CrossRef] [Green Version]
- Kroetsch, D.; Wang, C. Particle size distribution. Soil Sampl. Methods Anal. 2008, 2, 713–725. [Google Scholar]
- Wolf, M.; Lehndorff, E.; Wiesenberg, G.L.B.; Stockhausen, M.; Schward, L.; Amelung, W. Towards reconstruction of past fire regimes from geochemical analysis of charcoal. Org. Geochem. 2013, 55, 11–21. [Google Scholar] [CrossRef] [Green Version]
- Ippolito, J.A.; Spokas, K.A.; Novak, J.M.; Lentz, R.D.; Cantrell, K.B. Biochar elemental composition and factors influencing nutrient retention. In Biochar for Environmental Management: Science, Technology and Implementation, 2nd ed.; Lehmann, J., Joseph, S., Eds.; Routledge: New York, NY, USA, 2015; pp. 137–161. [Google Scholar]
- Ippolito, J.A.; Cui, L.; Kammann, C.; Wrage-Mönnig, N.; Estavillo, J.M.; Fuertes-Mendizabal, T.; Cayuela, M.L.; Sigua, G.; Novak, J.; Spokas, K.; et al. Feedstock choice, pyrolysis temperature and type influence biochar characteristics: A comprehensive meta-data analysis review. Biochar 2020, 2, 421–438. [Google Scholar] [CrossRef]
- Ippolito, J.A.; Berry, C.M.; Strawn, D.G.; Novak, J.M.; Levine, J.; Harley, A. Biochars reduce mine land soil bioavailable metals. J. Environ. Qual. 2017, 46, 411–419. [Google Scholar] [CrossRef]
- Khan, N.; Clark, I.; Sánchez-Monedero, M.A.; Shea, S.; Meier, S.; Bolan, N. Maturity indices in co-composting of chicken manure and sawdust with biochar. Bioresour. Technol. 2014, 168, 245–251. [Google Scholar] [CrossRef]
- Zhang, J.; Zeng, G.; Chen, Y.; Yu, M.; Yu, Z.; Li, H.; Yu, Y.; Huang, H. Effects of physico-chemical parameters on the bacterial and fungal communities during agricultural waste composting. Bioresour. Technol. 2011, 102, 2950–2956. [Google Scholar] [CrossRef]
- Reyes-Torres, M.; Oviedo-Ocaña, E.R.; Dominguez, I.; Komilis, D.; Sánchez, A. A systematic review on the composting of green waste: Feedstock quality and optimization strategies. Waste Manag. 2018, 77, 486–499. [Google Scholar] [CrossRef]
- Järvis, J.; Ivask, M.; Nei, L.; Kuu, A.; Luud, A. Effect of green waste compost application on afforestation success. Balt. For. 2016, 22, 90–97. [Google Scholar]
- Heiskanen, J.; Hagner, M.; Ruhanen, H.; Mäkitalo, K. Addition of recyclable biochar, compost and fibre clay to the growth medium layer for the cover system of mine tailings: A bioassay in a greenhouse. Environ. Earth Sci. 2020, 79, 422. [Google Scholar] [CrossRef]
- Lazdina, D.; Bardule, A.; Lazdins, A.; Stola, J. Use of waste water sludge and wood ash as fertiliser for Salix cultivation in acid peat soils. Agron. Res. 2011, 9, 305–314. [Google Scholar]
- Gul, S.; Whalen, J.K. Biochemical cycling of nitrogen and phosphorus in biochar amended soils. Soil Biol. Biochem. 2016, 103, 1–15. [Google Scholar] [CrossRef]
- Seehausen, M.L.; Gale, N.V.; Dranga, S.; Hudson, V.; Liu, N.; Michener, J.; Thurston, E.; Williams, C.; Smith, S.M.; Thomas, S.C. Is there a positive synergistic effect of biochar and compost soil amendments on plant growth and physiological performance? Agronomy 2017, 7, 13. [Google Scholar] [CrossRef]
- Tessier, J.T.; Raynal, D.J. Use of nitrogen to phosphorus ratios in plant tissue as an indicator of nutrient limitation and nitrogen saturation. J. Appl. Ecol. 2003, 40, 523–534. [Google Scholar] [CrossRef]
- Tisdale, S.L.; Nelson, W.L.; Beaton, J.D. Soil Fertility and Fertilizers; Collier Macmillan Publishers: London, UK, 1985. [Google Scholar]
- Xu, G.; Zhang, Y.; Sun, J.; Shao, H. Negative interactive effects between biochar and phosphorus fertilization on phosphorus availability and plant yield in saline sodic soil. Sci. Total Environ. 2016, 568, 910–915. [Google Scholar] [CrossRef] [PubMed]
- Joseph, S.; Kammann, C.I.; Shepherd, J.G.; Conte, P.; Schmidt, H.P.; Hagemann, N.; Rich, A.M.; Marjo, C.E.; Allen, J.; Munroe, P.; et al. Microstructural and associated chemical changes during the composting of a high temperature biochar: Mechanisms for nitrate, phosphate and other nutrient retention and release. Sci. Total Environ. 2018, 618, 1210–1223. [Google Scholar] [CrossRef] [Green Version]
- Hossain, M.Z.; Bahar, M.M.; Sarkar, B.; Donne, S.W.; Ok, Y.S.; Palansooriya, K.N.; Kirkham, M.B.; Chowdhury, S.; Bolan, N. Biochar and its importance on nutrient dynamics in soil and plant. Biochar 2020, 2, 379–420. [Google Scholar] [CrossRef]
- Biederman, L.A.; Harpole, W.S. Biochar and its effects on plant productivity and nutrient cycling: A meta-analysis. GCB Bioenergy 2013, 5, 202–214. [Google Scholar] [CrossRef]
- Liu, Z.; He, T.; Cao, T.; Yang, T.; Meng, J.; Chen, W. Effects of biochar application on nitrogen leaching, ammonia volatilization and nitrogen use efficiency in two distinct soils. J. Soil Sci. Plant Nutr. 2017, 17, 515–528. [Google Scholar] [CrossRef] [Green Version]
- Spokas, K.A.; Novak, J.M.; Venterea, R.T. Biochar’s role as an alternative N-fertilizer: Ammonia capture. Plant Soil 2012, 350, 35–42. [Google Scholar] [CrossRef]
- Nguyen, T.T.N.; Xu, C.Y.; Tahmasbian, I.; Che, R.; Xu, Z.; Zhou, X.; Wallace, H.M.; Bai, S.H. Effects of biochar on soil available inorganic nitrogen: A review and metaanalysis. Geoderma 2017, 288, 79–96. [Google Scholar] [CrossRef] [Green Version]
- Sadowska, U.; Domagała-Świątkiewicz, I.; Żabiński, A. Biochar and its effects on plant–soil macronutrient cycling during a three-year field trial on sandy soil with peppermint (Mentha piperita L.). Part I: Yield and macro element content in soil and plant biomass. Agronomy 2020, 10, 1950. [Google Scholar] [CrossRef]
- Bista, P.; Ghimire, R.; Machado, S.; Pritchett, L. Biochar effects on soil properties and wheat biomass vary with fertility management. Agronomy 2019, 9, 623. [Google Scholar] [CrossRef] [Green Version]
- Marks, A.N.; Mattan, S.; Alcañiz, J.M.; Pérez-Herrero, E.; Domene, X. Gasifier biochar effects on nutrient availability, organic matter mineralization, and soil fauna activity in a multi-year Mediterranean trial. Agric. Ecosyst. Environ. 2016, 215, 30–39. [Google Scholar] [CrossRef]
- Wang, J.; Xiong, Z.; Kuzyakov, Y. Biochar stability in soil: Metaanalysis of decomposition and priming effects. Glob. Chang. Biol. Bioenergy 2016, 8, 512–523. [Google Scholar] [CrossRef] [Green Version]
- Ventura, M.; Alberti, G.; Panzacchi, P.; Vedove, G.D.; Miglietta, F.; Tonon, G. Biochar mineralization and priming effect in a poplar short rotation coppice from a 3-year field experiment. Biol. Fertil. Soils 2019, 55, 67–78. [Google Scholar] [CrossRef]
- Mertens, J.; Germer, S.; Germer, J.; Sauerborn, J. Comparison of soil amendments for reforestation with a native multipurpose tree under semiarid climate: Root and root tuber response of Spondias tuberosa. For. Ecol. Manag. 2017, 396, 1–10. [Google Scholar] [CrossRef]
- Mašková, T.; Herben, T. Root: Shoot ratio in developing seedlings: How seedlings change their allocation in response to seed mass and ambient nutrient supply. Ecol. Evol. 2018, 8, 7143–7150. [Google Scholar] [CrossRef] [Green Version]
- Song, X.; Wan, F.; Chang, X.; Zhang, J.; Sun, M.; Liu, Y. Effects of nutrient deficiency on root morphology and nutrient allocation in Pistacia chinensis Bunge seedlings. Forests 2019, 10, 1035. [Google Scholar] [CrossRef] [Green Version]
- Jordan-Meille, L.; Martineau, E.; Bornot, Y.; Lavres, J.; Abreu-Junior, C.H.; Domec, J.C. How does water-stressed corn respond to potassium nutrition? A shoot-root scale approach study under controlled conditions. Agriculture 2018, 8, 180. [Google Scholar] [CrossRef] [Green Version]
- West, J.B.; Espeleta, J.F.; Donovan, L.A. Fine root production and turnover across a complex edaphic gradient of a Pinus palustris–Aristida stricta savanna ecosystem. For. Ecol. Manag. 2004, 189, 397–406. [Google Scholar] [CrossRef]
- Cheng, Y.; Han, Y.; Wang, Q.; Wang, Z. Seasonal dynamics of fine root biomass, root length density, specific root length, and soil resource availability in a Larix gmelinii plantation. Front. Biol. China 2006, 1, 310–317. [Google Scholar] [CrossRef]
- Bjork, R.G.; Majdi, M.; Klemedtsson, L.; Jonsson, L.L.; Molau, U. Long-term warming effects on root morphology, root mass distribution, and microbial activity in two dry tundra plant communities in northern Sweden. New Phytol. 2007, 176, 862–873. [Google Scholar] [CrossRef] [PubMed]
- Makita, N.; Hirano, Y.; Mizoguchi, T.; Kominami, Y.; Dannoura, M.; Ishii, H.; Finer, L.; Kanazawa, Y. Very fine roots respond to soil depth: Biomass allocation, morphology, and physiology in a broad-leaved temperate forest. Ecol. Res. 2011, 26, 95–104. [Google Scholar] [CrossRef]
- Amendola, C.; Montagnoli, A.; Terzaghi, M.; Trupiano, D.; Oliva, F.; Baronti, S.; Miglietta, F.; Chiatante, D.; Scippa, G.S. Short-term effects of biochar on grapevine fine root dynamics and arbuscular mycorrhizae production. Agric. Ecosyst. Environ. 2017, 239, 236–245. [Google Scholar] [CrossRef]
- Hilty, J.; Muller, B.; Pantin, F.; Leuzinger, S. Tansley review: Plant growth: The what, the how, and the why. New Phytol. 2021, 232, 25–41. [Google Scholar] [CrossRef]
- Lind, E.M.; Borer, E.; Seabloom, E.; Adler, P.; Bakker, J.D.; Blumenthal, D.M.; Crawley, M.; Davies, K.; Firn, J.; Gruner, D.S.; et al. Life-history constraints in grassland plant species: A growth-defence trade-off is the norm. Ecol. Lett. 2013, 16, 513–521. [Google Scholar] [CrossRef] [Green Version]
- Agegnehu, G.; Bass, A.M.; Nelson, P.N.; Bird, M.I. Benefits of biochar, compost and biochar–compost for soil quality, maize yield and greenhouse gas emissions in a tropical agricultural soil. Sci. Total Environ. 2016, 543, 295–306. [Google Scholar] [CrossRef]
- Zhang, D.; Pan, G.; Wu, G.; Kibue, G.W.; Li, L.; Zhang, X.; Zheng, J.; Zheng, J.; Cheng, K.; Joseph, S. Biochar helps enhance maize productivity and reduce greenhouse gas emissions under balanced fertilization in a rainfed low fertility inceptisol. Chemosphere 2016, 142, 106–113. [Google Scholar] [CrossRef]
- Viger, M.; Hancock, R.D.; Miglietta, F.; Taylor, G. More plant growth but less plant defence? First global gene expression data for plants grown in soil amended with biochar. Glob. Chang. Biol. Bioenergy 2015, 7, 658–672. [Google Scholar] [CrossRef]
- Polzella, A.; De Zio, E.; Arena, S.; Scippa, G.S.; Scaloni, A.; Montagnoli, A.; Chiatante, D.; Trupiano, D. Toward an understanding of mechanisms regulating plant response to biochar application. Plant Biosyst. 2019, 153, 163–172. [Google Scholar] [CrossRef]
- Simiele, M.; Sferra, G.; Lebrun, M.; Renzone, G.; Bourgerie, S.; Scippa, G.S.; Morabito, D.; Scaloni, A.; Trupiano, D. In-depth study to decipher mechanisms underlying Arabidopsis thaliana tolerance to metal (loid) soil contamination in association with biochar and/or bacteria. Environ. Exp. Bot. 2021, 182, 104335. [Google Scholar] [CrossRef]
- Amthor, J.S. Efficiency of lignin biosynthesis: A quantitative analysis. Ann. Bot. 2003, 91, 673–695. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rogers, L.A.; Campbell, M.M. The genetic control of lignin deposition during plant growth and development. New Phytol. 2004, 164, 17–30. [Google Scholar] [CrossRef] [PubMed]
- Ithal, N.; Recknor, J.; Nettleton, D.; Maier, T.; Baum, T.J.; Mitchum, M.G. Developmental transcript profiling of cyst nematode feeding cells in soybean roots. Mol. Plant Microbe Interact. 2007, 20, 510–525. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vanholme, R.; Storme, V.; Vanholme, B.; Sundin, L.; Christensen, J.H.; Goeminne, G.; Halpin, C.; Rohde, A.; Morreel, K.; Boerjan, W. A systems biology view of responses to lignin biosynthesis perturbations in Arabidopsis. Plant Cell 2012, 24, 3506–3529. [Google Scholar] [CrossRef] [Green Version]
- Jaiswal, A.K.; Alkan, N.; Elad, Y.; Sela, N.; Philosoph, A.M.; Graber, E.R.; Frenkel, O. Molecular insights into biochar-mediated plant growth promotion and systemic resistance in tomato against Fusarium crown and root rot disease. Sci. Rep. 2020, 10, 13934. [Google Scholar] [CrossRef]
- Liu, Q.; Luo, L.; Zheng, L. Lignins: Biosynthesis and biological functions in plants. Int. J. Mol. Sci. 2018, 19, 335. [Google Scholar] [CrossRef] [Green Version]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Ctrl | B | Co | BCo | |
|---|---|---|---|---|
| Moisture content (g·kg−1) | 1.52 ± 0.06 a | 1.43 ± 0.05 ab | 1.15 ± 0.01 b | 1.57 ± 0.14 a |
| pH | 7.4 ± 0.0 a | 7.4 ± 0.0 a | 7.4 ± 0.0 a | 7.4 ± 0.0 a |
| Alkalinity (% CaCO3) | 10.00 ± 0.29 a | 10.57 ± 0.49 a | 9.53 ± 0.33 a | 10.83 ± 0.24 a |
| EC (dS·m−1) | 0.233 ± 0.005 c | 0.247 ± 0.006 bc | 0.447 ± 0.002 a | 0.340 ± 0.025 b |
| CEC (cmol(+)·kg−1) | 6.49 ± 0.36 b | 8.62 ± 0.27 a | 9.69 ± 0.18 a | 9.60 ± 0.23 a |
| TOC (g·kg−1) | 12.26 ± 0.94 b | 13.38 ± 1.27 b | 23.41 ± 1.28 a | 23.54 ± 0.69 a |
| Ntot (g·kg−1) | 0.23 ± 0.01 d | 0.52 ± 0.01 c | 1.60 ± 0.05 b | 2.06 ± 0.01 a |
| Ptot (mg·kg−1) | 129.33 ± 4.52 c | 215.00 ± 3.24 b | 301.00 ± 2.94 a | 324.00 ± 5.93 a |
| Pav (mg·kg−1) | <12 b | <12 b | 16.90 ± 1.49 a | 18.63 ± 1.01 a |
| Particle size distribution *: | ||||
| ø < 2 µm (%) (clay) | 8.90 ± 0.36 b | 6.27 ± 0.18 c | 13.50 ± 0.18 a | 9.63 ± 0.96 b |
| 2 < ø < 20 µm (%) (silt) | 53.80 ± 1.61 a | 52.70 ± 1.19 a | 53.17 ± 1.72 a | 54.63 ± 1.70 a |
| 20 < ø < 50 µm (%) (very fine sand) | 2.63 ± 0.91 a | 1.93 ± 0.05 a | 1.90 ± 0.16 a | 1.60 ± 0.29 a |
| 50 < ø < 200 µm (%) (fine sand) | 8.93 ± 0.55 ab | 9.50 ± 0.35 a | 7.90 ± 1.38 ab | 6.73 ± 0.33 b |
| 200 µm < ø < 2 mm (%) (coarse sand) | 6.83 ± 0.49 a | 7.20 ± 0.41 a | 8.67 ± 0.23 a | 9.10 ± 0.79 a |
| ø > 2 mm (%) (gravel) | 27.80 ± 0.89 a | 28.67 ± 0.55 a | 28.33 ± 0.58 a | 28.00 ± 0.36 a |
| Ctrl * | B | Co | BCo | |
|---|---|---|---|---|
| Chl | 7.91 ± 0.13 b | 11.51 ± 1.99 a | 5.47 ± 0.40 b | 11.58 ± 1.70 a |
| Chl a | 6.17 ± 0.40 b | 9.02 ± 0.94 a | 4.37 ± 0.37 b | 9.04 ± 1.15 a |
| Chl b | 1.74 ± 0.27 b | 2.48 ± 0.34 a | 1.10 ± 0.03 b | 2.54 ± 0.55 a |
| Chl a/Chl b | 3.61 ± 0.79 a | 3.63 ± 1.99 a | 3.93 ± 0.22 a | 3.60 ± 0.33 a |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Simiele, M.; De Zio, E.; Montagnoli, A.; Terzaghi, M.; Chiatante, D.; Scippa, G.S.; Trupiano, D. Biochar and/or Compost to Enhance Nursery-Produced Seedling Performance: A Potential Tool for Forest Restoration Programs. Forests 2022, 13, 550. https://0-doi-org.brum.beds.ac.uk/10.3390/f13040550
Simiele M, De Zio E, Montagnoli A, Terzaghi M, Chiatante D, Scippa GS, Trupiano D. Biochar and/or Compost to Enhance Nursery-Produced Seedling Performance: A Potential Tool for Forest Restoration Programs. Forests. 2022; 13(4):550. https://0-doi-org.brum.beds.ac.uk/10.3390/f13040550
Chicago/Turabian StyleSimiele, Melissa, Elena De Zio, Antonio Montagnoli, Mattia Terzaghi, Donato Chiatante, Gabriella Stefania Scippa, and Dalila Trupiano. 2022. "Biochar and/or Compost to Enhance Nursery-Produced Seedling Performance: A Potential Tool for Forest Restoration Programs" Forests 13, no. 4: 550. https://0-doi-org.brum.beds.ac.uk/10.3390/f13040550