1. Introduction
Forests that develop after coppicing from the stumps of felled trees and from buried seeds after both artificial and natural disturbances are defined as secondary forest [
1]. In Japan, forests constitute almost 70% of the land area, and around 60% of them are secondary [
2]. In general, forests can fix CO
2 even at their climax stage, as extremely old-growth forests, though the amount of CO
2 fixation is thought to be less in older forests than in younger forests [
3]. Since large parts of the secondary forests have not reached such climax stage as compared to primary forests, they may contribute substantially to the global uptake of CO
2 [
4].
The gross uptake of CO
2 through photosynthesis is defined as the gross primary production (GPP), which is the largest CO
2 flux between the atmosphere and terrestrial ecosystems [
5]. In addition to the absolute size of GPP, it is important to accurately estimate it in secondary forests, as GPP is a source of energy for all organisms. Elucidation of the factors that control it is also important t for the prediction of GPP under climate change. GPP is often estimated by meteorological techniques, such as the Eddy Covariance (EC) method, in forest ecosystems [
6]. The EC method can not only estimate the GPP of an entire forest, but also identify the factors that control it [
7,
8]. However, a forest structure is highly complex, both vertically and horizontally, and is characterized by several distinct layers with very different functional properties [
9]. Therefore, to better understand the GPP of forest ecosystems, it is important to evaluate the GPP of each layer, especially that of the understory vegetation, which is reported to possibly contribute to a large extent to the overall GPP of a forest [
10,
11,
12]. However, it is difficult to estimate the GPP of each layer and to identify the factors that control it by the EC method [
9]. In the forest understory layer, in particular, it is usually not practical to use the EC method alone because low wind speeds and intermittent turbulence can reduce data reliability [
9,
13,
14].
Dwarf bamboo species of the genus
Sasa are dominant in the understory of many secondary forests in Japan, especially in snow-covered areas [
12,
15]. The exclusive dominance of
Sasa decreases the amount of light that reaches the ground, leading to lower survival rates and densities of seedlings and saplings [
16,
17]. Therefore, the abundant growth of
Sasa can prevent forest regeneration in Japan [
18]. Moreover, it also has a negative effect on forest function through the reduction of understory biodiversity [
19]. However, recent studies demonstrated that
Sasa, which was previously thought to only hinder forest regeneration and functions, contributed 16% to 25% of the forest GPP in an old-growth beech forest [
12]. In addition, Cai et al. (2021) demonstrated that the seasonal dynamics of
Sasa GPP and its controlling factors, as well as the annual
Sasa GPP, differed between sites covered by tree canopy and gaps created by large disturbances in the same forest [
12]. Thus,
Sasa GPP and its controlling factors could be influenced by the structure of the forest canopy, both temporally and spatially, which would create different environmental conditions on the forest floor. Since
Sasa is widely distributed in forests in Japan and could contribute significantly to forest ecosystem GPP, it is of importance to accurately estimate its GPP and its seasonal trends under different canopy structure. In addition, understanding the factors that control the seasonal changes of
Sasa GPP in these forests is essential for predicting how
Sasa GPP will change under global warming.
This study was conducted in three types of secondary forests: deciduous broadleaved, evergreen coniferous, and mixed forest consisting of both coniferous and deciduous trees. These three types of forest were chosen because they are the typical forest with
Sasa-dominated understory. By combining data of the vegetation distribution (data of vegetation survey conducted by the Ministry of the Environment in Japan) and the data of the distribution of
Sasa senanensis [
20], the targeted dwarf bamboo species in this study, it appeared that deciduous broadleaved forest, mixed forest, and evergreen coniferous forest occupy 43.8%, 2.2%, and 24.4%, respectively. Even though the ratio of mixed forest was relatively low, it was higher than that of other types of forest, i.e., deciduous coniferous forest and evergreen broadleaved forest. These three forests have different canopy structures both temporally and spatially, which could largely influence the understory environment. In the deciduous broadleaved forest dominated by mature beech trees, there were two areas where the forest floor environment was significantly different, so we established two study areas: a closed-canopy area and a canopy gap area (canopy area and gap area, [
12]). In this study, we tested two hypotheses: (1) the seasonal trends of
Sasa community GPP and
Sasa annual GPP would differ among forest types, (2) in addition to light intensity, the seasonal changes of
Sasa community GPP would be controlled by different factors in different forest types.
2. Materials and Methods
2.1. Study Area
The three secondary forests were an evergreen coniferous forest (about 60 years old), a mixed forest with coniferous and deciduous trees (about 90 years old), both at the Sugadaira Montane Research Center (SMRC), University of Tsukuba, Nagano (36°31′ N, 138°21′ E, 1315 m elevation; [
21]), and a deciduous broadleaved forest (more than 300 years old), in Kayanodaira, Nagano (36°50′ N, 138°30′ E, 1495 m elevation; [
22];
Figure 1). Observations were carried out in a 1 ha permanent monitoring site in each forest. The two 1 ha sites (200 m × 50 m) at SMRC (evergreen coniferous forest and mixed forest) were established in 2007, and the 1 ha site (100 m × 100 m) at Kayanodaira in 2005 (this is one of the Japanese Ministry of the Environment’s “Monitoring Sites 1000” program sites; [
23]). Each plot was divided into 10 m × 10 m sub-plots. At each site, in addition to the annual survey of diameter at breast height (DBH) and survival status of all target trees with DBH ≥ 5 cm, litter fall was recorded monthly. The dominant species were
Fagus crenata in the deciduous broadleaved forest,
Pinus densiflora in the evergreen coniferous forest, and
Quercus crispula and
P. densiflora in the mixed forest. The stand density was 2400 ha
−1, 984 ha
−1, and 1210 ha
−1 in the evergreen coniferous forest, mixed forest, and deciduous forest, respectively, and there was no forest management for at least 70 years. The forest understory at all sites was dominated by
Sasa senanensis [
21,
22], with a mean height of around 2 m. The mean annual temperature and average precipitation from 1981 to 2010 were 4.9 °C and 1678 mm year
−1 at Kayanodaira [
22] and 6.6 °C and 1343 mm year
−1 at SMRC (
https://www.sugadaira.tsukuba.ac.jp/nature/weather.html; accessed date: 28 February 2022). The maximum snow depth was 4 m at Kayanodaira and 1 m at SMRC. Hereinafter, the sites in this paper indicate canopy area and gap area in the deciduous broadleaved forest, evergreen coniferous forest, and mixed forest.
2.2. Measurement of Metrological Variables
Photosynthetic photon flux density (PPFD), air temperature (TA), relative humidity (RH) at 2 m above the ground, soil temperature (TS) at 5 cm depth, and soil water content (SWC) at 10 cm depth were measured from 2 April to 29 October in the evergreen coniferous forest and mixed forest and from 7 May to 25 October in the deciduous broadleaved forest. PPFD was measured at 5 min intervals, and other factors at 30 min intervals. All measurements in the deciduous broadleaved forest were conducted in both canopy and gap areas. PPFD was measured with a quantum sensor (SE-SQ110, Apogee Electronics, Santa Monica, CA, USA), TA and RH with a temperature sensor and an RH datalogger (HOBO MX2300, Onset Computer, Bourne, MA, USA), TS with temperature dataloggers (HOBO MX2204, Onset Computer), and SWC with a soil moisture sensor (SM150, Delta-T, Cambridge, UK). A solar radiation shield (RS3-B, Onset Computer) shaded the TA and RH sensor from direct sunlight. The quantum sensor in the deciduous broadleaved forest canopy area broke immediately after installation, so we used data acquired from a sensor built into a leaf area index analyzer (MIJ-15LAI/P, Environmental Measurement Japan, Co., Ltd., Fukuoka, Japan) established 2 m above the ground from 7 May.
2.3. CO2 Flux Measurement and Calculation
The CO
2 flux of the
S. senanensis community was measured by using a static chamber system (
Figure 2) at four measurement points at each site. The chamber system is described in detail in Cai et al. (2021). In brief, it consists of a lid, a body, and a bottom frame (
Figure 2). We used a new small multiple sensors for CO
2, temperature, and RH with a datalogger (HOBO MX1102A, Onset Computer), whose measurement accuracy and response time were verified beforehand. The bottom frame was set into the soil right after the snow melted at each site (2 April in the evergreen coniferous forest and mixed forest, 7 May in the canopy area and gap area in the deciduous broadleaved forest), and the lid and body were placed only during CO
2 flux measurement.
CO
2 uptake from the atmosphere within the chamber was defined as positive, and emission as negative. To estimate the GPP at the community scale and to obtain GPP-light response curves at each measurement point, right after we measured the CO
2 flux under 100% light (CO
2Flight), we measured it under 0% light (CO
2Fdark) by placing a black plastic sheet over the chamber and measuring PPFD outside the chamber. All data were stored at 2 s intervals, and each measurement took 3 to 5 min to hold the increase of TA within 2 °C during the measurements. To maintain data quality, we excluded measurements in which T
A increased by <2 °C or decreased or RH decreased. We calculated CO
2Flight and CO
2Fdark (μmol CO
2 m
−2 s
−1) as [
12]:
where ΔCO
2/Δt is the slope of the chamber CO
2 concentration against time (μmol CO
2 mol
−1 s
−1), P is the atmospheric pressure held constant (kPa), R is the gas constant (8.314 kPa m
−2 K
−1 mol
−1), T is the average temperature during the measurement (°C), V is the chamber volume (m
3), and A is the surface area under the chamber (m
2). The slope (S = ΔCO
2/Δt) was obtained by fitting a linear model (C = St + b) to the sampling data, where C is the momentary CO
2 concentration (μmol mol
−1) at time t, and b is the intercept.
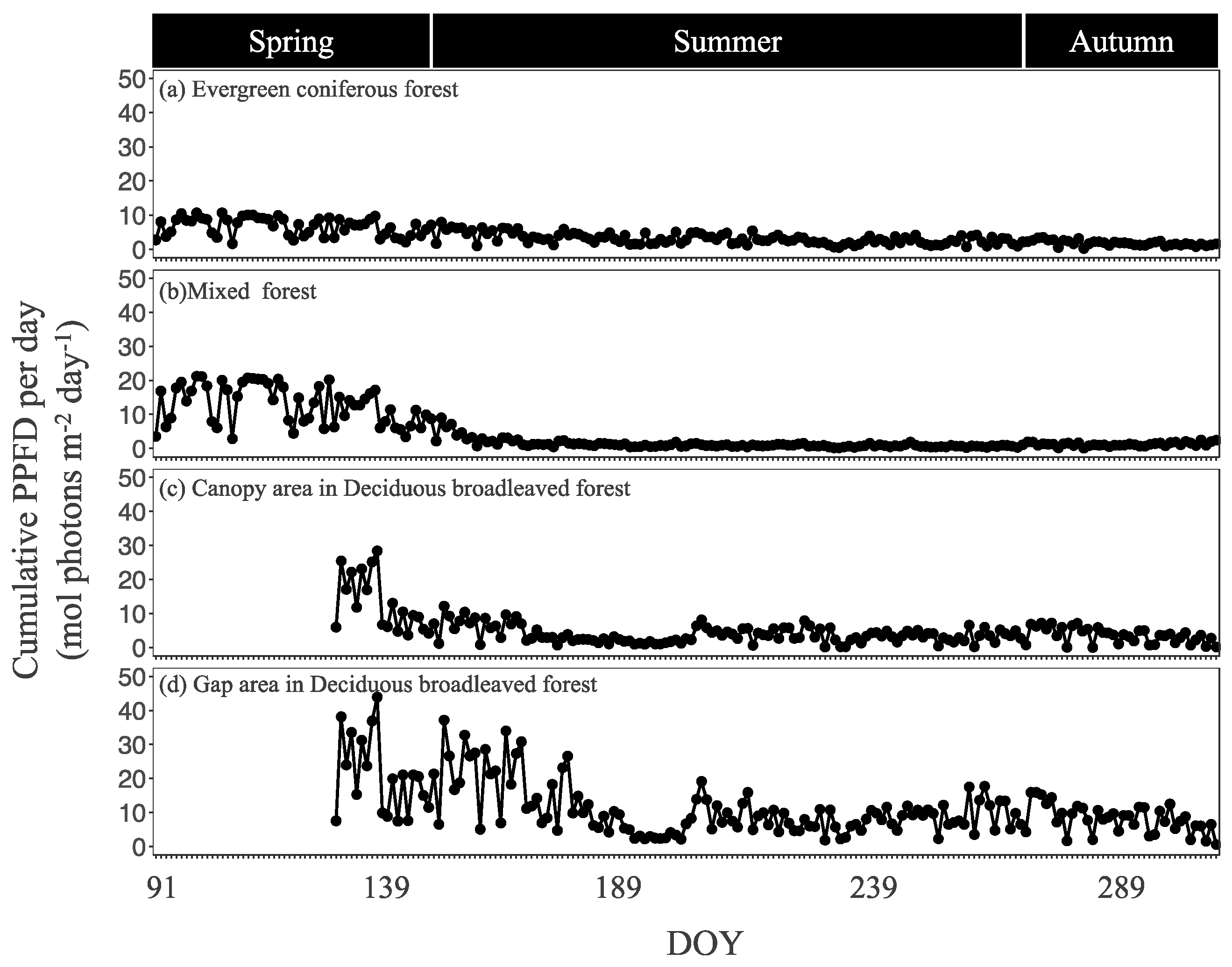
Measurements were taken from 28 April to 2 May (spring), 31 August to 3 September (summer), and 26 to 29 October (autumn) in 2021 in the evergreen coniferous forest and mixed forest, and from 22 to 25 May, 28 to 31 July, and 22 to 25 October in the canopy area and gap area in the deciduous broadleaved forest.
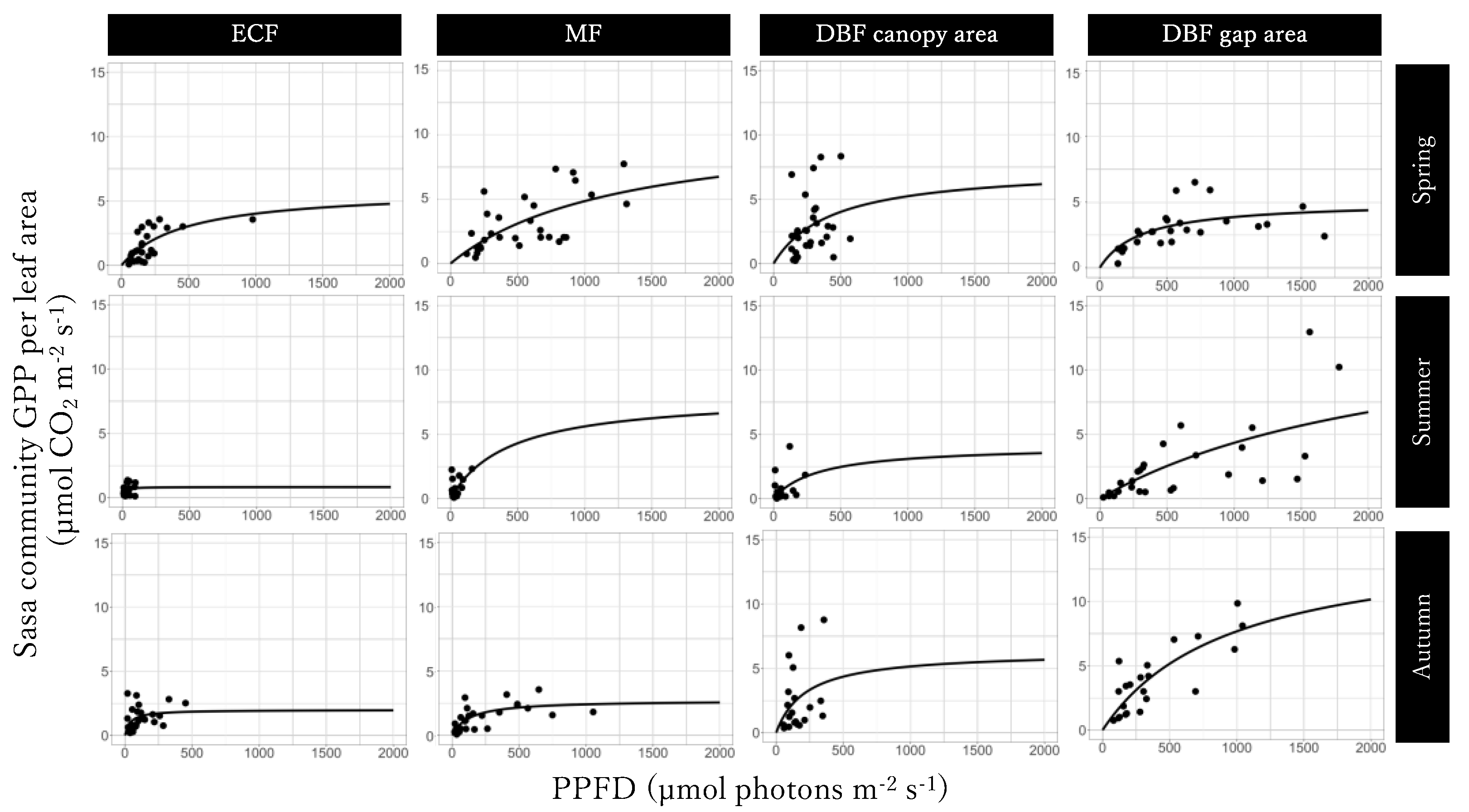
2.4. Sasa Senanensis GPP at the Community Scale and its Relationship with PPFD
The GPP of
S. senanensis at the community scale (μmol CO
2 m
−2 s
−1) was estimated as [
12]:
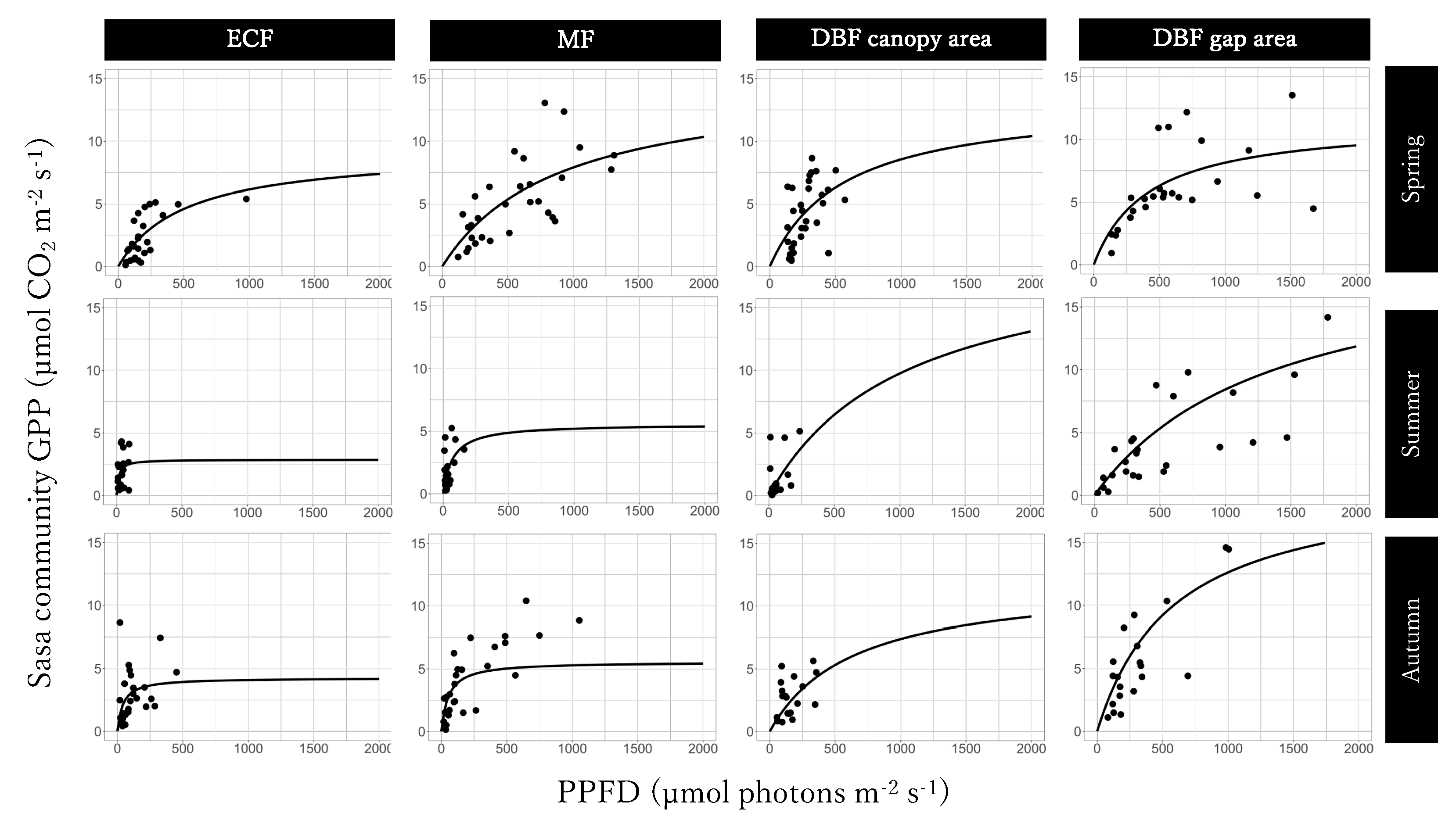
To depict the relationship with PPFD, we drew GPP–light response curves as a rectangular hyperbola [
12,
24]:
where α is the initial slope of the rectangular hyperbola (also called the apparent quantum yield of GPP), and GPP
max is the maximum GPP under light saturation. These parameters were calculated separately at each site in each season.
Furthermore, we calculated an index of photosynthetic ability of
Sasa by dividing the obtained GPP
Sasa-community by the
Sasa community leaf area (see
Section 2.5 as follows).
2.5. Biomass and Leaf Area of S. senanensis
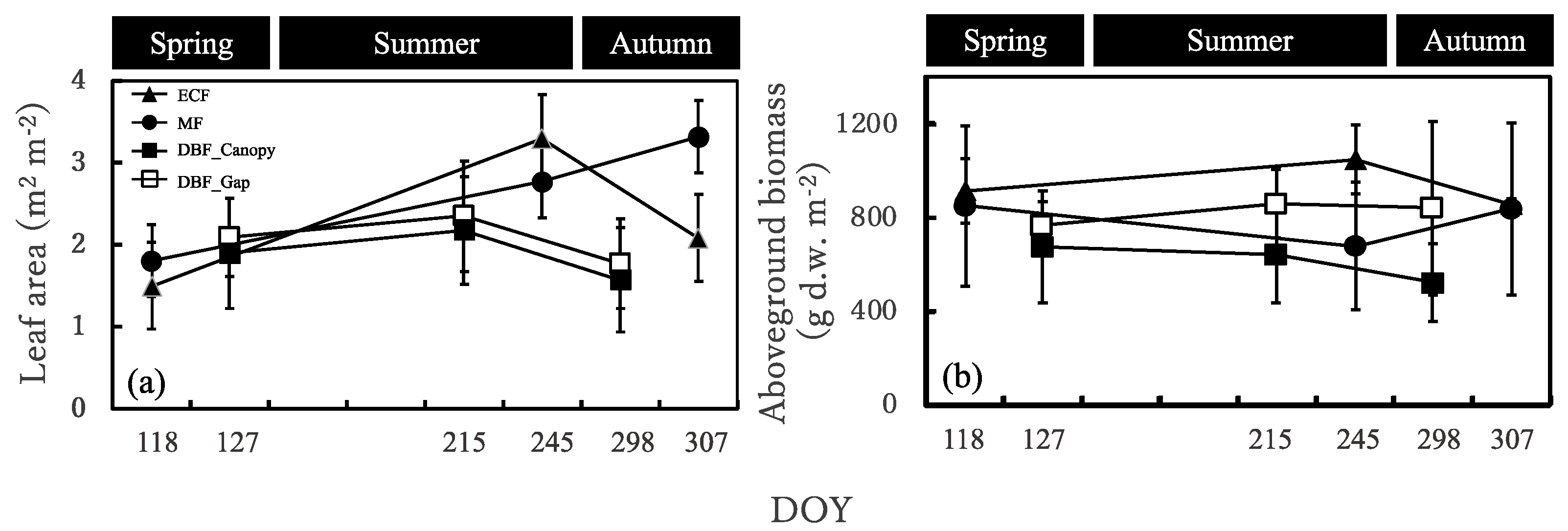
Allometric equations relating the length of each culm to its dry weight, the number of leaves per shoot to their biomass, and the number of leaves per shoot to their leaf area were estimated at each site in each season. For this, at least 15 shoots were harvested each time. After we measured the shoot length, leaf number, and leaf area per shoot, we oven-dried the samples for 48 h at 75 °C and then weighed culms and leaves separately. The leaf areas were measured using a potable leaf area meter (LI-3000CAP, LI-COR, Lincoln, NE, USA). The allometric equations were all significant at the 0.01 level (data not shown). They were then used to non-destructively estimate the biomass and leaf area of the S. senanensis community within each chamber.
2.6. Sasa GPP at the Forest Ecosystem Scale and Its Contribution to Forest GPP
To scale the
Sasa GPP from the community scale up to the forest ecosystem scale, we assumed that
Sasa was distributed evenly in the evergreen coniferous forest and mixed forest after visually checking its distribution. In the deciduous broadleaved forest, we took the area proportions of the canopy area (64.2%) and gap area (35.8%) into consideration [
12].
To estimate the contribution of
Sasa GPP to the forest ecosystem GPP, we estimated the tree GPP in each forest using the same procedure as in our previous study [
12]. First, we used the relationship between the net primary production (NPP) of trees and the GPP of trees [
25,
26]. Since we had access to an annual survey of DBH and litterfall in each forest, it was possible to calculate NPP as the sum of the increase of tree biomass and litterfall [
27]. The biomass of trees, both above and below ground, was calculated with an allometric formula [
28]. Second, we used the relationship between annual mean T
A and GPP of trees [
29,
30].
2.7. Statistical Analysis
To elucidate the factors that control the seasonal dynamics of the
Sasa community GPP, we conducted multiple linear regressions with daily cumulative
Sasa community GPP as the response variable and PPFD, T
A, RH, T
S, SWC, and leaf area of the
Sasa community GPP as the explanatory variables. All data were standardized, and variance inflation factors (VIFs) were calculated to detect collinearity among explanatory variables. We thereby excluded T
S and SWC from the analysis of the evergreen coniferous forest, T
A and T
S from the analysis of mixed forest, canopy area from the analysis of the deciduous broadleaved forest, and T
S and leaf area from the analysis of the deciduous broadleaved forest gap area (VIFs of other parameters <10). All analyses were performed in R (v. 4.0.3 software [
31]).
5. Conclusions
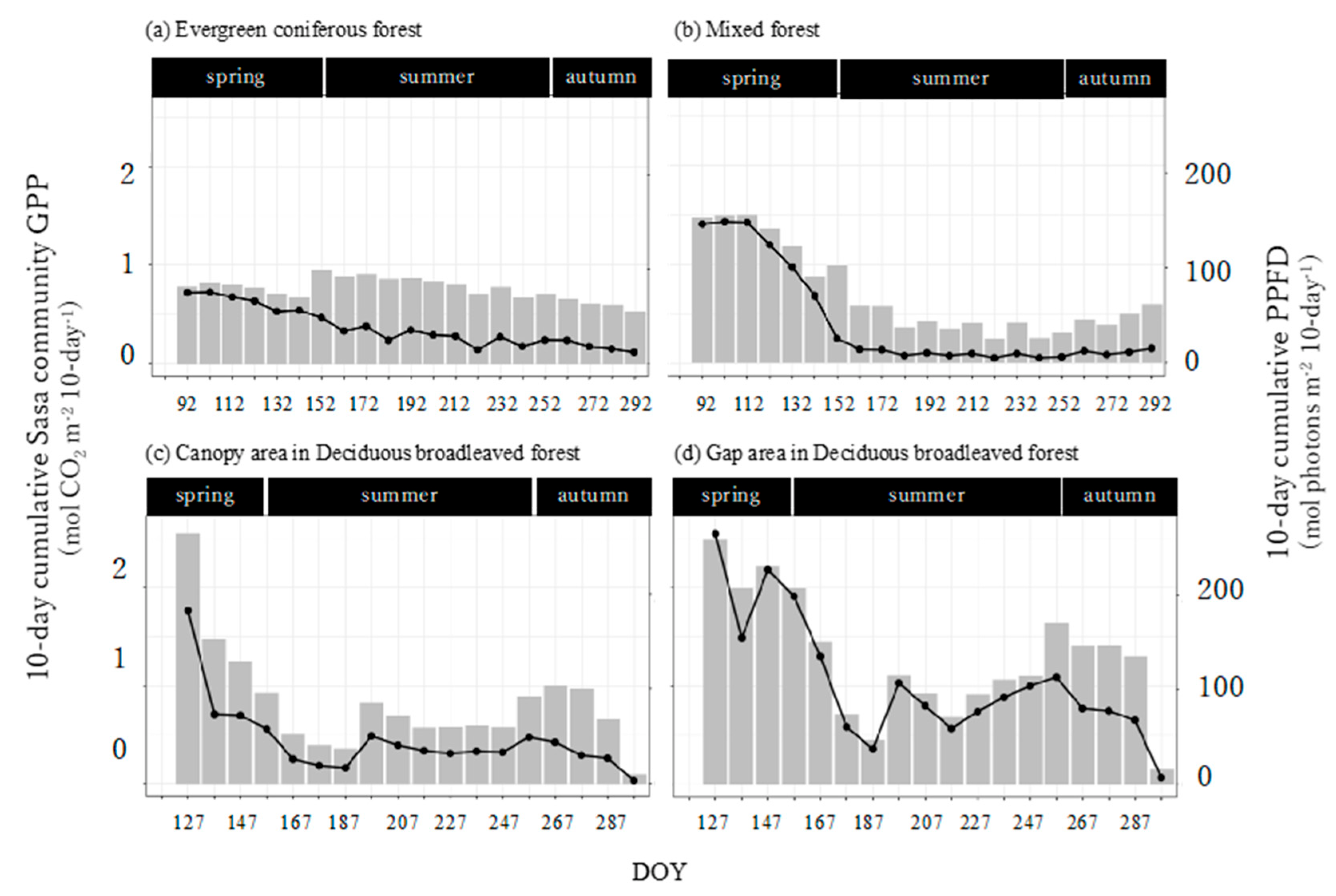
The seasonal trends of the Sasa community GPP and its controlling factors were different in the evergreen coniferous forest, mixed forest, and canopy and gap areas of the deciduous broadleaved forest. Further research is necessary, also taking into consideration stand structure and forest management, in addition to canopy structure.
The annual Sasa GPP was almost similar for evergreen coniferous forest, mixed forest, and deciduous broadleaved forest, which was different from what we expected. It also turned out that the annual Sasa GPP would be influenced not only by the understory environment or the photosynthesis ability of the Sasa community, but also by the length of the growing periods. Since Sasa dominates the understory of many secondary forests in Japan and could contribute greatly to the GPP, further research on annual changes in Sasa GPP is necessary.


 and
and 







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}