
Phenotypic Diversity Analysis and Superior Family Selection of Industrial Raw Material Forest Species-Pinus yunnanensis Franch

Abstract
:1. Introduction
2. Materials and Methods
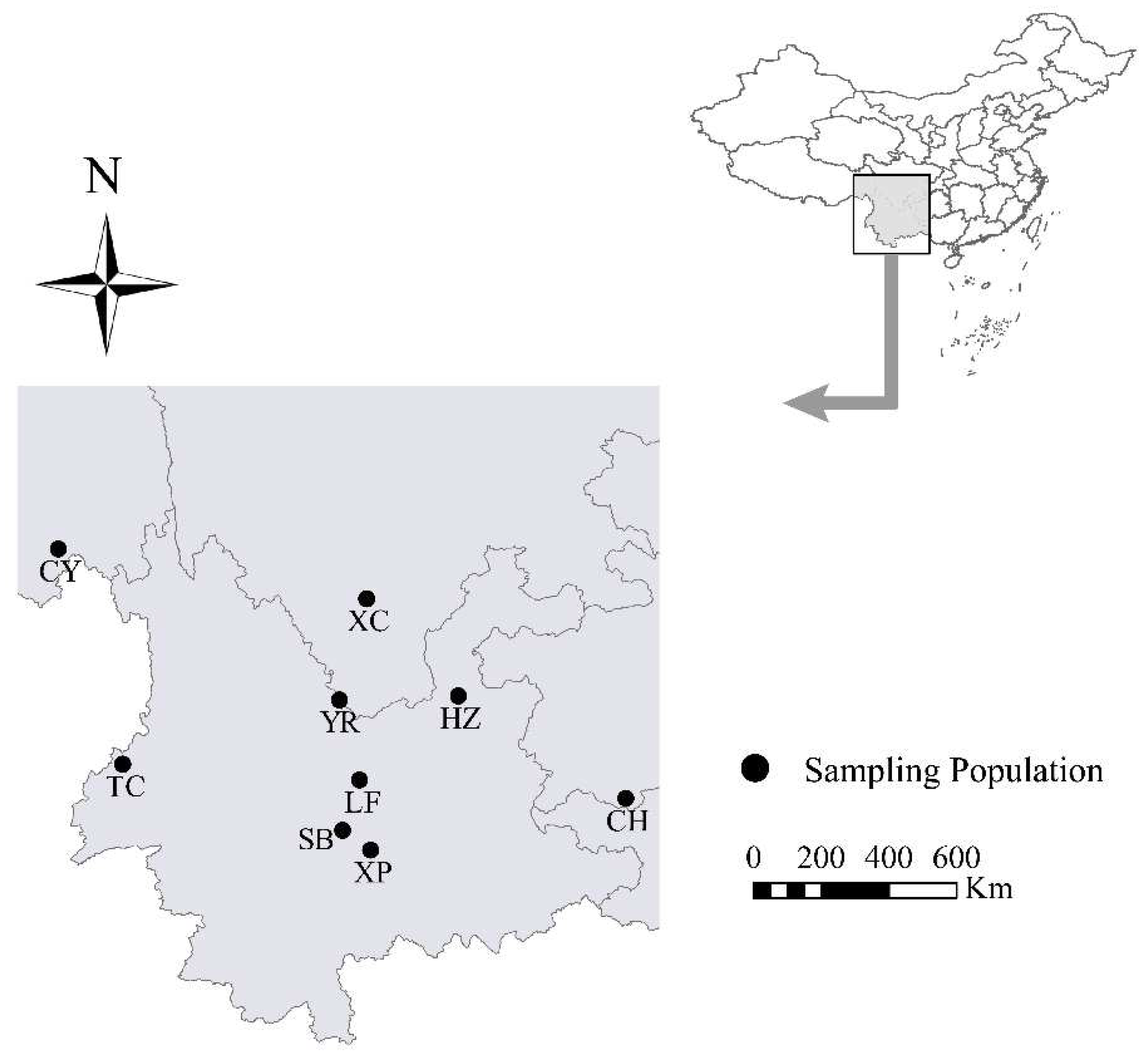
2.1. Site Description and Stand Characteristics
2.2. Measurement of Phenotypic Traits
2.3. Standardized Data
2.4. Data Analysis
2.5. Superior Family Selection Method
3. Results
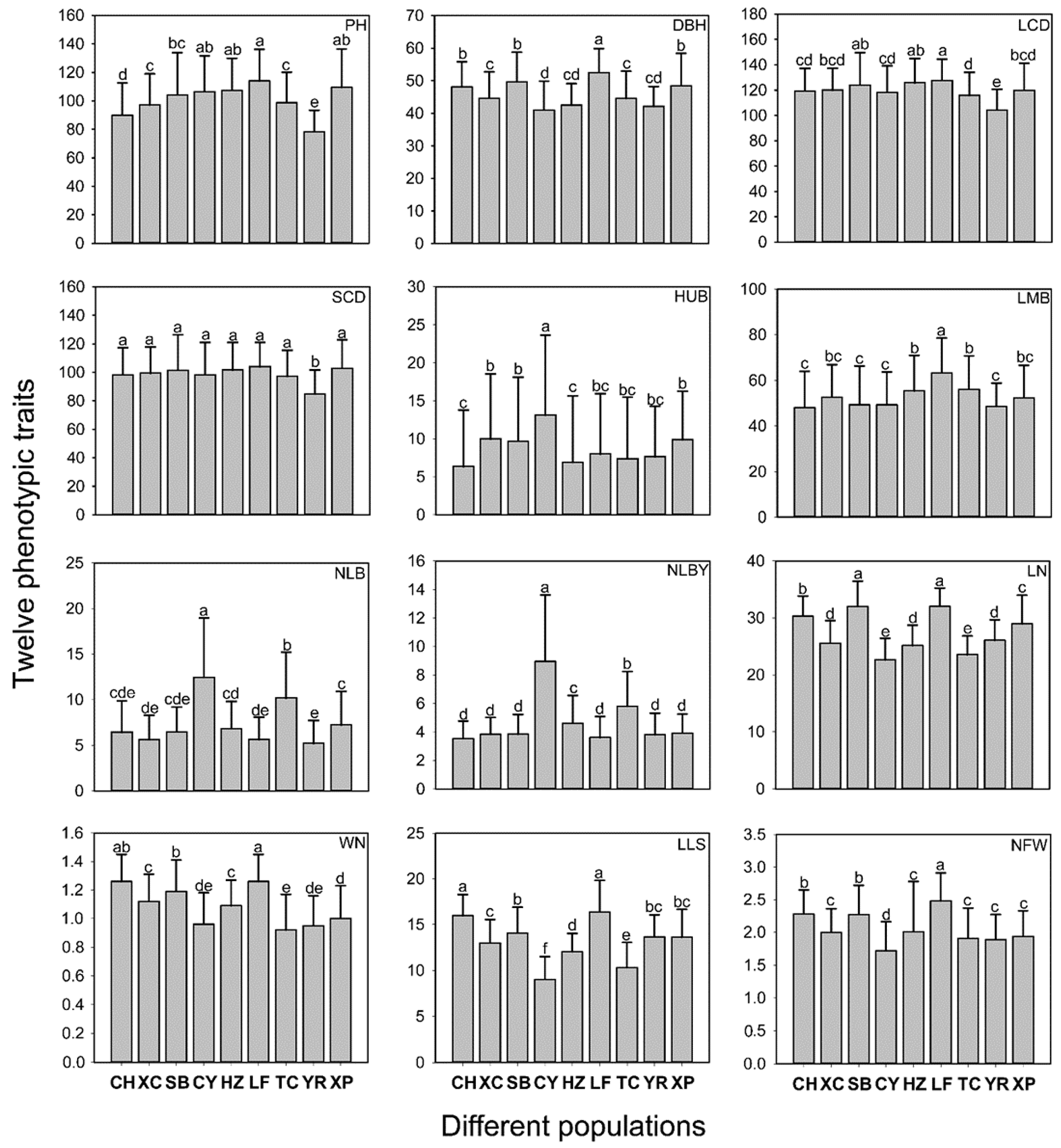
3.1. Phenotypic Variation among and within Populations
3.2. Phenotypic Differentiation among Populations
3.3. Variation Characteristics of Phenotypic Traits
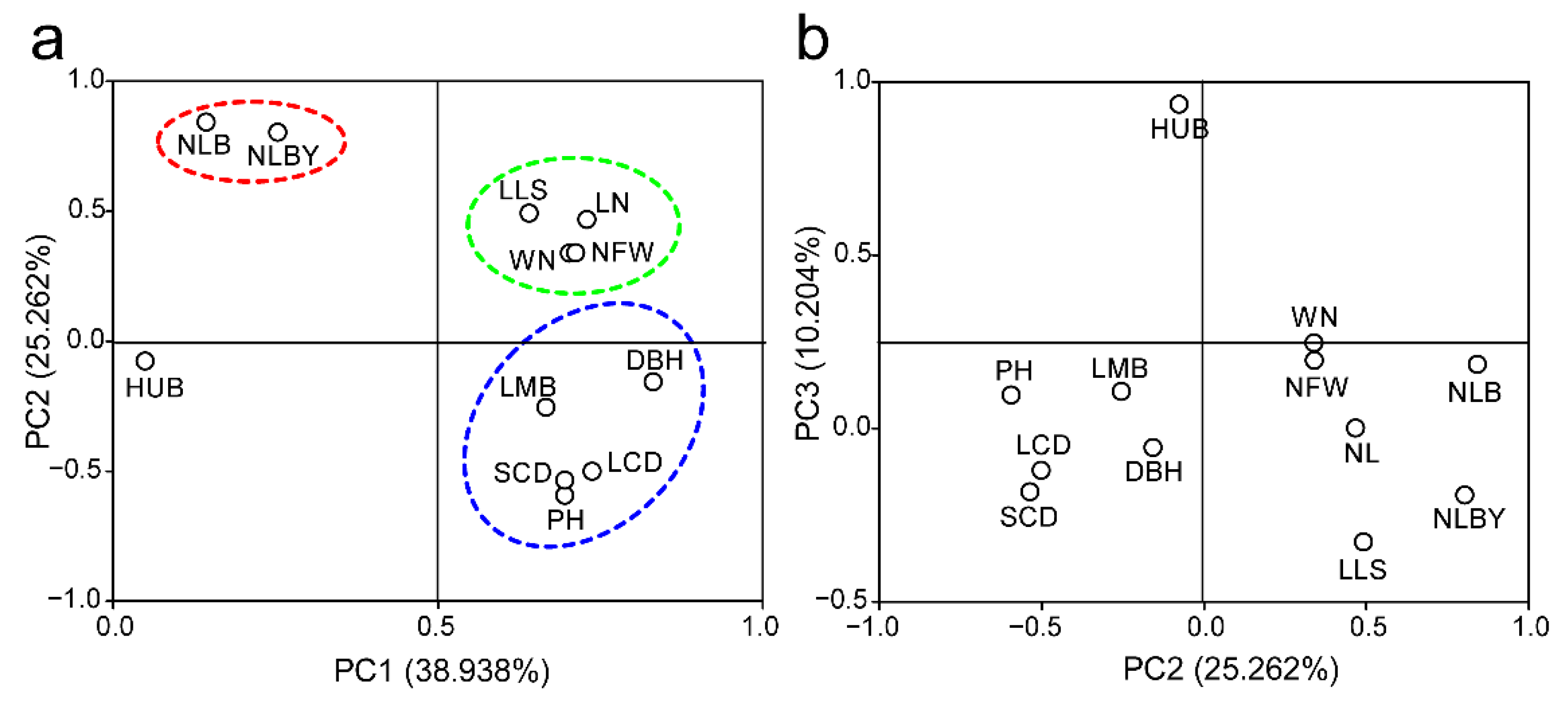
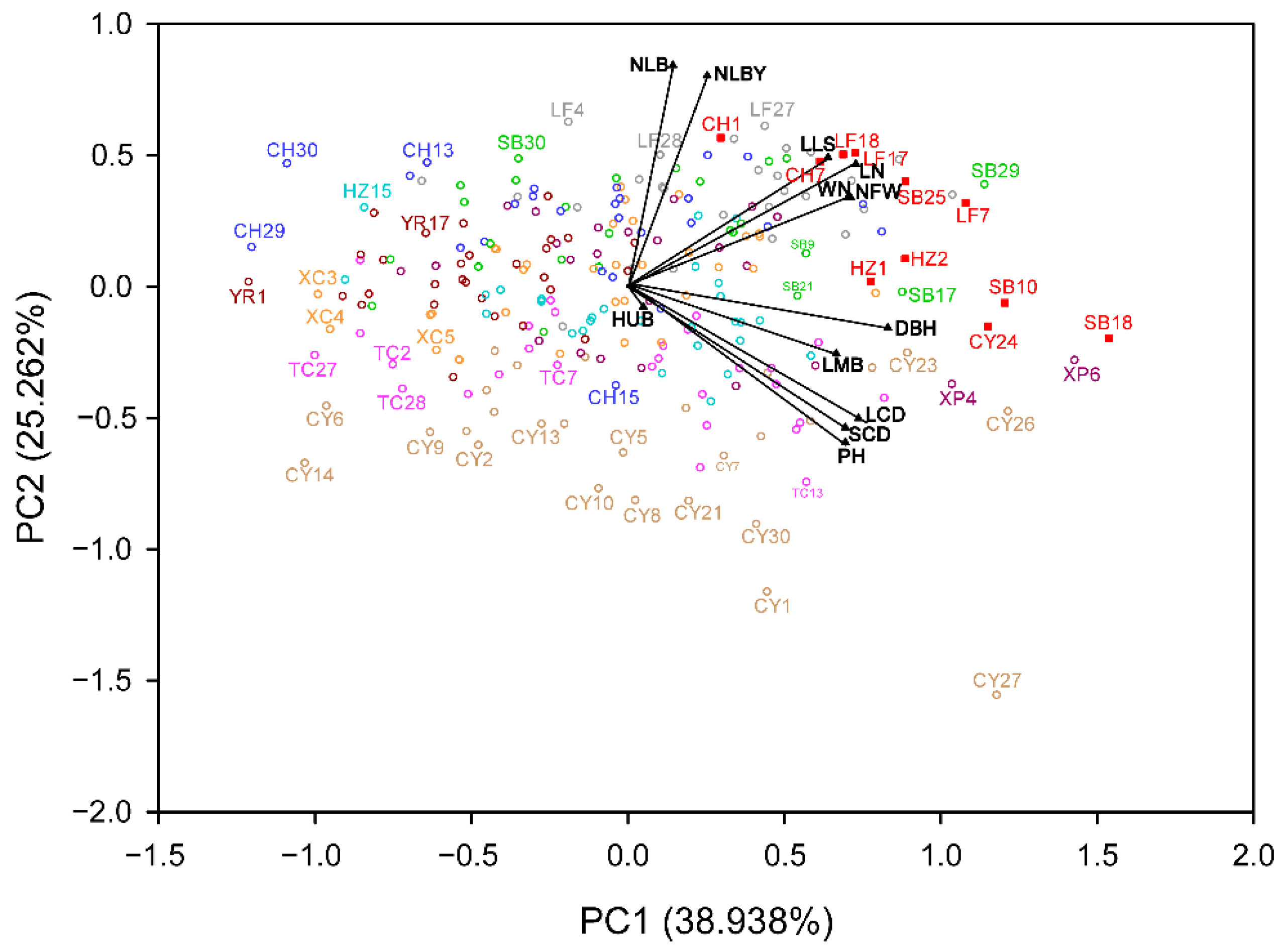
3.4. Principal Component Analysis of Phenotypic Traits
3.5. Correlation between Phenotypic Traits and Spatial Arrangement and Ecological Factors
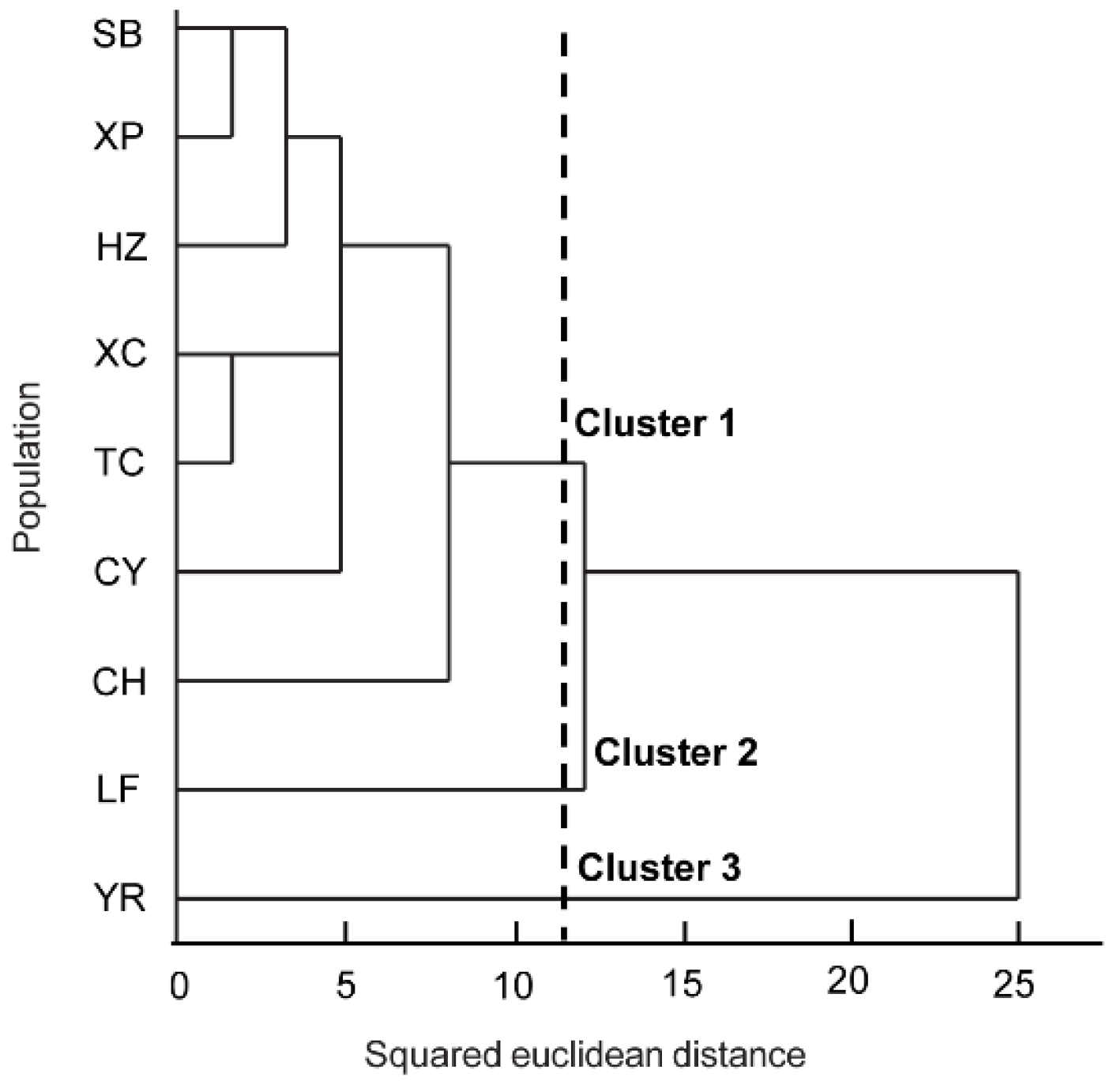
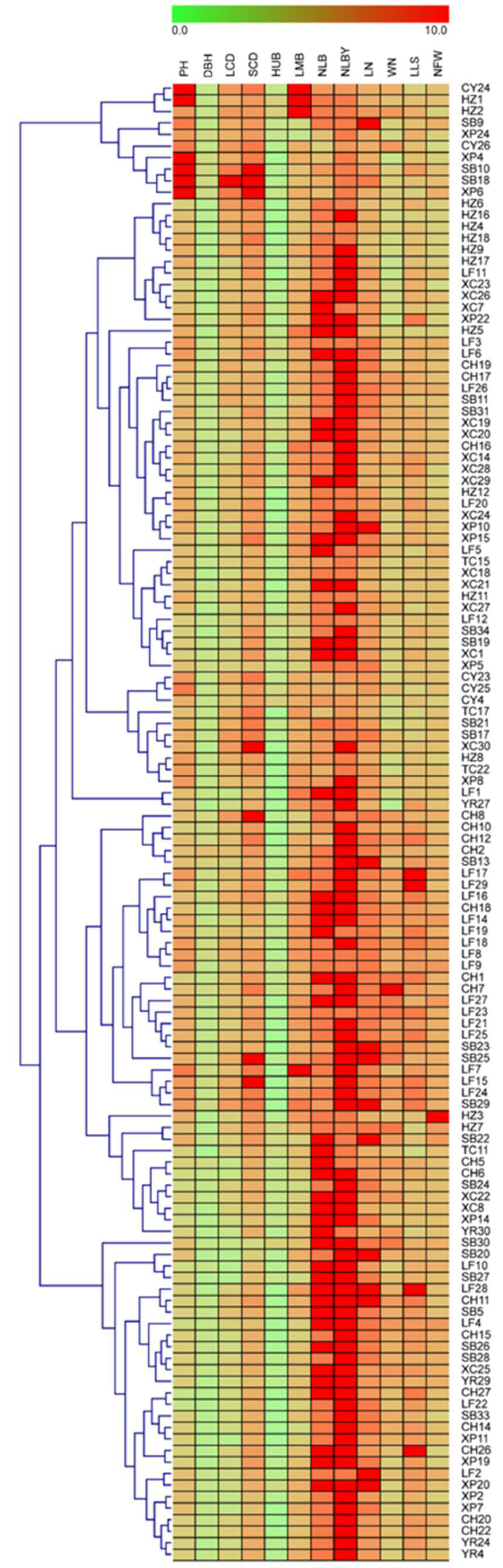
3.6. Cluster Analysis of P. yunanensis Population
3.7. Superior Family Selection of P. yunnanensis
4. Discussion
4.1. Sources of Phenotypic Variation in Different Populations
4.2. Phenotypic Variation Characteristics of Populations
4.3. Correlation between Phenotypic Traits and Spatial Arrangement and Ecological Factors
4.4. Superior Family Selection
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
References
- Sun, H.-L.; Li, S.-C.; Xiong, W.-L.; Yang, Z.-R.; Cui, B.-S.; Yang, T. Influence of slope on root system anchorage of Pinus yunnanensis. Ecol. Eng. 2008, 32, 60–67. [Google Scholar] [CrossRef]
- Li, S.; Huang, X.; Lang, X.; Xu, F.; Li, H.; Zheng, M.; Su, J. Effect of selective logging on soil microbial communities in a Pinus yunnanensis forest. Land Degrad. Dev. 2020, 31, 2268–2280. [Google Scholar] [CrossRef]
- Yang, J.; Zhang, Z.; Dawazhaxi; Wang, B.; Li, Q.; Yu, Q.; Ou, X.; Ali, K. Spatial distribution patterns and intra-specific competition of pine (Pinus yunnanensis) in abandoned farmland under the Sloping Land Conservation Program. Ecol. Eng. 2019, 135, 17–27. [Google Scholar] [CrossRef]
- Gao, X.; Zhou, X.; Wang, H.; Kong, X.; Zhang, S.; Wang, Z.; Wu, W.; Zhang, Z.; Lieutier, F. Influence of severe drought on the resistance of Pinus yunnanensis to a bark beetle-associated fungus. For. Pathol. 2017, 47, e12345. [Google Scholar] [CrossRef]
- Cai, N.; Xu, Y.; Chen, S.; He, B.; Li, G.; Li, Y.; Duan, A. Variation in seed and seedling traits and their relations to geo-climatic factors among populations in Yunnan Pine (Pinus yunnanensis). J. For. Res. 2016, 27, 1009–1017. [Google Scholar] [CrossRef]
- EFSA Panel on Food Additives and Nutrient Sources Added to Food (ANS); Younes, M.; Aggett, P.; Aguilar, F.; Crebelli, R.; Filipič, M.; Frutos, M.J.; Galtier, P.; Gott, D.; Gundert-Remy, U.; et al. Re-evaluation of glycerol esters of wood rosin (E 445) as a food additive. EFSA J. 2018, 16, e05370. [Google Scholar] [CrossRef] [Green Version]
- Zhu, S.; Xu, S.; Jing, W.; Zhao, Z.; Jiang, J. Synthesis and Herbicidal Activities of p-Menth-3-en-1-amine and Its Schiff Base Derivatives. J. Agric. Food Chem. 2016, 64, 9702–9707. [Google Scholar] [CrossRef]
- Xu, S.; Zeng, X.; Dai, S.; Wang, J.; Chen, Y.; Song, J.; Shi, Y.; Cheng, X.; Liao, S.; Zhao, Z. Turpentine Derived Secondary Amines for Sustainable Crop Protection: Synthesis, Activity Evaluation and QSAR Study. J. Agric. Food Chem. 2020, 68, 11829–11838. [Google Scholar] [CrossRef]
- Donoso, D.; Ballesteros, R.; Bolonio, D.; García-Martínez, M.-J.; Lapuerta, M.; Canoira, L. Hydrogenated Turpentine: A Biobased Component for Jet Fuel. Energy Fuels 2020, 35, 1465–1475. [Google Scholar] [CrossRef]
- Edelaar, P.; Jovani, R.; Gomez-Mestre, I. Should I Change or Should I Go? Phenotypic Plasticity and Matching Habitat Choice in the Adaptation to Environmental Heterogeneity. Am. Nat. 2017, 190, 506–520. [Google Scholar] [CrossRef]
- Stöcklin, J.; Kuss, P.; Pluess, A.R. Genetic diversity, phenotypic variation and local adaptation in the alpine landscape: Case studies with alpine plant species. Bot. Helv. 2009, 119, 125–133. [Google Scholar] [CrossRef]
- Khadivi-Khub, A.; Ebrahimi, A.; Sheibani, F.; Esmaeili, A. Phenological and pomological characterization of Persian walnut to select promising trees. Euphytica 2015, 205, 557–567. [Google Scholar] [CrossRef]
- Xu, Y.; Zhou, L.; Cai, N.; Deng, L.; Wang, D.; Duan, A.; He, C.; Xu, Y. Needle Phenotypic Variation among Populations of Pinus yunnanensis at Different Altitude. J. Yunnan Agric. Univ. Nat. Sci. 2016, 31, 109–114. [Google Scholar]
- Kuehne, C.; Pyttel, P.; Modrow, T.; Kohnle, U.; Bauhus, J. Seedling development and regeneration success after 10 years following group selection harvesting in a sessile oak (Quercus petraea [Mattuschka] Liebl.) stand. Ann. For. Sci. 2020, 77, 71. [Google Scholar] [CrossRef]
- Khadivi-Khub, A.; Ebrahimi, A.; Mohammadi, A.; Kari, A. Characterization and selection of walnut (Juglans regia L.) genotypes from seedling origin trees. Tree Genet. Genomes 2015, 11, 54. [Google Scholar] [CrossRef]
- Mwase, W.F.; Savill, P.S.; Hemery, G. Genetic parameter estimates for growth and form traits in common ash (Fraxinus excelsior, L.) in a breeding seedling orchard at Little Wittenham in England. New For. 2008, 36, 225–238. [Google Scholar] [CrossRef]
- Bogdan, S.; Katičić-Trupčević, I.; Kajba, D. Genetic Variation in Growth Traits in a Quercus robur L. Open-Pollinated Progeny Test of the Slavonian Provenance. Silvae Genet. 2004, 53, 198–201. [Google Scholar] [CrossRef] [Green Version]
- Cornelius, J.P.; Mesen, J.F. Provenance and family variation in growth rate, stem straightness, and foliar mineral concentration in Vochysia guatemalensis. Can. J. For. Res. 1997, 27, 1103–1109. [Google Scholar] [CrossRef]
- Espinoza, S.E.; Quiroz, I.A.; Magni, C.R.; Yáñez, M.A. Variation in early growth and survival of Pinus ponderosa provenances established in Southern Chile. New For. 2021, 52, 959–970. [Google Scholar] [CrossRef]
- Kavaliauskas, D.; Šeho, M.; Baier, R.; Fussi, B. Genetic variability to assist in the delineation of provenance regions and selection of seed stands and gene conservation units of wild service tree (Sorbus torminalis (L.) Crantz) in southern Germany. Eur. J. For. Res. 2021, 140, 551–565. [Google Scholar] [CrossRef]
- Buras, A.; Sass-Klaassen, U.; Verbeek, I.; Copini, P. Provenance selection and site conditions determine growth performance of pedunculate oak. Dendrochronologia 2020, 61, 125705. [Google Scholar] [CrossRef]
- Zhang, X.-M.; Gao, L.-M.; Möller, M.; Li, D.-Z. Molecular evidence for fragmentation among populations of Taxus wallichiana var. mairei, a highly endangered conifer in China. Can. J. For. Res. 2009, 39, 755–764. [Google Scholar] [CrossRef]
- Gandour, M.; Khouja, M.L.; Toumi, L.; Triki, S. Morphological evaluation of cork oak (Quercus suber): Mediterranean provenance variability in Tunisia. Ann. For. Sci. 2007, 64, 549–555. [Google Scholar] [CrossRef] [Green Version]
- Du, Q.; Xu, B.; Gong, C.; Yang, X.; Pan, W.; Tian, J.; Li, B.; Zhang, D. Variation in growth, leaf, and wood property traits of Chinese white poplar (Populus tomentosa), a major industrial tree species in Northern China. Can. J. For. Res. 2014, 44, 326–339. [Google Scholar] [CrossRef]
- Zhang, Z.; Zhang, H.; Yang, C.; Zhang, L.; Du, J.; Jiang, Y. Clonal variations in nutritional components of Pinus koreansis seeds collected from seed orchards in Northeastern China. J. For. Res. 2015, 27, 295–311. [Google Scholar] [CrossRef]
- Chaix, G.; Monteuuis, O.; Garcia, C.; Alloysius, D.; Gidiman, J.; Bacilieri, R.; Goh, D.K.S. Genetic variation in major phenotypic traits among diverse genetic origins of teak (Tectona grandis L.f.) planted in Taliwas, Sabah, East Malaysia. Ann. For. Sci. 2011, 68, 1015–1026. [Google Scholar] [CrossRef]
- Jimu, L.; Chen, S.F.; Wingfield, M.J.; Mwenje, E.; Roux, J. The Eucalyptus stem canker pathogen Teratosphaeria gauchensis represents distinct genetic groups in Africa and South America. For. Pathol. 2016, 46, 229–239. [Google Scholar] [CrossRef]
- Wee, A.K.S.; Li, C.; Dvorak, W.S.; Hong, Y. Genetic diversity in natural populations of Gmelina arborea: Implications for breeding and conservation. New For. 2012, 43, 411–428. [Google Scholar] [CrossRef]
- Belalia, N.; Lupini, A.; Djemel, A.; Morsli, A.; Mauceri, A.; Lotti, C.; Khelifi-Slaoui, M.; Khelifi, L.; Sunseri, F. Analysis of genetic diversity and population structure in Saharan maize (Zea mays L.) populations using phenotypic traits and SSR markers. Genet. Resour. Crop Evol. 2018, 66, 243–257. [Google Scholar] [CrossRef]
- Garcia, R.; Siepielski, A.M.; Benkman, C.W. Cone and seed trait variation in whitebark pine (Pinus albicaulis; Pinaceae) and the potential for phenotypic selection. Am. J. Bot. 2009, 96, 1050–1054. [Google Scholar] [CrossRef] [Green Version]
- Wei, Z.-Z.; Zhao, X.; Pan, W.; Zhang, J.-F.; Li, B.-L.; Zhang, D.-Q. Phenotypic variation among five provenances of Populus simonii in northern China. For. Stud. China 2011, 13, 97–103. [Google Scholar] [CrossRef]
- Jensen, J.S. Provenance Variation in Phenotypic Traits in Quercus robur and Quercus petraea in Danish Provenance Trials. Scand. J. For. Res. 2000, 15, 297–308. [Google Scholar] [CrossRef]
- Duan, H.; Cao, S.; Zheng, H.; Hu, D.; Lin, J.; Lin, H.; Hu, R.; Sun, Y.; Li, Y. Variation in the Growth Traits and Wood Properties of Chinese Fir from Six Provinces of Southern China. Forests 2016, 7, 192. [Google Scholar] [CrossRef] [Green Version]
- Ebrahimi, A.; Asadi, A.; Monfared, S.R.; Sahebi, M.; Rezaee, S.; Khaledian, Y. Evaluation of phenotypic diversity of the endangered orchid (Orchis mascula): Emphasizing on breeding, conservation and development. S. Afr. J. Bot. 2020, 132, 304–315. [Google Scholar] [CrossRef]
- Yang, S.; Guo, N.; Ge, H. Morphological and AFLP-Based Genetic Diversity in Rosa platyacantha Population in Eastern Tianshan Mountains of Northwestern China. Hortic. Plant J. 2016, 2, 55–60. [Google Scholar] [CrossRef] [Green Version]
- Han, H.; Li, S.; Gan, X.; Zhang, X. Phenotypic diversity in natural populations of an endangered plant Tetracentron sinense. Bot. Sci. 2017, 95, 283. [Google Scholar] [CrossRef] [Green Version]
- Li, W.; Wang, P.; Qi, Q.; Zhang, Q.; Gao, X.; Lin, M.; Cui, Y. Phenotypic Diversity and Variation of Lonicera caerulea Populations in the Changbai Mountain Alongside the Elevation Gradient. Pol. J. Environ. Stud. 2020, 30, 705–716. [Google Scholar] [CrossRef]
- Wang, B.; Mao, J.-F.; Zhao, W.; Wang, X.-R. Impact of Geography and Climate on the Genetic Differentiation of the Subtropical Pine Pinus yunnanensis. PLoS ONE 2013, 8, e67345. [Google Scholar] [CrossRef]
- Xu, Y.L.; Cai, N.H.; Shi, C.; Wang, D.W.; He, C.Z.; Duan, A.A.; Kang, X.Y. Relationships between the genetic diversity of Pinus yunnanensis Franch natural populations and ecological factors. Chin. J. Ecol. 2016, 35, 1767–1775. [Google Scholar] [CrossRef]
- Pretzsch, H. Genetic diversity reduces competition and increases tree growth on a Norway spruce (Picea abies [L.] Karst.) provenance mixing experiment. For. Ecol. Manag. 2021, 497, 119498. [Google Scholar] [CrossRef]
- Hai, P.H.; Jansson, G.; Harwood, C.; Hannrup, B.; Thinh, H.H. Genetic variation in growth, stem straightness and branch thickness in clonal trials of Acacia auriculiformis at three contrasting sites in Vietnam. For. Ecol. Manag. 2008, 255, 156–167. [Google Scholar] [CrossRef]
- Kostić, S.; Orlović, S.; Karaklić, V.; Kesić, L.; Zorić, M.; Stojanović, D. Allometry and Post-Drought Growth Resilience of Pedunculate Oak (Quercus robur L.) Varieties. Forests 2021, 12, 930. [Google Scholar] [CrossRef]
- Shen, J.; Li, Z.; Gao, C.; Li, S.; Huang, X.; Lang, X.; Su, J. Radial growth response of Pinus yunnanensis to rising temperature and drought stress on the Yunnan Plateau, southwestern China. For. Ecol. Manag. 2020, 474, 118357. [Google Scholar] [CrossRef]
- Huang, Y.; Mao, J.; Chen, Z.; Meng, J.; Xu, Y.; Duan, A.; Li, Y. Genetic structure of needle morphological and anatomical traits of Pinus yunnanensis. J. For. Res. 2016, 27, 13–25. [Google Scholar] [CrossRef]
- Amodei, T.; Guibal, F.; Fady, B. Relationships between climate and radial growth in black pine (Pinus nigra Arnold ssp. salzmannii (Dunal) Franco) from the south of France. Ann. For. Sci. 2013, 70, 41–47. [Google Scholar] [CrossRef] [Green Version]
- Stajić, B.; Kazimirović, M.; Dukić, V.; Radaković, N. First Dendroclimatological Insight into Austrian Pine (Pinus nigra Arnold) Climate-Growth Relationship in Belgrade Area, Serbia. Seefor-South-East Eur. For. 2020, 11, 127–134. [Google Scholar] [CrossRef]
- Lo, Y.-H.; Blanco, J.A.; Seely, B.; Welham, C.; Kimmins, J. Relationships between climate and tree radial growth in interior British Columbia, Canada. For. Ecol. Manag. 2010, 259, 932–942. [Google Scholar] [CrossRef]
- Zhang, H.; Zhou, X.; Gu, W.; Wang, L.; Li, W.; Gao, Y.; Wu, L.; Guo, X.; Tigabu, M.; Xia, D.; et al. Genetic stability of Larix olgensis provenances planted in different sites in northeast China. For. Ecol. Manag. 2021, 485, 118988. [Google Scholar] [CrossRef]
- Xu, Y.; Cai, N.; Woeste, K.; Kang, X.; Chengzhong, H.; Li, G.; Chen, S.; Duan, A.; Yulan, X.; Nianhui, C.; et al. Genetic Diversity and Population Structure of Pinus yunnanensis by Simple Sequence Repeat Markers. For. Sci. 2016, 62, 38–47. [Google Scholar] [CrossRef]
- Gauli, A.; Gailing, O.; Stefenon, V.M.; Finkeldey, R. Genetic similarity of natural populations and plantations of Pinus roxburghii Sarg. in Nepal. Ann. For. Sci. 2009, 66, 703. [Google Scholar] [CrossRef] [Green Version]
- Rajeb, C.; Messaoud, C.; Chograni, H.; Bejaoui, A.; Boulila, A.; Rejeb, M.N.; Boussaid, M. Genetic diversity in Tunisian Crataegus azarolus L. var. aronia L. populations assessed using RAPD markers. Ann. For. Sci. 2010, 67, 512. [Google Scholar] [CrossRef] [Green Version]
- Wahid, N.; Gonzalez-Martinez, S.C.; El Hadrami, I.; Boulli, A. Variation of morphological traits in natural populations of maritime pine (Pinus pinaster Ait.) in Morocco. Ann. For. Sci. 2006, 63, 83–92. [Google Scholar] [CrossRef] [Green Version]
- Toft, B.D.; Alam, M.; Topp, B. Estimating genetic parameters of architectural and reproductive traits in young macadamia cultivars. Tree Genet. Genomes 2018, 14, 50. [Google Scholar] [CrossRef]
- Padi, F.K.; Opoku, S.Y.; Adomako, B.; Adu-Ampomah, Y. Effectiveness of juvenile tree growth rate as an index for selecting high yielding cocoa families. Sci. Hortic. 2012, 139, 14–20. [Google Scholar] [CrossRef]
- Verardi, C.K.; de Oliveira, A.L.B.; Silva, G.A.P.; Gouvêa, L.R.L.; Gonçalves, P.D.S. Comparison between different selection methods of rubber trees. Ind. Crop. Prod. 2014, 52, 255–263. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Population | Individual Samples | LON (°E) | LAT (°N) | EL (m) | MAP (mm) | MAT (°C) | WP (mm) | WT (°C) | DP (mm) | DT (°C) |
|---|---|---|---|---|---|---|---|---|---|---|
| Ceheng (CH) | 90 | 105.93 | 24.85 | 800 | 1256 | 17.9 | 705 | 24.6 | 29.5 | 9.9 |
| Xichang (XC) | 87 | 102.01 | 27.87 | 2610 | 918 | 7.8 | 518 | 14.0 | 22 | 0.3 |
| Shuangbai (SB) | 84 | 101.65 | 24.37 | 1655 | 1004 | 20.7 | 567 | 24.9 | 29.4 | 16.8 |
| Chayu (CY) | 84 | 97.35 | 28.62 | 2050 | 791 | 4.9 | 524 | 11.5 | 42 | −0.9 |
| Huize (HZ) | 90 | 103.40 | 26.40 | 2320 | 928 | 10.5 | 516 | 16.4 | 28 | 3.7 |
| Lufeng (LF) | 87 | 101.90 | 25.13 | 1925 | 817 | 15.9 | 450 | 20.9 | 39 | 11.4 |
| Tianchi (TC) | 90 | 98.32 | 25.37 | 2125 | 1442 | 12.6 | 743 | 17.4 | 76 | 6.8 |
| Yongren (YR) | 90 | 101.60 | 26.34 | 2055 | 825 | 13.9 | 467 | 19.0 | 29 | 7.9 |
| Xinping (XP) | 72 | 102.07 | 24.07 | 1600 | 974 | 18.2 | 550 | 22.5 | 27.2 | 12.3 |
| Traits | F Value | Proportion of Variance Components (%) | Phenotypic Differentiation Coefficients% | |||
|---|---|---|---|---|---|---|
| Among Populations | Within Population | Among Populations | Within Population | Random Error | ||
| PH | 19.92 ** | 2.12 ** | 115.01 | 22.49 | 6.15 | 90.39 |
| DBH | 20.91 ** | 1.90 ** | 14.61 | 2.19 | 62.54 | 91.69 |
| LCD | 11.01 ** | 1.69 * | 42.45 | 9.80 | 2.83 | 86.67 |
| SCD | 7.23 ** | 1.38 | 28.06 | 6.06 | 2.45 | 83.97 |
| HUB | 5.40 ** | 1.67 * | 3.60 | 1.76 | 65.73 | 76.39 |
| LMB | 10.04 ** | 1.64 * | 21.79 | 4.75 | 3.47 | 85.98 |
| NLB | 35.11 ** | 1.40 | 5.63 | 0.23 | 14.13 | 96.18 |
| NLBY | 54.59 ** | 0.92 | 3.04 | 0.00 | 4.75 | 98.34 |
| LN | 76.91 ** | 2.12 ** | 12.51 | 0.61 | 14.12 | 97.32 |
| WN | 34.38 ** | 1.26 | 0.02 | 0.01 | 0.04 | 96.45 |
| LLS | 74.72 ** | 2.68 ** | 5.71 | 0.44 | 6.73 | 96.54 |
| NFW | 22.38 ** | 1.22 | 0.05 | 0.02 | 0.22 | 94.82 |
| Mean | 21.04 | 4.03 | 15.26 | 91.23 | ||
| Traits | Population | Mean | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| CH | XC | SB | CY | HZ | LF | TC | YR | XP | ||
| PH | 25.19 | 22.66 | 28.66 | 23.76 | 21.10 | 19.48 | 21.78 | 19.35 | 24.73 | 22.97 |
| DBH | 16.13 | 18.07 | 18.45 | 21.86 | 15.71 | 14.05 | 18.84 | 14.56 | 20.52 | 17.58 |
| LCD | 15.08 | 14.50 | 20.73 | 17.67 | 15.31 | 13.14 | 15.77 | 15.87 | 17.93 | 16.22 |
| SCD | 19.42 | 18.69 | 24.64 | 23.20 | 19.22 | 16.57 | 18.90 | 20.10 | 19.42 | 20.02 |
| HUB | 66.46 | 70.63 | 62.38 | 78.16 | 103.10 | 71.15 | 75.88 | 47.48 | 56.42 | 70.18 |
| LMB | 32.93 | 27.17 | 34.67 | 29.44 | 28.30 | 24.27 | 26.24 | 21.26 | 27.50 | 27.98 |
| NLB | 53.60 | 47.84 | 42.23 | 52.79 | 43.78 | 43.65 | 49.35 | 47.59 | 51.16 | 48.00 |
| NLBY | 34.92 | 30.99 | 35.99 | 52.31 | 42.23 | 40.45 | 42.25 | 39.90 | 34.21 | 39.25 |
| LN | 11.70 | 15.46 | 13.97 | 16.78 | 14.29 | 9.98 | 13.97 | 13.85 | 17.55 | 14.17 |
| WN | 15.32 | 17.28 | 18.76 | 23.24 | 16.68 | 15.51 | 26.88 | 22.31 | 23.24 | 19.91 |
| LLS | 14.63 | 19.92 | 20.19 | 27.49 | 16.65 | 21.06 | 26.73 | 17.74 | 22.53 | 20.77 |
| NFW | 16.20 | 18.11 | 20.13 | 25.72 | 38.33 | 17.57 | 24.31 | 20.47 | 19.99 | 22.31 |
| Mean | 26.80 | 26.78 | 28.40 | 32.70 | 31.22 | 25.57 | 30.08 | 25.04 | 27.93 | 28.28 |
| Traits | Mean | Range | SD | Basic Value | CV (%) |
|---|---|---|---|---|---|
| PH | 100.20 | 126.82 | 21.52 | 121.72 | 21.48 |
| DBH | 46.26 | 93.10 | 8.46 | 54.72 | 18.29 |
| LCD | 119.45 | 87.33 | 15.21 | 134.66 | 12.73 |
| SCD | 98.22 | 121.00 | 15.55 | 113.77 | 15.83 |
| HUB | 8.83 | 56.33 | 6.64 | 15.47 | 75.20 |
| LMB | 52.55 | 85.67 | 12.04 | 64.59 | 22.91 |
| NLB | 0.82 | 0.90 | 0.12 | 0.94 | 14.63 |
| NLBY | 0.89 | 0.82 | 0.09 | 0.98 | 10.11 |
| LN | 27.21 | 36.85 | 4.65 | 31.86 | 17.09 |
| WN | 1.08 | 0.97 | 0.20 | 1.28 | 18.52 |
| LLS | 13.10 | 19.01 | 3.02 | 16.12 | 23.05 |
| NFW | 2.05 | 3.97 | 0.40 | 2.45 | 19.51 |
| Code | PH | DBH | LCD | SCD | HUB | LMB | NLB | NLBY | LN | WN | LLS | NFW |
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| CY24 | 156.00 | 51.05 | 145.33 | 121.33 | 14.33 | 83.67 | 0.74 | 0.76 | 26.83 | 1.33 | 11.37 | 2.39 |
| HZ1 | 157.67 | 49.61 | 139.67 | 108.67 | 27.33 | 84.33 | 0.87 | 0.90 | 25.70 | 1.20 | 12.16 | 2.28 |
| HZ2 | 133.67 | 51.38 | 145.67 | 113.67 | 24.33 | 90.33 | 0.86 | 0.82 | 30.00 | 1.39 | 14.92 | 1.94 |
| SB10 | 151.67 | 59.03 | 159.67 | 134.33 | 7.33 | 54.67 | 0.73 | 0.86 | 27.87 | 1.12 | 16.38 | 2.16 |
| SB18 | 164.33 | 64.70 | 171.00 | 141.00 | 10.33 | 55.00 | 0.67 | 0.83 | 30.77 | 1.13 | 11.95 | 2.13 |
| SB25 | 121.00 | 54.95 | 144.67 | 129.33 | 8.33 | 68.67 | 0.82 | 0.94 | 35.13 | 1.48 | 15.39 | 2.58 |
| LF7 | 142.33 | 60.12 | 139.00 | 117.33 | 1.67 | 90.67 | 0.81 | 0.94 | 33.50 | 1.22 | 16.24 | 2.52 |
| CH1 | 98.00 | 52.13 | 129.67 | 109.67 | 13.33 | 57.67 | 0.93 | 0.93 | 33.17 | 1.46 | 17.96 | 2.78 |
| CH7 | 113.00 | 56.20 | 135.33 | 120.67 | 16.33 | 57.67 | 0.86 | 0.92 | 33.90 | 1.59 | 16.08 | 2.63 |
| LF17 | 133.00 | 57.02 | 135.33 | 106.00 | 12.00 | 75.67 | 0.87 | 0.94 | 32.57 | 1.42 | 19.95 | 2.53 |
| LF18 | 129.67 | 57.21 | 141.00 | 97.00 | 8.00 | 72.33 | 0.87 | 0.92 | 33.73 | 1.42 | 17.17 | 2.84 |
| Mean | 136.39 | 55.76 | 144.21 | 118.90 | 13.03 | 71.88 | 0.82 | 0.89 | 31.20 | 1.34 | 15.42 | 2.43 |
| ΔG% | 26.54 | 17.04 | 17.17 | 16.83 | 32.23 | 26.89 | 0.02 | 0.02 | 12.78 | 19.53 | 15.02 | 15.81 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liu, Z.; Gao, C.; Li, J.; Miao, Y.; Cui, K. Phenotypic Diversity Analysis and Superior Family Selection of Industrial Raw Material Forest Species-Pinus yunnanensis Franch. Forests 2022, 13, 618. https://0-doi-org.brum.beds.ac.uk/10.3390/f13040618
Liu Z, Gao C, Li J, Miao Y, Cui K. Phenotypic Diversity Analysis and Superior Family Selection of Industrial Raw Material Forest Species-Pinus yunnanensis Franch. Forests. 2022; 13(4):618. https://0-doi-org.brum.beds.ac.uk/10.3390/f13040618
Chicago/Turabian StyleLiu, Zirui, Chengjie Gao, Jin Li, Yingchun Miao, and Kai Cui. 2022. "Phenotypic Diversity Analysis and Superior Family Selection of Industrial Raw Material Forest Species-Pinus yunnanensis Franch" Forests 13, no. 4: 618. https://0-doi-org.brum.beds.ac.uk/10.3390/f13040618