Variation in Soil Bacterial and Fungal Community Composition at Different Successional Stages of a Broad-Leaved Korean Pine Forest in the Lesser Hinggan Mountains

Abstract
:1. Introduction
2. Materials and Methods
2.1. Description of the Study Area
2.2. Plots Setting, Survey and Sampling
2.3. Soil Characteristics and Enzyme Activity Measurements
2.4. DNA Extraction, PCR Amplification, and Illumina Sequencing
2.5. Statistical Analyses
3. Results
3.1. Change in Soil Properties in the Forest with Different Successional Phases
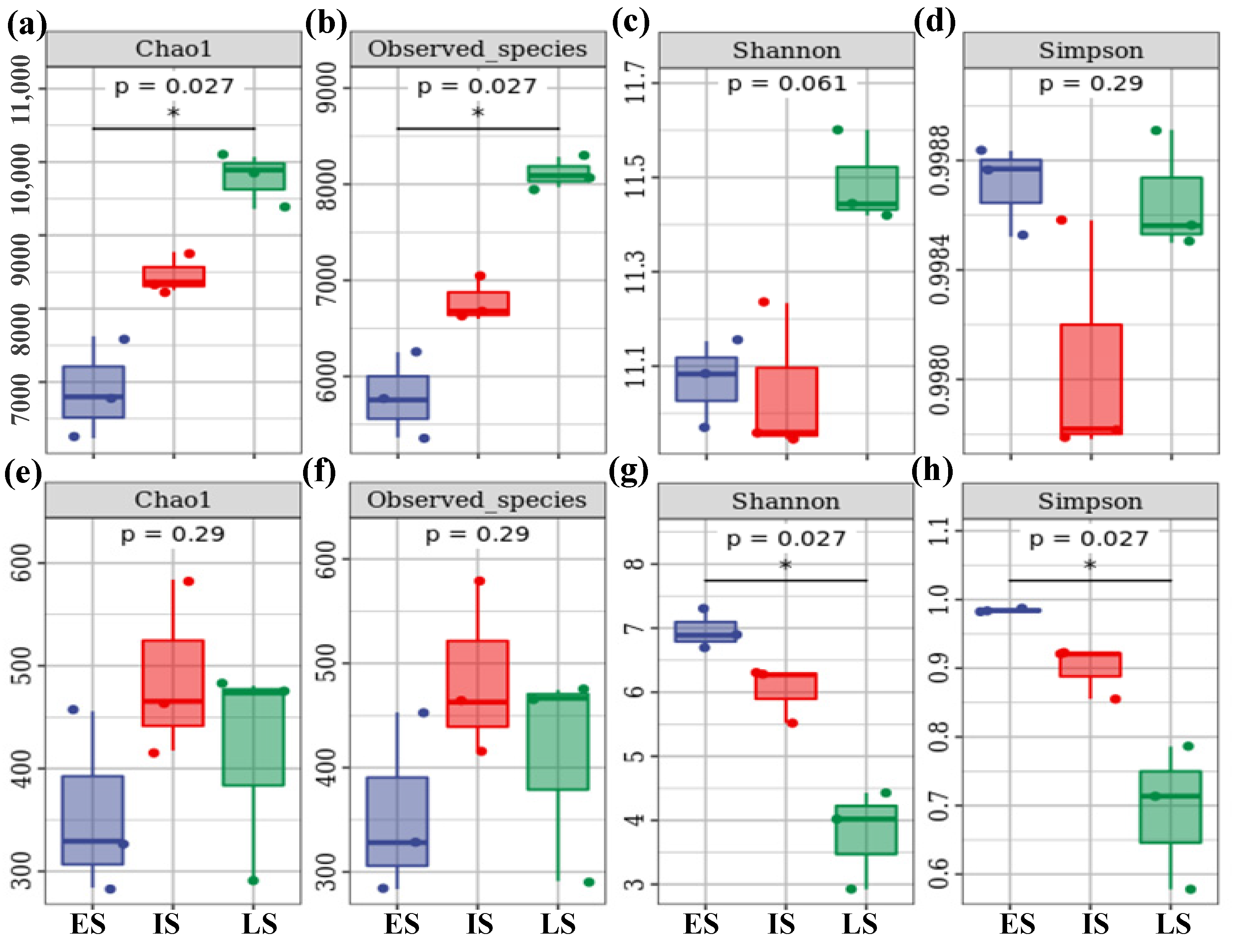
3.2. The Variation in Soil Bacterial and Fungal Alpha-Diversity at Various Successional Phases
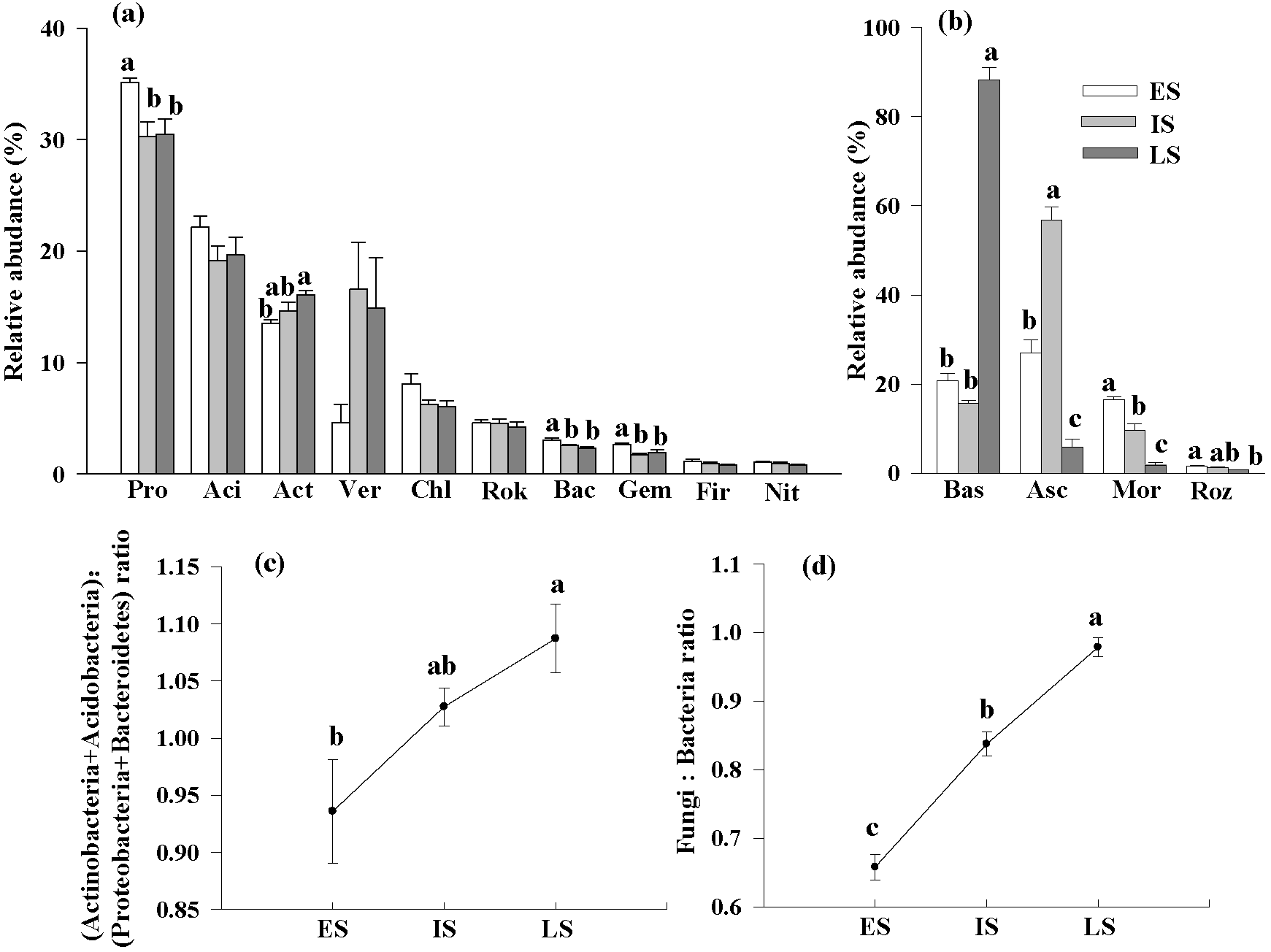
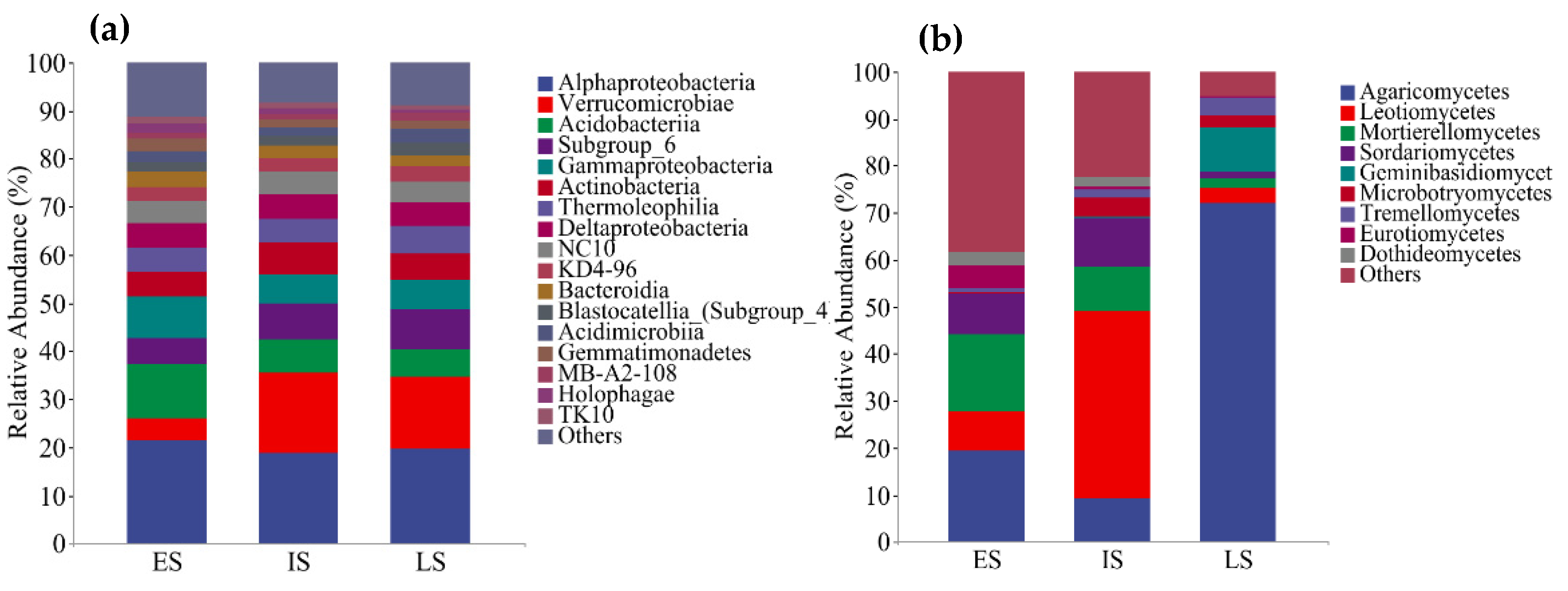
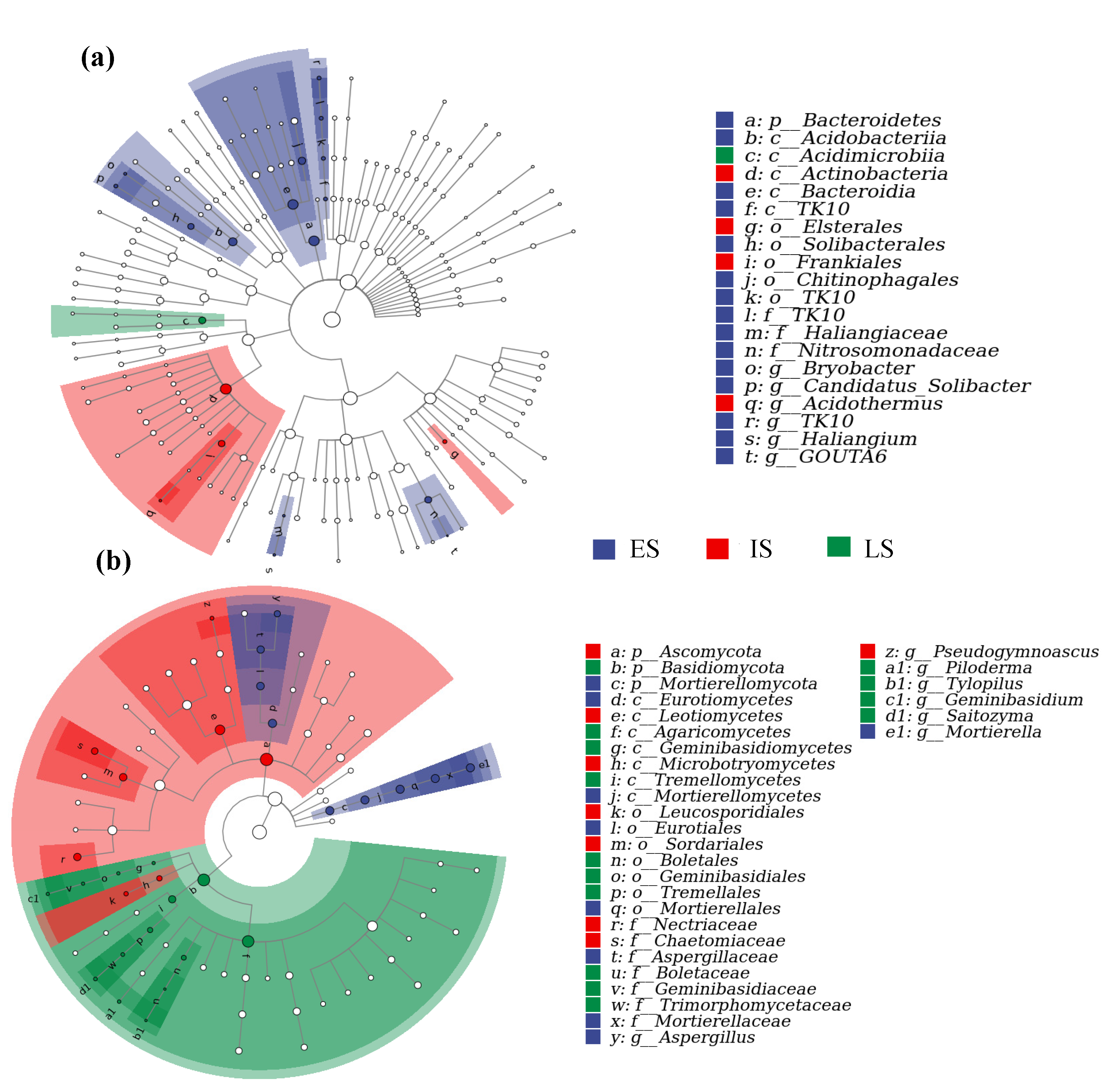
3.3. The Variation in Bacterial and Fungal Community Composition at Various Successional Phases
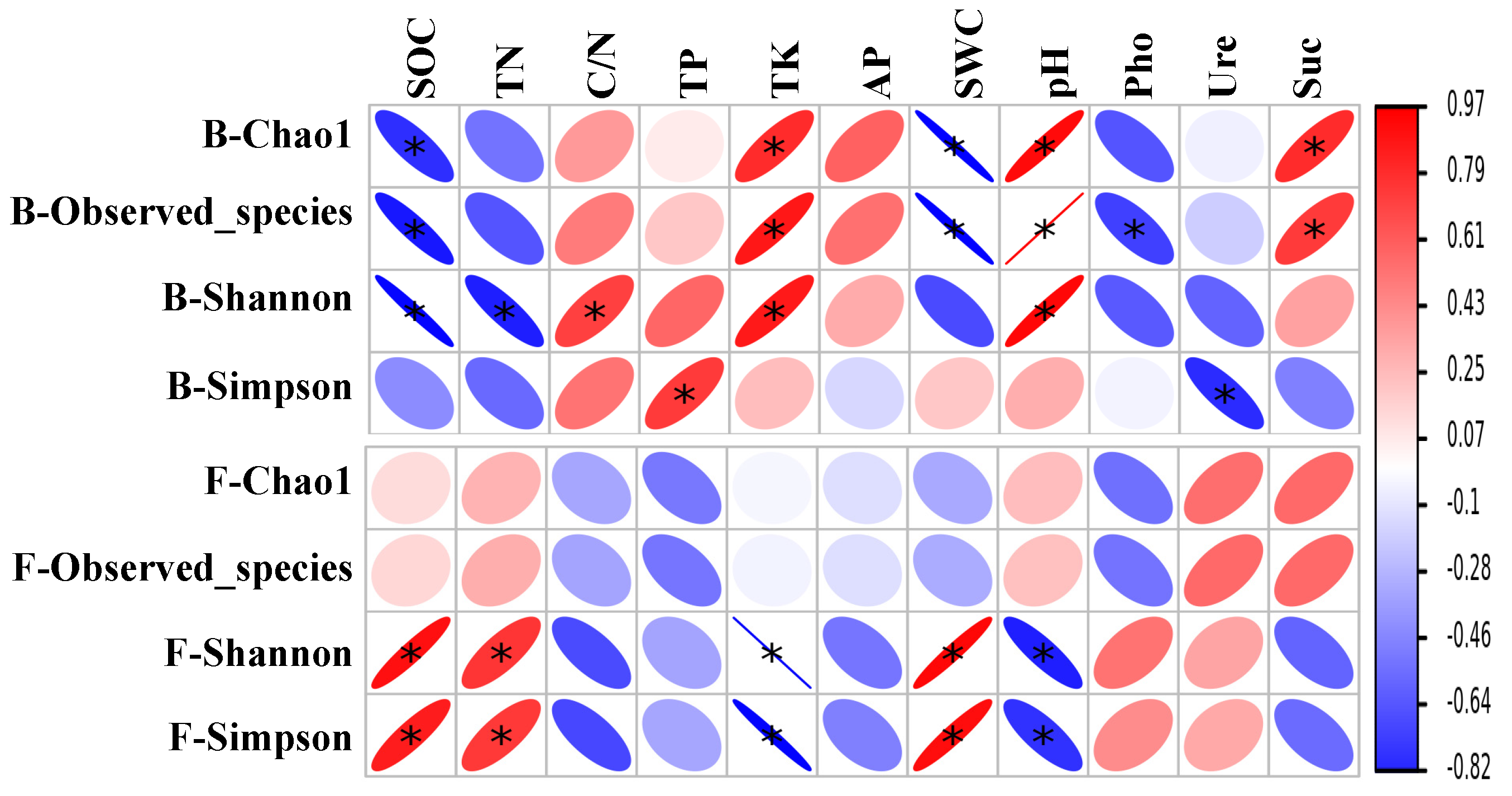
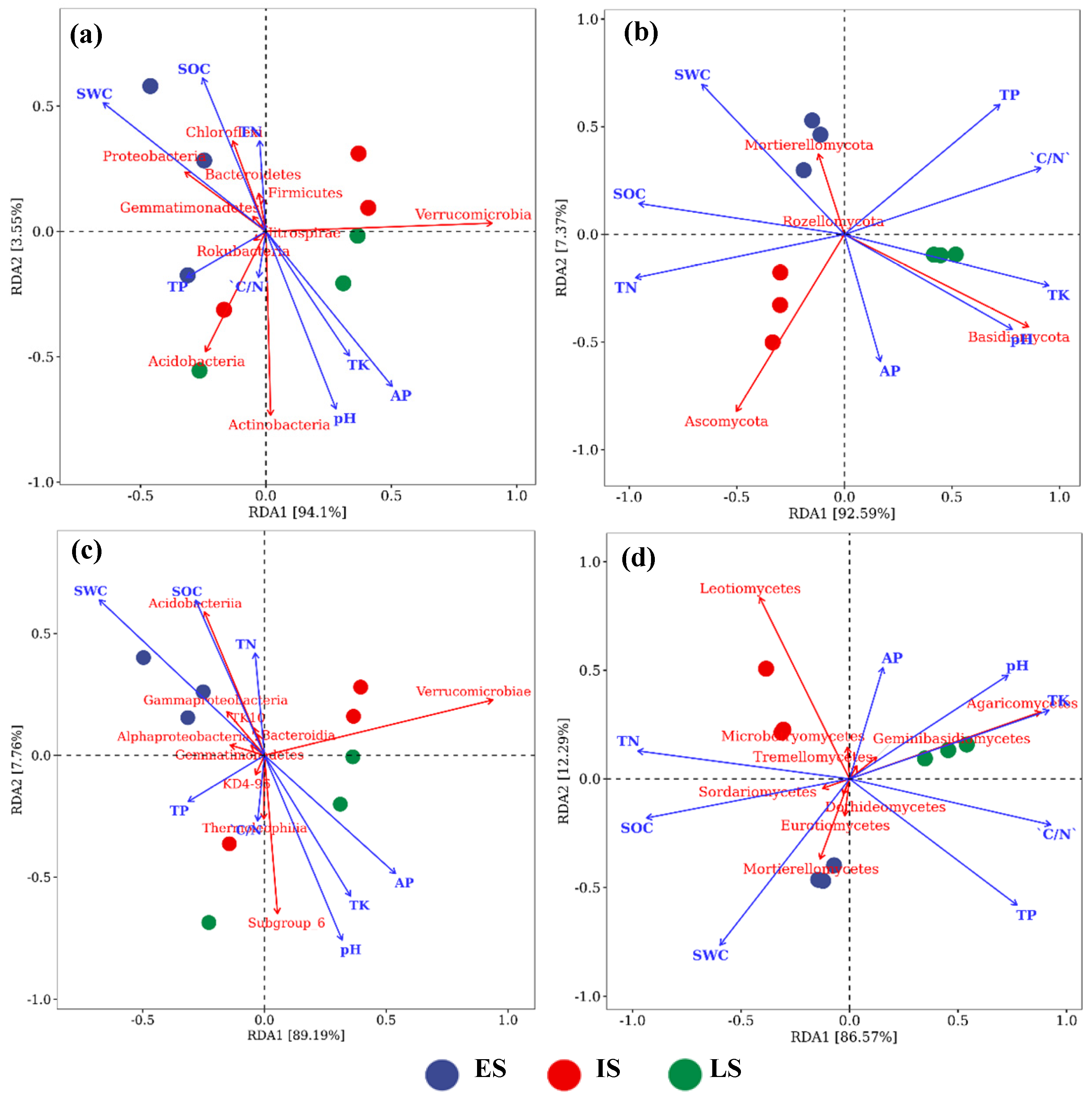
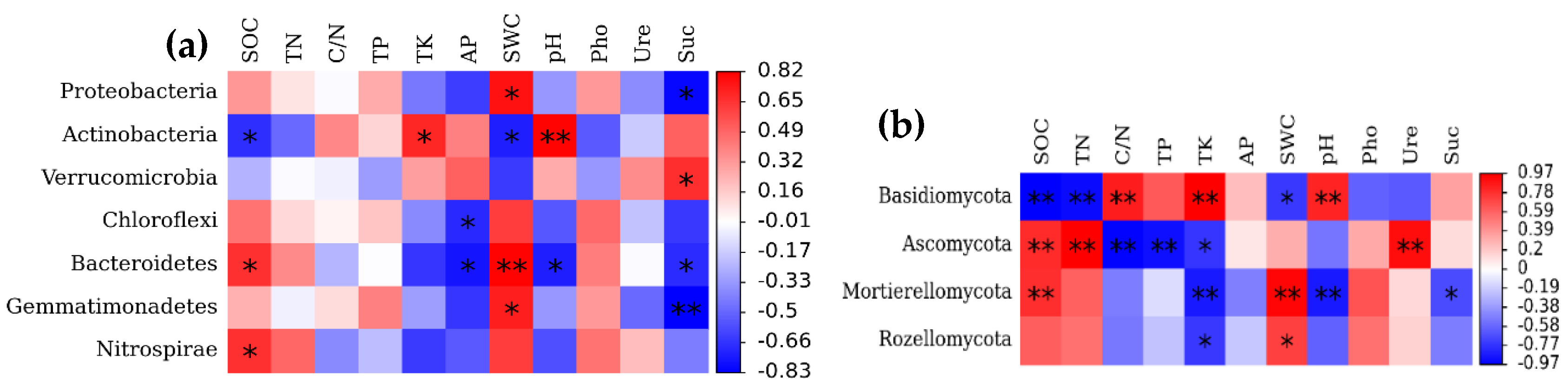
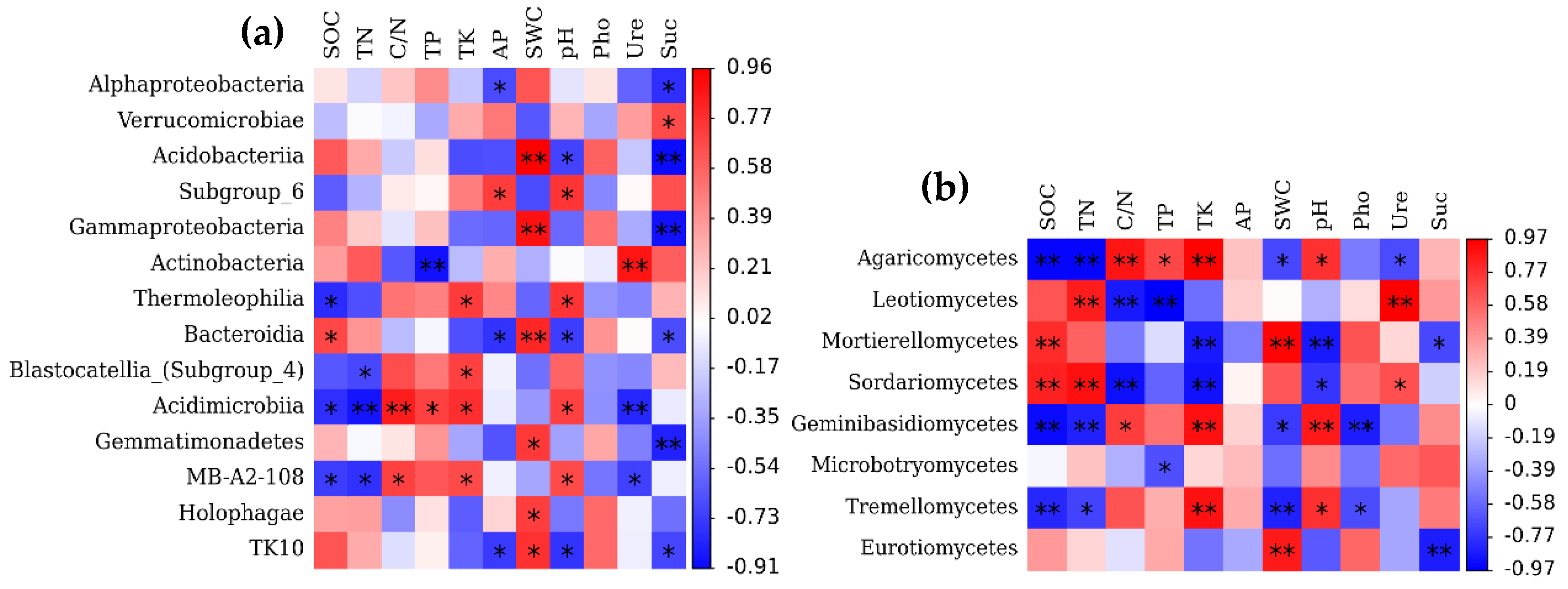
3.4. Effects of Soil Variables on the Makeup of the Bacterial and Fungal Community Composition
4. Discussion
4.1. The Variation in Soil Microbial Community Diversity in Forests across Successional Stages
4.2. The Variation of Soil Microbial Community Composition in Forests across Successional Stages
4.3. Effects of Soil Factors on Soil Microbial Community Composition in Forests across Successional Stages
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Kingdom. | Phylum | Class | Average Relative Abundance (%) | R2 | Slope ± SE | p-Value |
|---|---|---|---|---|---|---|
| Bacteria | Proteobacteria | Alphaproteobacteria | 20.03 | 0.273 | −0.809 ± 0.499 | 0.149 |
| Gammaproteobacteria | 6.81 | 0.689 | −1.333 ± 0.339 | 0.006 ** | ||
| Deltaproteobacteria | 5.10 | 0.087 | −0.186 ± 0.228 | 0.441 | ||
| Acidobacteria | Acidobacteriia | 8.02 | 0.868 | −2.741 ± 0.404 | <0.001 ** | |
| Subgroup_6 | 7.13 | 0.513 | 1.444 ± 0.532 | 0.03 * | ||
| Subgroup_4 | 2.29 | 0.320 | 0.323 ± 0.178 | 0.112 | ||
| Holophagae | 1.31 | 0.459 | −0.514 ± 0.211 | 0.045 * | ||
| Actinobacteria | Actinobacteria | 5.79 | 0.039 | 0.171 ± 0.321 | 0.611 | |
| Thermoleophilia | 5.19 | 0.443 | 0.569 ± 0.241 | 0.051 | ||
| Acidimicrobiia | 2.25 | 0.253 | 0.228 ± 0.148 | 0.167 | ||
| MB-A2-108 | 1.34 | 0.205 | 0.122 ± 0.091 | 0.221 | ||
| Bacteroidetes | Bacteroidia | 2.58 | 0.698 | −0.34 ± 0.085 | 0.005 ** | |
| Chloroflexi | KD4-96 | 3.00 | 0.003 | 0.03 ± 0.203 | 0.886 | |
| TK10 | 1.04 | 0.643 | −0.465 ± 0.131 | 0.009 ** | ||
| Rokubacteria | NC10 | 4.45 | 0.069 | −0.178 ± 0.248 | 0.496 | |
| Gemmatimonadetes | Gemmatimonadetes | 2.10 | 0.453 | −0.385 ± 0.160 | 0.047 * | |
| Verrucomicrobia | Verrucomicrobiae | 12.02 | 0.318 | 5.128 ± 2.838 | 0.114 | |
| Fungi | Ascomycota | Leotiomycetes | 17.12 | 0.015 | −0.006 ± 0.019 | 0.75 |
| Sordariomycetes | 6.90 | 0.500 | −0.138 ± 0.052 | 0.033 * | ||
| Eurotiomycetes | 1.90 | 0.674 | −0.284 ± 0.075 | 0.007 * | ||
| Dothideomycetes | 1.69 | 0.348 | −0.245 ± 0.127 | 0.095 | ||
| Mortierellomycota | Mortierellomycetes | 9.31 | 0.947 | −0.129 ± 0.012 | <0.001 ** | |
| Basidiomycota | Agaricomycetes | 33.50 | 0.593 | 0.022 ± 0.007 | 0.015 * | |
| Geminibasidiomycetes | 3.23 | 0.668 | 0.141 ± 0.038 | 0.007 ** | ||
| Microbotryomycetes | 2.26 | 0.267 | 0.214 ± 0.134 | 0.155 | ||
| Tremellomycetes | 2.15 | 0.771 | 0.507 ± 0.104 | 0.002 ** |


References
- Finegan, B. Forest succession (reply). Nature 1985, 315, 692. [Google Scholar] [CrossRef]
- Walker, L.R.; Walker, J.; Hobbs, R.J. Linking Restoration and Ecological Succession; Springer: Berlin, Germany, 2007. [Google Scholar]
- Zhang, C.; Liu, G.; Xue, S.; Wang, G. Soil bacterial community dynamics reflect changes in plant community and soil properties during the secondary succession of abandoned farmland in the Loess Plateau. Soil Biol. Biochem. 2016, 97, 40–49. [Google Scholar] [CrossRef]
- Zhao, C.; Long, J.; Liao, H.; Zheng, C.; Li, J.; Liu, L.; Zhang, M. Dynamics of soil microbial communities following vegetation succession in a karst mountain ecosystem, Southwest China. Sci. Rep. 2019, 9, 2160. [Google Scholar] [CrossRef] [PubMed]
- Bernard, L.; Mougel, C.; Maron, P.A.; Nowak, V.; Leveque, J.; Henault, C.; Haichar, F.Z.; Berge, O.; Marol, C.; Balesdent, J.; et al. Dynamics and identification of soil microbial populations actively assimilating carbon from 13C-labelled wheat residue as estimated by DNA- and RNA-SIP techniques. Environ. Microbiol. 2007, 9, 752–764. [Google Scholar] [CrossRef] [PubMed]
- Fang, X.; Yu, D.; Zhou, W.; Zhou, L.; Dai, L. The effects of forest type on soil microbial activity in Changbai Mountain, Northeast China. Ann. For. Sci. 2016, 73, 473–482. [Google Scholar] [CrossRef] [Green Version]
- Morrissey, E.M.; Mau, R.L.; Schwartz, E.; McHugh, T.A.; Dijkstra, P.; Koch, B.J.; Marks, J.C.; Hungate, B.A. Bacterial carbon use plasticity, phylogenetic diversity and the priming of soil organic matter. ISME J. 2017, 11, 1890–1899. [Google Scholar] [CrossRef]
- Fierer, N.; Bradford, M.A.; Jackson, R.B. Toward an ecological classification of soil bacteria. Ecology 2007, 88, 354–1364. [Google Scholar] [CrossRef]
- Zhou, Z.; Wang, C.; Jiang, L.; Luo, Y. Trends in soil microbial communities during secondary succession. Soil Biol. Biochem. 2017, 115, 92–99. [Google Scholar] [CrossRef]
- Yan, B.; Sun, L.; Li, J.; Liang, C.; Wei, F.; Xue, S.; Wang, G. Change in composition and potential functional genes of soil bacterial and fungal communities with secondary succession in Quercus liaotungensis forests of the Loess Plateau, western China. Geoderma 2020, 364, 114199. [Google Scholar] [CrossRef]
- Kielak, A.M.; Barreto, C.C.; Kowalchuk, G.A.; van Veen, J.A.; Kuramae, E.E. The Ecology of Acidobacteria: Moving beyond Genes and Genomes. Front. Microbiol. 2016, 7, 744. [Google Scholar] [CrossRef] [Green Version]
- Liu, G.; Chen, L.; Shi, X.; Yuan, Z.; Yuan, L.Y.; Lock, T.R.; Kallenbach, R.L. Changes in rhizosphere bacterial and fungal community composition with vegetation restoration in planted forests. Land Degrad. Dev. 2019, 30, 1147–1157. [Google Scholar] [CrossRef]
- Cai, Z.; Zhang, Y.; Yang, C.; Wang, S. Land-use type strongly shapes community composition, but not always diversity of soil microbes in tropical China. Catena 2018, 165, 369–380. [Google Scholar] [CrossRef]
- Jia, G.; Zhang, P.; Wang, G.; Cao, J.; Han, J.; Huang, Y. Relationship Between Microbial Community and Soil Properties During Natural Succession of Abandoned Agricultural Land.pdf. For. Ecol. Manag. 2010, 20, 352–360. [Google Scholar]
- Lin, Q.; Baldrian, P.; Li, L.; Novotny, V.; Hedenec, P.; Kukla, J.; Umari, R.; Meszarosova, L.; Frouz, J. Dynamics of Soil Bacterial and Fungal Communities During the Secondary Succession Following Swidden Agriculture IN Lowland Forests. Front. Microbiol. 2021, 12, 676251. [Google Scholar] [CrossRef]
- Zhang, X.; Liu, S.; Hong, Y.; Fu, S. Changes on Community Structure and Diversity of Soil Bacterial Community during the Succession of Quercus wutaishanica. Sci. Silvae Sin. 2019, 55, 193–202. [Google Scholar] [CrossRef]
- Nakayama, M.; Imamura, S.; Taniguchi, T.; Tateno, R. Does conversion from natural forest to plantation affect fungal and bacterial biodiversity, community structure, and co-occurrence networks in the organic horizon and mineral soil? For. Ecol. Manag. 2019, 446, 238–250. [Google Scholar] [CrossRef]
- Piao, Z.; Yang, L.; Zhao, L.; Yin, S. Actinobacterial community structure in soils receiving long-term organic and inorganic amendments. Appl. Environ. Microbiol. 2008, 74, 526–530. [Google Scholar] [CrossRef] [Green Version]
- Jiang, S.; Xing, Y.; Liu, G.; Hu, C.; Wang, X.; Yan, G.; Wang, Q. Changes in soil bacterial and fungal community composition and functional groups during the succession of boreal forests. Soil Biol. Biochem. 2021, 161, 108393. [Google Scholar] [CrossRef]
- Ren, C.; Liu, W.; Zhao, F.; Zhong, Z.; Deng, J.; Han, X.; Yang, G.; Feng, Y.; Ren, G. Soil bacterial and fungal diversity and compositions respond differently to forest development. Catena 2019, 181, 104071. [Google Scholar] [CrossRef]
- Zhong, Z.; Wang, X.; Zhang, X.; Zhang, W.; Xu, Y.; Ren, C.; Han, X.; Yang, G. Edaphic factors but not plant characteristics mainly alter soil microbial properties along a restoration chronosequence of Pinus tabulaeformis stands on Mt. Ziwuling, China. For. Ecol. Manag. 2019, 453, 117625. [Google Scholar] [CrossRef]
- Li, S.; Huang, X.; Shen, J.; Xu, F.; Su, J. Effects of plant diversity and soil properties on soil fungal community structure with secondary succession in the Pinus yunnanensis forest. Geoderma 2020, 379, 114646. [Google Scholar] [CrossRef]
- Wang, Z.; Bai, Y.; Hou, J.; Li, F.; Li, X.; Cao, R.; Deng, Y.; Wang, H.; Jiang, Y.; Yang, W. The Changes in Soil Microbial Communities across a Subalpine Forest Successional Series. Forests 2022, 13, 289. [Google Scholar] [CrossRef]
- Cline, L.C.; Zak, D.R. Soil microbial communities are shaped by plant-driven changes in resource availability during secondary succession. Ecology 2015, 96, 3374–3385. [Google Scholar] [CrossRef] [PubMed]
- Fierer, N.; Lauber, C.L.; Ramirez, K.S.; Zaneveld, J.; Bradford, M.A.; Knight, R. Comparative metagenomic, phylogenetic and physiological analyses of soil microbial communities across nitrogen gradients. ISME J. 2012, 6, 1007–1017. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, H.; Ye, D.; Wang, X.; Settles, M.L.; Wang, J.; Hao, Z.; Zhou, L.; Dong, P.; Jiang, Y.; Ma, Z. Soil bacterial communities of different natural forest types in Northeast China. Plant Soil 2014, 383, 203–216. [Google Scholar] [CrossRef]
- Lauber, C.L.; Hamady, M.; Knight, R.; Fierer, N. Pyrosequencing-based assessment of soil pH as a predictor of soil bacterial community structure at the continental scale. Appl. Environ. Microbiol. 2009, 75, 5111–5120. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thoms, C.; Gattinger, A.; Jacob, M.; Thomas, F.M.; Gleixner, G. Direct and indirect effects of tree diversity drive soil microbial diversity in temperate deciduous forest. Soil Biol. Biochem. 2010, 42, 1558–1565. [Google Scholar] [CrossRef]
- van der Heijden, M.G.; Bardgett, R.D.; van Straalen, N.M. The unseen majority: Soil microbes as drivers of plant diversity and productivity in terrestrial ecosystems. Ecol. Lett. 2008, 11, 296–310. [Google Scholar] [CrossRef]
- Sun, Y.; Wang, C.; Yang, J.; Liao, J.; Chen, H.Y.H.; Ruan, H.; Xu, X. Elevated CO2 shifts soil microbial communities from k- to r-strategists. Glob. Ecol. Biogeogr. 2021, 30, 961–972. [Google Scholar] [CrossRef]
- Jones, R.T.; Robeson, M.S.; Lauber, C.L.; Hamady, M.; Knight, R.; Fierer, N. A comprehensive survey of soil acidobacterial diversity using pyrosequencing and clone library analyses. ISME J. 2009, 3, 442–453. [Google Scholar] [CrossRef]
- Chen, Y.-L.; Xu, T.-L.; Veresoglou, S.D.; Hu, H.-W.; Hao, Z.-P.; Hu, Y.-J.; Liu, L.; Deng, Y.; Rillig, M.C.; Chen, B.-D. Plant diversity represents the prevalent determinant of soil fungal community structure across temperate grasslands in northern China. Soil Biol. Biochem. 2017, 110, 12–21. [Google Scholar] [CrossRef]
- Liu, C.; Xing, S.; Ma, H.; Du, Z.; MA, B. Effects of inoculating plant growth-promoting rhizobacteria on the biological characteristics of walnut (Juglans regia) rhizosphere soil under drought condition. Chin. J. Appl. Ecol. 2014, 25, 1475–1482. [Google Scholar] [CrossRef]
- Liao, H.; Huang, F.; Li, D.; Kang, L.; Chen, B.; Zhou, T.; Peng, S. Soil microbes regulate forest succession in a subtropical ecosystem in China: Evidence from a mesocosm experiment. Plant Soil 2018, 430, 277–289. [Google Scholar] [CrossRef]
- Liu, Y.; Zhang, W. Biodivers. Anal. Differ. Communities Broad-Leaved Korean Pine For. Xiaoxing’an Mountains. For. Eng. 2022, 38, 27–33. [Google Scholar] [CrossRef]
- Yang, X.; Lin, W.; Wu, Z. The species-abundance distribution pattern of broad-leaved Korean pine forest in the Lesser Khingan mountains. J. Cent. South Univ. For. Technol. 2020, 40, 104–113. [Google Scholar] [CrossRef]
- Shi, B.; Jin, G.; Wang, Z. Temporal and spatial variability in soil respiration in five temperate forests in Xiaoxing’an Mountains, China. Acta Ecol. Sin. 2012, 32, 5416–5428. [Google Scholar] [CrossRef]
- Gao, F.; Cui, X.; Sang, Y.; Song, J. Changes in soil organic carbon and total nitrogen as affected by primary forest conversion. For. Ecol. Manag. 2020, 463, 118013. [Google Scholar] [CrossRef]
- Bunnell, F.L.; Vales, D.J. Comparison of methods for estimating forest overstory cover: Differences among techniques. Can. J. For. Res. 1990, 20, 101–107. [Google Scholar] [CrossRef]
- Zhang, L.; Zhang, D.; Mao, Z. Charicateristic of Soil Oranic Carbon and Its Components in Different Successional Series of Broadleaved Korean Pine Forest in Xiaoxing’an Mountains. Sci. Silvae Sin. 2017, 53, 11–17. [Google Scholar]
- Zhang, W.; Qiao, W.; Gao, D.; Dai, Y.; Deng, J.; Yang, G.; Han, X.; Ren, G. Relationship between soil nutrient properties and biological activities along a restoration chronosequence of Pinus tabulaeformis plantation forests in the Ziwuling Mountains, China. Catena 2018, 161, 85–95. [Google Scholar] [CrossRef]
- Sparks, D.L.; Page, A.L.; Helmke, P.A.; Loeppert, R.H.; Soltanpour, P.N.; Tabatabai, M.A.; Johnston, C.T.; Sumner, M.E. Methods of Soil Analysis. Part. 3. Chemical Methods; John Wiley & Sons: Hoboken, NJ, USA, 1996. [Google Scholar] [CrossRef] [Green Version]
- Yao, H.; Huang, C. Soil Microbiology Ecology and Experimental Technology; China Science Publishing and Media Ltd.: Beijing, China, 2006. [Google Scholar]
- Bolyen, E.; Rideout, J.R.; Dillon, M.R.; Bokulich, N.A.; Abnet, C.; Al-Ghalith, G.A.; Alexander, H.; Alm, E.J.; Arumugam, M.; Asnicar, F.; et al. QIIME 2: Reproducible, Interactive, Scalable, and Extensible Microbiome Data Scienc. PeerJ Prepr. 2018, 6, e27295v2. [Google Scholar]
- Callahan, B.J.; McMurdie, P.J.; Rosen, M.J.; Han, A.W.; Johnson, A.J.; Holmes, S.P. DADA2: High-resolution sample inference from Illumina amplicon data. Nat. Methods 2016, 13, 581–583. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chao, A. Nonparametric estimation of the number of classes in a population. Scand. J. Stat. 1984, 4, 265–270. [Google Scholar]
- Shannon, P.; Markiel, A.; Ozier, O.; Baliga, N.S.; Wang, J.T.; Ramage, D.; Amin, N.; Schwikowski, B.; Ideker, T. Cytoscape: A software environment for integrated models of biomolecular interaction networks. Genome Res. 2003, 13, 2498–2504. [Google Scholar] [CrossRef] [PubMed]
- Simpson, E.H. Measurement of diversity. Nature 1949, 163, 688. [Google Scholar] [CrossRef]
- Segata, N.; Izard, J.; Waldron, L.; Biology, D.G.J.G. Metagenomic biomarker discovery and explanation. Genome Biol. 2011, 12, R60. [Google Scholar] [CrossRef] [Green Version]
- Team, R.C. R: A Language and Environment for Statistical Computing; Team, R.C: Vienna, Austria, 2013. [Google Scholar]
- Aanderud, Z.T.; Lennon, J.T. Validation of heavy-water stable isotope probing for the characterization of rapidly responding soil bacteria. Appl. Environ. Microbiol. 2011, 77, 4589–4596. [Google Scholar] [CrossRef] [Green Version]
- Siles, J.A.; Margesin, R. Abundance and Diversity of Bacterial, Archaeal, and Fungal Communities Along an Altitudinal Gradient in Alpine Forest Soils: What Are the Driving Factors? Microb. Ecol. 2016, 72, 207–220. [Google Scholar] [CrossRef] [Green Version]
- Shen, C.; Xiong, J.; Zhang, H.; Feng, Y.; Lin, X.; Li, X.; Liang, W.; Chu, H. Soil pH drives the spatial distribution of bacterial communities along elevation on Changbai Mountain. Soil Biol. Biochem. 2013, 57, 204–211. [Google Scholar] [CrossRef]
- Hooper, D.U.; Bignell, D.E.; Brown, V.K.; Brussard, L.; Dangerfield, J.M.; Wall, D.H.; Wardle, D.A.; Coleman, D.C.; Giller, K.E.; Lavelle, P.J.B. Interactions between Aboveground and Belowground Biodiversity in Terrestrial Ecosystems: Patterns, Mechanisms, and Feedbacks. Bioscience 2000, 50, 1049–1061. [Google Scholar] [CrossRef]
- Sterner, R.W.; Elser, J.J. Ecological Stoichiometry: The Biology of Elements from Molecules to the Biosphere; Princeton University Press: Princeton, NJ, USA, 2003. [Google Scholar] [CrossRef] [Green Version]
- Elser, J.J.; Sterner, R.W.; Gorokhova, E.; Fagan, W.F.; Markow, T.A.; Cotner, J.B.; Harrison, J.F.; Hobbie, S.E.; Odell, G.M.; Weider, L.W. Biological stoichiometry from genes to ecosystems. Ecol. Lett. 2008, 3, 540–550. [Google Scholar] [CrossRef]
- Su, Q. Growth rate hypothesis research progresses: Implications for zooplankton. Adv. Earth Sci. 2012, 27, 1204–1210. [Google Scholar]
- Yan, B.; Sun, L.; Li, J.; Gao, L.; Qiao, Z.; Gao, X.; Wu, C.; Wang, G. Changes of soil enzyme activity and their stoichiometric characteristics during secondary succession of Quercus liaotungensis forests. Chin. J. Ecol. 2021, 2021, 1–9. [Google Scholar] [CrossRef]
- Bai, X.; Shi, R.; You, Y.; Shen, H.; Han, S.; Zhang, Y. Bacterial community structure and diversity in soils of different forest ages and types in Baotianman forest, Henan Province, China. Chin. J. Appl. Ecol. 2015, 26, 2273–2281. [Google Scholar] [CrossRef]
- Chi, Y.; Liu, Z.; Bao, P. Decomposition and succession of wood by microflora on wood. J. Fungal Res. 2004, 2, 51–57. [Google Scholar] [CrossRef]
- Li, Q.; Wei, Y. Species composition and distribution characteristics of wood-decaying fungi in tropical areas of China. Res. Pap. 2021, 40, 2653–2672. [Google Scholar] [CrossRef]
- Urbanová, M.; Šnajdr, J.; Baldrian, P. Composition of fungal and bacterial communities in forest litter and soil is largely determined by dominant trees. Soil Biol. Biochem. 2015, 84, 53–64. [Google Scholar] [CrossRef]
- Chase, J.M. Community assembly: When should history matter? Oecologia 2003, 136, 489–498. [Google Scholar] [CrossRef]
- Padmanabhan, P.; Padmanabhan, S.; DeRito, C.; Gray, A.; Gannon, D.; Snape, J.R.; Tsai, C.S.; Park, W.; Jeon, C.; Madsen, E.L. Respiration of 13C-labeled substrates added to soil in the field and subsequent 16S rRNA gene analysis of 13C-labeled soil DNA. Appl. Env. Microbiol. 2003, 69, 1614–1622. [Google Scholar] [CrossRef] [Green Version]
- Liu, C.; Dong, Y.; Jiao, R. Research Progress in Acidobacteria Diversity in Forest Soil. World For. Res. 2016, 29, 17–22. [Google Scholar] [CrossRef]
- Barns, S.M.; Cain, E.C.; Sommerville, L.; Kuske, C.R. Acidobacteria phylum sequences in uranium-contaminated subsurface sediments greatly expand the known diversity within the phylum. Appl. Environ. Microbiol. 2007, 73, 3113–3116. [Google Scholar] [CrossRef] [Green Version]
- Staudt, K.; Serafimovich, A.; Siebicke, L.; Pyles, R.D.; Falge, E. Vertical structure of evapotranspiration at a forest site (a case study). Agric. For. Meteorol. 2011, 151, 709–729. [Google Scholar] [CrossRef]
- Liu, Z.; Wang, Y.; Tian, A.; Yu, P.; Xiong, W.; Xu, L.; Wang, Y. Intra-Annual Variation of Stem Radius of Larix principis-rupprechtii and Its Response to Environmental Factors in Liupan Mountains of Northwest China. Forests 2017, 8, 382. [Google Scholar] [CrossRef] [Green Version]
- Caldwell, B.A. Enzyme activities as a component of soil biodiversity: A review. Pedobiologia 2005, 49, 637–644. [Google Scholar] [CrossRef]






| Forest Stand Features | Forest Successional Stages | ||
|---|---|---|---|
| Early-Stage | Intermediate-Stage | Late-Stage | |
| Tree composition * | 60% Betula platyphylla, 20% Populus davidiana, 10% Abies nephlolepis, 10% Ulmus davidiana and others | 40% Fraxinus mandshurica, 20% Ulmus davidiana, 20% Populus davidiana,10% Quercus mongolica, 10% Acer mono and others | 60% Pinus koraiensis, 20% Fraxinus mandshurica, 10% Abies nephlolepis,10% Tilia amurensis and others |
| Mean height (m) | 22.03 | 20.57 | 24.50 |
| Mean DBH (cm) | 20.68 | 22.76 | 33.90 |
| Canopy closure | 0.8 | 0.8 | 0.8 |
| Forest type | Secondary birch forest | Selection cutting forest | Mixed broad-leaved Korean pine forest |
| Age of stand | <60 | 80–120 | >300 |
| Soil Properties Index | Forest Successional Stages | ||
|---|---|---|---|
| Early-Stage | Intermediate-Stage | Late-Stage | |
| SOC(g/kg) | 80.28 ± 2.52 a | 81.94 ± 1.17 a | 60.28 ± 0.74 b |
| TN(g/kg) | 4.93 ± 0.075 b | 6.14 ± 0.07 a | 3.13 ± 0.07 c |
| C/N | 16.31 ± 0.70 b | 13.35 ± 0.14 c | 19.27 ± 0.56 a |
| TP(g/kg) | 0.83±0.02 a | 0.64±0.03 b | 0.87±0.01 a |
| TK(g/kg) | 8.32 ± 0.38 b | 8.53 ± 0.25 b | 13.08 ± 0.12 a |
| AP(mg/kg) | 4.22 ± 0.37 a | 4.79 ± 0.16 a | 4.76 ± 0.16 a |
| pH | 5.10 ± 0.09 b | 5.29 ± 0.09 b | 5.82 ± 0.13 a |
| SWC(%) | 39.43 ± 0.70 a | 29.30 ± 0.67 b | 22.63 ± 0.38 c |
| Pho(mg/g 24 h) | 3.03 ± 0.03 a | 2.84 ± 0.04 ab | 2.53 ± 0.23 b |
| Ure(mg/g 24 h) | 2.42 ± 0.23 b | 8.54 ± 0.23 a | 1.69 ± 0.03 c |
| Suc(mg/g 24 h) | 2.75 ± 0.71 b | 8.45 ± 9.67 a | 8.06 ± 2.06 a |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhu, K.; Wang, Q.; Zhang, Y.; Zarif, N.; Ma, S.; Xu, L. Variation in Soil Bacterial and Fungal Community Composition at Different Successional Stages of a Broad-Leaved Korean Pine Forest in the Lesser Hinggan Mountains. Forests 2022, 13, 625. https://0-doi-org.brum.beds.ac.uk/10.3390/f13040625
Zhu K, Wang Q, Zhang Y, Zarif N, Ma S, Xu L. Variation in Soil Bacterial and Fungal Community Composition at Different Successional Stages of a Broad-Leaved Korean Pine Forest in the Lesser Hinggan Mountains. Forests. 2022; 13(4):625. https://0-doi-org.brum.beds.ac.uk/10.3390/f13040625
Chicago/Turabian StyleZhu, Kaiyue, Qingcheng Wang, Yong Zhang, Nowsherwan Zarif, Shuangjiao Ma, and Liqing Xu. 2022. "Variation in Soil Bacterial and Fungal Community Composition at Different Successional Stages of a Broad-Leaved Korean Pine Forest in the Lesser Hinggan Mountains" Forests 13, no. 4: 625. https://0-doi-org.brum.beds.ac.uk/10.3390/f13040625