
Parsing Long-Term Tree Recruitment, Growth, and Mortality to Identify Hurricane Effects on Structural and Compositional Change in a Tropical Forest



Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Site
2.2. Diameter and Species Observations
2.3. Plant Functional Types (PFT) of Species
2.4. Data Analyses
3. Results
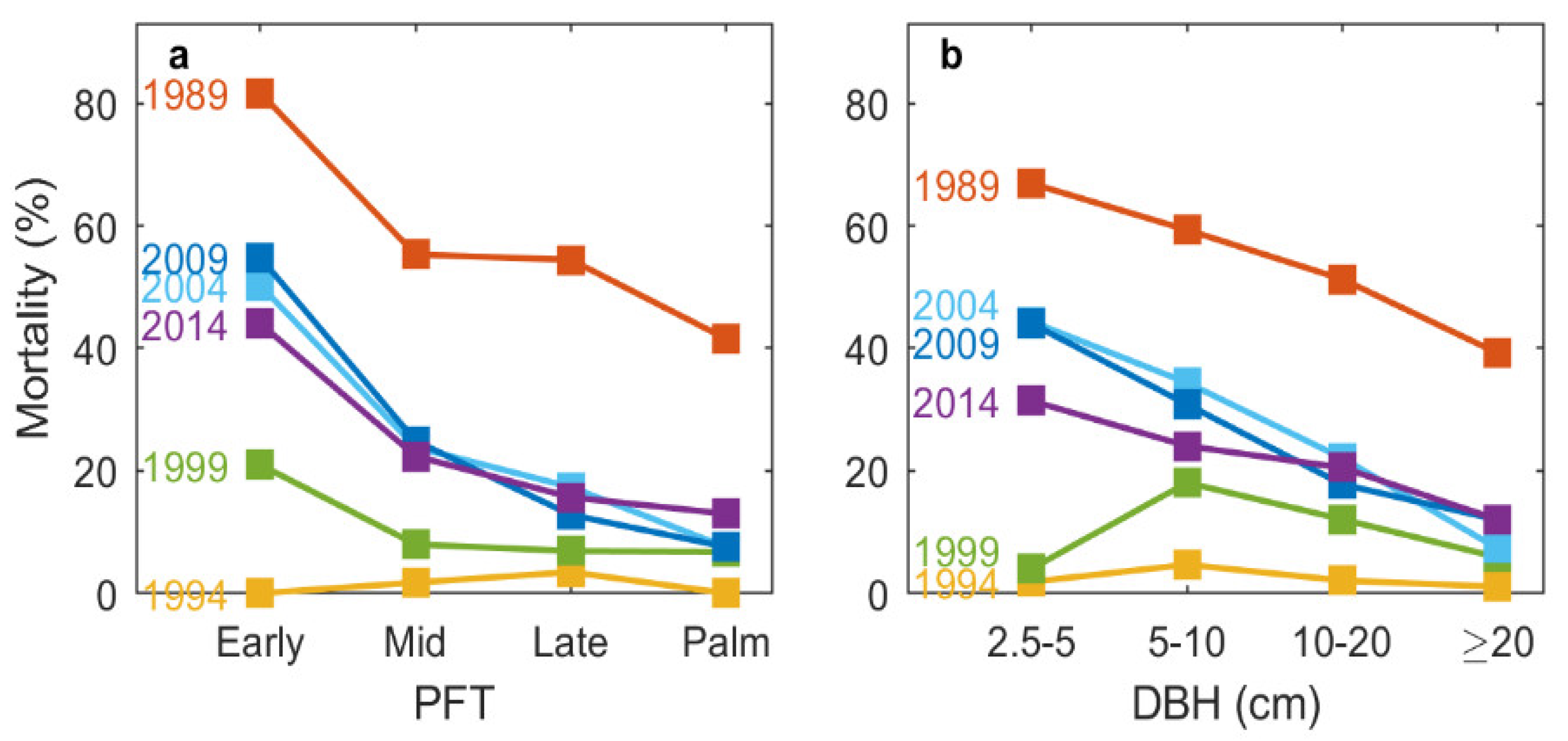
3.1. Mortality and Recruitment over the Course of 25 Years
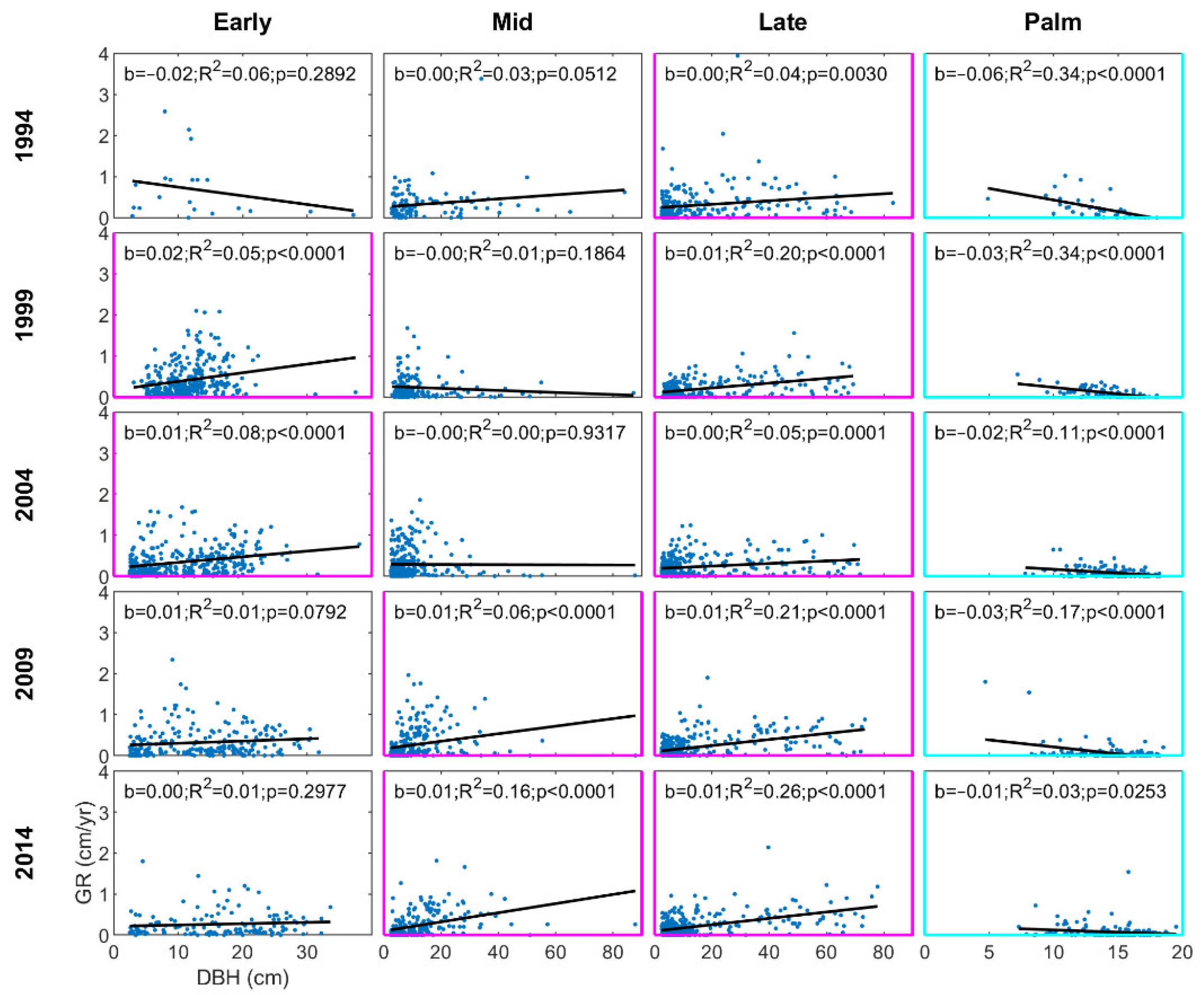
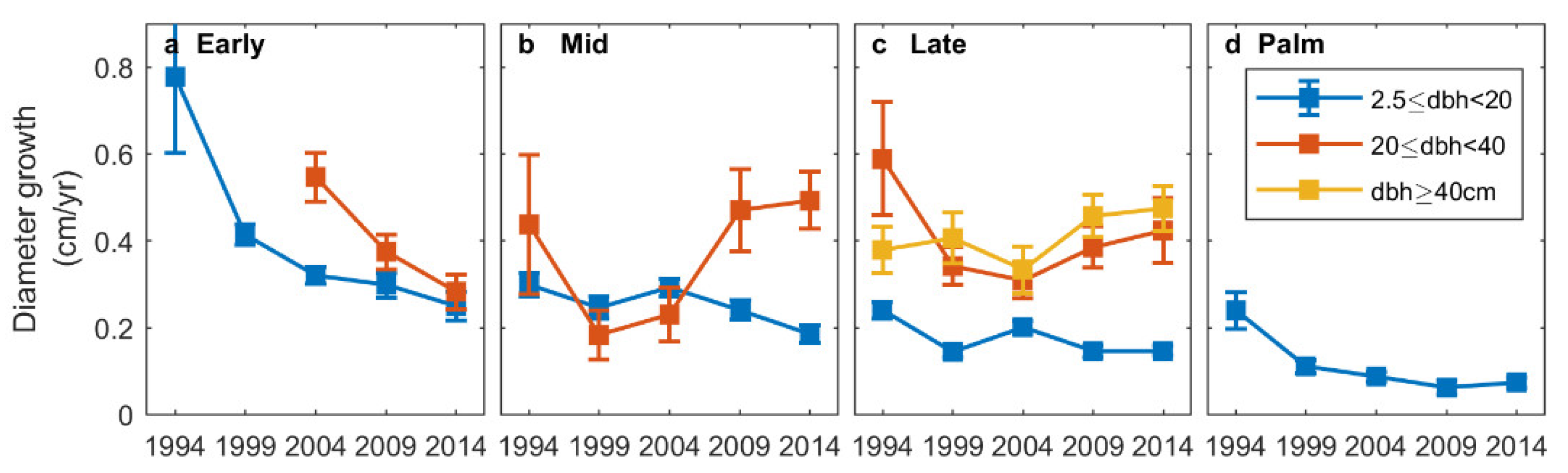
3.2. Diameter Growth Rates
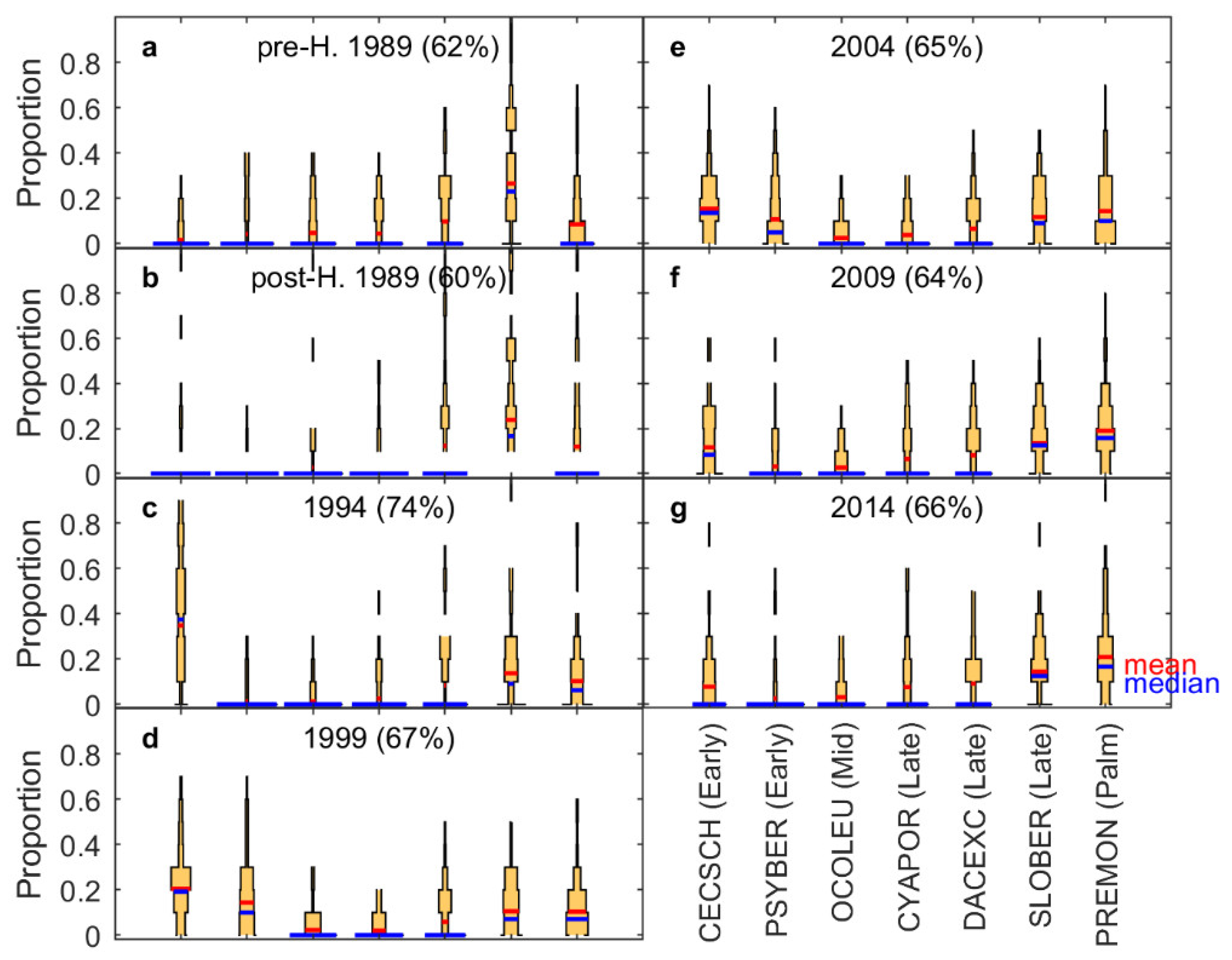
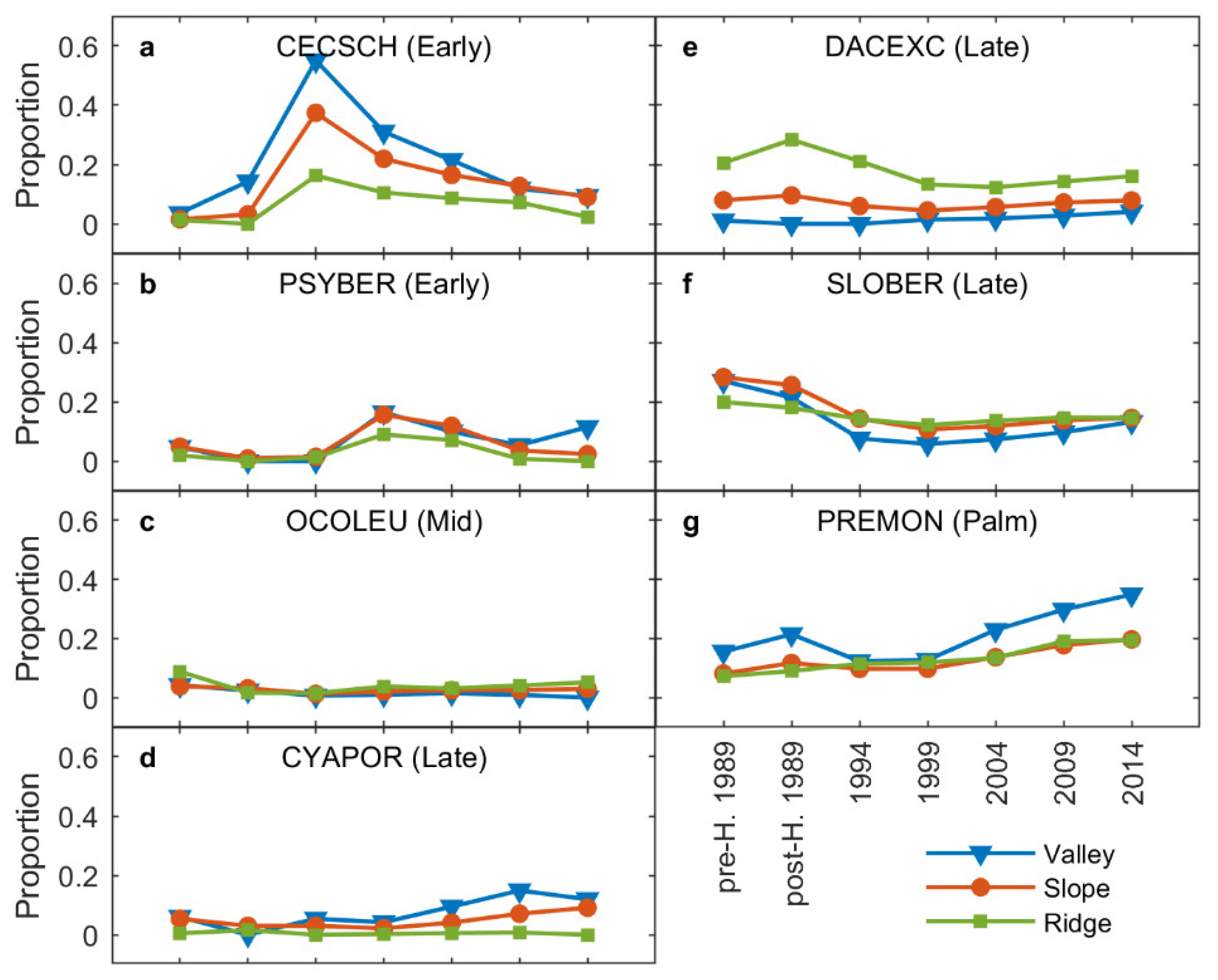
3.3. Shifts in Species Composition
3.4. Impact of Topography
4. Discussion
4.1. Patterns of Forest Structure following a Hurricane Disturbance
4.2. Composition Dynamics of Dominant Species and Plant Functional Types
4.3. Concluding Remarks
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Lugo, A.E. Visible and invisible effects of hurricanes on forest ecosystems: An international review. Austral Ecol. 2008, 33, 368–398. [Google Scholar] [CrossRef]
- Flynn, D.F.; Uriarte, M.; Crk, T.; Pascarella, J.B.; Zimmerman, J.K.; Aide, T.M.; Ortiz, M.A.C. Hurricane disturbance alters secondary forest recovery in Puerto Rico. Biotropica 2010, 42, 149–157. [Google Scholar] [CrossRef]
- Shiels, A.B.; Gonzalez, G.; Lodge, D.L.; Willing, M.R.; Zimmerman, J.K. Cascading effects of canopy opening and debris deposition from a large-scale hurricane experiment in a tropical rain forest. BioScience 2015, 65, 871–881. [Google Scholar] [CrossRef] [Green Version]
- Uriarte, M.; Thompson, J.; Zimmerman, J.K. Hurricane Maria tripled stem breaks and doubled tree mortality relative to other major storms. Nat. Commun. 2019, 10, 1362. [Google Scholar] [CrossRef] [Green Version]
- Hogan, J.A.; A Feagin, R.; Starr, G.; Ross, M.; Lin, T.-C.; O’Connell, C.; Huff, T.P.; A Stauffer, B.; Robinson, K.L.; Lara, M.C.; et al. A research framework to integrate cross-ecosystem responses to tropical cyclones. BioScience 2020, 70, 477–489. [Google Scholar] [CrossRef]
- Lin, T.C.; Hogan, J.A.; Chang, C.T. Tropical cyclone ecology: A scale-link perspective. Trends Ecol. Evol. 2020, 35, 594–604. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hall, J.; Muscarella, R.; Quebbeman, A.; Arellano, G.; Thompson, J.; Zimmerman, J.K.; Uriarte, M. Hurricane-induced rainfall is a stronger predictor of tropical forest damage in Puerto Rico than maximum wind speeds. Sci. Rep. 2020, 10, 4318. [Google Scholar] [CrossRef]
- Everham, M.E., III; Brokaw, N.V.L. Forest damage and recovery from catastrophic wind. Bot. Rev. 1996, 62, 113–185. [Google Scholar] [CrossRef]
- Frangi, J.L.; Lugo, A.E. Hurricane damage to a flood plain forest in the Luquillo Mountains of Puerto Rico. Biotropica 1991, 23, 324–335. [Google Scholar] [CrossRef]
- Walker, L.R. Tree damage and recovery from hurricane Hugo in Luquillo Experimental Forest, Puerto Rico. Part A. special issue: Ecosystem, plant, and animal responses to hurricanes in the Caribbean. Biotropica 1991, 23, 379–385. [Google Scholar] [CrossRef]
- Pascarella, J.B.; Aide, T.M.; Zimmerman, J.K. Short-term response of secondary forests to hurricane disturbance in Puerto Rico, USA. For. Ecol. Manag. 2004, 199, 379–393. [Google Scholar] [CrossRef]
- Bellingham, P.J.; Tanner, E.V.J. The influence of topography on tree growth, mortality, and recruitment in a tropical montane forest. Biotropica 2000, 32, 378–384. [Google Scholar] [CrossRef]
- Feeley, K.J.; Davies, S.J.; Perez, R.; Hubbell, S.P.; Foster, R.B. Directional changes in the species composition of a tropical forest. Ecology 2011, 92, 871–882. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Heartsill Scalley, T.; Scatena, F.N.; Lugo, A.E.; Moya, S.; Estrada, C.R. Changes in structure, composition, and nutrients during 15 years of hurricane-induced succession in a subtropical wet forest in Puerto Rico. Biotropica 2010, 42, 455–463. [Google Scholar] [CrossRef]
- Tanner, E.V.J.; Rodriguez-Sanchez, F.; Healey, J.R.; Holdaway, R.J.; Bellingham, P.J. Long-term hurricane damage effects on tropical forest tree growth and mortality. Ecology 2014, 95, 2974–2983. [Google Scholar] [CrossRef] [Green Version]
- Weaver, P.L. Forest changes after hurricanes in Puerto Rico Luquillo Mountains. Interciencia 1989, 14, 181–192. [Google Scholar]
- Burslem, D.F.; Whitmore, T.C.; Brown, G.C. Short-term effects of cyclone impact and long-term recovery of tropical rain forest on Kolombangara, Solomon Islands. J. Ecol. 2000, 88, 1063–1078. [Google Scholar] [CrossRef]
- Vandermeer, J.; de la Cerda, I.G. Height dynamics of the thinning canopy of a tropical rain forest: 14 years of succession in a post-hurricane forest in Nicaragua. For. Ecol. Manag. 2004, 199, 125–135. [Google Scholar] [CrossRef]
- Bonan, G. “[Plant Strategies]” in Ecological Climatology: Concepts and Applications, 3rd ed.; Cambridge University Press: New York, NY, USA, 2016; pp. 291–314. [Google Scholar]
- Zhang, J.; Heartsill-Scalley, T.; Bras, R.L. Forest Structure and Composition Are Critical to Hurricane Mortality. Forests 2022, 13, 202. [Google Scholar] [CrossRef]
- Bender, M.A.; Knutson, T.R.; Tuleya, R.E.; Sirutis, J.J.; Vecchi, G.A.; Garner, S.T.; Held, I.M. Modeled impact of anthropogenic warming on the frequency of intense Atlantic hurricanes. Science 2010, 327, 454–458. [Google Scholar] [CrossRef] [Green Version]
- Wang, S.; Toumi, R. More tropical cyclones are striking coasts with major intensities at landfall. Sci. Rep. 2022, 12, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Rozendaal, D.M.A.; Chazdon, R.L. Demographic drivers of tree biomass change during secondary succession in northeastern Costa Rica. Ecol. Appl. 2015, 25, 506–516. [Google Scholar] [CrossRef] [PubMed]
- Beard, J.S. The Natural Vegetation of the Windward and Leeward Islands; Oxford Forestry Memoirs; 21 Clarendon Press: Oxford, UK, 1949; p. 192. [Google Scholar]
- Gregory, F.A.; Sabat, A.M. The effect of hurricane disturbance on the fecundity of Sierra palms (Pretoea montana). Bios 1996, 67, 135–139. [Google Scholar]
- Ma, R.-Y.; Zhang, J.-L.; Cavaleri, M.A.; Sterck, F.; Strijk, J.S.; Cao, K.-F. Convergent evolution towards high net carbon gain efficiency contributes to the shade tolerance of palms (Arecaceae). PLoS ONE 2015, 10, e0140384. [Google Scholar] [CrossRef] [Green Version]
- Muscarella, R.; Emilio, T.; Phillips, O.L.; Lewis, S.L.; Slik, F.; Baker, W.J.; Couvreur, T.L.P.; Eiserhardt, W.L.; Svenning, J.C.; Affum-Baffoe, K.; et al. The global abundance of tree palms. Glob. Ecol. Biogeogr. 2020, 29, 1495–1514. [Google Scholar] [CrossRef]
- McDowell, W.H.; Leon, M.C.; Shattuck, M.D.; Potter, J.D.; Heartsill-Scalley, T.; González, G.; Shanley, J.B.; Wymore, A.S. Luquillo Experimental Forest: Catchment science in the montane tropics. Hydrol. Process. 2021, 35, e14146. [Google Scholar] [CrossRef]
- Scatena, F.N.; Silver, W.; Siccama, T.; Johnson, A.; Sanchez, M.J. Biomass and nutrient content of the Bisley Experimental Watersheds, Luquillo Experimental Forest, Puerto Rico, before and after hurricane Hugo, 1989. Biotropica 1993, 25, 15–27. [Google Scholar] [CrossRef]
- Heartsill Scalley, T.; Scatena, F.N.; Estrada, C.; McDowell, W.H.; Lugo, A.E. Disturbance and long-term patterns of rainfall and throughfall nutrient fluxes in a subtropical forest in Puerto Rico. J. Hydrol. 2007, 33, 472–485. [Google Scholar] [CrossRef]
- DeWalt, S.J.; Ickes, K.; James, A. Forest and community structure of tropical sub-montane rain forests on the island of Dominica, Lesser Antilles. Caribb. Nat. 2016, 1, 116–137. [Google Scholar]
- Scatena, F.N.; Lugo, A.E. Geomorphology, disturbance, and the soil and vegetation of two subtropical wet steepland watersheds of Puerto Rico. Geomorphology 1995, 13, 199–213. [Google Scholar] [CrossRef]
- Scatena, F.N. An introduction to the physiography and history of the Bisley Experimental Watersheds in the Luquillo Mountains of Puerto Rico. In General Technical Report SO-72; U.S. Department of Agriculture, Forest Service, Southern Forest Experiment Station: New Orleans, LA, USA, 1989; 22p. [Google Scholar] [CrossRef]
- García-Montiel, D.C.; Scatena, F.N. The effect of human activity on the structure and composition of a tropical forest in Puerto Rico. For. Ecol. Manag. 1994, 63, 57–78. [Google Scholar] [CrossRef]
- Royo, A.A.; Heartsill-Scalley, T.; Moya, S.; Scatena, F.N. Non-arborescent vegetation trajectories following repeated hurricane disturbance: Ephemeral versus enduring responses. Ecosphere 2011, 2, 77. [Google Scholar] [CrossRef] [Green Version]
- Lugo, A.E.; Helmer, E. Emerging forests on abandoned land: Puerto Rico’s new forests. For. Ecol. Manag. 2004, 190, 145–161. [Google Scholar] [CrossRef] [Green Version]
- Heartsill Scalley, T.; Scatena, F.N.; Moya, S.; Lugo, A.E. Long-term dynamics of organic matter and elements exported as coarse particulates from two Caribbean montane watersheds. J. Trop. Ecol. 2012, 28, 127–139. [Google Scholar] [CrossRef]
- Ostertag, R.; Silver, W.L.; Lugo, A.E. Factors affecting mortality and resistance to damage following hurricanes in a rehabilitated subtropical moist forest. Biotropica 2005, 37, 16–24. [Google Scholar] [CrossRef]
- Scatena, F.N.; Moya, S.; Estrada, C.; Chinea, J.D. The first five years in the reorganization of aboveground biomass and nutrient use following hurricane Hugo in the Bisley Experimental Watersheds, Luquillo Experimental Forest, Puerto Rico. Biotropica 1996, 28, 424–440. [Google Scholar] [CrossRef]
- Scalley, T.H. Insights on forest structure and composition from long-term research in the Luquillo mountains. Forests 2017, 8, 204. [Google Scholar] [CrossRef] [Green Version]
- Schowalter, T.D.; Ganio, L.M. Invertebrate communities in a tropical rain forest canopy in Puerto Rico following hurricane Hugo. Ecol. Entomol. 1999, 24, 191–201. [Google Scholar] [CrossRef]
- Uriarte, M.; Canham, C.D.; Thompson, J.; Zimmerman, J.K.; Brokaw, N. Seedling recruitment in a hurricane-driven tropical forest: Light limitation, density-dependence and the spatial distribution of parent trees. J. Ecol. 2005, 93, 291–304. [Google Scholar] [CrossRef]
- Muscarella, R.; Uriarte, M.; Forero-Montaña, J.; Comita, L.; Swenson, N.G.; Thompson, J.; Nytch, C.J.; Jonckheere, I.; Zimmerman, J.K. Life-history trade-offs during the seed-to-seedling transition in a subtropical wet forest community. J. Ecol. 2013, 101, 171–182. [Google Scholar] [CrossRef]
- Feng, X.; Uriarte, M.; González, G.; Reed, S.; Thompson, J.; Zimmerman, J.K.; Murphy, L. Improving predictions of tropical forest response to climate change through integration of field studies and ecosystem modeling. Glob. Chang. Biol. 2018, 24, e213–e232. [Google Scholar] [CrossRef] [PubMed]
- Köhler, P.; Ditzer, T.; Huth, A. Concepts for the aggregation of tropical tree species into functional types and the application to Sabah’s lowland rain forests. J. Trop. Ecol. 2000, 16, 591–602. [Google Scholar] [CrossRef]
- Duckworth, J.C.; Kent, M.; Ramsay, P.M. Plant functional types: An alternative to taxonomic plant community description in biogeography? Prog. Phys. Geogr. 2000, 24, 515–542. [Google Scholar] [CrossRef]
- Semenova, G.V.; van der Maarel, E. Plant functional types–a strategic perspective. J. Veg. Sci. 2000, 11, 917–922. [Google Scholar] [CrossRef]
- Verheijen, L.M.; Aerts, R.; Bönisch, G.; Kattge, J.; Van Bodegom, P.M. Variation in trait trade-offs allows differentiation among predefined plant functional types: Implications for predictive ecology. New Phytol. 2016, 209, 563–575. [Google Scholar] [CrossRef] [PubMed]
- Wigley, B.J.; Charles-Dominique, T.; Hempson, G.P.; Stevens, N.; te Beest, M.; Archibald, S.; Bond, W.J.; Bunney, K.; Coetsee, C.; Donaldson, J.; et al. A handbook for the standardised sampling of plant functional traits in disturbance-prone ecosystems, with a focus on open ecosystems. Aust. J. Bot. 2020, 68, 473–531. [Google Scholar] [CrossRef]
- Devoe, N.N. Differential Seeding and Regeneration in Openings and Beneath Closed Canopy in Sub-Tropical Wet Forest. Ph.D. Dissertation, Yale University, New Haven, CT, USA, 1989; 307p. [Google Scholar]
- Lugo, A.E.; Zimmerman, J.K. Ecological life histories. In Tropical Tree Seed Manual. Agriculture Handbook; Vozzo, J.A., Ed.; USDA Forest Service: Washington, DC, USA, 2002; pp. 191–213. [Google Scholar]
- Kunstler, G.; Falster, D.; Coomes, D.A.; Hui, F.; Kooyman, R.M.; Laughlin, D.C.; Poorter, L.; Vanderwel, M.; Vieilledent, G.; Wright, S.J.; et al. Plant functional traits have globally consistent effects on competition. Nature 2016, 529, 204–207. [Google Scholar] [CrossRef] [Green Version]
- Lugo, A.E.; Batlle, C.T.R. Leaf production, growth rate, and age of the palm Prestoea montana in the Luquillo Experimental Forest, Puerto Rico. J. Trop. Ecol. 1987, 3, 151–161. [Google Scholar] [CrossRef]
- Lugo, A.E.; Francis, J.K.; Frangi, J.L. Prestoea montana (R. Graham) Nichols. Sierra palm. Palmaceae Palm Family; Technical Report SO-ITF-SM-82; US Department of Agriculture, Forest Service, International Institute of Tropical Forestry: Rio Piedras, Puerto Rico, USA, 1998.
- Montgomery, D.C.; Runger, G.C. Large-sample tests on the difference in population proportions. In Applied Statistics and Probability for Engineers, 7th ed.; Wiley: New York, NY, USA, 2018; pp. 274–275. ISBN 978-1-119-40036-3. [Google Scholar]
- Crow, T.R. A rainforest chronicle: A 30-year record of change in structure and composition at El Verde, Puerto Rico. Biotropica 1980, 12, 42–55. [Google Scholar] [CrossRef]
- Zimmerman, J.K.; Everham, E.M., III; Waide, R.B.; Lodge, D.J.; Taylor, C.M.; Brokaw, N.V.L. Responses of tree species to hurricane winds in subtropical wet forest in Puerto Rico: Implications for tropical tree life histories. J. Ecol. 1994, 82, 911–922. [Google Scholar] [CrossRef]
- Condit, R.; Hubbell, S.P.; Lafrankie, J.V.; Sukumar, R.; Manokaran, N.; Foster, R.B.; Ashton, P.S. Species-area and species-individual relationships for tropical trees: A comparison of three 50-ha plots. J. Ecol. 1996, 84, 549–562. [Google Scholar] [CrossRef] [Green Version]
- Basnet, K.; Scatena, F.N.; Likens, G.E.; Lugo, A.E. Ecological consequences of root grafting in tabonuco (Dacryodes excelsa) trees in the Luquillo Experimental Forest, Puerto Rico. Biotropica 1993, 25, 28–35. [Google Scholar] [CrossRef]
- Yih, K.; Boucher, D.H.; Vandermeer, J.H.; Zamora, N. Recovery of the rain forest of southeastern Nicaragua after destruction by hurricane Joan. Biotropica 1991, 23, 106–113. [Google Scholar] [CrossRef]
- Lugo, A.E.; Frangi, J.L. Long-term response of Caribbean palm forests to hurricanes. Caribb. Nat. 2016, 1, 157–175. [Google Scholar]
- Lohbeck, M.; Poorter, L.; Martínez-Ramos, M.; Rodriguez-Velázquez, J.; van Breugel, M.; Bongers, F. Changing drivers of species dominance during tropical forest succession. Funct. Ecol. 2014, 28, 1052–1058. [Google Scholar] [CrossRef]
- Jucker, T.; Caspersen, J.; Chave, J.; Antin, C.; Barbier, N.; Bongers, F.; Dalponte, M.; van Ewijk, K.Y.; Forrester, D.I.; Haeni, M.; et al. Allometric equations for integrating remote sensing imagery into forest monitoring programmes. Glob. Chang. Biol. 2017, 23, 177–190. [Google Scholar] [CrossRef] [Green Version]
- Fortunel, C.; Lasky, J.R.; Uriarte, M.; Valencia, R.; Wright, S.J.; Garwood, N.C.; Kraft, N.J.B. Topography and neighborhood crowding can interact to shape species growth and distribution in a diverse Amazonian forest. Ecology 2018, 99, 2272–2283. [Google Scholar] [CrossRef]
- Muscarella, R.; Kolyaie, S.; Morton, D.C.; Zimmerman, J.K.; Uriarte, M. Effects of topography on tropical forest structure depend on climate context. J. Ecol. 2020, 108, 145–159. [Google Scholar] [CrossRef]
- Zambrano, J.; Arellano, G.; Swenson, N.G.; Staniczenko, P.P.; Thompson, J.; Fagan, W.F. Analyses of three-dimensional species associations reveal departures from neutrality in a tropical forest. Ecology 2022, e3681. [Google Scholar] [CrossRef]
- Lugo, A.E.; Scatena, F.N. Background and catastrophic tree mortality in tropical moist, wet, and rain forests. Biotropica 1996, 28, 585–599. [Google Scholar] [CrossRef]
- Baroni, T.J.; State University of New York College at Cortland, Cortland, New York, NY, USA; Larsson, K.-H.; University of Oslo, Natural History Museum, Oslo, Norway; Lodge, D.J.; USDA Forest Service, Luquillo, Puerto Rico, FL, USA. Personal communication, 2022.
- Xi, W. Synergistic effects of tropical cyclones on forest ecosystems: A global synthesis. J. For. Res. 2015, 26, 1–21. [Google Scholar] [CrossRef]
- Harris, N.L.; Gibbs, D.A.; Baccini, A.; Birdsey, R.A.; de Bruin, S.; Farina, M.; Fatoyinbo, L.; Hansen, M.C.; Herold, M.; Houghton, R.A.; et al. Global maps of twenty-first century forest carbon fluxes. Nat. Clim. Chang. 2021, 11, 234–240. [Google Scholar] [CrossRef]
- Chave, J.; Andalo, C.; Brown, S.; Cairns, M.A.; Chambers, J.Q.; Eamus, D.; Fölster, H.; Fromard, F.; Higuchi, N.; Kira, T.; et al. Tree allometry and improved estimation of carbon stocks and balance in tropical forests. Oecologia 2005, 145, 87–99. [Google Scholar] [CrossRef] [PubMed]
- Keller, M.; Palace, M.; Hurtt, G. Biomass estimation in the Tapajos National Forest, Brazil: Examination of sampling and allometric uncertainties. For. Ecol. Manag. 2001, 154, 371–382. [Google Scholar] [CrossRef]
- Baccini, A.; Walker, W.; Carvalho, L.; Farina, M.; Sulla-Menashe, D.; Houghton, R.A. Tropical forests are a net carbon source based on aboveground measurements of gain and loss. Science 2017, 358, 230–234. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Claeys, F.; Gourlet-Fleury, S.; Picard, N.; Ouédraogo, D.-Y.; Tadesse, M.G.; Hérault, B.; Baya, F.; Bénédet, F.; Cornu, G.; Mortier, F. Climate change would lead to a sharp acceleration of Central African forests dynamics by the end of the century. Environ. Res. Lett. 2019, 14, 044002. [Google Scholar] [CrossRef]
- McDowell, N.G.; Allen, C.D.; Anderson-Teixeira, K.; Aukema, B.H.; Bond-Lamberty, B.; Chini, L.; Clark, J.S.; Dietze, M.; Grossiord, C.; Hanbury-Brown, A.; et al. Pervasive shifts in forest dynamics in a changing world. Science 2020, 29, eaaz9463. [Google Scholar] [CrossRef]
- Luke, D.; McLaren, K.; Wilson, B. Modeling hurricane exposure in a Caribbean lower montane tropical wet forest: The effects of frequent, intermediate disturbances and topography on forest structural dynamics and composition. Ecosystems 2016, 19, 1178–1195. [Google Scholar] [CrossRef]
- Yap, S.L.; Davies, S.J.; Condit, R. Dynamic response of a Philippine dipterocarp forest to typhoon disturbance. J. Veg. Sci. 2016, 27, 133–143. [Google Scholar] [CrossRef]
- McLaren, K.; Luke, D.; Tanner, E.; Bellingham, P.J.; Healey, J.R. Reconstructing the effects of hurricanes over 155 years on the structure and diversity of trees in two tropical montane rainforests in Jamaica. Agric. For. Meteorol. 2019, 276, 107621. [Google Scholar] [CrossRef]
- Michener, W.K.; Jones, M.B. Ecoinformatics: supporting ecology as a data-intensive science. Trends Ecol. Evol. 2012, 27, 85–93. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Michener, W.K.; Porter, J.; Servilla, M.; Vanderbilt, K. Long term ecological research and information management. Ecol. Inform. 2011, 6, 13–24. [Google Scholar] [CrossRef]
- Condit, R.; Ashton, P.; Bunyavejchewin, S.; Dattaraja, H.S.; Davies, S.; Esufali, S.; Ewango, C.; Foster, R.; Gunatilleke, I.A.U.N.; Gunatilleke, C.V.S.; et al. The importance of demographic niches to tree diversity. Science 2006, 313, 98–101. [Google Scholar] [CrossRef] [Green Version]
- Clark, J.S.; Wolosin, M.; Dietze, M.; Ibanez, I.; LaDeau, S.; Welsh, M.; Kloeppel, B. Tree growth inference and prediction from diameter censuses and ring widths. Ecol. Appl. 2007, 17, 1942–1953. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Scalley, T.H.; Bras, R.L. Tree censuses at Bisley Experimental Watersheds before and after Hurricane Hugo. In Fort Collins, CO: Forest Service Research Data Archive; Forest Service: Washington, DC, USA, 2022. [Google Scholar] [CrossRef]
- Uriarte, M.; Canham, C.D.; Thompson, J.; Zimmerman, J.K. A neighborhood analysis of tree growth and survival in a hurricane-driven tropical forest. Ecol. Monogr. 2004, 74, 591–614. [Google Scholar] [CrossRef]
- Kozlowski, T.T.; Winget, C.H. Diurnal and seasonal variation in radii of tree stems. Ecology 1964, 45, 149–155. [Google Scholar] [CrossRef]
- Raffelsbauer, V.; Spannl, S.; Peña, K.; Pucha-Cofrep, D.; Steppe, K.; Bräuning, A. Tree circumference changes and species-specific growth recovery after extreme dry events in a montane rainforest in Southern Ecuador. Front. Plant Sci. 2019, 10, 342. [Google Scholar] [CrossRef]
- Hogan, J.A.; McMahon, S.M.; Buzzard, V.; Michaletz, S.T.; Enquist, B.J.; Thompson, J.; Swenson, N.G.; Zimmerman, J.K. Drought and the interannual variability of stem growth in an aseasonal, everwet forest. Biotropica 2009, 51, 139–154. [Google Scholar] [CrossRef] [Green Version]
- Condit, R.; Hubbell, S.P.; Foster, R.B. Mortality and growth of a commercial hardwood ‘el cativo’, Prioria copaifera, in Panama. For. Ecol. Manage. 1993, 62, 107–122. [Google Scholar] [CrossRef]
- Forest Service. USDA National Core Field Guide; Volume I: Field Data Collection Procedures for Phase 2 Plots; Forest Inventory and Analysis; Version 8.0; Forest Service: Washington, DC, USA, 2018. [Google Scholar]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Early | Mid | Late | Palm | |
|---|---|---|---|---|
| 1989–1994 | 1665.00 | 44.74 | 16.45 | 94.74 |
| 1994–1999 | 104.82 | 131.90 | 44.06 | 86.49 |
| 1999–2004 | 19.72 | 18.63 | 32.96 | 36.84 |
| 2004–2009 | 4.66 | 7.51 | 9.90 | 15.12 |
| 2009–2014 | 13.33 | 8.04 | 10.45 | 9.73 |
| Census Year | Stem Density (# ha−1) | Basal Area (m2 ha−1) | Aboveground Biomass (Mg ha−1) |
|---|---|---|---|
| Pre-Hugo 1989 | 1440 (702) | 38 (33) | 255 (271) |
| Post-Hugo 1989 | 604 (398) | 22 (26) | 152 (211) |
| 1994 | 1275 (569) | 32 (28) | 199 (226) |
| 1999 | 2254 (928) | 36 (26) | 217 (205) |
| 2004 | 2072 (756) | 38 (25) | 228 (210) |
| 2009 | 1645 (648) | 39 (27) | 238 (224) |
| 2014 | 1440 (642) | 39 (29) | 246 (240) |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, J.; Heartsill-Scalley, T.; Bras, R.L. Parsing Long-Term Tree Recruitment, Growth, and Mortality to Identify Hurricane Effects on Structural and Compositional Change in a Tropical Forest. Forests 2022, 13, 796. https://0-doi-org.brum.beds.ac.uk/10.3390/f13050796
Zhang J, Heartsill-Scalley T, Bras RL. Parsing Long-Term Tree Recruitment, Growth, and Mortality to Identify Hurricane Effects on Structural and Compositional Change in a Tropical Forest. Forests. 2022; 13(5):796. https://0-doi-org.brum.beds.ac.uk/10.3390/f13050796
Chicago/Turabian StyleZhang, Jiaying, Tamara Heartsill-Scalley, and Rafael L. Bras. 2022. "Parsing Long-Term Tree Recruitment, Growth, and Mortality to Identify Hurricane Effects on Structural and Compositional Change in a Tropical Forest" Forests 13, no. 5: 796. https://0-doi-org.brum.beds.ac.uk/10.3390/f13050796