
Fine Root Production and Soil Available Nutrients in Rubber Monoculture versus Rubber–Flemingia macrophylla Agroforestry

 , , , and
, , , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Site Description
2.2. Experimental Design
2.3. Estimation of Fine Root Production
2.4. Soil Chemistry Analysis
2.5. Data Analysis
3. Results
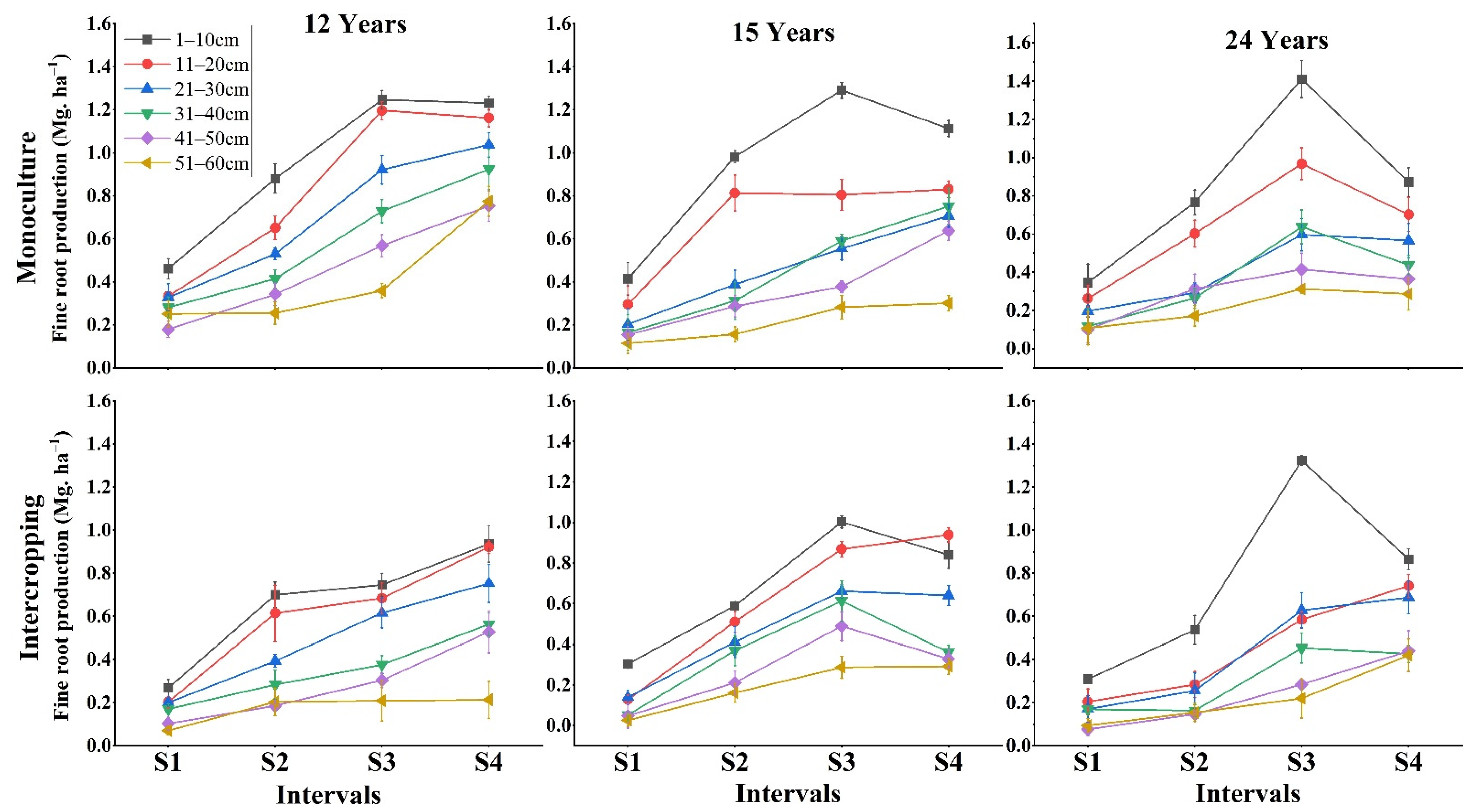
3.1. Impact of Depth, Season and Stand Age on Fine Root Production
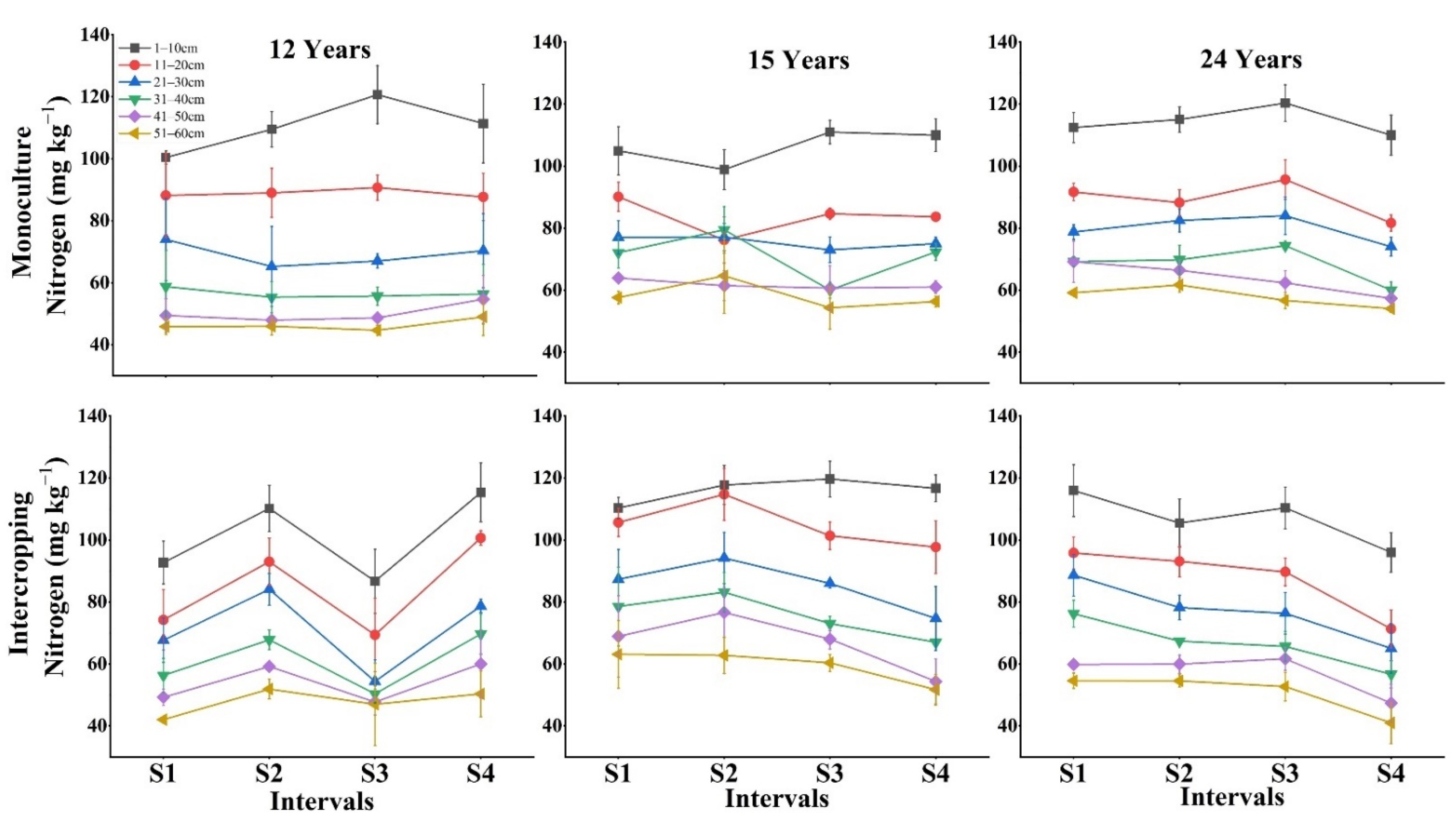
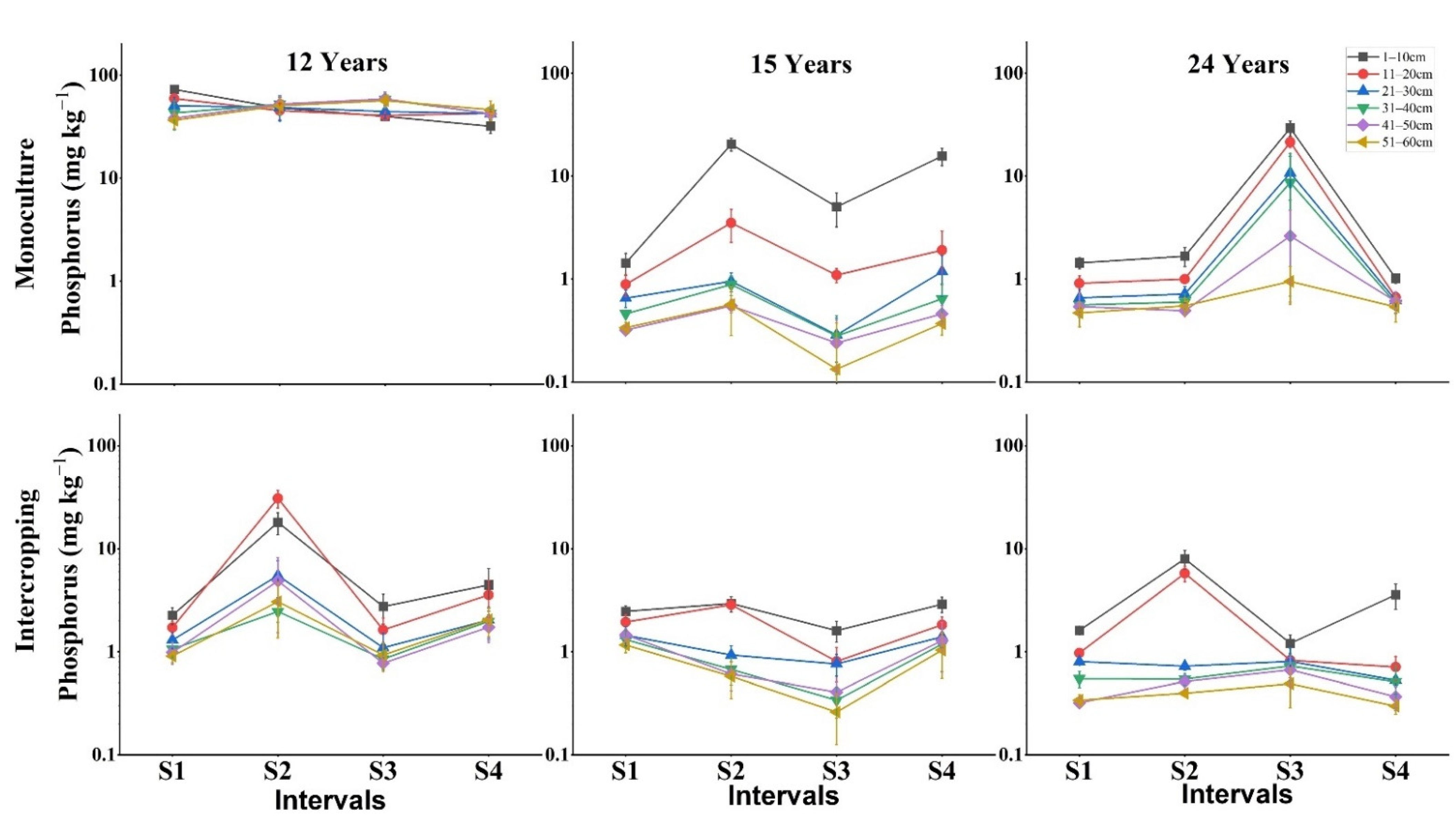
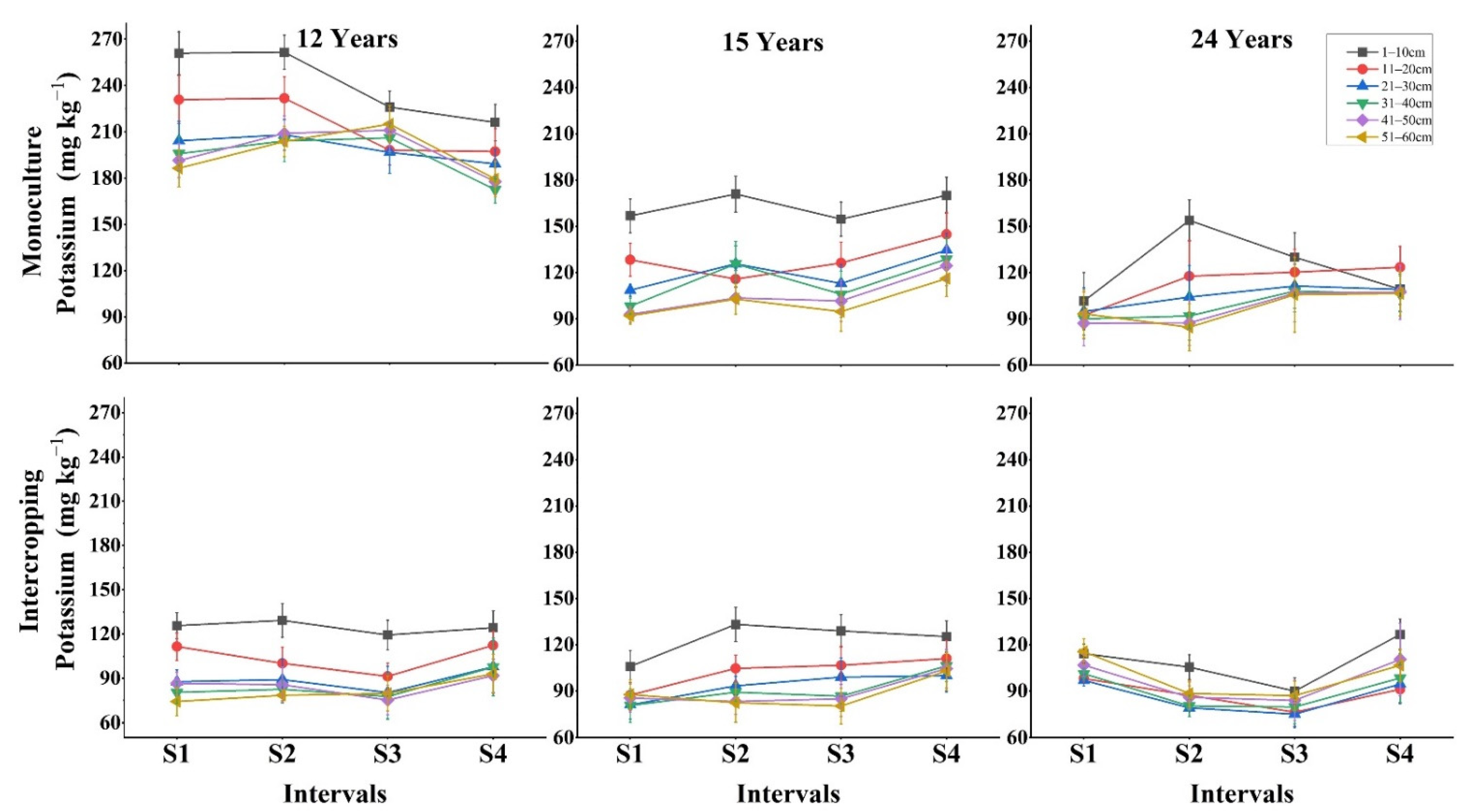
3.2. Impact of Depth, Season and Stand Age on Soil Available Nutrients
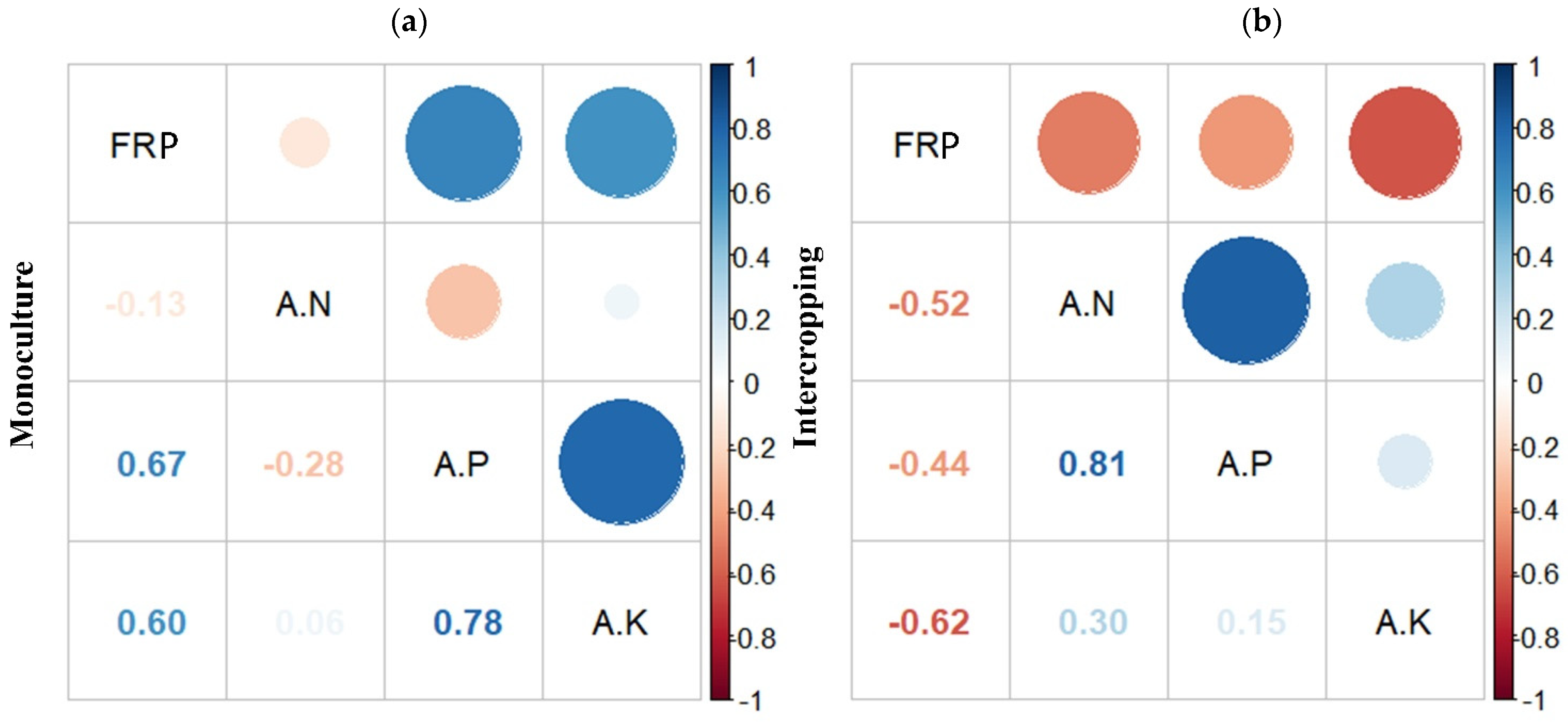
3.3. Correlations between Fine Root Mass Production and Soil Nutrients across Management Types
4. Discussion
4.1. Management Type Effects on Fine Root Production and Soil Nutrients
4.2. Stand Age Effects on Fine Root Production and Available Soil Nutrients
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Acknowledgments
Conflicts of Interest
References
- Rao, X.; Liu, C.-A.; Tang, J.-W.; Nie, Y.; Liang, M.-Y.; Shen, W.-J.; Siddique, K.H.M. Rubber-leguminous shrub systems stimulate soil N2O but reduce CO2 and CH4 emissions. For. Ecol. Manag. 2021, 480, 118665. [Google Scholar] [CrossRef]
- Edwards, D.P.; Laurance, S.G. Green labelling, sustainability and the expansion of tropical agriculture: Critical issues for certification schemes. Biol. Conserv. 2012, 151, 60–64. [Google Scholar] [CrossRef]
- Liu, C.; Pang, J.; Jepsen, M.R.; Lü, X.; Tang, J. Carbon Stocks across a Fifty Year Chronosequence of Rubber Plantations in Tropical China. Forests 2017, 8, 209. [Google Scholar] [CrossRef] [Green Version]
- Ziegler, A.D.; Fox, J.M.; Xu, J. The rubber juggernaut. Science 2009, 324, 1024–1025. [Google Scholar] [CrossRef] [PubMed]
- Chen, C.; Liu, W.; Wu, J.; Jiang, X.; Zhu, X. Can intercropping with the cash crop help improve the soil physico-chemical properties of rubber plantations? Geoderma 2019, 335, 149–160. [Google Scholar] [CrossRef]
- Liu, C.-A.; Nie, Y.; Rao, X.; Tang, J.-W.; Siddique, K.H.M. The effects of introducing Flemingia macrophylla to rubber plantations on soil water content and exchangeable cations. Catena 2019, 172, 480–487. [Google Scholar] [CrossRef]
- Singh, A.K. Prospective of rubber-based agroforestry on Swidden fallow for synergizing climate change mitigation and adaptation. Trop. Ecol. 2018, 59, 605–618. [Google Scholar]
- McCormack, M.L.; Dickie, I.A.; Eissenstat, D.M.; Fahey, T.J.; Fernandez, C.W.; Guo, D.; Helmisaari, H.; Hobbie, E.A.; Iversen, C.M.; Jackson, R.B. Redefining fine roots improves understanding of below-ground contributions to terrestrial biosphere processes. New Phytol. 2015, 207, 505–518. [Google Scholar] [CrossRef]
- Pierret, A.; Moran, C.J.; Doussan, C. Conventional detection methodology is limiting our ability to understand the roles and functions of fine roots. New Phytol. 2005, 166, 967–980. [Google Scholar] [CrossRef]
- Pregitzer, K.S.; DeForest, J.L.; Burton, A.J.; Allen, M.F.; Ruess, R.W.; Hendrick, R.L. Fine root architecture of nine North American trees. Ecol. Monogr. 2002, 72, 293–309. [Google Scholar] [CrossRef]
- Hishi, T. Heterogeneity of individual roots within the fine root architecture: Causal links between physiological and ecosystem functions. J. For. Res. 2007, 12, 126–133. [Google Scholar] [CrossRef]
- Raizada, A.; Jayaprakash, J.; Rathore, A.C.; Tomar, J.M.S. Distribution of fine root biomass of fruit and forest tree species raised on old river bed lands in the north west Himalaya. Trop. Ecol. 2013, 54, 251–261. [Google Scholar]
- Singha, D.; Tripathi, S.K. Variations in fine root growth during age chronosequence of moist tropical forest following shifting cultivation in Mizoram, northeast India. Trop. Ecol. 2017, 58, 769–779. [Google Scholar]
- Ma, X.; Wang, X.; Jin, Y.; Zhang, Y. Fine root production, turnover of Reaumuria songarica and soil carbon and nitrogen in Alxa steppe desert of NW China. Restor. Ecol. 2021, 29, e13362. [Google Scholar] [CrossRef]
- Lai, Z.; Zhang, Y.; Liu, J.; Wu, B.; Qin, S.; Fa, K. Fine-root distribution, production, decomposition, and effect on soil organic carbon of three revegetation shrub species in northwest China. For. Ecol. Manag. 2016, 359, 381–388. [Google Scholar] [CrossRef]
- Eissenstat, D.M.; Wells, C.E.; Yanai, R.D.; Whitbeck, J.L. Building roots in a changing environment: Implications for root longevity. New Phytol. 2000, 147, 33–42. [Google Scholar] [CrossRef] [Green Version]
- Yuan, Z.Y.; Chen, H.Y.H. Fine Root Biomass, Production, Turnover Rates, and Nutrient Contents in Boreal Forest Ecosystems in Relation to Species, Climate, Fertility, and Stand Age: Literature Review and Meta-Analyses. Crit. Rev. Plant Sci. 2010, 29, 204–221. [Google Scholar] [CrossRef]
- Tripathi, S.K. Fine root growth and soil nutrient dynamics during shifting cultivation in tropical semi-evergreen forests of northeast India. J. Environ. Biol. 2019, 40, 45–52. [Google Scholar]
- Zhang, Z.; Huang, M.; Zhang, Y. Vertical distribution of fine-root area in relation to stand age and environmental factors in black locust (Robinia pseudoacacia) forests of the Chinese Loess Plateau. Can. J. For. Res. 2018, 48, 1148–1158. [Google Scholar] [CrossRef]
- He, L.; Jia, Z.; Li, Q.; Zhang, Y.; Wu, R.; Dai, J.; Gao, Y. Fine root dynamic characteristics and effect on plantation’s carbon sequestration of three Salix shrub plantations in Tibetan Plateau alpine sandy land. Ecol. Evol. 2021, 11, 2645–2659. [Google Scholar] [CrossRef]
- Langenberger, G.; Cadisch, G.; Martin, K.; Min, S.; Waibel, H. Rubber intercropping: A viable concept for the 21st century? Agrofor. Syst. 2017, 91, 577–596. [Google Scholar] [CrossRef]
- Liu, C.; Jin, Y.; Hu, Y.; Tang, J.; Xiong, Q.; Xu, M.; Bibi, F.; Beng, K.C. Drivers of soil bacterial community structure and diversity in tropical agroforestry systems. Agric. Ecosyst. Environ. 2019, 278, 24–34. [Google Scholar] [CrossRef]
- Yang, B.; Meng, X.; Zhu, X.; Zakari, S.; Singh, A.K.; Bibi, F.; Mei, N.; Song, L.; Liu, W. Coffee performs better than amomum as a candidate in the rubber agroforestry system: Insights from water relations. Agric. Water Manag. 2021, 244, 106593. [Google Scholar] [CrossRef]
- Orwa, C.; Mutua, A.; Kindt, R.; Jamnadass, R.; Simons, A. Agroforestree Database: A Tree Reference and Selection Guide, Version 4; World Agroforestry Centre: Nairobi, Kenya, 2009. [Google Scholar]
- Pang, J.P.; Chen, M.Y.; Tang, J.W.; Guo, X.M.; Zeng, R. The dynamics of plant growth and soil moisture and nutrient in the rubber plantation and rubber–Flemingia macrophylla agroforestry system in Xishuangbanna, Southwest China. J. Mt. Sci. 2009, 27, 433–441. [Google Scholar]
- Luo, Q.P.; Liu, W.J. Comparison of the effect of different canopy of rubber plantation on soil splash erosion and through fall erosivity. Chin. J. Soil Sci. 2012, 43, 1348–1354. (In Chinese) [Google Scholar]
- Zhu, C.J.; Liu, W.J.; Wu, J.E. Rainfall erosivity and rainfall kinetic energy under rubber agroforestry system in Xishuangbanna. Chin. J. Soil Sci. 2014, 45, 1218–1224. [Google Scholar]
- Fukuzawa, S.; Oki, H.; Hosaka, M.; Sugasawa, J.; Kikuchi, S. ClickFerrophos: New chiral ferrocenyl phosphine ligands synthesized by click chemistry and the use of their metal complexes as catalysts for asymmetric hydrogenation and allylic substitution. Org. Lett. 2007, 9, 5557–5560. [Google Scholar] [CrossRef]
- Dybzinski, R.; Fargione, J.E.; Zak, D.R.; Fornara, D.; Tilman, D. Soil fertility increases with plant species diversity in a long-term biodiversity experiment. Oecologia 2008, 158, 85–93. [Google Scholar] [CrossRef]
- Liu, G.; Yu, Y.; Hou, J.; Xue, W.; Liu, X.; Liu, Y.; Wang, W.; Alsaedi, A.; Hayat, T.; Liu, Z. An ecological risk assessment of heavy metal pollution of the agricultural ecosystem near a lead-acid battery factory. Ecol. Indic. 2014, 47, 210–218. [Google Scholar] [CrossRef]
- Cordeiro, A.L.; Norby, R.J.; Andersen, K.M.; Valverde-Barrantes, O.; Fuchslueger, L.; Oblitas, E.; Hartley, I.P.; Iversen, C.M.; Gonçalves, N.B.; Takeshi, B. Fine-root dynamics vary with soil depth and precipitation in a lown-utrient tropical forest in the Central Amazonia. Plant-Environ. Interact. 2020, 1, 3–16. [Google Scholar] [CrossRef] [Green Version]
- Vitousek, P. Nutrient cycling and nutrient use efficiency. Am. Nat. 1982, 119, 553–572. [Google Scholar] [CrossRef]
- Vitousek, P.M. Litterfall, nutrient cycling, and nutrient limitation in tropical forests. Ecology 1984, 65, 285–298. [Google Scholar] [CrossRef]
- Brassard, B.W.; Chen, H.Y.H.; Cavard, X.; Ere, L.; Reich, P.B.; Bergeron, Y.; Par, D.; Yuan, Z. Tree species diversity increases fine root productivity through increased soil volume filling. J. Ecol. 2013, 101, 210–219. [Google Scholar] [CrossRef]
- Mueller, K.E.; Tilman, D.; Fornara, D.A.; Hobbie, S.E. Root depth distribution and the diversity–productivity relationship in a long-term grassland experiment. Ecology 2013, 94, 787–793. [Google Scholar] [CrossRef] [Green Version]
- Joslin, J.D.; Wolfe, M.H.; Hanson, P.J. Effects of altered water regimes on forest root systems. New Phytol. 2000, 147, 117–129. [Google Scholar] [CrossRef]
- Vasconcelos, A.C.M.; Casagrande, A.A.; Perecin, D.; Jorge, L.A.C.; Landell, M.G.A. Avaliação do sistema radicular da cana-de-açúcar por diferentes métodos. Rev. Bras. Ciência Solo 2003, 27, 849–858. [Google Scholar] [CrossRef] [Green Version]
- Walley, F.L.; Van Kessel, C.; Pennock, D.J. Landscape-scale variability of N mineralization in forest soils. Soil Biol. Biochem. 1996, 28, 383–391. [Google Scholar] [CrossRef]
- Singha, D.; Brearley, F.Q.; Tripathi, S.K. Fine root and soil nitrogen dynamics during stand development following shifting agriculture in Northeast India. Forests 2020, 11, 1236. [Google Scholar] [CrossRef]
- Bautista-Cruz, A.; Del Castillo, R.F. Soil changes during secondary succession in a tropical montane cloud forest area. Soil Sci. Soc. Am. J. 2005, 69, 906–914. [Google Scholar] [CrossRef]
- Eaton, J.M.; Lawrence, D. Loss of carbon sequestration potential after several decades of shifting cultivation in the Southern Yucatán. For. Ecol. Manag. 2009, 258, 949–958. [Google Scholar] [CrossRef]
- Powers, J.S.; Marín-Spiotta, E. Ecosystem processes and biogeochemical cycles in secondary tropical forest succession. Annu. Rev. Ecol. Evol. Syst. 2017, 48, 497–519. [Google Scholar] [CrossRef] [Green Version]
- Zhang, H.; Shi, L.; Wen, D.; Yu, K. Soil potential labile but not occluded phosphorus forms increase with forest succession. Biol. Fertil. Soils 2016, 52, 41–51. [Google Scholar] [CrossRef]
- Liu, X.; Wang, Z.; Huang, C.; Li, M.; Bibi, F.; Zhou, S.; Nakamura, A. Ant assemblage composition explains high predation pressure on artificial caterpillars during early night. Ecol. Entomol. 2020, 45, 547–554. [Google Scholar] [CrossRef]
- Balasubramanian, D.; Zhang, Y.-P.; Grace, J.; Sha, L.-Q.; Jin, Y.; Zhou, L.-G.; Lin, Y.-X.; Zhou, R.-W.; Gao, J.-B.; Song, Q.-H. Soil organic matter as affected by the conversion of natural tropical rainforest to monoculture rubber plantations under acric ferralsols. Catena 2020, 195, 104753. [Google Scholar] [CrossRef]
- Liu, C.; Jin, Y.; Liu, C.; Tang, J.; Wang, Q.; Xu, M. Phosphorous fractions in soils of rubber-based agroforestry systems: Influence of season, management and stand age. Sci. Total Environ. 2018, 616, 1576–1588. [Google Scholar] [CrossRef]
- Adamek, M.; Corre, M.D.; Hölscher, D. Responses of fine roots to experimental nitrogen addition in a tropical lower montane rain forest, Panama. J. Trop. Ecol. 2011, 27, 73–81. [Google Scholar] [CrossRef]
- Kubisch, P.; Leuschner, C.; Coners, H.; Gruber, A.; Hertel, D. Fine root abundance and dynamics of stone pine (Pinus cembra) at the alpine treeline is not impaired by self-shading. Front. Plant Sci. 2017, 8, 602. [Google Scholar] [CrossRef] [Green Version]
- Cai, H.; Li, F.; Jin, G. Fine root biomass, production and turnover rates in plantations versus natural forests: Effects of stand characteristics and soil properties. Plant Soil 2019, 436, 463–474. [Google Scholar] [CrossRef]
- Zhong, Y.; Yan, W.; Shangguan, Z. Soil carbon and nitrogen fractions in the soil profile and their response to long-term nitrogen fertilization in a wheat field. Catena 2015, 135, 38–46. [Google Scholar] [CrossRef]
- Anderson, J.M.; Ingram, J.S.I. Tropical Soil Biology and Fertility; CAB International Wallingford: Wallingford, UK, 1989; ISBN 0851986358. [Google Scholar]
- Atapattu, A.J.; Xia, S.; Cao, M.; Zhang, W.; Mishra, S.; Yang, X. Can Dominant Canopy Species Leaf Litter Determine Soil Nutrient Heterogeneity? A Case Study in a Tropical Rainforest in Southwest China. J. Soil Sci. Plant Nutr. 2020, 20, 2479–2489. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment For Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2013. [Google Scholar]
- Bates, D.; Maechler, M.; Bolker, B.M.; Walker, S. Fitting linear mixed-effects models using lme4. J. Stat. Softw. 2015, 67, 1–48. [Google Scholar] [CrossRef]
- Jacob, A.; Hertel, D.; Leuschner, C. On the significance of belowground overyielding in temperate mixed forests: Separating species identity and species diversity effects. Oikos 2013, 122, 463–473. [Google Scholar] [CrossRef]
- Brassard, B.W.; Chen, H.Y.H.; Bergeron, Y.; Paré, D. Differences in fine root productivity between mixed-and single-species stands. Funct. Ecol. 2011, 25, 238–246. [Google Scholar] [CrossRef]
- Domisch, T.; Finér, L.; Dawud, S.M.; Vesterdal, L.; Raulund-Rasmussen, K. Does species richness affect fine root biomass and production in young forest plantations? Oecologia 2015, 177, 581–594. [Google Scholar] [CrossRef]
- Sun, Z.; Liu, X.; Schmid, B.; Bruelheide, H.; Bu, W.; Ma, K. Positive effects of tree species richness on fine-root production in a subtropical forest in SE-China. J. Plant Ecol. 2017, 10, 146–157. [Google Scholar] [CrossRef] [Green Version]
- Archambault, C.; Paquette, A.; Messier, C.; Khlifa, R.; Munson, A.D.; Handa, I.T. Evergreenness influences fine root growth more than tree diversity in a common garden experiment. Oecologia 2019, 189, 1027–1039. [Google Scholar] [CrossRef]
- Pramanik, P.; Safique, S.; Zahan, A.; Phukan, M.; Ghosh, S. Cellulolytic microorganisms control the availability of nitrogen in microcosm of shredded pruning litter treated highly acidic tea-growing soils of Assam in Northeast India. Appl. Soil Ecol. 2017, 120, 30–34. [Google Scholar] [CrossRef]
- Petraglia, A.; Cacciatori, C.; Chelli, S.; Fenu, G.; Calderisi, G.; Gargano, D.; Abeli, T.; Orsenigo, S.; Carbognani, M. Litter decomposition: Effects of temperature driven by soil moisture and vegetation type. Plant Soil 2019, 435, 187–200. [Google Scholar] [CrossRef]
- Liu, C.-A.; Nie, Y.; Zhang, Y.-M.; Tang, J.-W.; Siddique, K.H.M. Introduction of a leguminous shrub to a rubber plantation changed the soil carbon and nitrogen fractions and ameliorated soil environments. Sci. Rep. 2018, 8, 17324. [Google Scholar] [CrossRef]
- Sardans, J.; Peñuelas, J. Drought changes phosphorus and potassium accumulation patterns in an evergreen Mediterranean forest. Funct. Ecol. 2007, 21, 191–201. [Google Scholar] [CrossRef]
- Rodrigues, M.; Pavinato, P.S.; Withers, P.J.A.; Teles, A.P.B.; Herrera, W.F.B. Legacy phosphorus and no tillage agriculture in tropical oxisols of the Brazilian savanna. Sci. Total Environ. 2016, 542, 1050–1061. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Effects | Fine Root Production (Mg ha−1) | Available Nitrogen (mg kg−1) | Available Phosphorus (mg kg−1) | Available Potassium (mg kg−1) | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Fixed Effect | df | Error df | SSq | F | p | SSq | F | p | SSq | F | p | SSq | F | p |
| Age (A) | 2 | 0 | 0.42 | 1.11 | 1.000 | 0.07 | 1.43 | 1.000 | 1.74 | 1.94 | 0.146 | 0.02 | 0.21 | 1.000 |
| Intervals (I) | 3 | 276 | 127.18 | 221.18 | 0.001 | 0.41 | 5.62 | 0.001 | 10.88 | 8.07 | 0.001 | 0.56 | 4.31 | 0.005 |
| Managements (M) | 1 | 0 | 0.95 | 4.96 | 1.000 | 0.00 | 0.02 | 1.000 | 0.60 | 1.34 | 0.248 | 0.05 | 1.19 | 1.000 |
| Depths (D) | 5 | 276 | 88.06 | 91.89 | 0.001 | 26.35 | 215.84 | 0.000 | 89.38 | 39.76 | 0.001 | 4.91 | 22.84 | 0.001 |
| I × M | 3 | 276 | 1.16 | 2.01 | 0.112 | 0.25 | 3.44 | 0.017 | 13.00 | 9.64 | 0.001 | 0.46 | 3.56 | 0.015 |
| I × A | 6 | 276 | 5.11 | 4.44 | 0.001 | 1.15 | 7.85 | 0.000 | 30.21 | 11.20 | 0.001 | 0.52 | 2.03 | 0.062 |
| M × A | 2 | 0 | 0.20 | 0.53 | 1.000 | 0.02 | 0.46 | 1.000 | 1.23 | 1.37 | 0.256 | 0.03 | 0.38 | 1.000 |
| I × M × A | 6 | 276 | 5.09 | 4.42 | 0.001 | 0.74 | 5.05 | 0.000 | 17.23 | 6.39 | 0.001 | 0.95 | 3.68 | 0.002 |
| A × D | 10 | 276 | 2.17 | 1.13 | 0.337 | 0.34 | 1.41 | 0.175 | 16.36 | 3.64 | 0.001 | 0.53 | 1.24 | 0.267 |
| M × D | 5 | 276 | 0.93 | 0.97 | 0.437 | 0.12 | 0.96 | 0.445 | 1.39 | 0.62 | 0.686 | 0.13 | 0.59 | 0.707 |
| I × D | 15 | 276 | 2.79 | 0.97 | 0.486 | 0.18 | 0.50 | 0.937 | 4.75 | 0.70 | 0.779 | 0.27 | 0.42 | 0.973 |
| I × M × D | 15 | 276 | 2.04 | 0.71 | 0.775 | 0.20 | 0.55 | 0.912 | 4.24 | 0.63 | 0.851 | 0.08 | 0.13 | 1.000 |
| I × A × D | 30 | 276 | 3.78 | 0.66 | 0.917 | 0.18 | 0.24 | 1.000 | 6.33 | 0.47 | 0.993 | 0.40 | 0.31 | 1.000 |
| M × A × D | 10 | 276 | 2.21 | 1.16 | 0.321 | 0.26 | 1.08 | 0.377 | 13.98 | 3.11 | 0.001 | 0.37 | 0.86 | 0.574 |
| I × M × A × D | 30 | 276 | 2.87 | 0.50 | 0.988 | 0.19 | 0.25 | 1.000 | 6.58 | 0.49 | 0.990 | 0.33 | 0.26 | 1.000 |
| Variation explained | ||||||||||||||
| R2m | 74 | 0.77 | 0.85 | 0.64 | ||||||||||
| R2c | 77 | 0.79 | 0.89 | 0.77 | ||||||||||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bibi, F.; Tomlinson, K.W.; Liu, C.; Liu, C.; Jin, Y.; Tang, J. Fine Root Production and Soil Available Nutrients in Rubber Monoculture versus Rubber–Flemingia macrophylla Agroforestry. Forests 2022, 13, 830. https://0-doi-org.brum.beds.ac.uk/10.3390/f13060830
Bibi F, Tomlinson KW, Liu C, Liu C, Jin Y, Tang J. Fine Root Production and Soil Available Nutrients in Rubber Monoculture versus Rubber–Flemingia macrophylla Agroforestry. Forests. 2022; 13(6):830. https://0-doi-org.brum.beds.ac.uk/10.3390/f13060830
Chicago/Turabian StyleBibi, Farkhanda, Kyle W. Tomlinson, Chenggang Liu, Chang’an Liu, Yanqiang Jin, and Jianwei Tang. 2022. "Fine Root Production and Soil Available Nutrients in Rubber Monoculture versus Rubber–Flemingia macrophylla Agroforestry" Forests 13, no. 6: 830. https://0-doi-org.brum.beds.ac.uk/10.3390/f13060830