Mean Leaf Angles Affect Irrigation Efficiency and Physiological Responses of Tropical Species Seedling


 , , , and
, , , and
Abstract
:1. Introduction
2. Materials and Methods
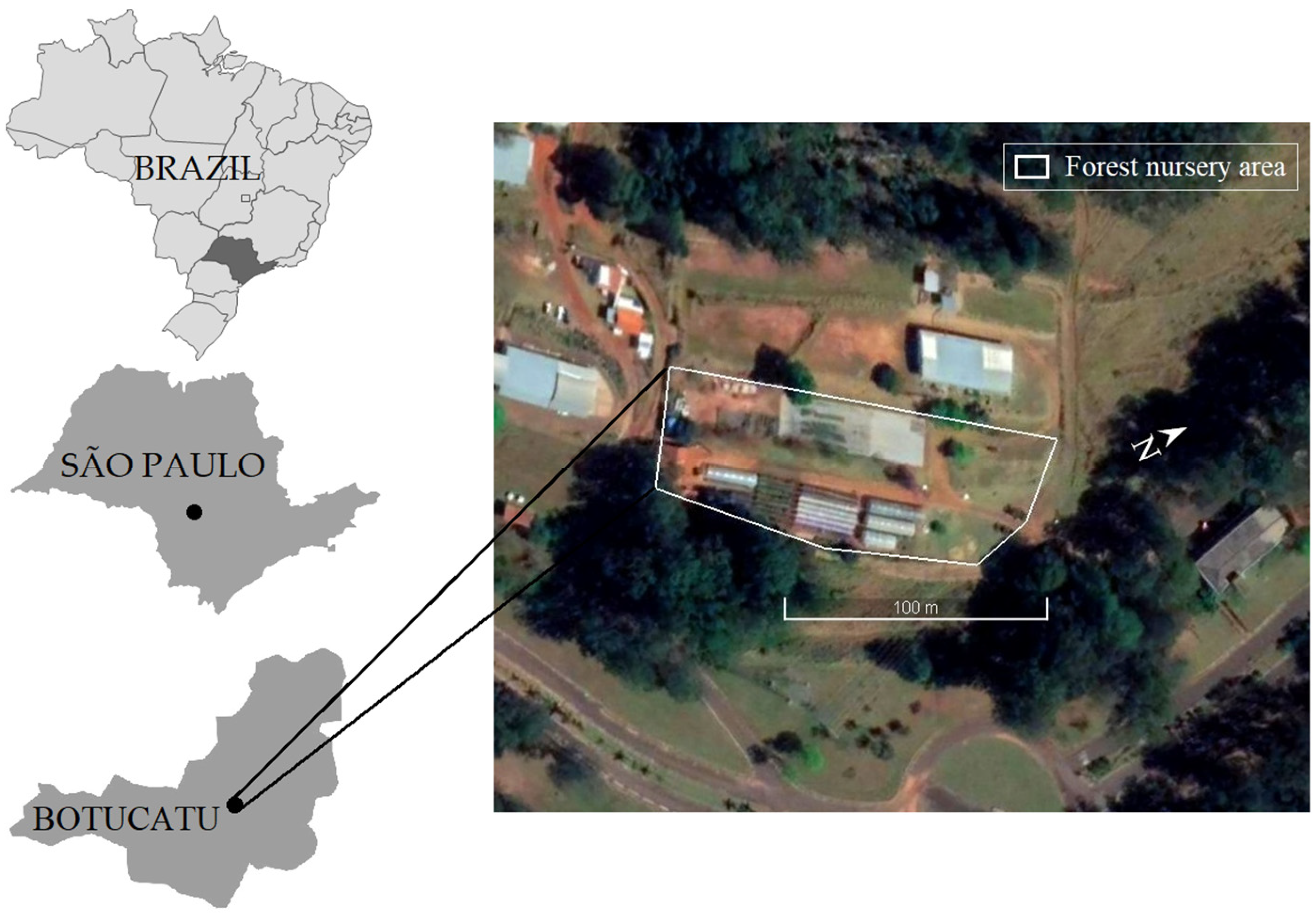
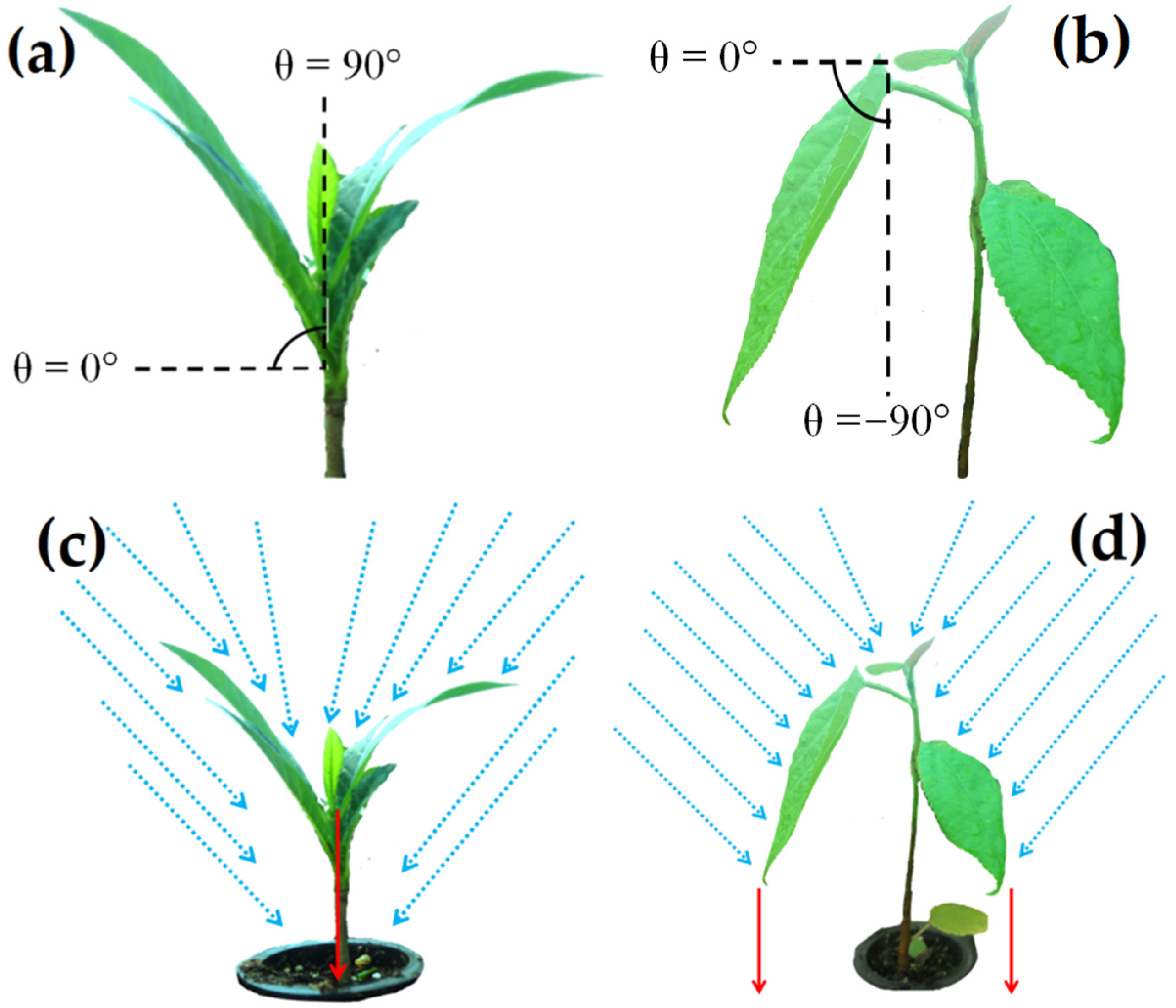
2.1. Study Site and Mean Leaf Angle Characterization
2.2. Experimental Design and Treatments
2.3. Nursery Culture
2.4. Physiological Analysis
2.5. Leaching Fraction
2.6. Total Dry Mass
2.7. Data Analysis
3. Results
3.1. Leaching Fraction
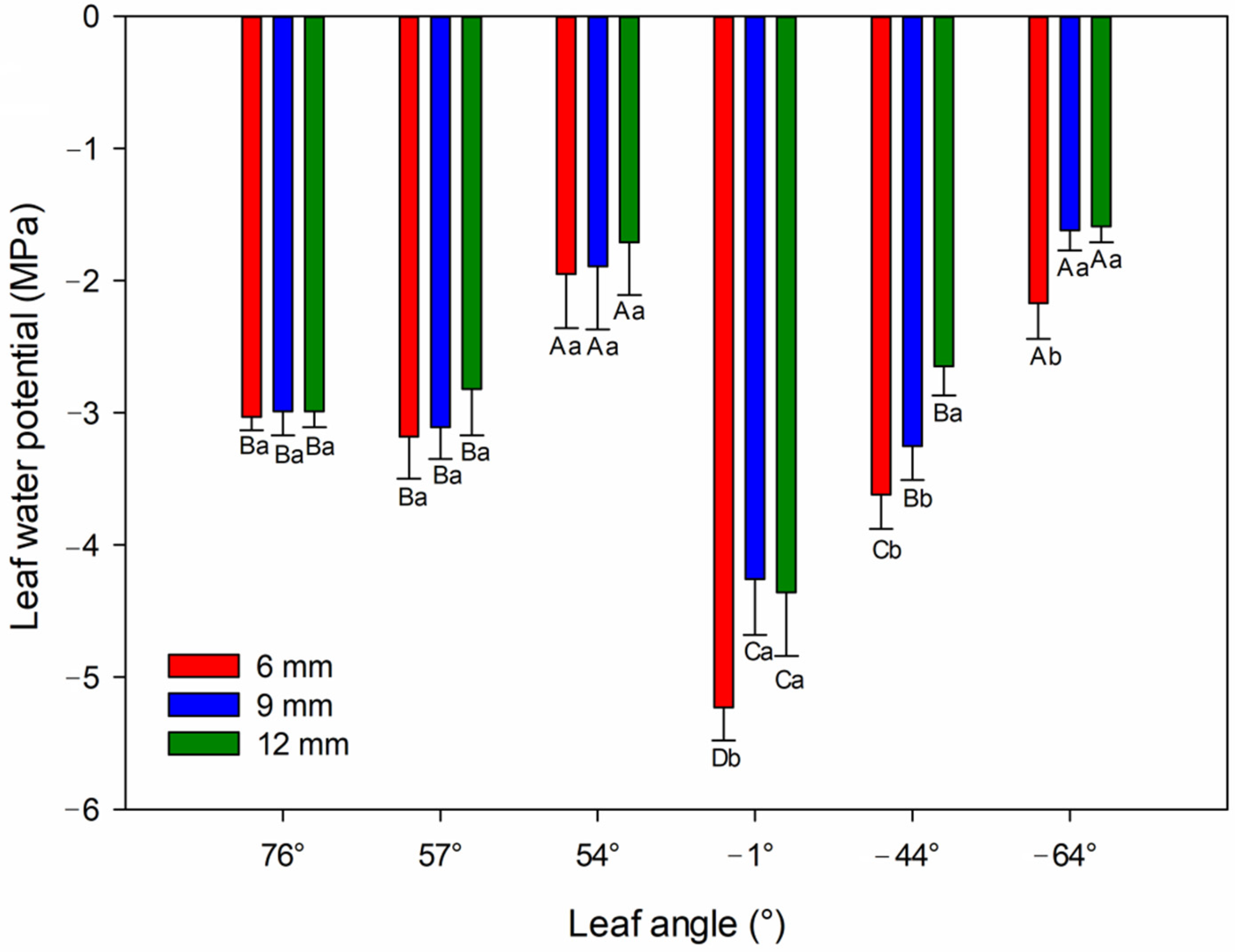
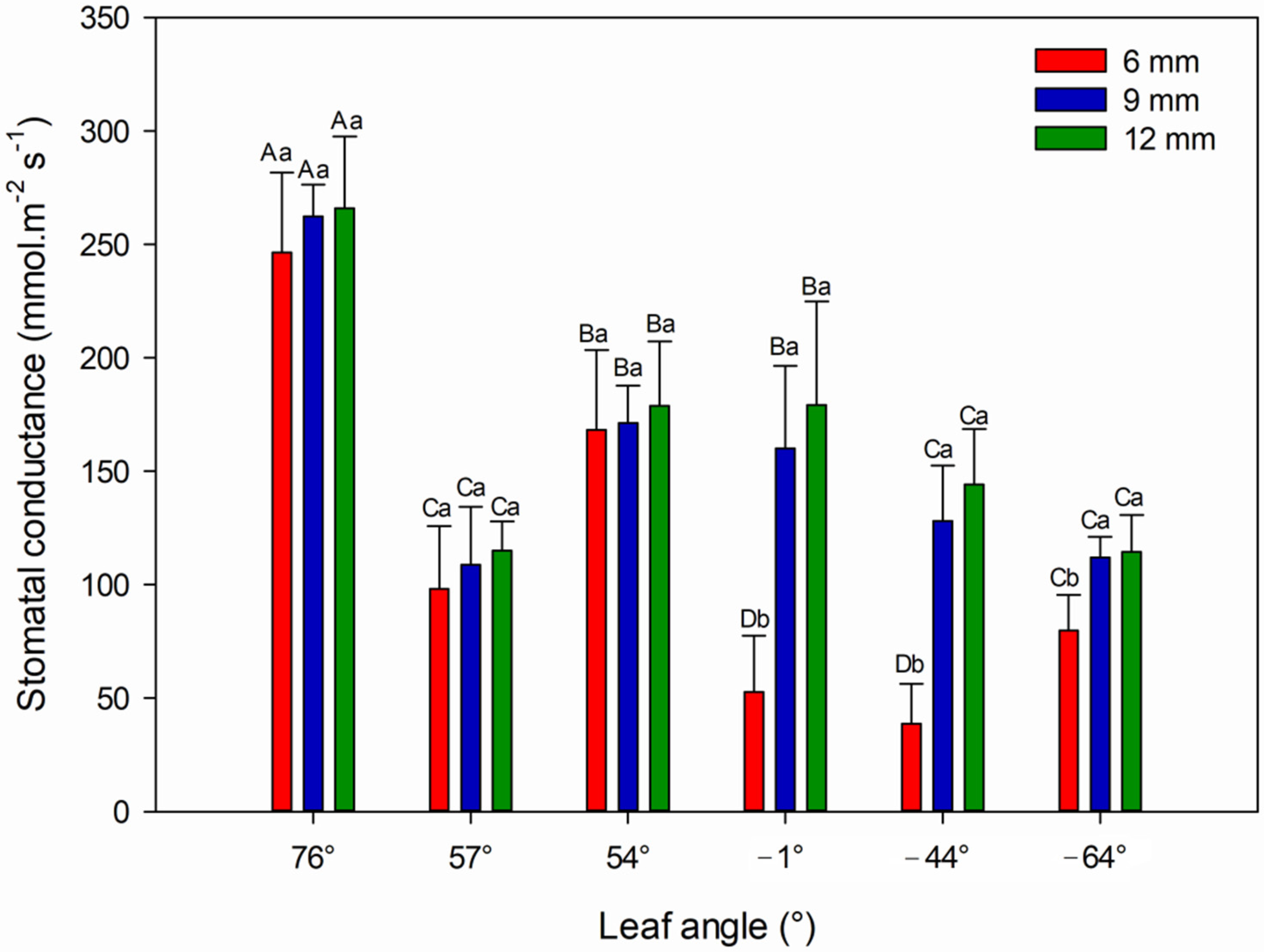
3.2. Physiological Analysis
3.3. Total Dry Mass
4. Discussion
4.1. Leaching Fraction
4.2. Leaf Water Potential
4.3. Stomatal Conductance
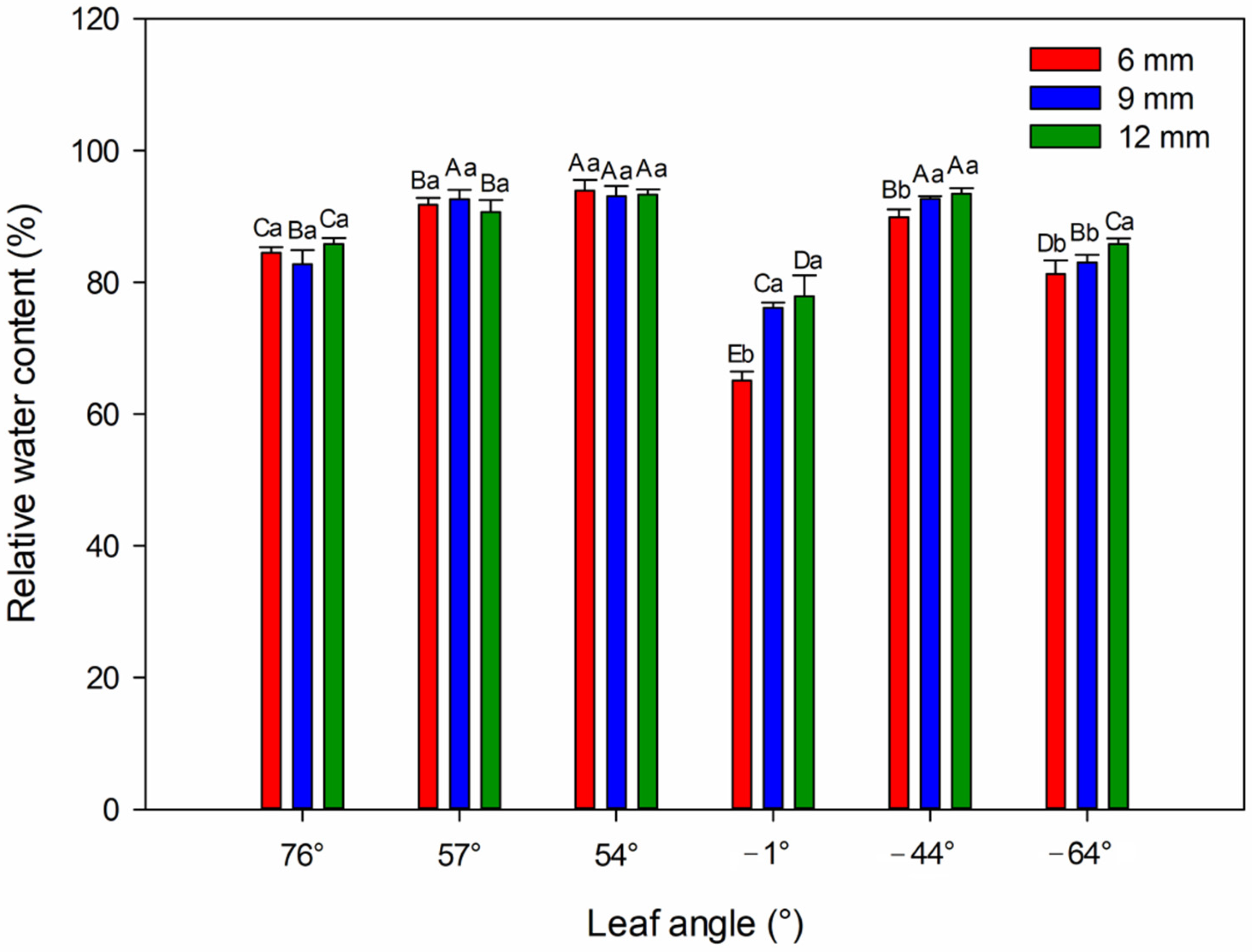
4.4. Relative Water Content
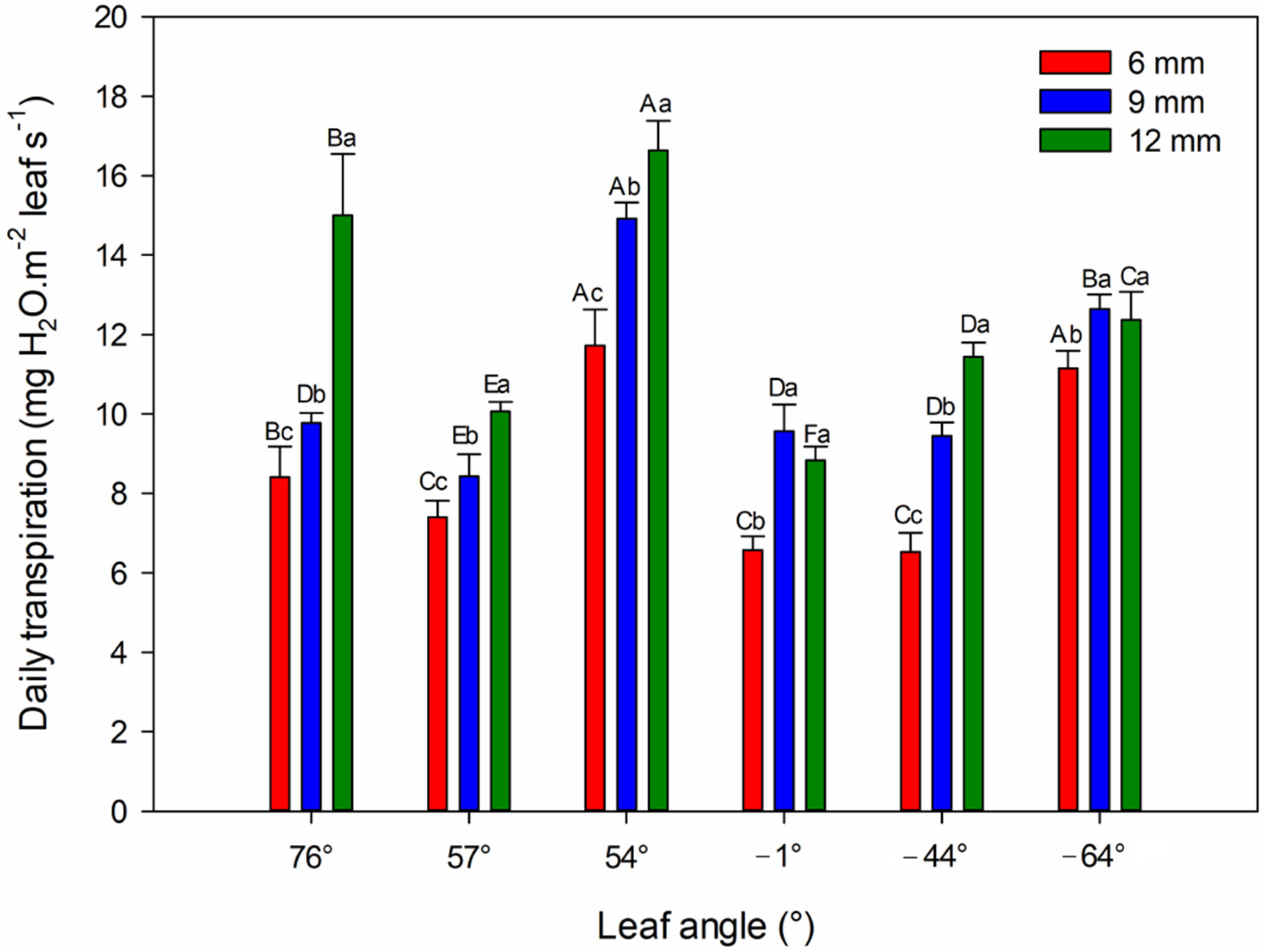
4.5. Daily Transpiration
4.6. Total Dry Mass
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Thebaldi, M.S.; Lima, L.A.; Braga Colares, M.D.F.; da Silva, A.C.; Lima, P.L.T. Dinâmica das características químicas de um substrato florestal exposto à irrigação. Ciência Florest. 2015, 25, 375–384. [Google Scholar] [CrossRef] [Green Version]
- Riikonen, J.; Luoranen, J. Seedling production and the field performance of seedlings. Forests 2018, 9, 740. [Google Scholar] [CrossRef] [Green Version]
- Wilson, P.C.; Albano, J.P. Impact of Fertigation versus Controlled-release Fertilizer Formulations on Nitrate Concentrations in Nursery Drainage Water. HortTechnol. 2011, 21, 176–180. [Google Scholar] [CrossRef] [Green Version]
- Ciavatta, S.F.; da Silva, M.R.; Simões, D. Fertirrigação na produção de mudas de Eucalyptus grandis nos períodos de inverno e verão. CERNE 2014, 20, 217–222. [Google Scholar] [CrossRef] [Green Version]
- Kormanek, M.; Małek, S.; Banach, J.; Durło, G.; Jagiełło-Leńczuk, K.; Dudek, K. Seasonal changes of perlite–peat substrate properties in seedlings grown in different sized container trays. New For. 2021, 52, 271–283. [Google Scholar] [CrossRef]
- Fulcher, A.; LeBude, A.V.; Owen, J.S.; White, S.A.; Beeson, R.C. The next ten years: Strategic vision of water resources for nursery producers. Horttechnology 2016, 26, 121–132. [Google Scholar] [CrossRef] [Green Version]
- Lea-Cox, J.D.; Bauerle, W.L.; van Iersel, M.W.; Kantor, G.F.; Bauerle, T.L.; Lichtenberg, E.; King, D.M.; Crawford, L. Advancing Wireless Sensor Networks for Irrigation Management of Ornamental Crops: An Overview. HortTechnol. 2013, 23, 717–724. [Google Scholar] [CrossRef] [Green Version]
- Incrocci, L.; Marzialetti, P.; Incrocci, G.; Di Vita, A.; Balendonck, J.; Bibbiani, C.; Spagnol, S.; Pardossi, A. Substrate water status and evapotranspiration irrigation scheduling in heterogenous container nursery crops. Agric. Water Manag. 2014, 131, 30–40. [Google Scholar] [CrossRef]
- Tawegoum, R.; Leroy, F.; Sintes, G.; Chassériaux, G. Forecasting hourly evapotranspiration for triggering irrigation in nurseries. Biosyst. Eng. 2015, 129, 237–247. [Google Scholar] [CrossRef]
- Davies, M.J.; Harrison-Murray, R.; Atkinson, C.J.; Grant, O.M. Application of deficit irrigation to container-grown hardy ornamental nursery stock via overhead irrigation, compared to drip irrigation. Agric. Water Manag. 2016, 163, 244–254. [Google Scholar] [CrossRef] [Green Version]
- Yeager, T.; Million, J.; Larsen, C.; Stamps, B. Florida nursery best management practices: Past, present, and future. Horttechnology 2010, 20, 82–88. [Google Scholar] [CrossRef] [Green Version]
- Reinhardt, D.; Kuhlemeier, C. Plant architecture. EMBO Rep. 2002, 3, 846–851. [Google Scholar] [CrossRef] [PubMed]
- Falster, D.S.; Westoby, M. Leaf size and angle vary widely across species: What consequences for light interception? New Phytol. 2003, 158, 509–525. [Google Scholar] [CrossRef] [Green Version]
- James, S.A.; Bell, D.T. Leaf orientation, light interception and stomatal conductance of Eucalyptus globulus ssp. globulus leaves. Tree Physiol. 2000, 20, 815–823. [Google Scholar] [CrossRef] [Green Version]
- Niinemets, Ü. A review of light interception in plant stands from leaf to canopy in different plant functional types and in species with varying shade tolerance. Ecol. Res. 2010, 25, 693–714. [Google Scholar] [CrossRef]
- Utsugi, H.; Araki, M.; Kawasaki, T.; Ishizuka, M. Vertical distributions of leaf area and inclination angle, and their relationship in a 46-year-old Chamaecyparis obtusa stand. For. Ecol. Manag. 2006, 225, 104–112. [Google Scholar] [CrossRef]
- Mantilla-Perez, M.B.; Salas Fernandez, M.G. Differential manipulation of leaf angle throughout the canopy: Current status and prospects. J. Exp. Bot. 2017, 68, 5699–5717. [Google Scholar] [CrossRef] [Green Version]
- Silva, R.B.; Gabira, M.M.; Prado, D.Z.; Uesugi, G.; Simões, D.; Silva, M.R. Influence of Mean Leaf Angles and Irrigation Volumes on Water Capture, Leaching, and Growth of Tropical Tree Seedlings. Forests 2020, 11, 1198. [Google Scholar] [CrossRef]
- Tonello, K.C.; Teixeira Filho, J. Transpiração e condutância estomática de Eucalyptus sp. em resposta à radiação global e diferentes condições hídricas. IRRIGA 2013, 18, 607–623. [Google Scholar] [CrossRef]
- Grossnickle, S.C.; MacDonald, J.E. Seedling Quality: History, Application, and Plant Attributes. Forests 2018, 9, 283. [Google Scholar] [CrossRef] [Green Version]
- Davis, A.S.; Jacobs, D.F. Quantifying root system quality of nursery seedlings and relationship to outplanting performance. New For. 2005, 30, 295–311. [Google Scholar] [CrossRef]
- Luis, V.C.; Puértolas, J.; Climent, J.; Peters, J.; González-Rodríguez, Á.M.; Morales, D.; Jiménez, M.S. Nursery fertilization enhances survival and physiological status in Canary Island pine (Pinus canariensis) seedlings planted in a semiarid environment. Eur. J. For. Res. 2009, 128, 221–229. [Google Scholar] [CrossRef]
- Da Cunha, A.R.; Martins, D. Classificação climática para os municípios de Botucatu e São Manuel, SP. IRRIGA 2009, 14, 1–11. [Google Scholar] [CrossRef]
- Elsheery, N.I.; Cao, K.-F. Gas exchange, chlorophyll fluorescence, and osmotic adjustment in two mango cultivars under drought stress. Acta Physiol. Plant. 2008, 30, 769–777. [Google Scholar] [CrossRef]
- Smart, R.E.; Bingham, G.E. Rapid Estimates of Relative Water Content. Plant Physiol. 1974, 53, 258–260. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aroca, R.; Porcel, R.; Ruiz-Lozano, J.M. How does arbuscular mycorrhizal symbiosis regulate root hydraulic properties and plasma membrane aquaporins in Phaseolus vulgaris under drought, cold or salinity stresses? New Phytol. 2007, 173, 808–816. [Google Scholar] [CrossRef]
- Lea-Cox, J.D.; Ross, D.S.; Teffeau, K.M. A water and nutrient management planning process for container nursery and greenhouse production systems in Maryland. J. Environ. Hortic. 2001, 19, 230–236. [Google Scholar] [CrossRef]
- Shapiro, S.S.; Wilk, M.B. An Analysis of Variance Test for Normality (Complete Samples). Biometrika 1965, 52, 591–611. [Google Scholar] [CrossRef]
- Fisher, R.A. Statistical Methods for Research Workers; Oliver and Boyd: London, UK, 1925. [Google Scholar]
- Scott, A.J.; Knott, M. A Cluster Analysis Method for Grouping Means in the Analysis of Variance. Biometrics 1974, 30, 507–512. [Google Scholar] [CrossRef] [Green Version]
- STATISTICA (Data Analysis Software System), version 8; StatSoft, Inc.: Tulsa, OK, USA, 2007.
- Karam, N.S.; Niemiera, A.X. Cyclic Sprinkler Irrigation and Pre-irrigation Substrate Water Content Affect Water and N Leaching from Containers. J. Environ. Hortic. 1994, 12, 198–202. [Google Scholar] [CrossRef]
- Beeson, R.C.; Yeager, T.H. Plant Canopy Affects Sprinkler Irrigation Application Efficiency of Container-grown Ornamentals. HortScience 2003, 38, 1373–1377. [Google Scholar] [CrossRef] [Green Version]
- Fochesato, M.L.; de Souza, P.V.D.; Schäfer, G.; Maciel, H.S. Alterações das características químicas de três substratos comerciais na produção de mudas cítricas. Ciência Agrotecnol. 2008, 32, 1040–1046. [Google Scholar] [CrossRef]
- Bumgarner, M.L.; Salifu, K.F.; Jacobs, D.F. Subirrigation of Quercus rubra Seedlings: Nursery Stock Quality, Media Chemistry, and Early Field Performance. HortScience 2008, 43, 2179–2185. [Google Scholar] [CrossRef] [Green Version]
- Da Ros, C.O.; Silvestrin, T.B.; Somavilla, L.; Perrando, E.R.; Silva, R.F. da Perdas de Nutrientes por Lixiviação na Produção de Mudas de Cedro Australiano. Floresta Ambient. 2017, 24, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Beeson, R.C. Modelling actual evapotranspiration of Ligustrum japonicum from rooted cuttings to commercially marketable plants in 12 liter black polyethylene containers. In Proceedings of the IV International Symposium on Irrigation of Horticultural Crops, Davis, CA, USA, 31 December 2004; pp. 71–77. [Google Scholar]
- De Rego, J.L.; de Oliveira, E.L.L.; Chaves, A.F.; Araújo, A.P.B.; Bezerra, F.M.L.; dos Santos, A.B.; Mota, S. Uso de esgoto doméstico tratado na irrigação da cultura da melancia. Rev. Bras. Eng. Agrícola Ambient. 2005, 9, 155–159. [Google Scholar] [CrossRef]
- Rodrigues, S.B.S.; Mantovani, E.C.; de Oliveira, R.A.; de Paiva, H.N.; Alves, M.E.B. Necessidades hídricas de mudas de eucalipto na região Centro-oeste de Minas Gerais. IRRIGA 2011, 16, 212–223. [Google Scholar] [CrossRef] [Green Version]
- Taiz, L.; Zeiger, E. Plant Physiology; ArtMed: Porto Alegre, Brazil, 2017. [Google Scholar]
- Kerbauy, G. Fisiologia Vegetal, 3rd ed.; Guanabara Koogan: Rio de Janeiro, Brazil, 2019; ISBN 9788527735339. [Google Scholar]
- Landsberg, J.; Sands, P. Physiological Ecology of Forest Production: Principles, Processes and Models; Academic Press: Cambridge, MA, USA, 2010; ISBN 9780080922546. [Google Scholar]
- Chen, X.; Zhao, P.; Ouyang, L.; Zhu, L.; Ni, G.; Schäfer, K.V.R. Whole-plant water hydraulic integrity to predict drought-induced Eucalyptus urophylla mortality under drought stress. For. Ecol. Manag. 2020, 468, 118179. [Google Scholar] [CrossRef]
- Rajan, L.J.; Santhoshkumar, A.V.; Surendra, G.K.; Kunhamu, T.K. Arbuscular Mycorrhizal Fungi Inoculation as a Climate Adaptation Strategy for Establishment of Swietenia macrophylla King. Seedlings. Forests 2020, 11, 488. [Google Scholar] [CrossRef]
- Ofori-Amanfo, K.K.; Klem, K.; Veselá, B.; Holub, P.; Agyei, T.; Marek, M.V.; Grace, J.; Urban, O. Interactive Effect of Elevated CO2 and Reduced Summer Precipitation on Photosynthesis is Species-Specific: The Case Study with Soil-Planted Norway Spruce and Sessile Oak in a Mountainous Forest Plot. Forests 2021, 12, 42. [Google Scholar] [CrossRef]
- Yu, M.-H.; Ding, G.-D.; Gao, G.-L.; Zhao, Y.-Y.; Yan, L.; Sai, K. Using Plant Temperature to Evaluate the Response of Stomatal Conductance to Soil Moisture Deficit. Forests 2015, 6, 3748–3762. [Google Scholar] [CrossRef] [Green Version]
- Kumarathunge, D.P.; Drake, J.E.; Tjoelker, M.G.; López, R.; Pfautsch, S.; Vårhammar, A.; Medlyn, B.E. The temperature optima for tree seedling photosynthesis and growth depend on water inputs. Glob. Chang. Biol. 2020, 26, 2544–2560. [Google Scholar] [CrossRef]
- Gabira, M.M.; Silva, R.; Bortolheiro, F.; Mateus, C.; Villas Boas, R.L.; Rossi, S.; Girona, M.M.; Silva, M.R.D. Composted sewage sludge as an alternative substrate for forest seedlings production. iForest—Biogeosci. For. 2021, 14, 569–575. [Google Scholar] [CrossRef]
- Gholinezhad, E. Impact of drought stress and stress modifiers on water use efficiency, membrane lipidation indices, and water relationship indices of pot marigold (Calendula officinalis L.). Braz. J. Bot. 2020, 43, 747–759. [Google Scholar] [CrossRef]
- Sofy, M. Application of salicylic acid and zinc improves wheat yield through physiological processes under different levels of irrigation intervals. Int. J. Plant Res. 2015, 5, 136–156. [Google Scholar]
- Sapes, G.; Sala, A. Relative water content consistently predicts drought mortality risk in seedling populations with different morphology, physiology and times to death. Plant Cell Environ. 2021, 44, 3322–3335. [Google Scholar] [CrossRef] [PubMed]
- Schock, A.A.; Ramm, A.; Martinazzo, E.G.; Silva, D.M.; Bacarin, M.A. Crescimento e fotossíntese de plantas de pinhão-manso cultivadas em diferentes condições de luminosidade. Rev. Bras. Eng. Agrícola Ambient. 2014, 18, 3–9. [Google Scholar] [CrossRef]
- San-Eufrasio, B.; Sánchez-Lucas, R.; López-Hidalgo, C.; Guerrero-Sánchez, V.M.; Castillejo, M.Á.; Maldonado-Alconada, A.M.; Jorrín-Novo, J.V.; Rey, M.-D. Responses and Differences in Tolerance to Water Shortage under Climatic Dryness Conditions in Seedlings from Quercus spp. and Andalusian Q. ilex Populations. Forests 2020, 11, 707. [Google Scholar] [CrossRef]
- Silva, P.H.M.; Campoe, O.C.; De Paula, R.C.; Lee, D.J. Seedling Growth and Physiological Responses of Sixteen Eucalypt Taxa under Controlled Water Regime. Forests 2016, 7, 110. [Google Scholar] [CrossRef] [Green Version]
- Vellini, A.L.T.T.; de Paula, N.F.; Alves, P.L.C.A.; Pavani, L.C.; Bonine, C.A.V.; Scarpinati, E.A.; Paula, R.C. de Respostas fisiológicas de diferentes clones de eucalipto sob diferentes regimes de irrigação. Rev. Árvore 2008, 32, 651–663. [Google Scholar] [CrossRef]
- Deng, X.; Xiao, W.; Shi, Z.; Zeng, L.; Lei, L. Combined Effects of Drought and Shading on Growth and Non-Structural Carbohydrates in Pinus massoniana Lamb Seedlings. Forests 2020, 11, 18. [Google Scholar] [CrossRef] [Green Version]
- Ruiz Gómez, F.J.; Pérez-de-Luque, A.; Sánchez-Cuesta, R.; Quero, J.L.; Navarro Cerrillo, R.M. Differences in the Response to Acute Drought and Phytophthora cinnamomi Rands Infection in Quercus ilex L. Seedlings. Forests 2018, 9, 634. [Google Scholar] [CrossRef] [Green Version]
- Zafar, Z.; Rasheed, F.; Atif, R.M.; Maqsood, M.; Gailing, O. Salicylic Acid-Induced Morpho-Physiological and Biochemical Changes Triggered Water Deficit Tolerance in Syzygium cumini L. Saplings. Forests 2021, 12, 491. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Mean Leaf Angles (°) | Leaching Fraction (%) | ||
|---|---|---|---|
| Irrigation Depths (mm) | |||
| 6 | 9 | 12 | |
| 76 | 22.1 ± 3.8 Bc 1 | 34.0 ± 2.2 Bb | 66.8 ± 1.7 Aa |
| 57 | 45.9 ± 0.8 Ab | 47.5 ± 0.7 Ab | 53.0 ± 0.2 Ba |
| 54 | 24.7 ± 1.6 Bb | 42.3 ± 0.5 Aa | 42.9 ± 1.4 Ba |
| −1 | 3.9 ± 0.4 Db | 13.5 ± 4.2 Ca | 16.8 ± 3.8 Ca |
| −44 | 11.3 ± 2.6 Cb | 17.6 ± 1.9 Ca | 19.2 ± 3.8 Ca |
| −64 | 6.5 ± 1.2 Da | 7.2 ± 3.7 Da | 7.3 ± 2.5 Da |
| Mean Leaf Angles (°) | Total Dry Mass (g) | ||
|---|---|---|---|
| Irrigation Depths (mm) | |||
| 6 | 9 | 12 | |
| 76 | 9.85 ± 0.75 Aa 1 | 9.26 ± 0.69 Aa | 7.66 ± 0.55 Bb |
| 57 | 8.52 ± 0.51 Ba | 8.22 ± 0.75 Ba | 7.09 ± 0.34 Cb |
| 54 | 4.49 ± 0.34 Da | 4.13 ± 0.19 Ea | 3.67 ± 0.19 Eb |
| −1 | 2.36 ± 0.50 Fb | 2.54 ± 0.22 Fb | 3.37 ± 0.53 Fa |
| −44 | 5.56 ± 0.54 Cc | 7.07 ± 0.43 Cb | 8.52 ± 0.65 Aa |
| −64 | 3.88 ± 0.34 Eb | 5.87 ± 0.60 Da | 6.43 ± 0.59 Da |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Delgado, L.G.M.; da Silva, R.B.G.; Gabira, M.M.; Rodrigues, A.L.; Simões, D.; de Almeida, L.F.R.; da Silva, M.R. Mean Leaf Angles Affect Irrigation Efficiency and Physiological Responses of Tropical Species Seedling. Forests 2022, 13, 832. https://0-doi-org.brum.beds.ac.uk/10.3390/f13060832
Delgado LGM, da Silva RBG, Gabira MM, Rodrigues AL, Simões D, de Almeida LFR, da Silva MR. Mean Leaf Angles Affect Irrigation Efficiency and Physiological Responses of Tropical Species Seedling. Forests. 2022; 13(6):832. https://0-doi-org.brum.beds.ac.uk/10.3390/f13060832
Chicago/Turabian StyleDelgado, Luiz Gustavo Martinelli, Richardson Barbosa Gomes da Silva, Mônica Moreno Gabira, Angélica Lino Rodrigues, Danilo Simões, Luiz Fernando Rolim de Almeida, and Magali Ribeiro da Silva. 2022. "Mean Leaf Angles Affect Irrigation Efficiency and Physiological Responses of Tropical Species Seedling" Forests 13, no. 6: 832. https://0-doi-org.brum.beds.ac.uk/10.3390/f13060832