
Differential Responses of Soil Extracellular Enzyme Activity and Stoichiometric Ratios under Different Slope Aspects and Slope Positions in Larix olgensis Plantations


Abstract
:1. Introduction
2. Materials and Methods
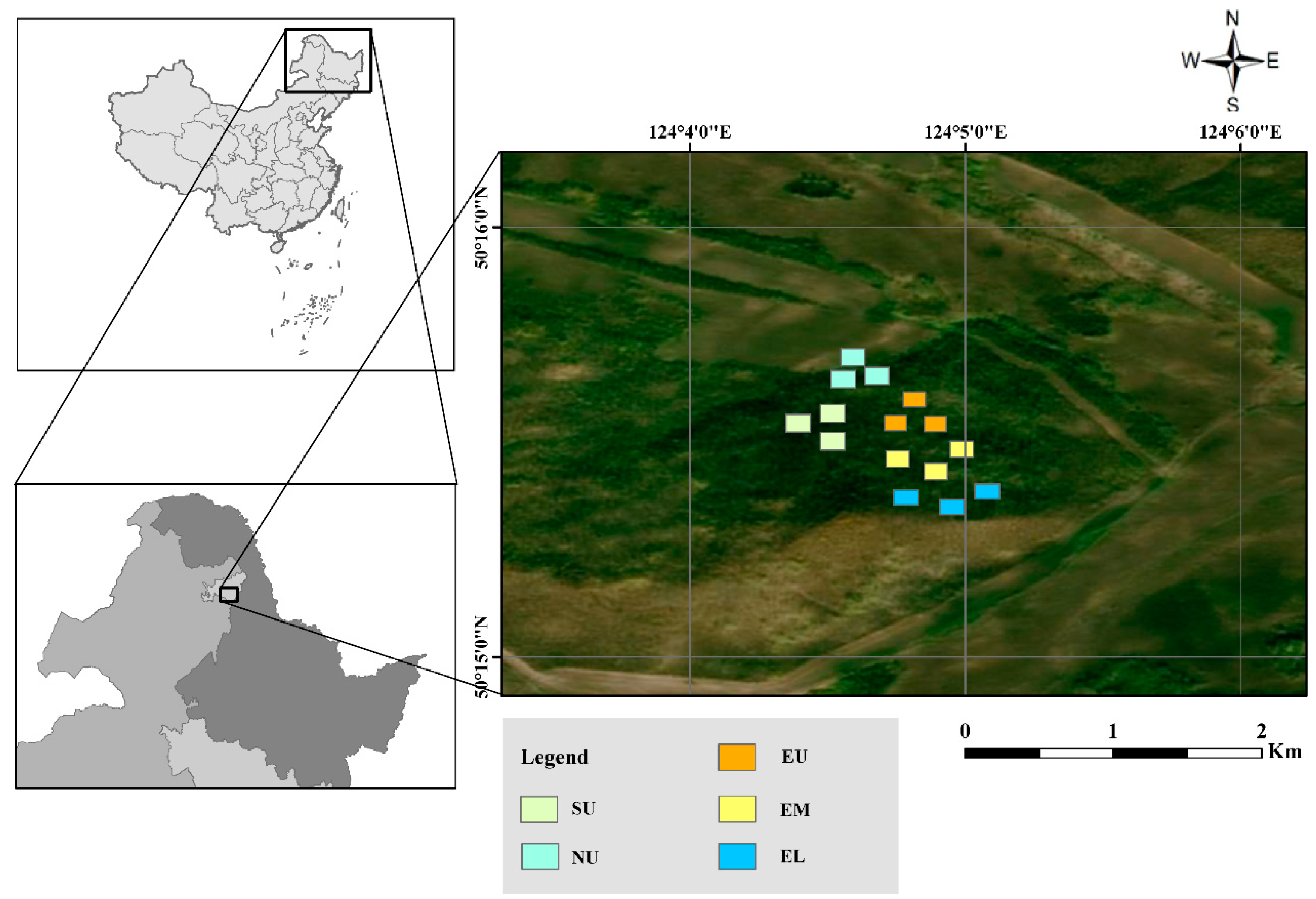
2.1. Study Site
2.2. Experimental Design
2.3. Soil Physicochemical Properties and Soil Enzyme Activity
2.4. Data Analysis
3. Results
3.1. Soil Physicochemical Properties at Different Slope Aspects and Slope Positions
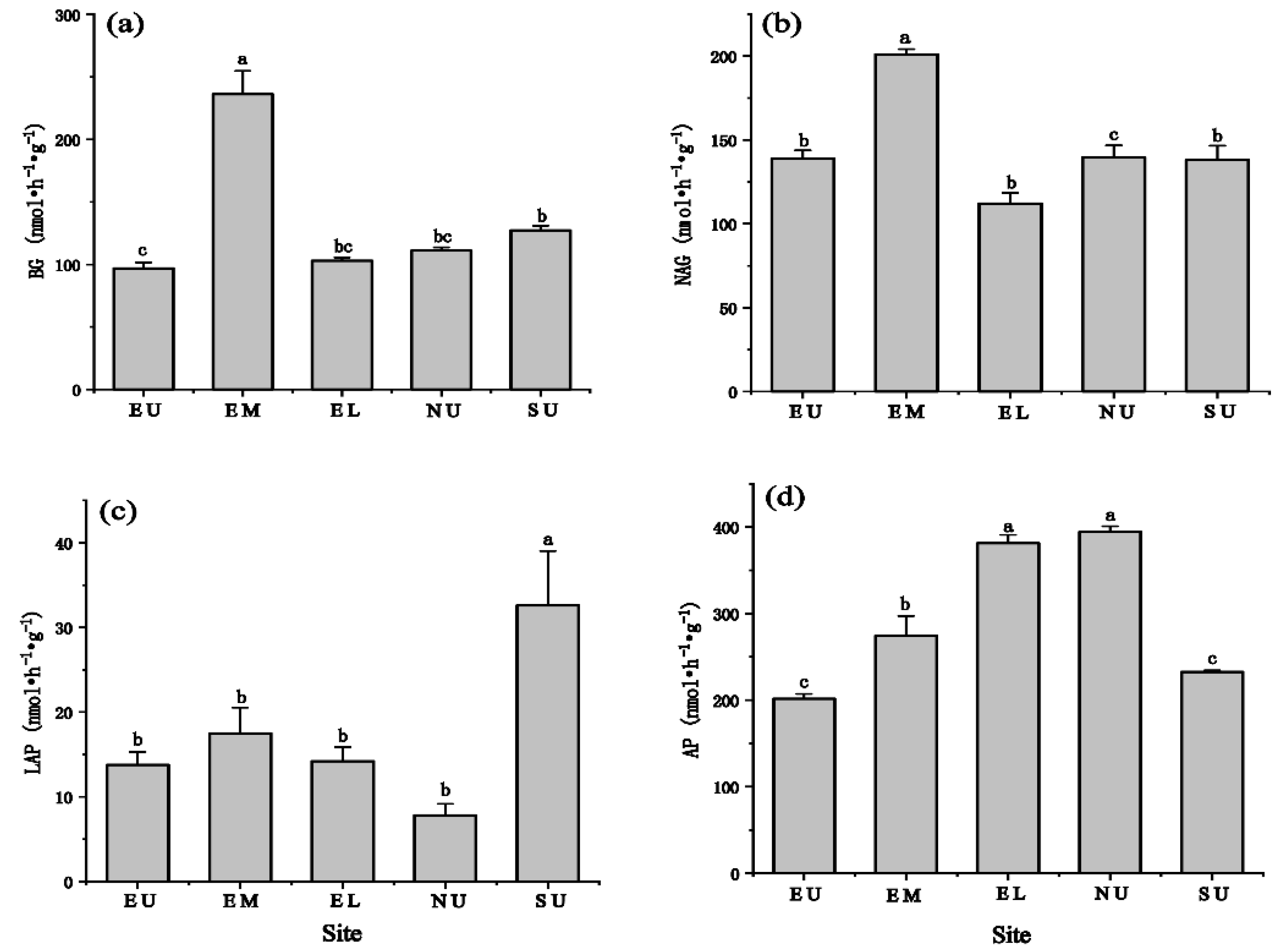
3.2. Characteristics of Soil Extracellular Enzyme Activity Changes at Different Slope Aspects and Slope Positions
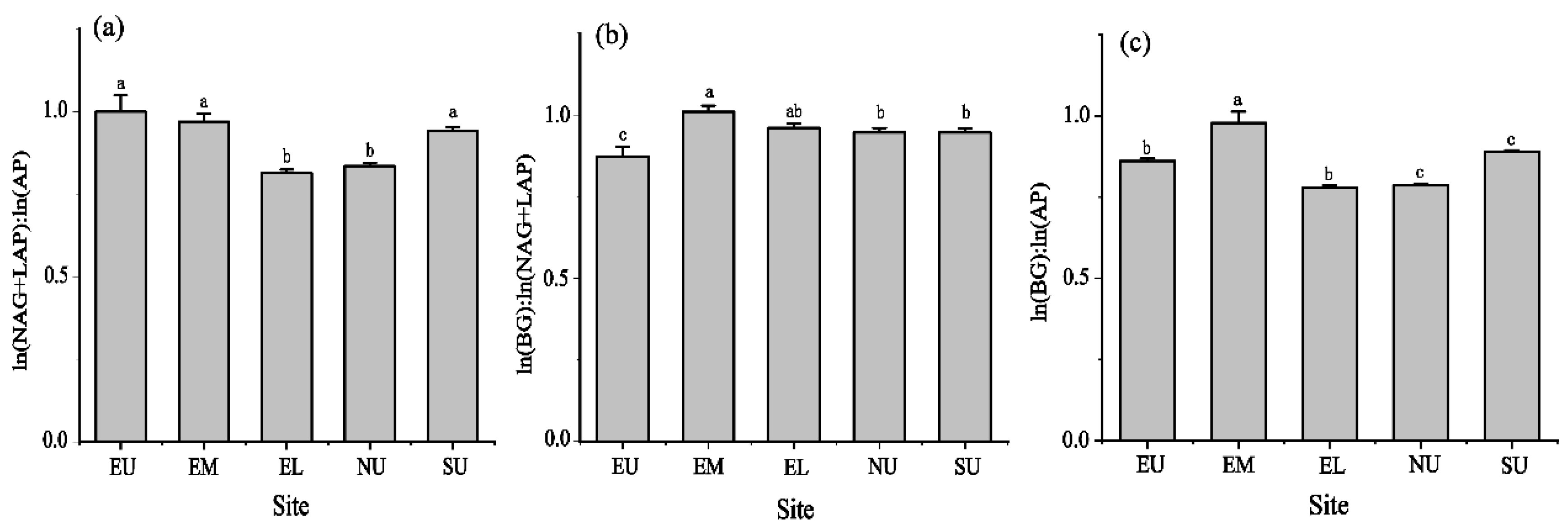
3.3. Soil Extracellular Enzyme Stoichiometry Ratios at Different Slope Aspects and Slope Positions
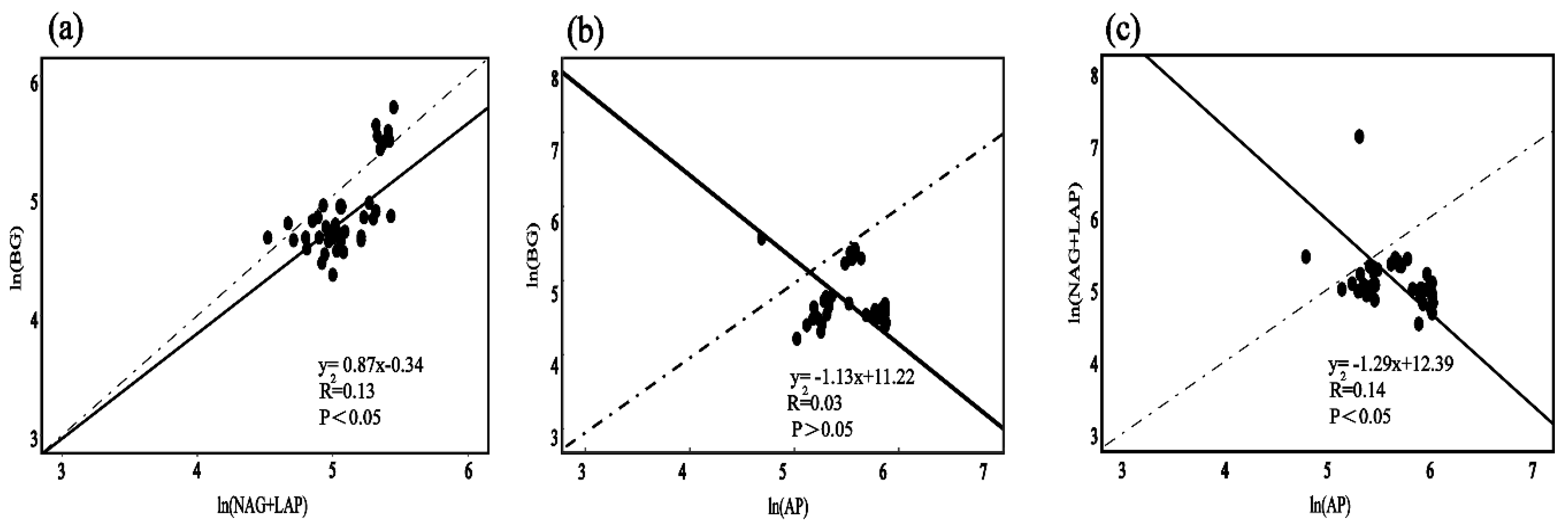
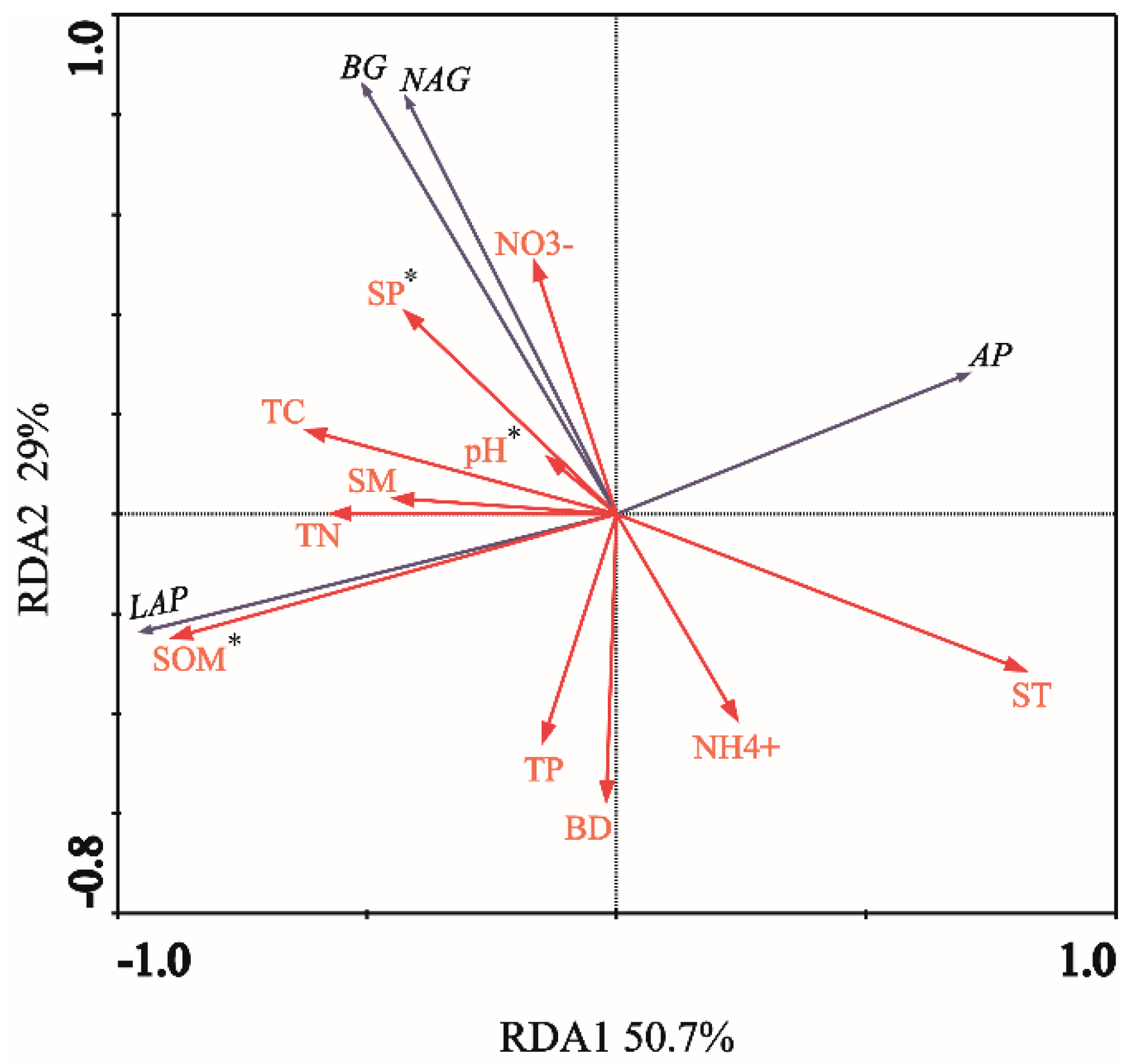
3.4. Relationship between Soil Extracellular Enzyme Activity and Stoichiometric Ratio and Soil Factors
4. Discussion
4.1. Response of Soil Extracellular Enzymes to Slope Aspects and Slope Positions in L. olgensis Plantation Forests
4.2. Soil Enzyme Stoichiometric Ratios Reveal Nutrient Limitation of Microorganisms in L. olgensis Plantation Forests
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Tegegne, Y.T.; Cramm, M.; Brusselen, J.V. Sustainable Forest Management, FLEGT, and REDD+: Exploring Interlinkages to Strengthen Forest Policy Coherence. Sustainability 2018, 10, 4841. [Google Scholar] [CrossRef] [Green Version]
- Lõhmus, P.; Lõhmus, A. The Potential of Production Forests for Sustaining Lichen Diversity: A Perspective on Sustainable Forest Management. Forests 2019, 10, 1063. [Google Scholar] [CrossRef] [Green Version]
- Torres-Rojo, J.M.; Moreno-Sánchez, R.; Mendoza-Briseño, M.A. Sustainable Forest Management in Mexico. Curr. For. Rep. 2016, 2, 93–105. [Google Scholar] [CrossRef] [Green Version]
- Jourgholami, M.; Ramineh, A.; Zahedi Amiri, G.; Labelle, E. The Influence of Slope Positions on the Recovery Response of Compacted Soil Properties and Enzyme Activity in an Oriental Beech Stand in the Hyrcanian Forests, Iran. Sustainability 2019, 11, 1940. [Google Scholar] [CrossRef] [Green Version]
- Li, T.; Liang, J.; Chen, X.; Wang, H.; Zhang, S.; Pu, Y.; Xu, X.; Li, H.; Xu, J.; Wu, X.; et al. The interacting roles and relative importance of climate, topography, soil properties and mineralogical composition on soil potassium variations at a national scale in China. Catena 2021, 196, 104875. [Google Scholar] [CrossRef]
- Zhu, M.; Feng, Q.; Zhang, M.; Liu, W.; Qin, Y.; Deo, R.C.; Zhang, C. Effects of topography on soil organic carbon stocks in grasslands of a semiarid alpine region, northwestern China. J. Soils Sediments 2018, 19, 1640–1650. [Google Scholar] [CrossRef]
- Seibert, J.; Stendahl, J.; Sørensen, R. Topographical influences on soil properties in boreal forests. Geoderma 2007, 141, 139–148. [Google Scholar] [CrossRef]
- Tian, L.; Zhao, L.; Wu, X.; Fang, H.; Zhao, Y.; Yue, G.; Liu, G.; Chen, H. Vertical patterns and controls of soil nutrients in alpine grassland: Implications for nutrient uptake. Sci. Total Environ. 2017, 607–608, 855–864. [Google Scholar] [CrossRef]
- Scholten, T.; Goebes, P.; Kühn, P.; Seitz, S.; Assmann, T.; Bauhus, J.; Bruelheide, H.; Buscot, F.; Erfmeier, A.; Fischer, M.; et al. On the combined effect of soil fertility and topography on tree growth in subtropical forest ecosystems—A study from SE China. J. Plant Ecol. 2017, 10, 111–127. [Google Scholar] [CrossRef]
- Lü, Q.; Yin, H.; He, P.; Li, X.; Wang, Y. Effects of early management of Pinus massoniana plantation target trees on soil physicochemical properties and plant diversity. Chin. J. Appl. Environ. Biol. 2018, 24, 500–507. [Google Scholar]
- Huang, Y.-M.; Liu, D.; An, S.-S. Effects of slope aspect on soil nitrogen and microbial properties in the Chinese Loess region. Catena 2015, 125, 135–145. [Google Scholar] [CrossRef]
- Yang, Y.; Liang, C.; Wang, Y.; Cheng, H.; An, S.; Chang, S.X. Soil extracellular enzyme stoichiometry reflects the shift from P- to N-limitation of microorganisms with grassland restoration. Soil Biol. Biochem. 2020, 149, 107928. [Google Scholar] [CrossRef]
- Finn, D.; Kopittke, P.M.; Dennis, P.G.; Dalal, R.C. Microbial energy and matter transformation in agricultural soils. Soil Biol. Biochem. 2017, 111, 176–192. [Google Scholar] [CrossRef] [Green Version]
- Wang, Q.; Wang, S. Response of labile soil organic matter to changes in forest vegetation in subtropical regions. Appl. Soil Ecol. 2011, 47, 210–216. [Google Scholar] [CrossRef]
- Cui, Y.; Bing, H.; Fang, L.; Jiang, M.; Shen, G.; Yu, J.; Wang, X.; Zhu, H.; Wu, Y.; Zhang, X. Extracellular enzyme stoichiometry reveals the carbon and phosphorus limitations of microbial metabolisms in the rhizosphere and bulk soils in alpine ecosystems. Plant Soil 2019, 458, 7–20. [Google Scholar] [CrossRef]
- Cui, Y.; Fang, L.; Guo, X.; Wang, X.; Zhang, Y.; Li, P.; Zhang, X. Ecoenzymatic stoichiometry and microbial nutrient limitation in rhizosphere soil in the arid area of the northern Loess Plateau, China. Soil Biol. Biochem. 2018, 116, 11–21. [Google Scholar] [CrossRef]
- Rosinger, C.; Rousk, J.; Sandén, H. Can enzymatic stoichiometry be used to determine growth-limiting nutrients for microorganisms?—A critical assessment in two subtropical soils. Soil Biol. Biochem. 2019, 128, 115–126. [Google Scholar] [CrossRef]
- Burns, R.G.; DeForest, J.L.; Marxsen, J.; Sinsabaugh, R.L.; Stromberger, M.E.; Wallenstein, M.D.; Weintraub, M.N.; Zoppini, A. Soil enzymes in a changing environment: Current knowledge and future directions. Soil Biol. Biochem. 2013, 58, 216–234. [Google Scholar] [CrossRef]
- Sinsabaugh, R.L.; Lauber, C.L.; Weintraub, M.N.; Ahmed, B.; Allison, S.D.; Crenshaw, C.; Contosta, A.R.; Cusack, D.; Frey, S.; Gallo, M.E.; et al. Stoichiometry of soil enzyme activity at global scale. Ecol. Lett. 2008, 11, 1252–1264. [Google Scholar] [CrossRef]
- Sinsabaugh, R.L.; Hill, B.H.; Shah, J.J.F. Ecoenzymatic stoichiometry of microbial organic nutrient acquisition in soil and sediment. Nature 2009, 462, 795–798. [Google Scholar] [CrossRef]
- Xiao, W.; Chen, X.; Jing, X.; Zhu, B. A meta-analysis of soil extracellular enzyme activities in response to global change. Soil Biol. Biochem. 2018, 123, 21–32. [Google Scholar] [CrossRef]
- Brockett, B.F.T.; Prescott, C.E.; Grayston, S.J. Soil moisture is the major factor influencing microbial community structure and enzyme activities across seven biogeoclimatic zones in western Canada. Soil Biol. Biochem. 2012, 44, 9–20. [Google Scholar] [CrossRef]
- Xu, Z.; Yu, G.; Zhang, X.; He, N.; Wang, Q.; Wang, S.; Wang, R.; Zhao, N.; Jia, Y.; Wang, C. Soil enzyme activity and stoichiometry in forest ecosystems along the North-South Transect in eastern China (NSTEC). Soil Biol. Biochem. 2017, 104, 152–163. [Google Scholar] [CrossRef]
- Gomez, E.J.; Delgado, J.A.; Gonzalez, J.M. Environmental factors affect the response of microbial extracellular enzyme activity in soils when determined as a function of water availability and temperature. Ecol. Evol. 2020, 10, 10105–10115. [Google Scholar] [CrossRef]
- Souza, L.F.T.; Billings, S.A. Temperature and pH mediate stoichiometric constraints of organically derived soil nutrients. Glob. Chang. Biol. 2022, 28, 1630–1642. [Google Scholar] [CrossRef]
- Puissant, J.; Jones, B.; Goodall, T.; Mang, D.; Blaud, A.; Gweon, H.S.; Malik, A.; Jones, D.L.; Clark, I.M.; Hirsch, P.R.; et al. The pH optimum of soil exoenzymes adapt to long term changes in soil pH. Soil Biol. Biochem. 2019, 138, 107601. [Google Scholar] [CrossRef]
- Fan, B.; Tao, W.; Qin, G.; Hopkins, I.; Zhang, Y.; Wang, Q.; Lin, H.; Guo, L. Soil micro-climate variation in relation to slope aspect, position, and curvature in a forested catchment. Agric. For. Meteorol. 2020, 290, 107999. [Google Scholar] [CrossRef]
- Zhou, L.; Liu, S.; Shen, H.; Zhao, M.; Xu, L.; Xing, A.; Fang, J. Soil extracellular enzyme activity and stoichiometry in China’s forests. Funct. Ecol. 2020, 34, 1461–1471. [Google Scholar] [CrossRef]
- Gianfreda, L.; Rao, M.A. The influence of pesticides on soil enzymes. In Soil Enzymology; Springer: Berlin/Heidelberg, Germany, 2010; pp. 293–312. [Google Scholar]
- Waring, B.G.; Weintraub, S.R.; Sinsabaugh, R.L. Ecoenzymatic stoichiometry of microbial nutrient acquisition in tropical soils. Biogeochemistry 2013, 117, 101–113. [Google Scholar] [CrossRef]
- Peng, X.; Wang, W. Stoichiometry of soil extracellular enzyme activity along a climatic transect in temperate grasslands of northern China. Soil Biol. Biochem. 2016, 98, 74–84. [Google Scholar] [CrossRef]
- Moorhead, D.L.; Sinsabaugh, R.L.; Hill, B.H.; Weintraub, M.N. Vector analysis of ecoenzyme activities reveal constraints on coupled C, N and P dynamics. Soil Biol. Biochem. 2016, 93, 1–7. [Google Scholar] [CrossRef]
- Hill, B.H.; Elonen, C.M.; Jicha, T.M.; Kolka, R.K.; Lehto, L.R.L.P.; Sebestyen, S.D.; Seifert-Monson, L.R. Ecoenzymatic stoichiometry and microbial processing of organic matter in northern bogs and fens reveals a common P-limitation between peatland types. Biogeochemistry 2014, 120, 203–224. [Google Scholar] [CrossRef]
- Burke, D.J.; Weintraub, M.N.; Hewins, C.R.; Kalisz, S. Relationship between soil enzyme activities, nutrient cycling and soil fungal communities in a northern hardwood forest. Soil Biol. Biochem. 2011, 43, 795–803. [Google Scholar] [CrossRef]
- Cui, Y.; Moorhead, D.L.; Wang, X.; Xu, M.; Wang, X.; Wei, X.; Zhu, Z.; Ge, T.; Peng, S.; Zhu, B.; et al. Decreasing microbial phosphorus limitation increases soil carbon release. Geoderma 2022, 419, 115868. [Google Scholar] [CrossRef]
- Yang, Y.; Wu, J.; Zhao, S.; Mao, Y.; Zhang, J.; Pan, X.; He, F.; Ploeg, M. Impact of long-term sub-soiling tillage on soil porosity and soil physical properties in the soil profile. Land Degrad. Dev. 2021, 32, 2892–2905. [Google Scholar] [CrossRef]
- Storer, D.A. A simple high sample volume ashing procedure for determination of soil organic matter. Commun. Soil Sci. Plant Anal. 2008, 15, 759–772. [Google Scholar] [CrossRef]
- German, D.P.; Weintraub, M.N.; Grandy, A.S.; Lauber, C.L.; Rinkes, Z.L.; Allison, S.D. Optimization of hydrolytic and oxidative enzyme methods for ecosystem studies. Soil Biol. Biochem. 2011, 43, 1387–1397. [Google Scholar] [CrossRef]
- Chang, E.-H.; Chen, T.-H.; Tian, G.; Chiu, C.-Y. The effect of altitudinal gradient on soil microbial community activity and structure in moso bamboo plantations. Appl. Soil Ecol. 2016, 98, 213–220. [Google Scholar] [CrossRef]
- Salehi, M.H.; Esfandiarpour, I.; Sarshogh, M. The Effect of Aspect on Soil Spatial Variability in Central Zagros, Iran. Procedia Environ. Sci. 2011, 7, 293–298. [Google Scholar] [CrossRef] [Green Version]
- Liu, J.; Qiu, L.; Wang, X.; Wei, X.; Gao, H.; Zhang, Y.; Cheng, J. Effects of wildfire and topography on soil nutrients in a semiarid restored grassland. Plant Soil 2018, 428, 123–136. [Google Scholar] [CrossRef]
- Fu, B.J.; Liu, S.L.; Ma, K.M.; Zhu, Y.G. Relationships between soil characteristics, topography and plant diversity in a heterogeneous deciduous broad-leaved forest near Beijing, China. Plant Soil 2004, 261, 47–54. [Google Scholar] [CrossRef]
- Ji, L.; Yu, J.; Zhang, X.; Liu, Y.; Yang, L. Differential Responses of Soil Bacterial and Fungal Community to Short-Term Crop Tree Management in a Larix gmelinii Plantation. Forests 2021, 12, 1411. [Google Scholar] [CrossRef]
- Zheng, T.; Xie, H.; Thompson, G.L.; Bao, X.; Deng, F.; Yan, E.; Zhou, X.; Liang, C. Shifts in microbial metabolic pathway for soil carbon accumulation along subtropical forest succession. Soil Biol. Biochem. 2021, 160, 108335. [Google Scholar] [CrossRef]
- DeForest, J.L.; Moorhead, D.L. Effects of elevated pH and phosphorus fertilizer on soil C, N and P enzyme stoichiometry in an acidic mixed mesophytic deciduous forest. Soil Biol. Biochem. 2020, 150, 107996. [Google Scholar] [CrossRef]
- Lyu, M.; Nie, Y.; Giardina, C.P.; Vadeboncoeur, M.A.; Ren, Y.; Fu, Z.; Wang, M.; Jin, C.; Liu, X.; Xie, J. Litter quality and site characteristics interact to affect the response of priming effect to temperature in subtropical forests. Funct. Ecol. 2019, 33, 2226–2238. [Google Scholar] [CrossRef]
- Xu, Z.; Yu, G.; Zhang, X.; Ge, J.; He, N.; Wang, Q.; Wang, D. The variations in soil microbial communities, enzyme activities and their relationships with soil organic matter decomposition along the northern slope of Changbai Mountain. Appl. Soil Ecol. 2015, 86, 19–29. [Google Scholar] [CrossRef]
- Zuo, Y.; Zhang, H.; Li, J.; Yao, X.; Chen, X.; Zeng, H.; Wang, W. The effect of soil depth on temperature sensitivity of extracellular enzyme activity decreased with elevation: Evidence from mountain grassland belts. Sci. Total Environ. 2021, 777, 146136. [Google Scholar] [CrossRef]
- Zhu, X.; Liu, M.; Kou, Y.; Liu, D.; Liu, Q.; Zhang, Z.; Jiang, Z.; Yin, H. Differential effects of N addition on the stoichiometry of microbes and extracellular enzymes in the rhizosphere and bulk soils of an alpine shrubland. Plant Soil 2020, 449, 285–301. [Google Scholar] [CrossRef]
- Jing, X.; Wang, Y.; Chung, H.; Mi, Z.; Wang, S.; Zeng, H.; He, J.-S. No temperature acclimation of soil extracellular enzymes to experimental warming in an alpine grassland ecosystem on the Tibetan Plateau. Biogeochemistry 2013, 117, 39–54. [Google Scholar] [CrossRef]
- Yan, D. Dynamics of soil nutrients in larch plantations. J. For. Res. 1999, 10, 239–242. [Google Scholar]
- He, Q.; Wu, Y.; Bing, H.; Zhou, J.; Wang, J. Vegetation type rather than climate modulates the variation in soil enzyme activities and stoichiometry in subalpine forests in the eastern Tibetan Plateau. Geoderma 2020, 374, 114424. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sample Site | Altitude (m) | Crown Density | Soil Thickness (cm) | Diameter (cm) | Height (m) | Basal Area (cm)2 |
|---|---|---|---|---|---|---|
| EL | 417 | 0.50 | 70 | 16.80 ± 0.52 | 11.56 ± 0.16 | 221.56 ± 0.62 |
| EL | 418 | 0.50 | 70 | 15.82 ± 0.92 | 11.85 ± 0.24 | 196.46 ± 1.02 |
| EL | 420 | 0.50 | 75 | 16.28 ± 0.44 | 13.25 ± 0.18 | 208.06 ± 0.54 |
| EM | 425 | 0.60 | 47 | 16.62 ± 1.17 | 14.27 ± 1.10 | 216.84 ± 1.27 |
| EM | 423 | 0.65 | 48 | 15.48 ± 0.37 | 12.07 ± 0.20 | 188.11 ± 0.47 |
| EM | 422 | 0.60 | 49 | 14.97 ± 0.38 | 12.54 ± 0.17 | 175.92 ± 0.48 |
| EU | 428 | 0.60 | 36 | 14.90 ± 0.41 | 12.48 ± 0.17 | 174.28 ± 0.51 |
| EU | 429 | 0.55 | 34 | 13.43 ± 0.54 | 12.45 ± 1.25 | 141.59 ± 0.64 |
| EU | 428 | 0.60 | 33 | 13.67 ± 0.49 | 13.18 ± 1.16 | 146.69 ± 0.59 |
| NU | 418 | 0.60 | 32 | 12.70 ± 0.36 | 11.77 ± 0.19 | 123.84 ± 0.69 |
| NU | 418 | 0.60 | 34 | 14.67 ± 0.37 | 13.70 ± 0.90 | 163.68 ± 0.68 |
| NU | 418 | 0.60 | 32 | 12.49 ± 0.34 | 11.96 ± 0.18 | 121.87 ± 0.59 |
| SU | 429 | 0.70 | 42 | 15.14 ± 0.59 | 12.31 ± 0.24 | 177.57 ± 0.46 |
| SU | 430 | 0.70 | 41 | 15.99 ± 0.58 | 12.10 ± 0.22 | 200.71 ± 0.47 |
| SU | 430 | 0.70 | 43 | 14.97 ± 0.49 | 12.27 ± 0.21 | 175.92 ± 0.44 |
| Enzyme | EC | Substrate Proxy | Substrate Concentration | Function |
|---|---|---|---|---|
| L-leucine aminopeptidase (LAP) | EC 3.4.11.1 | L–Leucine–7-amino–4–methylcoumarin | 200 μmol·L−1 | Catalyze the cleavage of amino acids from proteins or other peptide substrates. |
| β-glucosidase (BG) | EC 3.2.1.21 | 4–MUB–b–d–glucoside | 200 μmol·L−1 | Decomposition of readily degradable carbon and release of glucose from cellulose. |
| Acid phosphatase (AP) | EC 3.1.3.2 | 4–MUB–phosphate | 200 μmol·L−1 | Release of phosphate from Hydrolyzed phosphoglycans and phospholipids. |
| β-1,4-N-acetylglucosaminidase (NAG) | EC 3.2.1.61 | 4–MUB–N-acetyl–β–d–glucosaminide | 200 μmol·L−1 | Hydrolyze N-acetylglucosamine (an amino sugar) in chitin. |
| Site | SM % | BD g·cm3 | SP % | ST ℃ | pH | TC g·kg−1 | TN g·kg−1 | TP g·kg−1 | SOM g·kg−1 | NH4+-N mg·kg−1 | NO3−-N mg·kg−1 |
|---|---|---|---|---|---|---|---|---|---|---|---|
| EU | 55.23 ± 3.84 ab | 0.87 ± 0.09 b | 170.52 ± 15.33 a | 15.43 ± 0.52 a | 5.69 ± 0.11 a | 86.61 ± 5.59 b | 3.84 ± 0.32 a | 0.90 ± 0.19 a | 50.49 ± 4.46 a | 211.80 ± 9.88 a | 0.83 ± 0.06 b |
| EM | 61.95 ± 5.33 ab | 0.84 ± 0.08 b | 173.84 ± 15.87 a | 13.80 ± 0.36 ab | 5.73 ± 0.19 a | 104.20 ± 6.37 a | 4.85 ± 0.62 a | 0.75 ± 0.04 a | 67.81 ± 8.82 a | 157.45 ± 1.76 b | 1.96 ± 0.29 a |
| EL | 43.03 ± 3.57 b | 1.20 ± 0.05 a | 117.85 ± 6.75 b | 14.87 ± 0.87 ab | 5.76 ± 0.10 a | 109.69 ± 5.32 a | 5.20 ± 0.77 a | 0.75 ± 0.18 a | 66.93 ± 14.64 a | 155.3 ± 21.15 b | 1.89 ± 0.22 a |
| SU | 69.62 ± 11.89 ab | 1.00 ± 0.05 ab | 148.61 ± 9.34 ab | 15.13 ± 0.58 ab | 5.35 ± 0.14 a | 85.38 ± 6.68 b | 5.09 ± 0.71 a | 0.96 ± 0.06 a | 71.76 ± 9.82 a | 208.50 ± 15.32 a | 0.67 ± 0.05 b |
| NU | 85.02 ± 16.49 a | 1.18 ± 0.06 a | 153.32 ± 14.7 ab | 13.50 ± 0.30 b | 5.41 ± 0.14 a | 119.95 ± 6.46 a | 3.81 ± 0.74 a | 0.73 ± 0.16 a | 55.67 ± 13.04 a | 193.12 ± 11.66 ab | 0.78 ± 0.08 b |
| F | 2.63 * | 6.05 ** | 2.99 * | 2.28 ns | 1.88 ns | 0.70 ns | 1.08 ns | 0.54 ns | 6.01 ** | 4.05 ** | 15.20 ** |
| Site | VL | VA |
|---|---|---|
| EU | 1.23 ± 0.02 c | 45.25 ± 1.24 b |
| EM | 1.41 ± 0.03 a | 45.97 ± 0.67 b |
| EL | 1.24 ± 0.01 c | 50.91 ± 0.36 a |
| SU | 1.23 ± 0.01 c | 50.18 ± 0.35 a |
| NU | 1.30 ± 0.01 b | 46.76 ± 0.36 b |
| Soil Factor | BG | AP | NAG | LAP | C:Neea | C:Peea | N:Peea |
|---|---|---|---|---|---|---|---|
| Total nitrogen | 0.04 | 0.54 * | 0.02 | −0.04 | 0.05 | −0.22 | −0.22 |
| Total carbon | 0.08 | 0.513 * | 0.06 | −0.08 | 0.03 | −0.15 | −0.13 |
| Total phosphorus | 0.19 | 0.35 | −0.13 | 0.07 | 0.23 | 0.06 | −0.20 |
| NH4+-N | 0.44 | 0.49 | −0.28 | 0.08 | 0.52 * | 0.21 | −0.37 |
| NO3−-N | −0.44 | −0.31 | 0.50 | 0.01 | −0.60 * | −0.30 | 0.43 |
| Soil organic matter | 0.18 | 0.63 ** | −0.08 | −0.06 | 0.22 | −0.12 | −0.32 |
| Soil temperature | −0.23 | −0.21 | 0.01 | −0.22 | −0.18 | −0.22 | −0.01 |
| Soil moisture | 0.55* | 0.09 | −0.18 | 0.19 | 0.51 * | 0.53 * | −0.06 |
| Soil bulk density | 0.20 | 0.41 | 0.41 | −0.05 | −0.12 | −0.01 | 0.21 |
| Soil porosity | 0.01 | −0.2 | −0.38 | −0.17 | 0.27 | 0.09 | −0.27 |
| pH | −0.69 ** | −0.3 | 0.14 | 0.07 | −0.62** | −0.57 * | 0.12 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, M.; Ji, L.; Shen, F.; Meng, J.; Wang, J.; Shan, C.; Yang, L. Differential Responses of Soil Extracellular Enzyme Activity and Stoichiometric Ratios under Different Slope Aspects and Slope Positions in Larix olgensis Plantations. Forests 2022, 13, 845. https://0-doi-org.brum.beds.ac.uk/10.3390/f13060845
Wang M, Ji L, Shen F, Meng J, Wang J, Shan C, Yang L. Differential Responses of Soil Extracellular Enzyme Activity and Stoichiometric Ratios under Different Slope Aspects and Slope Positions in Larix olgensis Plantations. Forests. 2022; 13(6):845. https://0-doi-org.brum.beds.ac.uk/10.3390/f13060845
Chicago/Turabian StyleWang, Mingwei, Li Ji, Fangyuan Shen, Jun Meng, Junlu Wang, Chengfeng Shan, and Lixue Yang. 2022. "Differential Responses of Soil Extracellular Enzyme Activity and Stoichiometric Ratios under Different Slope Aspects and Slope Positions in Larix olgensis Plantations" Forests 13, no. 6: 845. https://0-doi-org.brum.beds.ac.uk/10.3390/f13060845