
Geomorphic and Climatic Drivers Are Key Determinants of Structural Variability of Mangrove Forests along the Kenyan Coast

 , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
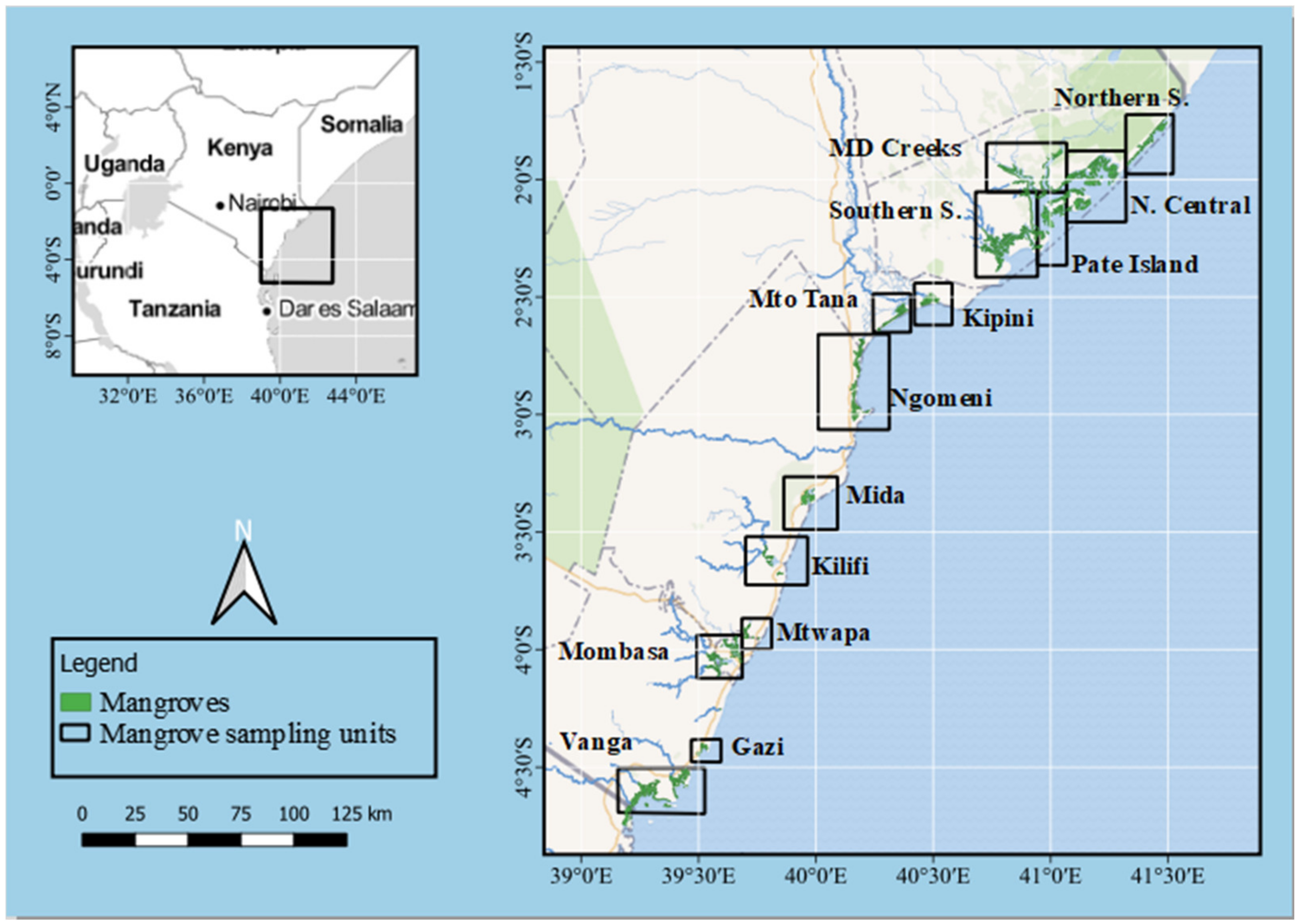
2.1. Description of the Study Area
2.1.1. Climate
2.1.2. Oceanography
2.1.3. Hydrology
2.1.4. Geomorphology
2.1.5. Demography
2.2. Assessment of Vegetation Structure
2.3. Factors Affecting Mangrove Structural Variability
2.4. Data Analysis
3. Results
3.1. Species Composition
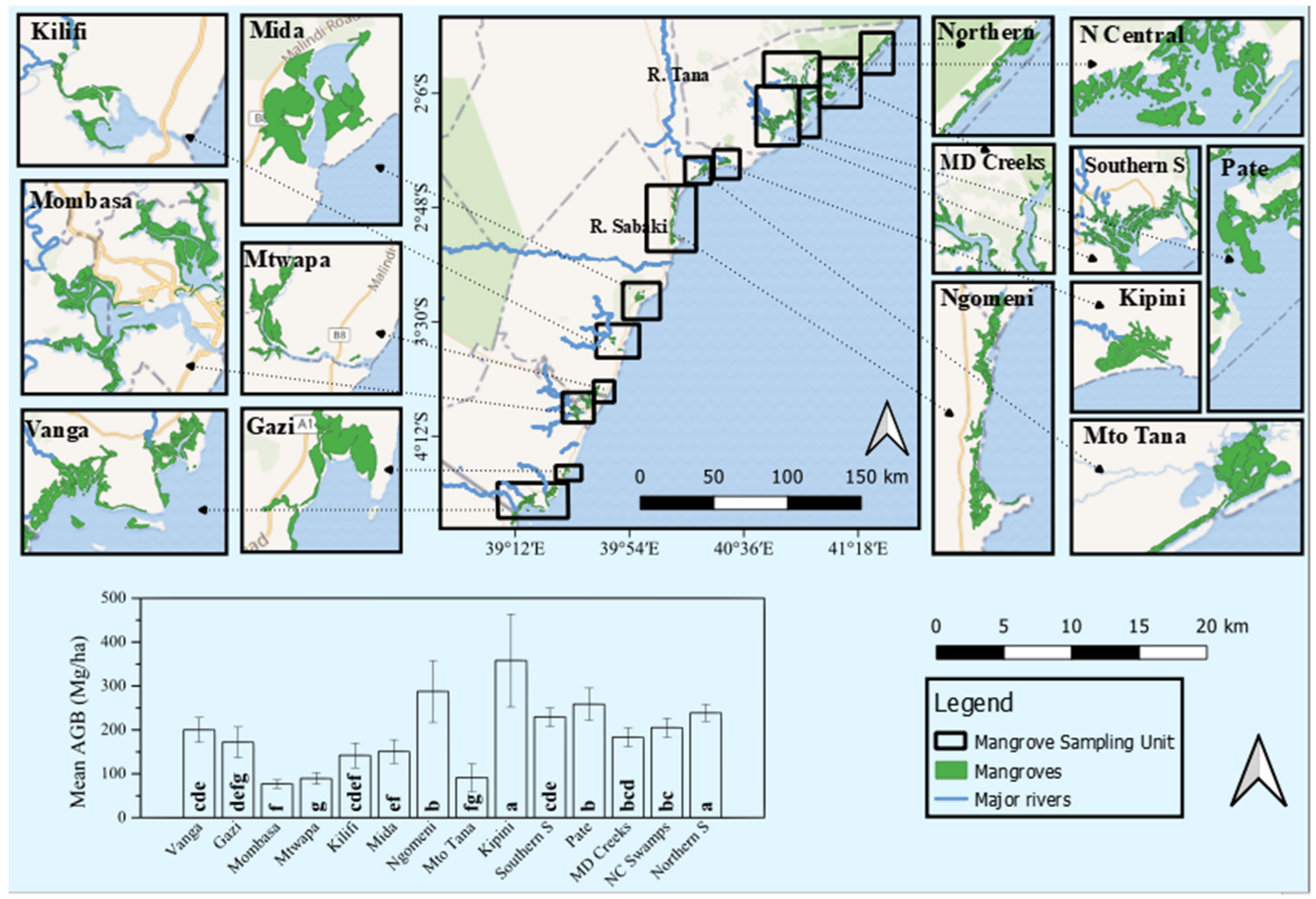
3.2. Mangrove Structure and Diversity across the Sites
3.3. Multivariate Analysis
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Site | Species | Mean DBH (cm) | SE | Mean Height (m) | SE | Basal Area (m2/ha) | IV |
|---|---|---|---|---|---|---|---|
| Vanga | Avicennia marina | 7.71 | 0.39 | 4.51 | 0.18 | 132.97 | 64.24 |
| Bruguiera gymnorhiza | 6.33 | 0.25 | 5.02 | 0.12 | 44.74 | 49 | |
| Ceriops tagal | 4.63 | 0.07 | 3.39 | 0.05 | 96.47 | 106.92 | |
| Lumnitzera racemosa | 3.4 | 0.35 | 1.43 | 0.07 | 0.07 | 1.95 | |
| Rhizophora mucronata | 7.63 | 0.13 | 6.06 | 0.07 | 612.86 | 187.68 | |
| Sonneratia alba | 18.32 | 0.99 | 10.36 | 0.45 | 129.58 | 34.04 | |
| Xylocarpus granatum | 7.18 | 0.34 | 5.01 | 0.15 | 30.78 | 29.75 | |
| Gazi | Avicennia marina | 16.41 | 2.06 | 8.81 | 0.73 | 15.81 | 35.87 |
| Bruguiera gymnorhiza | 7.94 | 0.46 | 5.11 | 0.26 | 31.54 | 87.11 | |
| Ceriops tagal | 4.22 | 0.08 | 2.75 | 0.05 | 30.3 | 130.4 | |
| Rhizophora mucronata | 7.6 | 0.48 | 6.53 | 0.33 | 98.15 | 145.44 | |
| Sonneratia alba | 11.41 | 1.14 | 6.28 | 0.37 | 22.25 | 24.04 | |
| Xylocarpus granatum | 8.65 | 0.84 | 5.24 | 0.35 | 30.15 | 68.05 | |
| Mombasa | Avicennia marina | 10.55 | 0.39 | 5.69 | 0.13 | 178.44 | 78.99 |
| Bruguiera gymnorhiza | 4.36 | 0.48 | 3.41 | 0.22 | 0.46 | 2.85 | |
| Ceriops tagal | 4.57 | 0.13 | 2.79 | 0.05 | 66.74 | 71.64 | |
| Lumnitzera racemosa | 7.55 | 0.25 | 3.2 | 0.4 | 0.22 | 1.37 | |
| Rhizophora mucronata | 4.57 | 0.05 | 4.07 | 0.03 | 277.84 | 196.97 | |
| Sonneratia alba | 11.85 | 0.33 | 7.11 | 0.13 | 145.23 | 45.24 | |
| Xylocarpus granatum | 4.68 | 0.67 | 3.52 | 0.51 | 0.63 | 2.89 | |
| Xylocarpus moluccensis | 6.5 | 0.5 | 7.45 | 0.05 | 0.17 | 1.36 | |
| Mtwapa | Avicennia marina | 11.65 | 1.37 | 5.03 | 0.36 | 5.15 | 16.16 |
| Ceriops tagal | 3.73 | 0.05 | 2.08 | 0.03 | 27.6 | 98.91 | |
| Rhizophora mucronata | 4.42 | 0.07 | 3.5 | 0.03 | 91.23 | 236.7 | |
| Xylocarpus granatum | 5.74 | 0.56 | 4.02 | 0.17 | 2.47 | 14.89 | |
| Kilifi | Avicennia marina | 9.93 | 0.5 | 5.52 | 0.28 | 81.14 | 108.87 |
| Bruguiera gymnorhiza | 4.09 | 0.32 | 3.15 | 0.21 | 1.28 | 23.76 | |
| Ceriops tagal | 3.77 | 0.06 | 2.08 | 0.04 | 17.84 | 81.18 | |
| Rhizophora mucronata | 6.49 | 0.29 | 4.43 | 0.12 | 103.23 | 157.68 | |
| Sonneratia alba | 8.67 | 0.66 | 4.89 | 0.21 | 16.44 | 28.5 | |
| Mida | Avicennia marina | 11.37 | 0.98 | 5.61 | 0.22 | 13.95 | 53.86 |
| Bruguiera gymnorhiza | 11.93 | 1.76 | 7.08 | 0.58 | 6.83 | 53.61 | |
| Ceriops tagal | 4.29 | 0.08 | 3.87 | 0.06 | 7.34 | 156.59 | |
| Rhizophora mucronata | 7.27 | 0.25 | 6.27 | 0.12 | 7.87 | 141.2 | |
| Ngomeni | Avicennia marina | 11.36 | 0.78 | 5.46 | 0.21 | 18.45 | 89.9 |
| Bruguiera gymnorhiza | 12.2 | 1.13 | 7.43 | 0.43 | 6.15 | 45.83 | |
| Ceriops tagal | 6.07 | 0.28 | 4.86 | 0.13 | 4.5 | 116.14 | |
| Rhizophora mucronata | 6.99 | 0.27 | 6.55 | 0.11 | 11.32 | 150.72 | |
| Sonneratia alba | 11.92 | 0.96 | 6.64 | 0.27 | 13.55 | 28.5 | |
| Xylocarpus granatum | 18.54 | 1.42 | 7.81 | 0.35 | 12.77 | 32.07 | |
| Mto Tana | Avicennia marina | 7.11 | 0.25 | 4.28 | 0.17 | 50.79 | 210.19 |
| Bruguiera gymnorhiza | 9.61 | 1.8 | 8.09 | 1.24 | 2.38 | 32.86 | |
| Ceriops tagal | 4.79 | 0.17 | 3.85 | 0.12 | 26.19 | 184.45 | |
| Rhizophora mucronata | 7.03 | 1.42 | 8 | 1.27 | 0.44 | 15.35 | |
| Kipini | Avicennia marina | 7.75 | 0.23 | 6.61 | 0.14 | 105.1 | 158.12 |
| Bruguiera gymnorhiza | 16.59 | 2.55 | 10.59 | 1.18 | 26.01 | 47.35 | |
| Ceriops tagal | 3.55 | 0.19 | 4.33 | 0.14 | 0.5 | 11.24 | |
| Heritiera littoralis | 13.85 | 0.96 | 9.11 | 0.4 | 151.03 | 112.99 | |
| Xylocarpus granatum | 10.32 | 1.02 | 6.96 | 0.34 | 44.91 | 61.21 | |
| Southern Swamps | Avicennia marina | 6.89 | 0.31 | 3.59 | 0.13 | 31.48 | 25.15 |
| Bruguiera gymnorhiza | 8.13 | 0.53 | 4.85 | 0.12 | 68.44 | 60.47 | |
| Ceriops tagal | 4.76 | 0.09 | 3.21 | 0.04 | 97.36 | 100.74 | |
| Rhizophora mucronata | 8.02 | 0.11 | 5.81 | 0.04 | 735.87 | 234.07 | |
| Sonneratia alba | 12.4 | 0.75 | 7.69 | 0.25 | 42.75 | 19.56 | |
| Pate Island | Avicennia marina | 12.64 | 0.83 | 10.76 | 0.35 | 30.59 | 35.35 |
| Bruguiera gymnorhiza | 4.89 | 0.14 | 7.32 | 0.09 | 13.58 | 50.98 | |
| Ceriops tagal | 4.49 | 0.29 | 7.21 | 0.21 | 2.39 | 40.23 | |
| Rhizophora mucronata | 9.18 | 0.27 | 9.99 | 0.12 | 190.65 | 199.72 | |
| Sonneratia alba | 16.34 | 0.8 | 12.2 | 0.26 | 100.24 | 73.63 | |
| Xylocarpus granatum | 14.94 | 1.2 | 10.76 | 0.43 | 36.32 | 21.51 | |
| Mongoni-Dodori Creek | Avicennia marina | 6.4 | 0.7 | 7.08 | 0.36 | 32.67 | 40.5 |
| Bruguiera gymnorhiza | 7.17 | 0.96 | 8.75 | 0.4 | 7.58 | 43.1 | |
| Ceriops tagal | 6.67 | 0.23 | 8.91 | 0.1 | 56.28 | 105.9 | |
| Rhizophora mucronata | 7.05 | 0.28 | 9.7 | 0.11 | 107.8 | 147.33 | |
| Sonneratia alba | 14.4 | 0.59 | 12 | 0.22 | 64.22 | 49.9 | |
| Xylocarpus granatum | 12.7 | 1.03 | 11.31 | 0.36 | 26.41 | 32.03 | |
| Northern Central Swamps | Avicennia marina | 10.35 | 1.08 | 9.76 | 0.26 | 61.81 | 24.15 |
| Bruguiera gymnorhiza | 10.65 | 1.03 | 12.08 | 0.3 | 28.87 | 33.94 | |
| Ceriops tagal | 5.24 | 0.08 | 10.03 | 0.05 | 106.78 | 103.71 | |
| Rhizophora mucronata | 8.14 | 0.16 | 12.52 | 0.09 | 467.67 | 204.26 | |
| Sonneratia alba | 15.48 | 0.75 | 14.52 | 0.25 | 92.95 | 31.38 | |
| Northern Swamps | Avicennia marina | 9.64 | 0.65 | 4.78 | 0.17 | 24.71 | 19.93 |
| Bruguiera gymnorhiza | 7.73 | 0.98 | 5.91 | 0.47 | 6.6 | 15.4 | |
| Ceriops tagal | 6.2 | 0.18 | 3.75 | 0.12 | 35.43 | 53.12 | |
| Rhizophora mucronata | 11.6 | 0.19 | 9.09 | 0.1 | 653.74 | 239.98 | |
| Sonneratia alba | 14.13 | 0.49 | 7.53 | 0.19 | 137.1 | 50.52 |
| Vanga | Gazi | Mombasa | Mtwapa | Kilifi | Mida | Ngomeni | Mto Tana | Kipini | Southern Swamps | Pate | Mongoni-Dodori Creeks | Northern Central | Northern Swamps | |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Vanga | 0.0217 | 0.0845 | 0.0009 | 0.0104 | 0.0011 | 0.0321 | 0.0002 | 0.0001 | 0.1558 | 0.2616 | 0.0902 | 0.1681 | 0.014 | |
| Gazi | 0.0217 | 0.0008 | 0.0241 | 0.009 | 0.0148 | 0.1242 | 0.0007 | 0.0004 | 0.0196 | 0.0517 | 0.0639 | 0.0222 | 0.0004 | |
| Mombasa | 0.0845 | 0.0008 | 0.0007 | 0.0284 | 0.0013 | 0.0004 | 0.0011 | 0.0002 | 0.0314 | 0.0047 | 0.0075 | 0.0482 | 0.0648 | |
| Mtwapa | 0.0009 | 0.0241 | 0.0007 | 0.0037 | 0.062 | 0.0066 | 0.0168 | 0.0002 | 0.0013 | 0.0009 | 0.0076 | 0.0058 | 0.0004 | |
| Kilifi | 0.0104 | 0.009 | 0.0284 | 0.0037 | 0.0192 | 0.0765 | 0.0951 | 0.0005 | 0.0203 | 0.006 | 0.0589 | 0.0418 | 0.0091 | |
| Mida | 0.0011 | 0.0148 | 0.0013 | 0.062 | 0.0192 | 0.0306 | 0.0999 | 0.0003 | 0.0131 | 0.0004 | 0.0021 | 0.0252 | 0.0005 | |
| Ngomeni | 0.0321 | 0.1242 | 0.0004 | 0.0066 | 0.0765 | 0.0306 | 0.0263 | 0.0002 | 0.0259 | 0.0088 | 0.0602 | 0.0169 | 0.0008 | |
| Mto Tana | 0.0002 | 0.0007 | 0.0011 | 0.0168 | 0.0951 | 0.0999 | 0.0263 | 0.0001 | 0.001 | 0.0001 | 0.0034 | 0.0013 | 0.0003 | |
| Kipini | 0.0001 | 0.0004 | 0.0002 | 0.0002 | 0.0005 | 0.0003 | 0.0002 | 0.0001 | 0.0002 | 0.0001 | 0.0004 | 0.0001 | 0.0003 | |
| Southern Swamps | 0.1558 | 0.0196 | 0.0314 | 0.0013 | 0.0203 | 0.0131 | 0.0259 | 0.001 | 0.0002 | 0.0448 | 0.0365 | 0.4916 | 0.0334 | |
| Pate Island | 0.2616 | 0.0517 | 0.0047 | 0.0009 | 0.006 | 0.0004 | 0.0088 | 0.0001 | 0.0001 | 0.0448 | 0.254 | 0.049 | 0.008 | |
| Mongoni-Dodori Creeks | 0.0902 | 0.0639 | 0.0075 | 0.0076 | 0.0589 | 0.0021 | 0.0602 | 0.0034 | 0.0004 | 0.0365 | 0.254 | 0.0709 | 0.0087 | |
| Northern Central | 0.1681 | 0.0222 | 0.0482 | 0.0058 | 0.0418 | 0.0252 | 0.0169 | 0.0013 | 0.0001 | 0.4916 | 0.049 | 0.0709 | 0.1647 | |
| Northern Swamps | 0.014 | 0.0004 | 0.0648 | 0.0004 | 0.0091 | 0.0005 | 0.0008 | 0.0003 | 0.0003 | 0.0334 | 0.008 | 0.0087 | 0.1647 |
| Mean DBH (cm) | Mean Height (m) | Basal Area (m2 ha−1) | Stand Density (trees ha−1) | |
|---|---|---|---|---|
| Mean Height (m) | 0.668 | |||
| 0.009 | ||||
| Basal Area (m2 ha−1) | 0.777 | 0.504 | ||
| 0.001 | 0.066 | |||
| Stand Density (Trees ha−1) | −0.688 | −0.549 | −0.307 | |
| 0.006 | 0.042 | 0.286 | ||
| Mean AGB (Mg ha−1) | 0.814 | 0.532 | 0.968 | −0.433 |
| 0.000 | 0.050 | 0.000 | 0.122 |
References
- Hogarth, P.J. The Biology of Mangroves and Seagrasses; Oxford University Press: Oxford, UK, 2015; ISBN 0-19-871654-0. [Google Scholar]
- Twilley, R.R. Properties of Mangrove Ecosystems Related to the Energy Signature of Coastal Environments. In Maximum Power: The Ideas and Applications of H.T. Odum; University Press of Colorado: Boulder, CO, USA, 1995; pp. 43–62. [Google Scholar]
- Woodroffe, C.D.; Rogers, K.; McKee, K.L.; Lovelock, C.E.; Mendelssohn, I.; Saintilan, N. Mangrove Sedimentation and Response to Relative Sea-Level Rise. Annu. Rev. Mar. Sci. 2016, 8, 243–266. [Google Scholar] [CrossRef] [Green Version]
- Tomlinson, P.B. The Botany of Mangroves; Cambridge University Press: Cambridge, UK, 2016; ISBN 1-316-79065-7. [Google Scholar]
- Osland, M.J.; Feher, L.C.; Griffith, K.T.; Cavanaugh, K.C.; Enwright, N.M.; Day, R.H.; Stagg, C.L.; Krauss, K.W.; Howard, R.J.; Grace, J.B.; et al. Climatic Controls on the Global Distribution, Abundance, and Species Richness of Mangrove Forests. Ecol. Monogr. 2017, 87, 341–359. [Google Scholar] [CrossRef] [Green Version]
- Thom, B. Mangrove Ecology: A Geomorphological Perspective. In Mangrove Ecosystem in Australia: Structure, Function, and Management: Proceedings of the Australian National Mangrove Workshop, Australian Institute of Marine Science, Cape Ferguson, 18–20 April 1979; Australian Institute of Marine Science: Townsville, Australia, 1982; pp. 3–17. [Google Scholar]
- Snedaker, S.C.; Snedaker, J.G. The Mangrove Ecosystem: Research Methods; UNESCO: London, UK, 1984; ISBN 92-3-102181-8. [Google Scholar]
- Woodroffe, C. Mangrove Sediments and Geomorphology. In Coastal and Estuarine Studie; American Geophysical Union: Washington, DC, USA, 1993; Volume 7. [Google Scholar]
- Lugo, A.E.; Snedaker, S.C. The Ecology of Mangroves. Annu. Rev. Ecol. Syst. 1974, 5, 39–64. [Google Scholar] [CrossRef]
- Schaeffer-Novelli, Y.; Cintrón-Molero, G.; Soares, M.; De-Rosa, T. Brazilian Mangroves. Aquat. Ecosyst. Health Manag. 2000, 3, 561–570. [Google Scholar] [CrossRef]
- Odum, H.T. An Introduction. In Systems Ecology; Wiley: New York, NY, USA, 1983; ISBN 0471652776 9780471652779. [Google Scholar]
- Thom, B.G. Coastal Landforms and Geomorphic Processes. Monogr. Oceanogr. Methodol. 1984, 8, 3–17. [Google Scholar]
- Twilley, R.R.; Rivera-Monroy, V.H. Developing Performance Measures of Mangrove Wetlands Using Simulation Models of Hydrology, Nutrient Biogeochemistry, and Community Dynamics. J. Coast. Res. 2005, 40, 79–93. [Google Scholar]
- Balke, T.; Friess, D.A. Geomorphic Knowledge for Mangrove Restoration: A Pan-tropical Categorization. Earth Surf. Process. Landf. 2016, 41, 231–239. [Google Scholar] [CrossRef]
- Twilley, R.R.; Rivera-Monroy, V.H.; Rovai, A.S.; Castañeda-Moya, E.; Davis, S. Mangrove Biogeochemistry at Local to Global Scales Using Ecogeomorphic Approaches. In Coastal Wetlands; Elsevier: Amsterdam, The Netherlands, 2019; pp. 717–785. [Google Scholar]
- Worthington, T.A.; Zu Ermgassen, P.S.; Friess, D.A.; Krauss, K.W.; Lovelock, C.E.; Thorley, J.; Tingey, R.; Woodroffe, C.D.; Bunting, P.; Cormier, N. A Global Biophysical Typology of Mangroves and Its Relevance for Ecosystem Structure and Deforestation. Sci. Rep. 2020, 10, 1–11. [Google Scholar] [CrossRef]
- GoK. National Mangrove Ecosystem Management Plan (2017–2027); Kenya Forest Service: Nairobi, Kenya, 2017.
- Kairo, J.G. Ecology and Restoration of Mangrove Systems in Kenya. Ph.D. Thesis, Vrije Universiteit Brussel, Brussels, Belgium, 2001. [Google Scholar]
- Ferguson, W. A Land (Scape) Ecological Survey of the Mangrove Resource of Kenya-Draft Report; Forest Department: Nairobi, Kenya, 1993. [Google Scholar]
- Lang’at, J.K.S. Variability of Mangrove Forests along the Kenyan Coast; Kenya Marine and Fisheries Research Institute: Mombasa, Kenya, 2008. [Google Scholar]
- Woodroffe, C.D. Pacific Island Mangroves: Distribution and Environmental Settings; University of Hawaii: Honolulu, HI, USA, 1987. [Google Scholar]
- Woodroffe, C.D. Coasts: Form, Process and Evolution; Cambridge University Press: Cambridge, UK, 2002; ISBN 0-521-01183-3. [Google Scholar]
- Schaeffer-Novelli, Y.; Cintrón-Molero, G.; Adaime, R.R.; de Camargo, T.M. Variability of Mangrove Ecosystems along the Brazilian Coast. Estuaries 1990, 13, 204–218. [Google Scholar] [CrossRef]
- Ellison, J.C. Biogeomorphology of Mangroves. In Coastal Wetlands; Elsevier: Amsterdam, The Netherlands, 2019; pp. 687–715. [Google Scholar]
- Government of Kenya. State of the Coast Report II: Enhancing Integrated Management of Coastal and Marine Resources in Kenya; National Environment Management Authority (NEMA): Nairobi, Kenya, 2017.
- ASCLME. National Marine Ecosystem Diagnostic Analysis—Kenya; Contribution to the Agulhas and Somali Current Large Marine Ecosystems Project (supported by UNDP with GEF grant financing); UNDP: Nairobi, Kenya, 2012. [Google Scholar]
- Tychsen, J.; Klinge, H. KenSea. Environmental Sensitivity Atlas for Coastal Area of Kenya. Geological Survey of Denmark and Greenland: Copenhagen, Denmark, 2006. [Google Scholar]
- Bosire, J.; Mangora, M.M.; Bandeira, S.; Rajkaran, A.; Ratsimbazafy, R.; Appadoo, C.; Kairo, J. Mangroves of the Western Indian Ocean: Status and Management; WIOMSA: Zanzibar, Tanzania, 2015; ISBN 9987-9559-4-0. [Google Scholar]
- Oosterom, A.P. The Geomorphology of Southeast Kenya. Ph.D. Thesis, Agricultural University, Wageningen, The Netherlands, 1988. [Google Scholar]
- Okoola, R.E. A Diagnostic Study of the Eastern Africa Monsoon Circulation during the Northern Hemisphere Spring Season. Int. J. Climatol. J. R. Meteorol. Soc. 1999, 19, 143–168. [Google Scholar] [CrossRef]
- Schott, F.A.; Xie, S.-P.; McCreary, J.P., Jr. Indian Ocean Circulation and Climate Variability. Rev. Geophys. 2009, 47, RG1002. [Google Scholar] [CrossRef]
- Kitheka, J.U.; Mavuti, K.M. Tana Delta and Sabaki Estuaries of Kenya: Freshwater and Sediment Input, Upstream Threats and Management Challenges. In Estuaries: A Lifeline of Ecosystem Services in the Western Indian Ocean; Springer: Cham, Switzerland, 2016; pp. 89–109. [Google Scholar]
- Kuria, Z. Groundwater Distribution and Aquifer Characteristics in Kenya. In Developments in Earth Surface Processes; Elsevier: Amsterdam, The Netherlands, 2013; Volume 16, ISBN 0928-2025. pp. 83–107. [Google Scholar]
- Mwango, F.K.; Muhangú, B.; Juma, C.; Githae, I. Groundwater Resources in Kenya. In Proceedings of the International Workshop on ‘Managing Shared Aquifer Resources in Africa’, Tripoli, Libya, 2–4 June 2002; pp. 93–100. [Google Scholar]
- Nijsten, G.-J.; Christelis, G.; Villholth, K.G.; Braune, E.; Gaye, C.B. Transboundary Aquifers of Africa: Review of the Current State of Knowledge and Progress towards Sustainable Development and Management. J. Hydrol. Reg. Stud. 2018, 20, 21–34. [Google Scholar] [CrossRef]
- Oiro, S.; Comte, J.-C. Drivers, Patterns and Velocity of Saltwater Intrusion in a Stressed Aquifer of the East African Coast: Joint Analysis of Groundwater and Geophysical Data in Southern Kenya. J. Afr. Earth Sci. 2019, 149, 334–347. [Google Scholar] [CrossRef] [Green Version]
- Kenya National Bureau of Statistics. Kenya National Bureau of Statistics. Volume I: Population by County and Sub-County; Kenya National Bureau of Statistics: Nairobi, Kenya, 2019; p. 49.
- Kenya National Bureau of Statistics. 2019 Kenya Population and Housing Census. Volume II: Distribution of Population by Administrative Units; Kenya National Bureau of Statistics: Nairobi, Kenya, 2019; p. 270.
- Kauffman, J.B.; Donato, D.C. Protocols for the Measurement, Monitoring and Reporting of Structure, Biomass and Carbon Stocks in Mangrove Forests; CIFOR: Bogor, Indonesia, 2012. [Google Scholar]
- Cintron, G.; Schaeffer Novelli, Y. Methods for Studying Mangrove Structure. Monogr. Oceanogr. Methodol. 1984, 8, 91–113. [Google Scholar]
- Komiyama, A.; Poungparn, S.; Kato, S. Common Allometric Equations for Estimating the Tree Weight of Mangroves. J. Trop. Ecol. 2005, 21, 471–477. [Google Scholar] [CrossRef]
- Gillerot, L.; Vlaminck, E.; De Ryck, D.J.; Mwasaru, D.M.; Beeckman, H.; Koedam, N. Inter-and Intraspecific Variation in Mangrove Carbon Fraction and Wood Specific Gravity in Gazi Bay, Kenya. Ecosphere 2018, 9, e02306. [Google Scholar] [CrossRef]
- Lewis, R.R., III; Milbrandt, E.C.; Brown, B.; Krauss, K.W.; Rovai, A.S.; Beever, J.W., III; Flynn, L.L. Stress in Mangrove Forests: Early Detection and Preemptive Rehabilitation Are Essential for Future Successful Worldwide Mangrove Forest Management. Mar. Pollut. Bull. 2016, 109, 764–771. [Google Scholar] [CrossRef]
- Kamau, J.; Ngisiange, N.; Ochola, O.; Kilionzi, J.; Kimeli, A.; Mahongo, S.B.; Onganda, H.; Mitto, C.; Ohowa, B.; Magori, C. Factors Influencing Spatial Patterns in Primary Productivity in Kenyan Territorial Waters. West. Indian Ocean J. Mar. Sci. 2020, 9–18. [Google Scholar] [CrossRef]
- Kitheka, J.U.; Obiero, M.; Nthenge, P. River Discharge, Sediment Transport and Exchange in the Tana Estuary, Kenya. Estuar. Coast. Shelf Sci. 2005, 63, 455–468. [Google Scholar] [CrossRef]
- Kairo, J.; Kivyatu, B.; Koedam, N. Application of Remote Sensing and GIS in the Management of Mangrove Forests within and Adjacent to Kiunga Marine Protected Area, Lamu, Kenya. Environ. Dev. Sustain. 2002, 4, 153–166. [Google Scholar] [CrossRef]
- Halo, I.; Sagero, P.; Manyilizu, M.; Mahongo, S.B. Biophysical Modelling of Coastal Upwelling Variability and Circulation along the Tanzanian and Kenyan Coasts. West. Indian Ocean J. Mar. Sci. 2020, 43–61. [Google Scholar] [CrossRef]
- Kairo, J.G.; Hamza, A.; Wanjiru, C. Mikoko Pamoja: A Demonstrably Effective Community-Based Blue Carbon Project in Kenya. In A Blue Carbon Primer; CRC Press: Boca Raton, FL, USA, 2018; pp. 341–350. ISBN 0-429-43536-3. [Google Scholar]
- Mohamed, M.O.S.; Neukermans, G.; Kairo, J.G.; Dahdouh-Guebas, F.; Koedam, N. Mangrove Forests in a Peri-Urban Setting: The Case of Mombasa (Kenya). Wetl. Ecol. Manag. 2009, 17, 243–255. [Google Scholar] [CrossRef] [Green Version]






| Site | Environmental Setting | Annual Ave. Precipitation (mm) | Population Density Per sq. km. | River Influence * |
|---|---|---|---|---|
| Vanga | Estuarine Coast | 1109 | 311 | Low |
| Gazi | Lagoonal Coast | 1236 | 115 | Low |
| Mombasa | Estuarine Coast | 1050 | 6964 | Moderate |
| Mtwapa | Tidal Coast | 1090 | 1627 | None |
| Kilifi | Tidal Coast | 954 | 676 | Low |
| Mida | Tidal Coast | 930 | 537 | None |
| Ngomeni | Deltaic Coast | 979 | 676 | High |
| Mto Tana | Tidal Coast | 575 | 22 | Moderate |
| Kipini | Deltaic Coast | 947 | 58 | High |
| Southern S | Lagoonal Coast | 960 | 77 | Low |
| Pate Island | Lagoonal Coast | 960 | 229 | None |
| MD Creeks | Tidal Coast | 960 | 10 | None |
| N Central | Lagoonal Coast | 863 | 136 | None |
| Northern | Open Coast | 560 | 5 | None |
| Taxon | Av. Dissim | Contrib. % | Cumulative % |
|---|---|---|---|
| Sonneratia alba | 4.395 | 20.05 | 20.05 |
| Ceriops tagal | 4.306 | 19.64 | 39.69 |
| Avicennia marina | 3.11 | 14.18 | 53.87 |
| Bruguiera gymnorhiza | 2.978 | 13.58 | 67.46 |
| Rhizophora mucronata | 2.473 | 11.28 | 78.74 |
| Xylocarpus granatum | 2.135 | 9.739 | 88.48 |
| Heritiera littoralis | 1.672 | 7.628 | 96.11 |
| Lumnitzera racemosa | 0.6135 | 2.798 | 98.91 |
| Xylocarpus moluccensis | 0.24 | 1.095 | 100 |
| Site | DBH (cm) | Group * | Height (m) | Group * | AGB (Mg ha−1) | Group * |
|---|---|---|---|---|---|---|
| Vanga | 7.05 ± 0.09 | ef | 5.21 ± 0.05 | f | 199.9 ± 28 | cde |
| Gazi | 6.28 ± 0.18 | g | 4.29 ± 0.10 | h | 171.9 ± 34.7 | defg |
| Mombasa | 5.55 ± 0.06 | h | 4.21 ± 0.03 | h | 76.77 ± 9.61 | f |
| Mtwapa | 4.29 ± 0.05 | i | 3.06 ± 0.03 | i | 89.2 ± 13.4 | g |
| Kilifi | 6.53 ± 0.18 | fg | 3.97 ± 0.09 | h | 141.9 ± 28.3 | cdef |
| Mida | 5.89 ± 0.13 | gh | 4.89 ± 0.06 | g | 150.7 ± 27 | ef |
| Ngomeni | 8.09 ± 0.22 | c | 6.04 ± 0.08 | e | 287.6 ± 70.6 | b |
| Mto Tana | 5.95 ± 0.15 | gh | 4.13 ± 0.10 | h | 91.2 ± 31.4 | fg |
| Kipini | 9.53 ± 0.30 | b | 7.26 ± 0.14 | d | 358 ± 106 | a |
| Southern Swamps | 7.20 ± 0.08 | def | 5.03 ± 0.03 | fg | 229.3 ± 21 | cde |
| Pate Island Swamps | 9.45 ± 0.21 | b | 9.71 ± 0.09 | b | 258.5 ± 36.5 | b |
| Mongoni Dodori Creek | 7.80 ± 0.18 | cd | 9.43 ± 0.08 | b | 183 ± 21.3 | bcd |
| Northern Central Swamps | 7.42 ± 0.11 | de | 11.54 ± 0.06 | a | 205.1 ± 21.5 | bc |
| Northern Swamps | 10.95 ± 0.16 | a | 7.92 ± 0.09 | c | 238.5 ± 19.4 | a |
| Site | Environmental Setting | Mangrove Structural Properties | |||||
|---|---|---|---|---|---|---|---|
| (1) Species | (2) BA | (3) Height | (4) SD | (5) AGB | (6) CI | ||
| Vanga | Estuarine coast | 7 | 19.7 ± 2.2 | 5.2 ± 0.1 | 2987 ± 279 | 199.9 ± 28 | 21.5 |
| Gazi | Lagoonal Coast | 6 | 20.8 ± 3.6 | 4.3 ± 0.1 | 3730 ± 561 | 171.9 ± 34.7 | 19.9 |
| Mombasa | Estuarine Coast | 8 | 8.7 ± 0.8 | 4.2 ± 0.0 | 2113 ± 154 | 76.8 ± 9.61 | 6.2 |
| Mtwapa | Tidal Creek | 4 | 14.1 ± 2.2 | 3.1 ± 0.0 | 7856 ± 2094 | 89.2 ± 13.4 | 13.5 |
| Kilifi | Tidal Creek | 5 | 15.7 ± 2.3 | 4.0 ± 0.1 | 3418 ± 627 | 141.9 ± 28.3 | 10.7 |
| Mida | Tidal Creek | 4 | 16.1 ± 1.9 | 4.9 ± 0.1 | 4913 ± 826 | 150.7 ± 27 | 15.4 |
| Ngomeni | Deltaic coast | 6 | 26.1 ± 5.6 | 6.0 ± 0.1 | 3179 ± 554 | 287.6 ± 70.6 | 30.0 |
| Mto Tana | Deltaic Coast | 4 | 11.4 ± 2.9 | 4.1 ± 0.1 | 2739 ± 749 | 91.2 ± 31.4 | 5.2 |
| Kipini | Deltaic Coast | 5 | 29.8 ± 5.9 | 7.3 ± 0.1 | 2164 ± 391 | 358 ± 106 | 23.4 |
| Southern | Lagoonal Coast | 5 | 21.7 ± 1.5 | 5.0 ± 0.0 | 3092 ± 213 | 229.3 ± 21 | 16.9 |
| Pate Island | Lagoonal Coast | 6 | 26.7 ± 3.4 | 9.7 ± 0.1 | 2302 ± 315 | 258.5 ± 36.5 | 35.8 |
| MD Creek | Tidal Creek | 6 | 18.4 ± 1.8 | 9.4 ± 0.1 | 2169 ± 296 | 183 ± 21.3 | 22.6 |
| NC | Lagoonal Coast | 5 | 19.4 ± 1.4 | 11.5 ± 0.1 | 2496 ± 272 | 205.1 ± 21.5 | 28.0 |
| Northern | Open Coast | 5 | 22.6 ± 1.5 | 7.9 ± 0.1 | 1607 ± 129 | 238.5 ± 19.4 | 14.4 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Njiru, D.M.; Githaiga, M.N.; Nyaga, J.M.; Lang’at, K.S.; Kairo, J.G. Geomorphic and Climatic Drivers Are Key Determinants of Structural Variability of Mangrove Forests along the Kenyan Coast. Forests 2022, 13, 870. https://0-doi-org.brum.beds.ac.uk/10.3390/f13060870
Njiru DM, Githaiga MN, Nyaga JM, Lang’at KS, Kairo JG. Geomorphic and Climatic Drivers Are Key Determinants of Structural Variability of Mangrove Forests along the Kenyan Coast. Forests. 2022; 13(6):870. https://0-doi-org.brum.beds.ac.uk/10.3390/f13060870
Chicago/Turabian StyleNjiru, Derrick Muthomi, Michael Njoroge Githaiga, Justine Muhoro Nyaga, Kipkorir Sigi Lang’at, and James Gitundu Kairo. 2022. "Geomorphic and Climatic Drivers Are Key Determinants of Structural Variability of Mangrove Forests along the Kenyan Coast" Forests 13, no. 6: 870. https://0-doi-org.brum.beds.ac.uk/10.3390/f13060870