
Does the Age of Pinus sylvestris Mother Trees Influence Reproductive Capacity and Offspring Seedling Survival?

 , , and
, , and
Abstract
:1. Introduction
2. Materials and Methods
3. Results
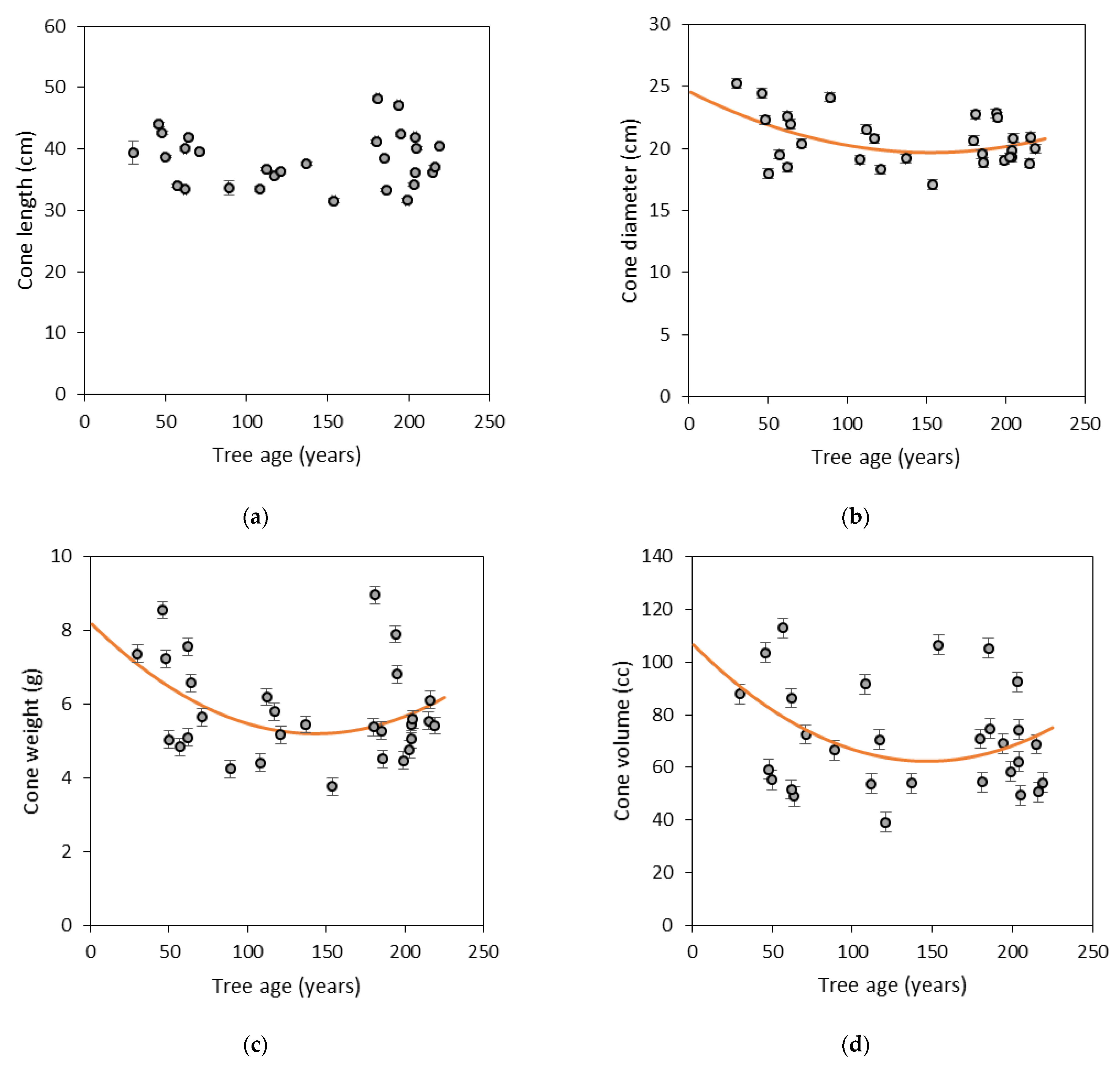
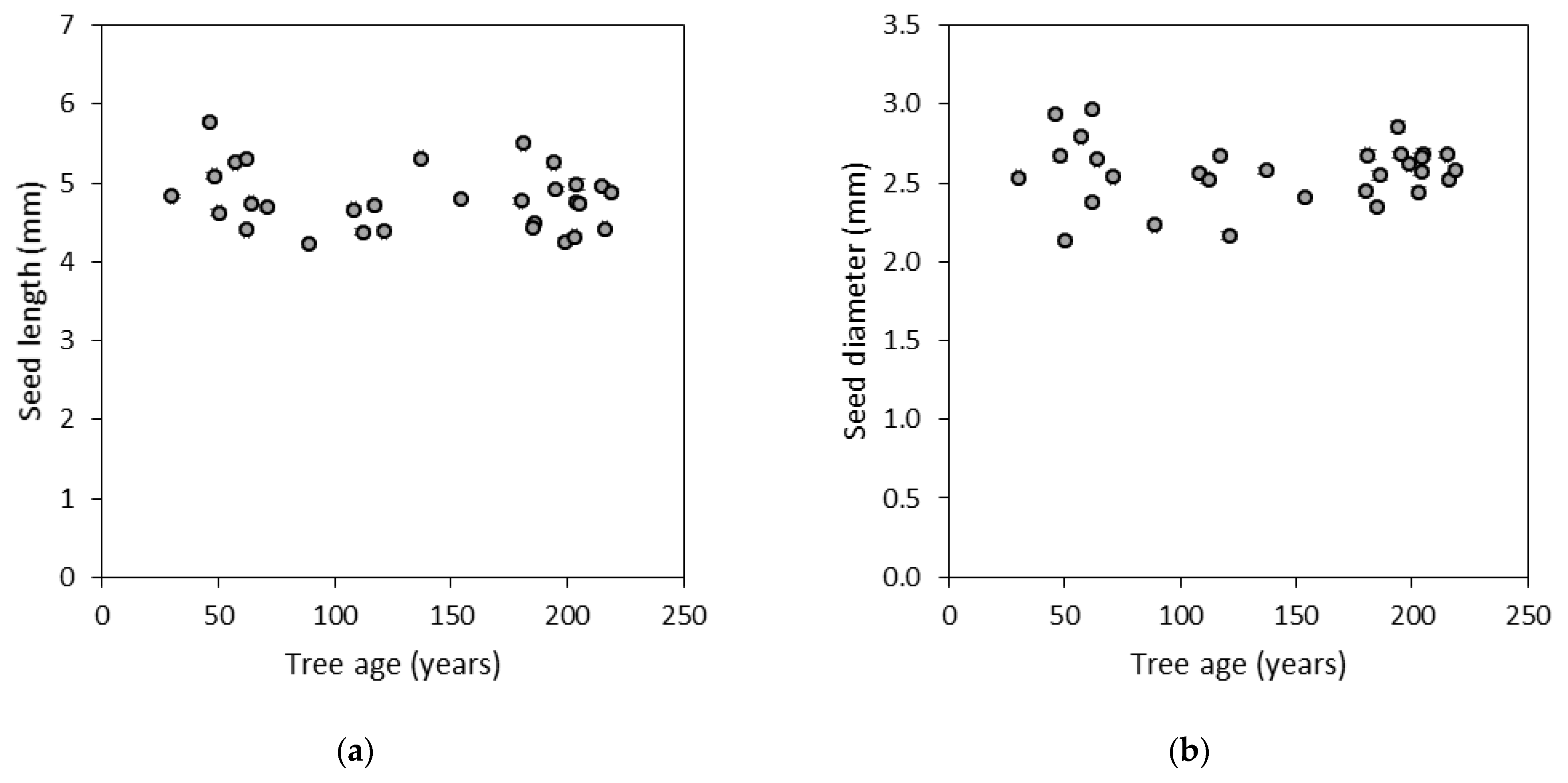
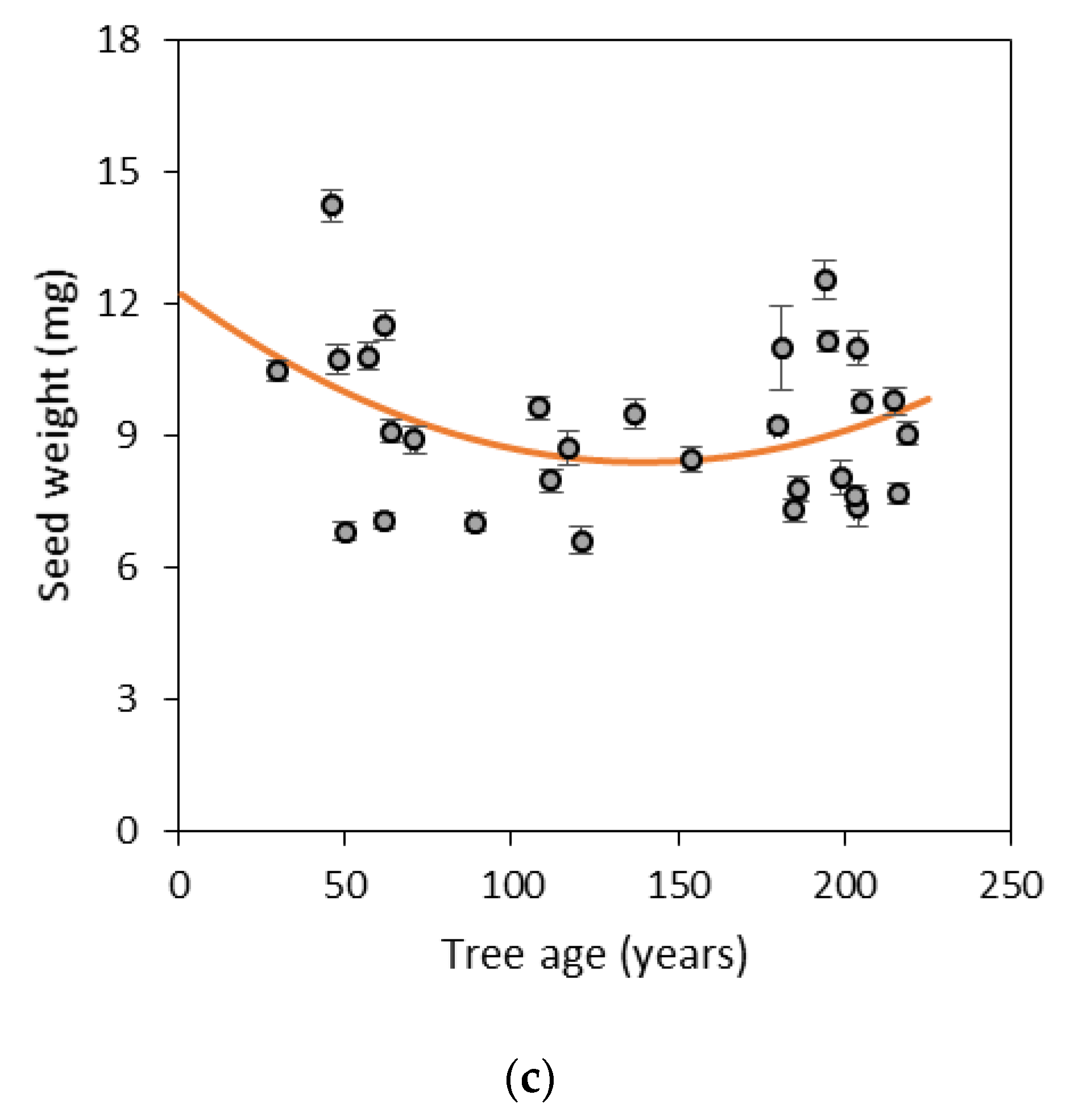
3.1. Morphological Traits of Cones and Seeds
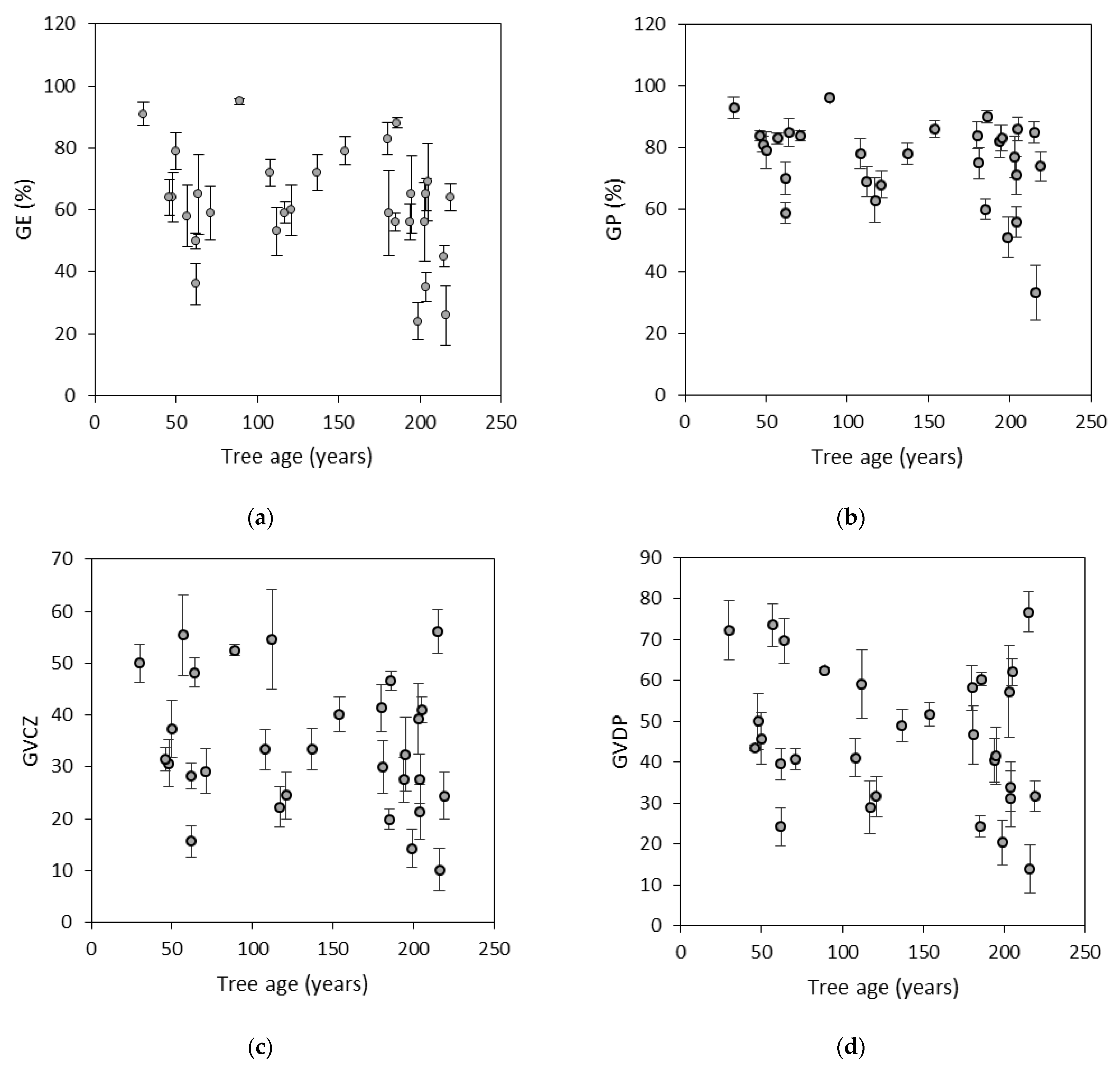
3.2. Germination Capacity and Seed Viability
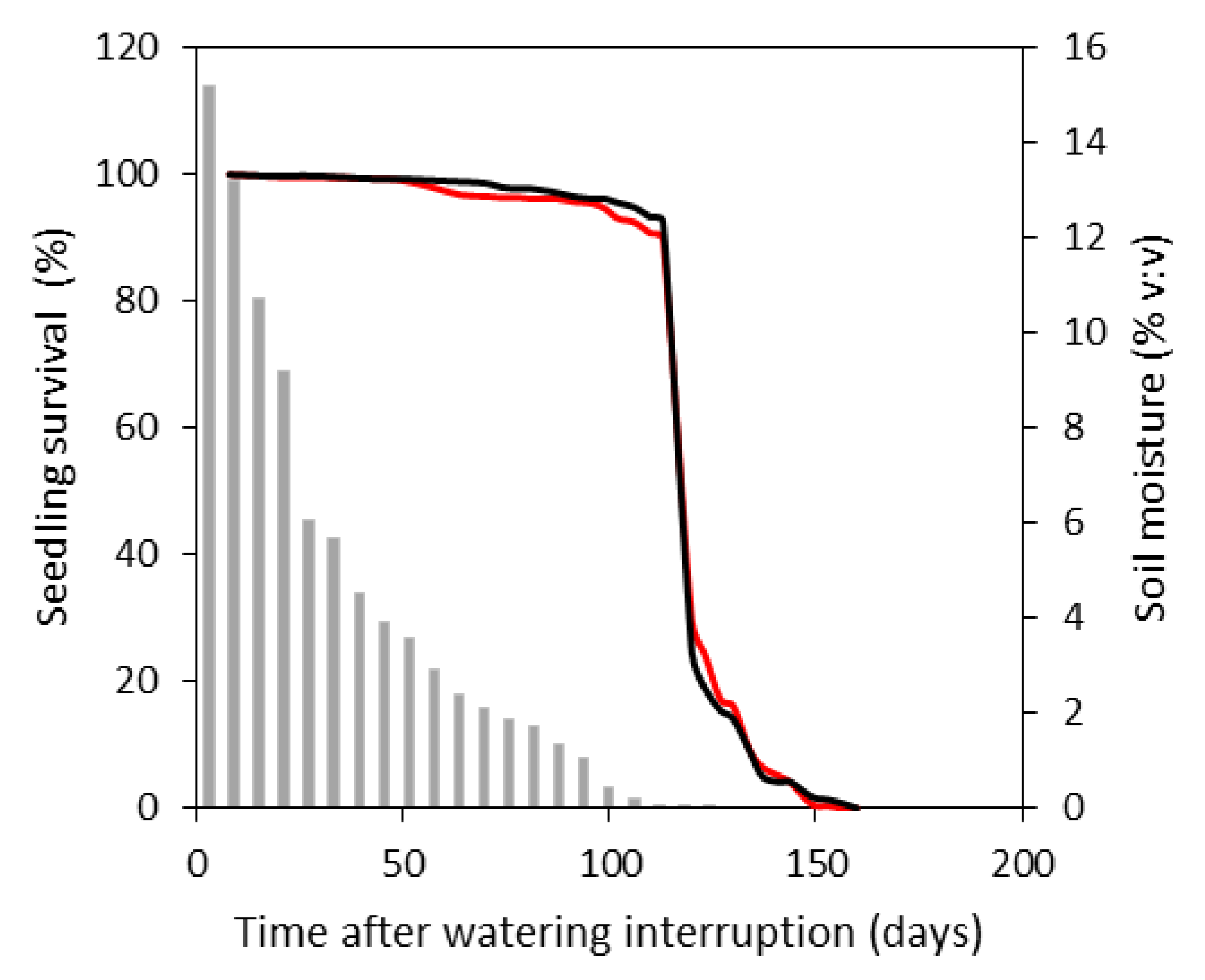
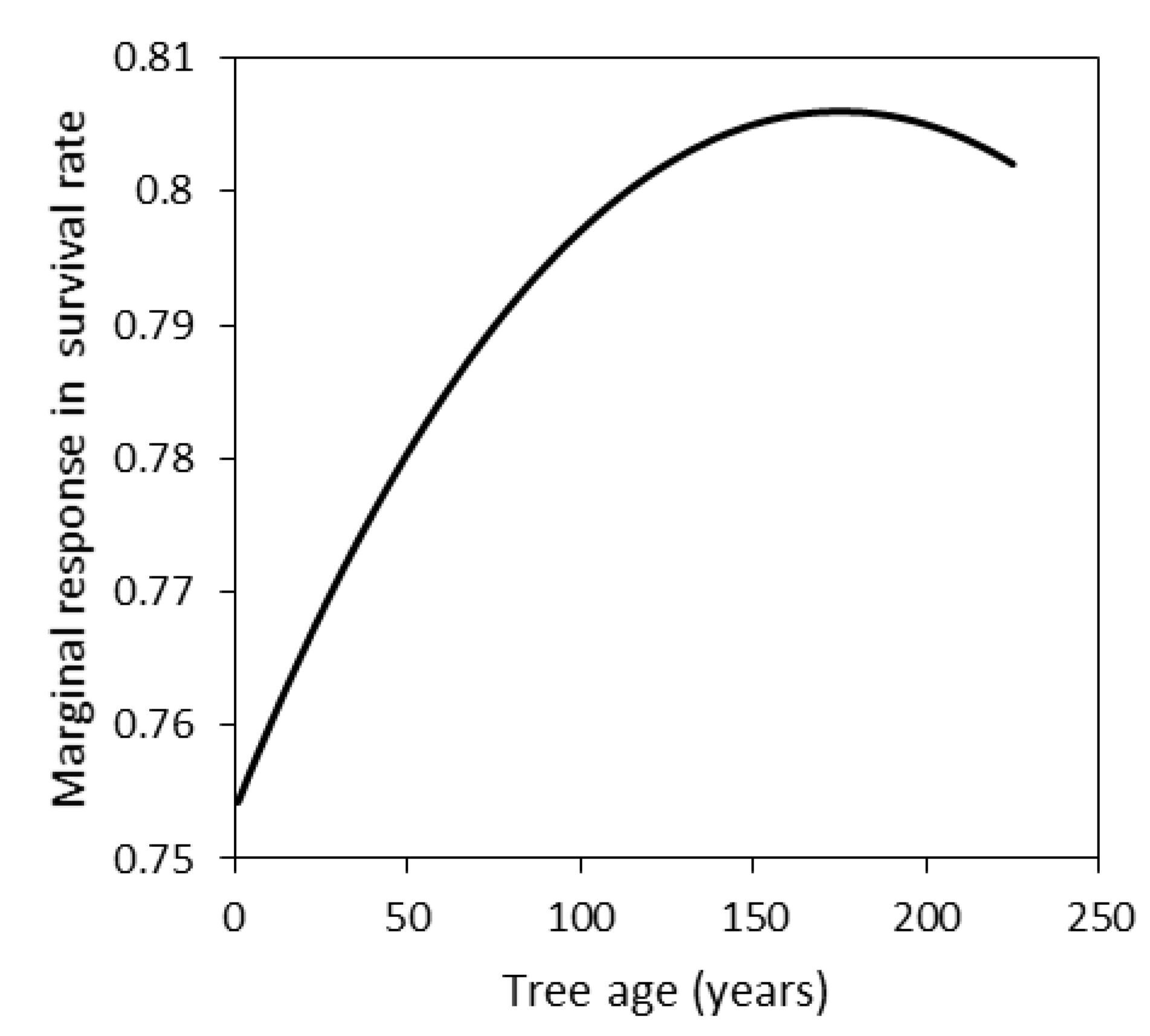
3.3. Seedling Survival in the Death Trial
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Grove, A.T.; Rackham, O. The Nature of Mediterranean Europe: An Ecological History; Yale University Press: New Haven, CT, USA; London, UK, 2001. [Google Scholar]
- Peterken, G.F. Natural Woodland: Ecology and Conservation in Northern Temperate Regions; Cambridge University Press: Cambridge, UK, 1996. [Google Scholar]
- Thomas, H. Senescence, ageing and death of the whole plant. New Phytol. 2013, 197, 696–711. [Google Scholar] [CrossRef] [PubMed]
- Oliver, C.D.; Larson, B.C. Forest Stand Dynamics; Wiley Ed.: Hoboken, NJ, USA, 1990; p. 467. [Google Scholar]
- Pardos, M.; Madrigal, G.; Robledo, J.J.; Climent, J.; Alía, R.; Montes, F.; Camarero, J.J.; Rubio, A.; Calama, R. Efecto del envejecimiento sobre la capacidad reproductiva de Quercus suber en los bosques maduros de espacios protegidos. Cuad. De La SECF 2019, 45, 19–36, ISSN 2386-83682019. [Google Scholar]
- Lindenmayer, D.B.; Franklin, J.F. Congruence between natural and human forest disturbance: A case study for Australian montane ash forest. For. Ecol. Manag. 2002, 155, 319–335. [Google Scholar] [CrossRef]
- Nadrowski, K.; Wirth, C.; Scherer-Lorenzen, M. Is forest diversity driving ecosystem function and service? Curr. Opin. Environ. Sustain. 2010, 524, 75–79. [Google Scholar] [CrossRef]
- Carey, E.V.; Sala, A.; Keane, M.; Callaway, R.M. Are old forests underestimated as global carbon sinks? Glob. Chang. Biol. 2001, 7, 339–344. [Google Scholar] [CrossRef]
- Luyssaert, S.; Detlef Schulze, E.; Börner, A.; Knohl, A.; Hessenmöller, D.; Law, B.E.; Ciais, P.; Grace, J. Old-growth forests as global carbon sinks. Nature 2008, 455, 213–215. [Google Scholar] [CrossRef]
- Alejano, R.; Domínguez-Delmás, M.; García-González, I.; Wazny, T.; Vázquez-Piqué, J.; Fernández-Martínez, M. The age of black pine (Pinus nigra Arn. ssp. salzmannii (Dunal) Franco) mother trees has no effect on seed germination and on offspring seedling performance. Ann. For. Sci. 2019, 76, 15. [Google Scholar] [CrossRef] [Green Version]
- Nyland, R. Silviculture: Concepts and Applications, 2nd ed.; The McGraw-Hill Companies: New York, NY, USA, 2002. [Google Scholar]
- Calama, R.; Manso, R.; Lucas-Borja, M.E.; Espelta, J.M.; Piqué, M.; Bravo, F.; Del Peso, C.; Pardos, M. Natural regeneration in Iberian pines: A review of dynamic processes and proposals for management. For. Syst. 2017, 26, eR02S. [Google Scholar] [CrossRef]
- Pukkala, T.; Kolström, T. A stochastic spatial regeneration model for Pinus sylvestris. Scand. J. For. Res. 1992, 7, 377–385. [Google Scholar] [CrossRef]
- Manso, R.; Pukkala, R.; Pardos, M.; Miina, J.; Calama, R. Modelling Pinus pinea forest management to attain natural regeneration under present and future climatic scenarios. Can. J. For. Res. 2014, 44, 250–262. [Google Scholar] [CrossRef] [Green Version]
- Petit, R.; Hampe, A. Some evolutionary consequences of being a tree. Annu. Rev. Ecol. Evol. Syst. 2006, 37, 187–214. [Google Scholar] [CrossRef] [Green Version]
- Mukassabi, T.A.; Polwart, A.; Coleshaw, T.; Thomas, P.A. Scots pine seed dynamics on a waterlogged site. Trees 2012, 26, 1305–1315. [Google Scholar] [CrossRef]
- Lanner, R.M.; Connor, K.F. Does bristlecone pine senesce? Exp. Gerontol. 2001, 36, 675–685. [Google Scholar] [CrossRef]
- Tíscar-Oliver, P.A. Capacidad reproductiva de Pinus nigra subsp. Salzmannii en relación con la edad de la planta madre. Investig. Agrar. Sist. Recur. For. 2002, 11, 357–371. [Google Scholar]
- Ramírez-Valiente, J.A.; Solé-Medina, A.; Pyhäjärvi, T.; Savolainen, O.; Heer, K.; Opgenoorth, L.; Danusevicius, D.; Robledo-Arnuncio, J.J. Adaptive responses to temperature and precipitation variation at the early-life stages of Pinus sylvestris. New Phytol. 2021, 232, 1632–1647. [Google Scholar] [CrossRef]
- Pardos, M.; Pérez, S.; Calama, R.; Alonso, R.; Lexer, M.J. Ecosystem service provision, management systems and climate change in Valsaín forest, central Spain. REEC 2017, 17, 17–32. [Google Scholar] [CrossRef]
- Martínez-Vilalta, J.; Cochard, H.; Mencuccini, M.; Sterck, F.; Herrero, A.; Korhonen, J.F.J.; Llorens, P.; Nikinmaa, E.; Nolè, A.; Poyatos, R.; et al. Hydraulic adjustment of Scots pine across Europe. New Phytol. 2009, 184, 353–364. [Google Scholar] [CrossRef]
- Speer, J.H. Fundamentals of Tree-Ring Research; University of Arizona Press: Tucson, AZ, USA, 2010. [Google Scholar]
- FAO. A Guide to Forest Seed Handling, 20/2nd ed.; FAO Forestry Paper: Rome, Italy, 1985; ISBN 92-5-102291-7. [Google Scholar]
- Czabator, F.J. Germination value: An index combining speed and completeness of pine seed germination. For. Sci. 1962, 8, 386–396. [Google Scholar]
- Djavanshir, K.; Pourbeik, H. Germination value-A new formula. Silvae Genet. 1976, 25, 79–83. [Google Scholar]
- Rojo, A.; Montero, G. El Pino Silvestre en la Sierra de Guadarrama; Ministerio de Agricultura, Pesca y Alimentación: Madrid, Spain, 1996; p. 293. [Google Scholar]
- Montero, G.; Del Río, M.; Roig, S.; Rojo, A. Selvicultura de Pinus sylvestris Mill. In Compendio de Selvicultura Aplicada en España; Serrada, R., Montero, G., Reque, J.A., Eds.; Inia-Fucovasa: Madrid, Spain, 2008; pp. 503–534. [Google Scholar]
- Calama, R.; Manso, R.; Barbeito, I.; Madrigal, G.; Garriga, E.; Gordo, F.J.; Montero, G.; Cañellas, I.; Pardos, M. Do inter-specific differences in seed size determine natural regeneration traits in Pinus pinea and Pinus sylvestris? Appl. Ecol. Environ. Res. 2015, 13, 387–404. [Google Scholar]
- Aizen, M.A.; Patterson, W.A. Acorn size and geographical range in the North American oaks (Quercus L.). J. Biogeogr. 1990, 17, 327–332. [Google Scholar] [CrossRef] [Green Version]
- Aizen, M.A.; Woodcock, H. Latitudinal trends in acorn size in eastern North American species of Quercus. Can. J. Bot. 1992, 70, 1218–1222. [Google Scholar] [CrossRef]
- Long, T.J.; Jones, R.H. Seedling growth strategies and seed size effects in fourteen oak species native to different soil moisture habitats. Trees 1996, 11, 1–8. [Google Scholar] [CrossRef]
- Parker, W.C.; Noland, T.L.; Morneault, A.E. The effects of seed mass on germination, seedlings emergence, and early seedling growth of eastern white pine (Pinus strobus L.). New For. 2006, 32, 33–49. [Google Scholar] [CrossRef]
- Kaliniewicz, Z.; Rawa, T.; Tylek, P.; Markowski, P.; Anders, A.; Fura, S. The effect of the age of Scots pie (Pinus sylvestris L.) stands on the physical properties of seeds and the operating parameters of cleaning machines. Tech. Sci. 2013, 16, 63–72. [Google Scholar]
- Brutovska, E.; Samelova, A.; Dusicka, J.; Micieta, K. Ageing of trees: Application of general ageing theories. Ageing Res. Rev. 2013, 12, 855–866. [Google Scholar] [CrossRef]
- Zhu, J.; Kang, H.; Tan, H.; Xu, M. Effect of drought stresses induced by polyethylene glycol on germination of Pinus sylvestris var. mongolica seeds from natural and plantation forests on sandy lands. J. For. Res. 2006, 11, 319–328. [Google Scholar] [CrossRef]
- Müller, M.; Siles, L.; Cela, J.; Munné-Bosch, S. Perennially young: Seed production and quality in controlled and natural populations of Cistus albidus reveal compensatory mechanisms that prevent senescence in terms of seed yield and viability. J. Exp. Bot. 2014, 65, 287–297. [Google Scholar] [CrossRef] [Green Version]
- Xu, Y.; Cail, N.; He, B.; Zhang, R.; Zhao, W.; Mao, J.; Duan, A.; Li, Y.; Woeste, K. Germination and early seedling growth of Pinus densata Mast. Provenances. J. For. Res. 2016, 27, 283–294. [Google Scholar] [CrossRef]
- Smith, C.C.; Fretwell, S.D. The optimal balance between size and number of offspring. Am. Nat. 1974, 108, 499–506. [Google Scholar] [CrossRef]
- Willson, M.F. Dispersal mode, seed shadows and colonization patterns. Vegetatio 1993, 15, 261–280. [Google Scholar] [CrossRef]
- Suárez-Vidal, E.; Sampedro, L.; Zas, R. Is the benefit of larger seed provisioning on seedling performance greater under abiotic stress? Environ. Exp. Bot. 2017, 134, 45–53. [Google Scholar] [CrossRef] [Green Version]
- Castro, J. Short delay in timing of emergence determines establishment success in Pinus sylvestris across microhabitats. Ann. Bot. 2006, 98, 1233–1240. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Westoby, M.; Jurado, E.; Leishman, M. Comparative evolutionary ecology of seed size. Trends Ecol. Evol. 1992, 7, 368–372. [Google Scholar] [CrossRef]
- Castro, J. Seed mass versus seedling performance in Scots pine: A maternally dependent trait. New Phytol. 1999, 144, 153–161. [Google Scholar] [CrossRef]
- Leishman, M.R.; Wright, I.J.; Moles, A.T.; Westoby, M. The evolutionary ecology of seed size. In Seeds-The Ecology of Regeneration in Plant Communities, 2nd ed.; Fenner, M., Ed.; CAB International: Wallingford, UK, 2000; pp. 31–57. [Google Scholar]
- Ramírez-Valiente, J.A.; Robledo-Arnuncio, J.J. Exotic gene flow affects fitness trait values but not levels of heritable trait variation in the southernmost population of Scots pine (Pinus sylvestris L. var nevadensis). Biol. Conserv. 2015, 192, 331–342. [Google Scholar] [CrossRef]
- Muilu-Mäkelä, R.; Vuosku, J.; Läärä, E.; Saarinen, M.; Heiskanen, J.; Häggman, H.; Sarjala, T. Water availability influences morphology, mycorrhizal associations, PSII efficiency and polyamine metabolism at early growth phase of Scots pine seedlings. Plant Physiol. Biochem. 2015, 88, 70–81. [Google Scholar] [CrossRef]
- Ivanov, Y.V.; Zlobin, I.E.; Kartashov, A.V.; Pashkovskiy, P.P.; Kuznetsov, V.V. Scale of physiological processes sensitivity to PEG-induced water stress in Scots pine seedlings. Russ. J. Plant Physiol. 2018, 65, 532–540. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Tree No. | Tree Age (yrs) | Diameter (cm) | Height (m) | Tree No. | Tree Age (yrs) | Diameter (cm) | Height (m) |
|---|---|---|---|---|---|---|---|
| 801 | 154 | 62.0 | 12.9 | 817 | 204 | 82.0 | 14.2 |
| 802 | 48 | 34.1 | 12.1 | 818 | 108 | 37.0 | 13.1 |
| 803 | 137 | 29.0 | 14.4 | 819 | 205 | 62.0 | 15.5 |
| 804 | 50 | 29.2 | 13.7 | 820 | 89 | 37.0 | 12.1 |
| 805 | 181 | 64.5 | 11.1 | 821 | 199 | 76.2 | 15 |
| 806 | 57 | 29.2 | 13.7 | 822 | 117 | 40.0 | 104 |
| 807 | 215 | 68.0 | 14.7 | 823 | 204 | 76.0 | 14.3 |
| 808 | 46 | 40.1 | 14.3 | 824 | 185 | 38.5 | 13.6 |
| 809 | 216 | 64.3 | 14.6 | 825 | 194 | 72.5 | 13.8 |
| 810 | 30 | 23.7 | 15 | 826 | 71 | 31.0 | 13.7 |
| 811 | 186 | 58.6 | 15.1 | 827 | 219 | 57 | 10 |
| 812 | 62 | 13.1 | 7.9 | 828 | 203 | 25 | 10.4 |
| 813 | 180 | 66.0 | 13.8 | 829 | 195 | 56.5 | 15.6 |
| 814 | 64 | 28.2 | 12.5 | 830 | 112 | 29.5 | 16.1 |
| 816 | 62 | 35.6 | 11.3 | 832 | 121 | 41.5 | 13.7 |
| Variables | Covariance Parameter Estimates | Variance Explained by Tree Effect (%) | p (σ2t = 0) | Tree Age Effect | |||
|---|---|---|---|---|---|---|---|
| Tree (σ2t) | Residual (σ2e) | Total | Age (p > F) | Age2 (p > F) | |||
| Length | 18.02 | 20.14 | 38.15 | 47 | 0.0001 | 0.16 | 0.15 |
| Diameter | 3.96 | 6.28 | 10.24 | 39 | 0.0001 | 0.07 | 0.19 |
| Weight | 1.61 | 2.01 | 3.62 | 44 | 0.0001 | 0.07 | 0.09 |
| Volume | 385.71 | 498.71 | 884.42 | 44 | 0.0001 | 0.08 | 0.12 |
| Variables | Covariance Parameter Estimates | Variance Explained by Tree Effect (%) | p (σ2t = 0) | Tree Age Effect | |||
|---|---|---|---|---|---|---|---|
| Tree (σ2t) | Residual (σ2e) | Total | Age (p > F) | Age2 (p > F) | |||
| Length | 0.15 | 0.18 | 0.33 | 45 | <0.0001 | 0.24 | 0.29 |
| Diameter | 0.04 | 0.07 | 0.11 | 36 | <0.0001 | 0.15 | 0.15 |
| Weight | 3.29 | 11.67 | 14.95 | 22 | 0.0001 | 0.09 | 0.12 |
| Variables | Covariance Parameter Estimates | Variance Explained by Tree Effect (%) | p (σ2t = 0) | Tree Age Effect | |||
|---|---|---|---|---|---|---|---|
| Tree (σ2t) | Residual (σ2e) | Total | Age (p > F) | Age2 (p > F) | |||
| GE | 234.3 | 225.5 | 459.8 | 51 | <0.0001 | 0.45 | 0.31 |
| GP | 157.9 | 85.8 | 242.7 | 65 | <0.0001 | 0.86 | 0.92 |
| GVCZ | 134.1 | 85.7 | 219.8 | 61 | <0.0001 | 0.94 | 0.89 |
| GVDP | 237.5 | 122.2 | 359.7 | 66 | <0.0001 | 0.49 | 0.63 |
| VS | 0.0698 | 0.74 | 0.72 | ||||
| Fixed Effect | Parameter Estimate | Standard Error | t Value | P > |t| | |
|---|---|---|---|---|---|
| Intercept | 1.1180 | 0.7036 | 1.59 | 0.1219 | |
| Age | 0.003500 | 0.001546 | 2.26 | 0.0238 | |
| Age2 | −0.00001 | 5.924 × 10−6 | −1.92 | 0.0557 | |
| Random effects | Covariance estimate | Standard error | Z | P > Z | |
| Date | 15.9918 | 4.1338 | 3.87 | <0.0001 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pardos, M.; Vázquez-Piqué, J.; Benito, L.; Madrigal, G.; Alejano, R.; Fernández, M.; Calama, R. Does the Age of Pinus sylvestris Mother Trees Influence Reproductive Capacity and Offspring Seedling Survival? Forests 2022, 13, 937. https://0-doi-org.brum.beds.ac.uk/10.3390/f13060937
Pardos M, Vázquez-Piqué J, Benito L, Madrigal G, Alejano R, Fernández M, Calama R. Does the Age of Pinus sylvestris Mother Trees Influence Reproductive Capacity and Offspring Seedling Survival? Forests. 2022; 13(6):937. https://0-doi-org.brum.beds.ac.uk/10.3390/f13060937
Chicago/Turabian StylePardos, Marta, Javier Vázquez-Piqué, Luis Benito, Guillermo Madrigal, Reyes Alejano, Manuel Fernández, and Rafael Calama. 2022. "Does the Age of Pinus sylvestris Mother Trees Influence Reproductive Capacity and Offspring Seedling Survival?" Forests 13, no. 6: 937. https://0-doi-org.brum.beds.ac.uk/10.3390/f13060937