
Spatial Scale Effects of Soil Respiration in Arid Desert Tugai Forest: Responses to Plant Functional Traits and Soil Abiotic Factors

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
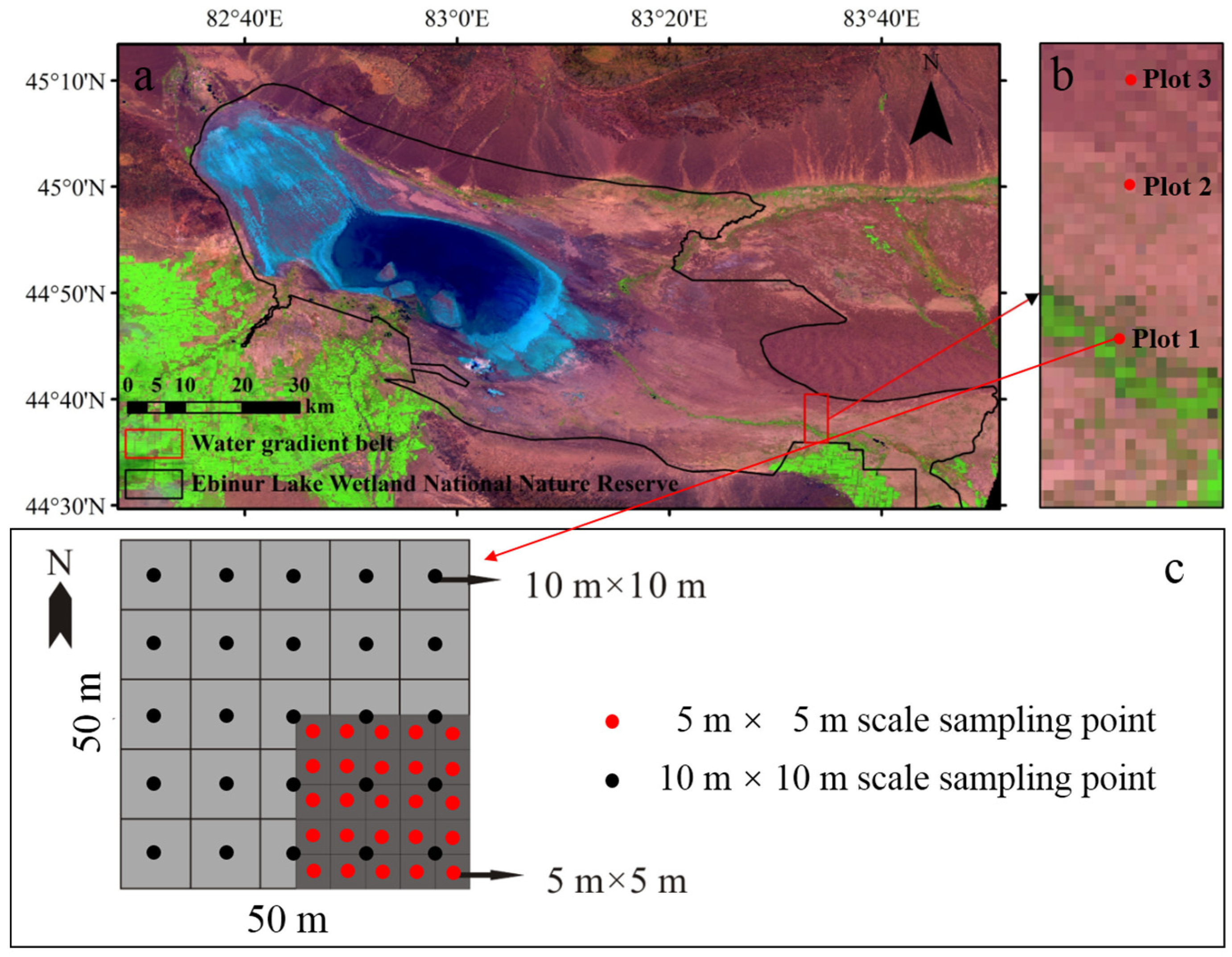
2.1. Study Site
2.2. Experimental Design
2.3. Rs and Soil Indicators
2.4. Community Survey and Plant Functional Trait Measurement
2.5. Statistical Analyses
3. Results
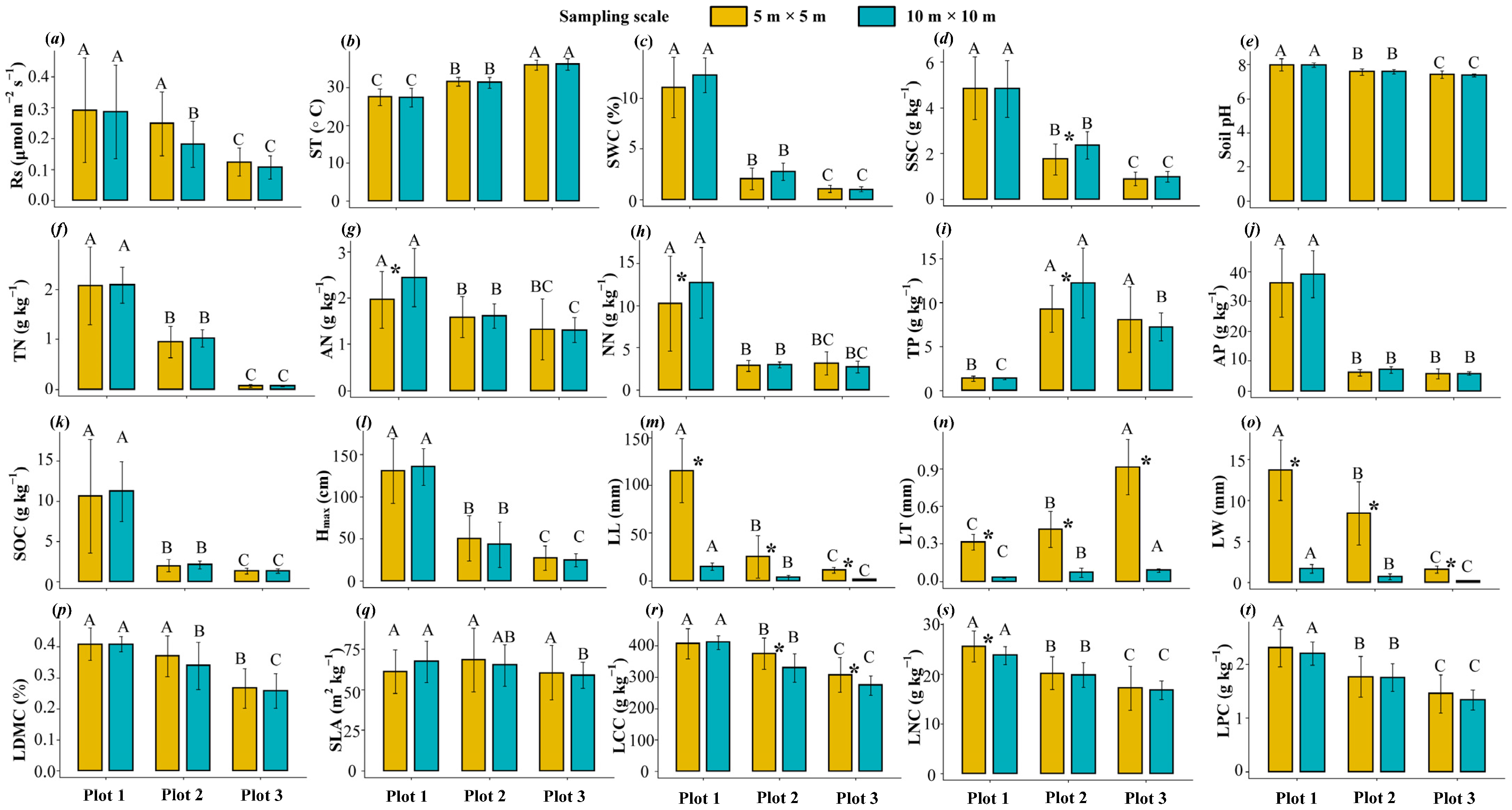
3.1. Changes in Rs and Environmental Factors
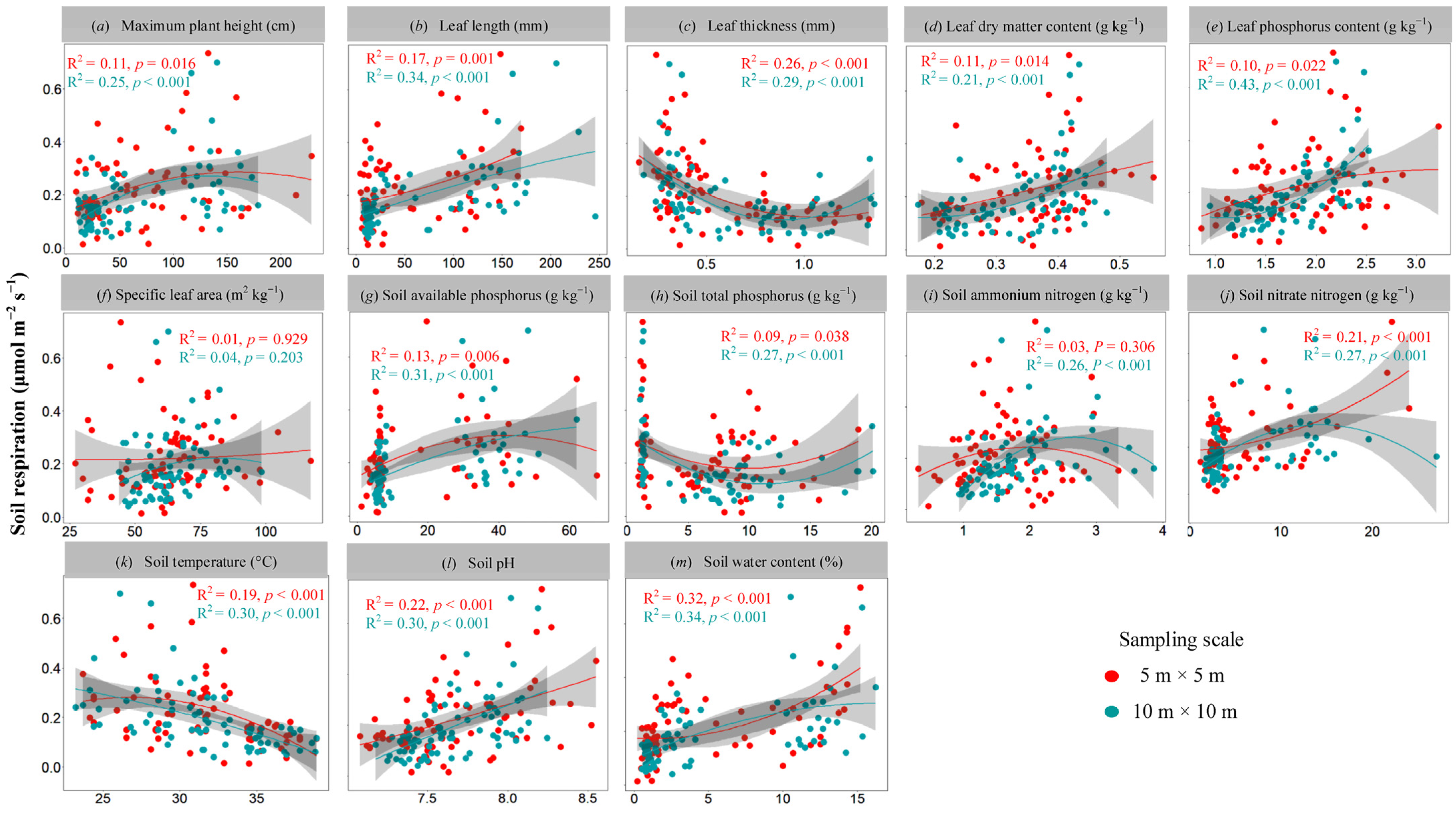
3.2. Multiple Regression Analysis of Rs and Environmental Factors in Specific Plot Types under Different Sampling Scales
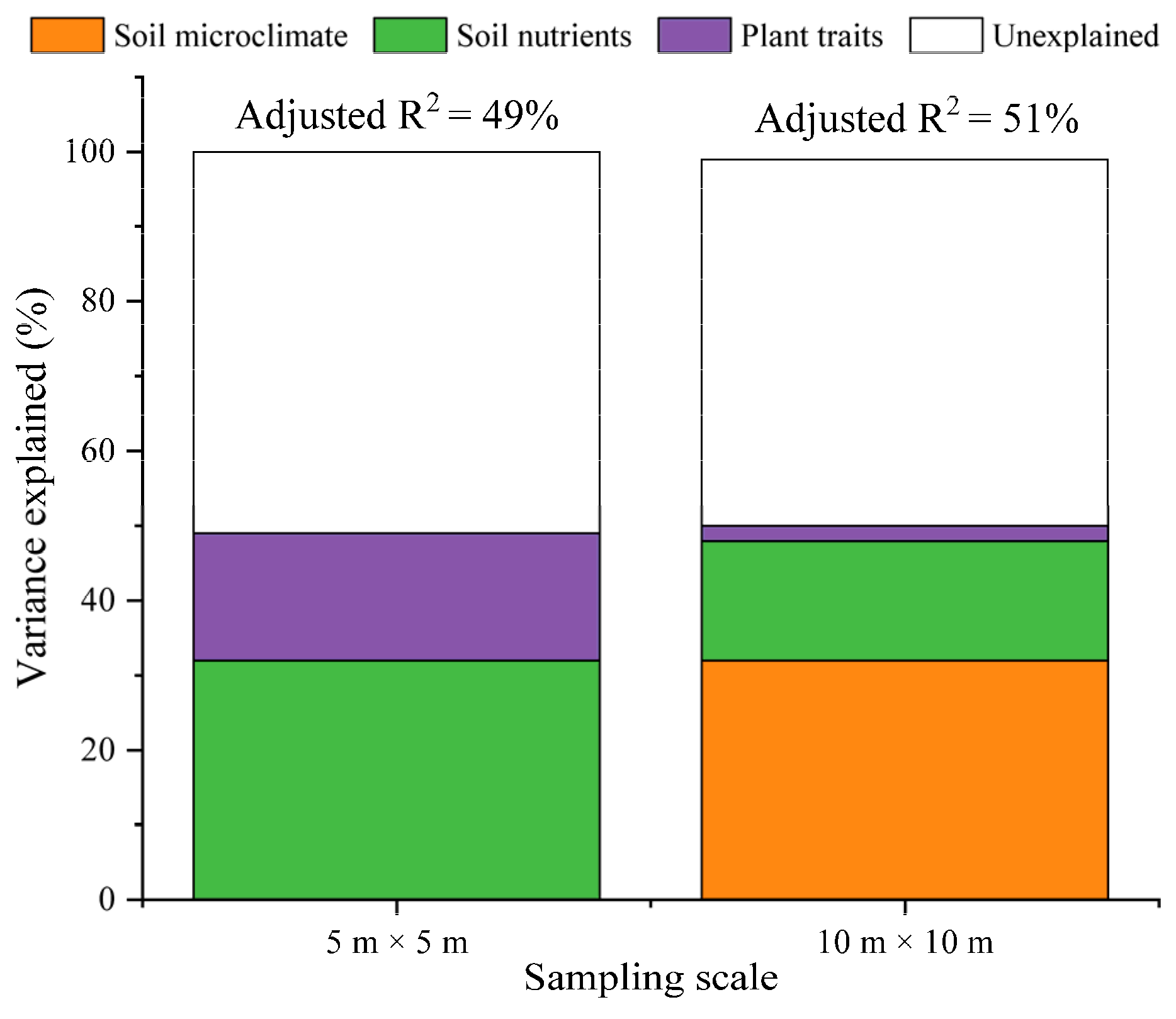
3.3. The Influencing Factors of Rs among Three Plot Types under Different Sampling Scales
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Hashimoto, S.; Carvalhais, N.; Ito, A.; Migliavacca, M.; Nishina, K.; Reichstein, M. Global spatiotemporal distribution of soil respiration modeled using a global database. Biogeosciences 2015, 12, 4121–4132. [Google Scholar] [CrossRef] [Green Version]
- Nunes, M.R.; Karlen, D.L.; Veum, K.S.; Moorman, T.B.; Cambardella, C.A. Biological soil health indicators respond to tillage intensity: A US meta-analysis. Geoderma 2020, 369, 114335. [Google Scholar] [CrossRef]
- Pries, C.E.H.; Castanha, C.; Porras, R.C.; Torn, M.S. The whole-soil carbon flux in response to warming. Science 2017, 355, 1420–1423. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lei, J.; Guo, X.; Zeng, Y.; Zhou, J.; Gao, Q.; Yang, Y. Temporal changes in global soil respiration since 1987. Nat. Commun. 2021, 12, 403. [Google Scholar] [CrossRef] [PubMed]
- Adachi, M.; Ito, A.; Yonemura, S.; Takeuchi, W. Estimation of global soil respiration by accounting for land-use changes derived from remote sensing data. J. Environ. Manag. 2017, 200, 97–104. [Google Scholar] [CrossRef] [Green Version]
- Fóti, S.; Balogh, J.; Herbst, M.; Papp, M.; Koncz, P.; Bartha, S.; Zimmermann, Z.; Komoly, C.; Szabó, G.; Margóczi, K.; et al. Meta-analysis of field scale spatial variability of grassland soil CO2 efflux: Interaction of biotic and abiotic drivers. CATENA 2016, 143, 78–89. [Google Scholar] [CrossRef] [Green Version]
- Schlesinger, W.H.; Andrews, J.A. Soil respiration and the global carbon cycle. Biogeochemistry 2000, 48, 7–20. [Google Scholar] [CrossRef]
- Hursh, A.; Ballantyne, A.; Cooper, L.; Maneta, M.; Kimball, J.; Watts, J. The sensitivity of soil respiration to soil temperature, moisture, and carbon supply at the global scale. Glob. Chang. Biol. 2016, 23, 2090–2103. [Google Scholar] [CrossRef]
- Tang, X.; Du, J.; Shi, Y.; Lei, N.; Chen, G.; Cao, L.; Pei, X. Global patterns of soil heterotrophic respiration—A meta-analysis of available dataset. CATENA 2020, 191, 104574. [Google Scholar] [CrossRef]
- Stephan, E.; Groffman, P.; Vidon, P.; Stella, J.C.; Endreny, T. Interacting drivers and their tradeoffs for predicting denitrification potential across a strong urban to rural gradient within heterogeneous landscapes. J. Environ. Manag. 2021, 294, 113021. [Google Scholar] [CrossRef]
- Riutta, T.; Kho, L.K.; Teh, Y.A.; Ewers, R.; Majalap, N.; Malhi, Y. Major and persistent shifts in below-ground carbon dynamics and soil respiration following logging in tropical forests. Glob. Chang. Biol. 2021, 27, 2225–2240. [Google Scholar] [CrossRef] [PubMed]
- Ouyang, W.; Lai, X.; Li, X.; Liu, H.; Lin, C.; Hao, F. Soil respiration and carbon loss relationship with temperature and land use conversion in freeze-thaw agricultural area. Sci. Total Environ. 2015, 533, 215–222. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Zou, J.; Dang, S.; Osborne, B.; Ren, Y.; Ju, X. Topography modifies the effect of land-use change on soil respiration: A meta-analysis. Ecosphere 2021, 12, e03845. [Google Scholar] [CrossRef]
- Han, G.; Yu, J.; Li, H.; Yang, L.; Wang, G.; Mao, P.; Gao, Y. Winter Soil Respiration from Different Vegetation Patches in the Yellow River Delta, China. Environ. Manag. 2012, 50, 39–49. [Google Scholar] [CrossRef]
- Jian, J.; Steele, M.K.; Zhang, L.; Bailey, V.L.; Zheng, J.; Patel, K.F.; Bond-Lamberty, B.P. On the use of air temperature and precipitation as surrogate predictors in soil respiration modelling. Eur. J. Soil Sci. 2021, 73, e13149. [Google Scholar] [CrossRef]
- Darenova, E.; Čater, M. Different Structure of Sessile Oak Stands Affects Soil Moisture and Soil CO2 Efflux. For. Sci. 2018, 64, 340–348. [Google Scholar] [CrossRef]
- Cai, Y.; Nishimura, T.; Ida, H.; Hirota, M. Spatial variation in soil respiration is determined by forest canopy structure through soil water content in a mature beech forest. For. Ecol. Manag. 2021, 501, 119673. [Google Scholar] [CrossRef]
- Lee, J.-S. Relationship of root biomass and soil respiration in a stand of deciduous broadleaved trees—A case study in a maple tree. J. Ecol. Environ. 2018, 42, 19. [Google Scholar] [CrossRef]
- Wang, J.; Teng, D.; He, X.; Qin, L.; Yang, X.; Lv, G. Spatial non-stationarity effects of driving factors on soil respiration in an arid desert region. CATENA 2021, 207, 105617. [Google Scholar] [CrossRef]
- Tang, Y.-S.; Wang, L.; Jia, J.-W.; Fu, X.-H.; Le, Y.-Q.; Chen, X.-Z.; Sun, Y. Response of soil microbial community in Jiuduansha wetland to different successional stages and its implications for soil microbial respiration and carbon turnover. Soil Biol. Biochem. 2011, 43, 638–646. [Google Scholar] [CrossRef]
- Yang, J.; Zhan, C.; Li, Y.; Zhou, D.; Yu, Y.; Yu, J. Effect of salinity on soil respiration in relation to dissolved organic carbon and microbial characteristics of a wetland in the Liaohe River estuary, Northeast China. Sci. Total Environ. 2018, 642, 946–953. [Google Scholar] [CrossRef] [PubMed]
- Singh, R.; Singh, A.K.; Singh, S.; Srivastava, P.; Singh, H.; Raghubanshi, A.S. Geomorphologic heterogeneity influences dry-season soil CO2 efflux by mediating soil biophysical variables in a tropical river valley. Aquat. Sci. 2019, 81, 43. [Google Scholar] [CrossRef]
- Raich, J.W.; Schlesinger, W.H. The global carbon dioxide flux in soil respiration and its relationship to vegetation and climate. Tellus B Chem. Phys. Meteorol. 1992, 44, 81–99. [Google Scholar] [CrossRef] [Green Version]
- Tang, J.; Baldocchi, D.D. Spatial-Temporal variation in soil respiration in an oak-grass savanna ecosystem in California and its partitioning into autotrophic and heterotrophic components. Biogeochemistry 2005, 73, 183–207. [Google Scholar] [CrossRef]
- Xu, M.; Qi, Y. Soil-surface CO2 efflux and its spatial and temporal variations in a young ponderosa pine plantation in northern California. Glob. Chang. Biol. 2001, 7, 667–677. [Google Scholar] [CrossRef] [Green Version]
- Janssens, I.A.; Pilegaard, K. Large seasonal changes in Q 10 of soil respiration in a beech forest. Glob. Chang. Biol. 2003, 9, 911–918. [Google Scholar] [CrossRef]
- Tang, J.; Baldocchi, D.; Qi, Y.; Xu, L. Assessing soil CO2 efflux using continuous measurements of CO2 profiles in soils with small solid-state sensors. Agric. For. Meteorol. 2003, 118, 207–220. [Google Scholar] [CrossRef]
- Metcalfe, D.B.; Fisher, R.A.; Wardle, D.A. Plant communities as drivers of soil respiration: Pathways, mechanisms, and significance for global change. Biogeosciences 2011, 8, 2047–2061. [Google Scholar] [CrossRef] [Green Version]
- Högberg, P.; Nordgren, A.; Ågren, G.I. Carbon allocation between tree root growth and root respiration in boreal pine forest. Oecologia 2002, 132, 579–581. [Google Scholar] [CrossRef]
- Raich, J.W.; Tufekciogul, A. Vegetation and soil respiration: Correlations and controls. Biogeochemistry 2000, 48, 71–90. [Google Scholar] [CrossRef]
- Kuzyakov, Y.; Gavrichkova, O. REVIEW: Time lag between photosynthesis and carbon dioxide efflux from soil: A review of mechanisms and controls. Glob. Chang. Biol. 2010, 16, 3386–3406. [Google Scholar] [CrossRef]
- Atkin, O.K.; Edwards, E.J.; Loveys, B.R. Response of root respiration to changes in temperature and its relevance to global warming. New Phytol. 2000, 147, 141–154. [Google Scholar] [CrossRef]
- Högberg, P.; Nordgren, A.; Buchmann, N.; Taylor, A.F.S.; Ekblad, A.; Högberg, M.N.; Nyberg, G.; Ottosson-Löfvenius, M.; Read, D.J. Large-scale forest girdling shows that current photosynthesis drives soil respiration. Nature 2001, 411, 789–792. [Google Scholar] [CrossRef] [PubMed]
- Moyano, F.E.; Kutsch, W.L.; Schulze, E.-D. Response of mycorrhizal, rhizosphere and soil basal respiration to temperature and photosynthesis in a barley field. Soil Biol. Biochem. 2007, 39, 843–853. [Google Scholar] [CrossRef]
- De Deyn, G.B.; Cornelissen, J.H.C.; Bardgett, R.D. Plant functional traits and soil carbon sequestration in contrasting biomes. Ecol. Lett. 2008, 11, 516–531. [Google Scholar] [CrossRef]
- Kumari, T.; Singh, R.; Verma, P.; Raghubanshi, A.S. Monsoon-phase regulates the decoupling of auto- and heterotrophic respiration by mediating soil nutrient availability and root biomass in tropical grassland. CATENA 2021, 209, 105808. [Google Scholar] [CrossRef]
- Shen, Y.; Gilbert, G.S.; Li, W.; Fang, M.; Lu, H.; Yu, S. Linking Aboveground Traits to Root Traits and Local Environment: Implications of the Plant Economics Spectrum. Front. Plant Sci. 2019, 10, 1412. [Google Scholar] [CrossRef]
- Mao, W.; Felton, A.J.; Ma, Y.; Zhang, T.; Sun, Z.; Zhao, X.; Smith, M.D. Relationships between aboveground and belowground trait responses of a dominant plant species to alterations in watertable depth. Land Degrad. Dev. 2018, 29, 4015–4024. [Google Scholar] [CrossRef]
- Raich, J.W. Temporal patterns of soil respiration in tropical forest plantations in lowland Costa Rica. In Proceedings of the 96th ESA Annual Meeting, Austin, TX, USA, 7–12 August 2011. [Google Scholar]
- Jiang, Y.; Huang, X.; Zhang, X.; Zhang, X.; Zhang, Y.; Zheng, C.; Deng, A.; Zhang, J.; Wu, L.; Hu, S.; et al. Optimizing rice plant photosynthate allocation reduces N2O emissions from paddy fields. Sci. Rep. 2016, 6, 29333. [Google Scholar] [CrossRef] [Green Version]
- Martin, J.G.; Bolstad, P.V. Variation of soil respiration at three spatial scales: Components within measurements, intra-site variation and patterns on the landscape. Soil Biol. Biochem. 2009, 41, 530–543. [Google Scholar] [CrossRef]
- Chen, Q.; Wang, Q.; Han, X.; Wan, S.; Li, L. Temporal and spatial variability and controls of soil respiration in a temperate steppe in northern China. Glob. Biogeochem. Cycles 2010, 24, GB2010. [Google Scholar] [CrossRef]
- Buczko, U.; Bachmann, S.; Gropp, M.; Jurasinski, G.; Glatzel, S. Spatial variability at different scales and sampling requirements for in situ soil CO2 efflux measurements on an arable soil. CATENA 2015, 131, 46–55. [Google Scholar] [CrossRef]
- Wu, J.; Jones, B.; Li, H.; Loucks, O.L. Scaling and Uncertainty Analysis in Ecology: Methods and Applications; Springer: Dordrecht, The Netherlands, 2006. [Google Scholar]
- Jenerette, D.; Wu, J. On the Definitions of Scale. Bull. Ecol. Soc. Am. 2000, 81, 104–105. [Google Scholar] [CrossRef]
- Siefert, A.; Ravenscroft, C.; Althoff, D.; Alvarez-Yepiz, J.C.; Carter, B.E.; Glennon, K.L.; Heberling, M.; Jo, I.S.; Pontes, A.; Sauer, A.; et al. Scale dependence of vegetation-environment relationships: A meta-analysis of multivariate data. J. Veg. Sci. 2012, 23, 942–951. [Google Scholar] [CrossRef]
- Tamme, R.; Hiiesalu, I.; Laanisto, L.; Szava-Kovats, R.; Pärtel, M. Environmental heterogeneity, species diversity and co-existence at different spatial scales. J. Veg. Sci. 2010, 21, 796–801. [Google Scholar] [CrossRef]
- Kosugi, Y.; Mitani, T.; Itoh, M.; Noguchi, S.; Tani, M.; Matsuo, N.; Takanashi, S.; Ohkubo, S.; Nik, A.R. Spatial and temporal variation in soil respiration in a Southeast Asian tropical rainforest. Agric. For. Meteorol. 2007, 147, 35–47. [Google Scholar] [CrossRef]
- Zhang, M.; Li, G.; Liu, B.; Liu, J.; Wang, L.; Wang, D. Effects of herbivore assemblage on the spatial heterogeneity of soil nitrogen in eastern Eurasian steppe. J. Appl. Ecol. 2020, 57, 1551–1560. [Google Scholar] [CrossRef]
- Gong, G.L.X. Species diversity and dominant species’ niches of eremophyte communities of the Tugai forest in the Ebinur basin of Xinjiang, China. Biodivers. Sci. 2017, 25, 29–35. [Google Scholar] [CrossRef] [Green Version]
- Zerbe, S.; Halik, Ü.; Küchler, J. Urban greening in the oases of continental arid Southern Xinjiang (NW China)—An interdisciplinary approach. Erde 2005, 136, 245–266. [Google Scholar]
- Thevs, N.; Zerbe, S.; Schnittler, M.; Abdusalih, N.; Succow, M. Structure, reproduction and flood-induced dynamics of riparian Tugai forests at the Tarim River in Xinjiang, NW China. Forestry 2008, 81, 45–57. [Google Scholar] [CrossRef] [Green Version]
- Zhang, X.; Yang, X.; Lv, G. Diversity patterns and response mechanisms of desert plants to the soil environment along soil water and salinity gradients. Acta Ecol. Sin. 2016, 36, 3206–3215. [Google Scholar] [CrossRef]
- Leemans, R.; Eickhout, B. Another reason for concern: Regional and global impacts on ecosystems for different levels of climate change. Glob. Environ. Chang. 2004, 14, 219–228. [Google Scholar] [CrossRef]
- Gong, Y.; Ling, H.; Lv, G.; Chen, Y.; Guo, Z.; Cao, J. Disentangling the influence of aridity and salinity on community functional and phylogenetic diversity in local dryland vegetation. Sci. Total Environ. 2018, 653, 409–422. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.-N.; Yang, X.-D.; Li, Y.; He, X.-M.; Lv, G.-H.; Yang, J.-J. Influence of edaphic factors on plant distribution and diversity in the arid area of Xinjiang, Northwest China. Arid Land Res. Manag. 2017, 32, 38–56. [Google Scholar] [CrossRef]
- Wang, H.; Cai, Y.; Yang, Q.; Gong, Y.; Lv, G. Factors that alter the relative importance of abiotic and biotic drivers on the fertile island in a desert-oasis ecotone. Sci. Total Environ. 2019, 697, 134096. [Google Scholar] [CrossRef]
- Shi, B.; Xu, W.; Zhu, Y.; Wang, C.; Loik, M.E.; Sun, W. Heterogeneity of grassland soil respiration: Antagonistic effects of grazing and nitrogen addition. Agric. For. Meteorol. 2019, 268, 215–223. [Google Scholar] [CrossRef]
- Barba, J.; Yuste, J.C.; Martínez-Vilalta, J.; Lloret, F. Drought-induced tree species replacement is reflected in the spatial variability of soil respiration in a mixed Mediterranean forest. For. Ecol. Manag. 2013, 306, 79–87. [Google Scholar] [CrossRef]
- Liu, C.; Song, X.; Wang, L.; Wang, D.; Zhou, X.; Liu, J.; Zhao, X.; Li, J.; Lin, H. Effects of grazing on soil nitrogen spatial heterogeneity depend on herbivore assemblage and pre-grazing plant diversity. J. Appl. Ecol. 2016, 53, 242–250. [Google Scholar] [CrossRef]
- Yan, J.-X.; Sun, Q.; Li, J.-J.; Li, H.-J. Effect of the Sampling Scale and Number on the Heterogeneity of Soil Respiration in a Mixed Broadleaf-conifer Forest. Huan Jing Ke Xue = Huanjing Kexue 2019, 40, 383–391. [Google Scholar]
- Jiang, Y.; Zhang, B.; Wang, W.; Li, B.; Wu, Z.; Chu, C. Topography and plant community structure contribute to spatial heterogeneity of soil respiration in a subtropical forest. Sci. Total Environ. 2020, 733, 139287. [Google Scholar] [CrossRef]
- Han, M.; Shi, B.; Jin, G. Spatial patterns of soil respiration in a spruce-fir valley forest, Northeast China. J. Soils Sediments 2018, 19, 10–22. [Google Scholar] [CrossRef]
- Darenova, E.; Čater, M. Effect of spatial scale and harvest on heterogeneity of forest floor CO2 efflux in a sessile oak forest. CATENA 2020, 188, 104455. [Google Scholar] [CrossRef]
- Russell, C.A.; Voroney, R.P. Carbon dioxide efflux from the floor of a boreal aspen forest. I. Relationship to environmental variables and estimates of C respired. Can. J. Soil Sci. 1998, 78, 301–310. [Google Scholar] [CrossRef]
- Rayment, M.; Jarvis, P. Temporal and spatial variation of soil CO2 efflux in a Canadian boreal forest. Soil Biol. Biochem. 2000, 32, 35–45. [Google Scholar] [CrossRef]
- Adachi, M.; Bekku, Y.S.; Konuma, A.; Kadir, W.R.; Okuda, T.; Koizumi, H. Required sample size for estimating soil respiration rates in large areas of two tropical forests and of two types of plantation in Malaysia. For. Ecol. Manag. 2005, 210, 455–459. [Google Scholar] [CrossRef]
- Shi, B.; Jin, G. Variability of soil respiration at different spatial scales in temperate forests. Biol. Fertil. Soils 2016, 52, 561–571. [Google Scholar] [CrossRef]
- Subke, J.-A.; Inglima, I.; Cotrufo, M.F. Trends and methodological impacts in soil CO2 efflux partitioning: A metaanalytical review. Glob. Chang. Biol. 2006, 12, 921–943. [Google Scholar] [CrossRef]
- Tang, X.; Pei, X.; Lei, N.; Luo, X.; Liu, L.; Shi, L.; Chen, G.; Liang, J. Global patterns of soil autotrophic respiration and its relation to climate, soil and vegetation characteristics. Geoderma 2020, 369, 114339. [Google Scholar] [CrossRef]
- Cassart, B.; Basia, A.A.; Jonard, M.; Ponette, Q. Functional traits drive the difference in soil respiration between Gilbertiodendron dewevrei monodominant forests patches and Scorodophloeus zenkeri mixed forests patches in the Central Congo basin. Plant Soil 2021, 460, 313–331. [Google Scholar] [CrossRef]
- Garnier, E.; Cortez, J.; Billès, G.; Navas, M.-L.; Roumet, C.; Debussche, M.; Laurent, G.; Blanchard, A.; Aubry, D.; Bellmann, A.; et al. Plant functional markers capture ecosystem properties during secondary succession. Ecology 2004, 85, 2630–2637. [Google Scholar] [CrossRef]
- Pei, Z.; Eichenberg, D.; Bruelheide, H.; Kröber, W.; Kühn, P.; Li, Y.; von Oheimb, G.; Purschke, O.; Scholten, T.; Buscot, F.; et al. Soil and tree species traits both shape soil microbial communities during early growth of Chinese subtropical forests. Soil Biol. Biochem. 2016, 96, 180–190. [Google Scholar] [CrossRef]
- Fortunel, C.; Garnier, E.; Joffre, R.; Kazakou, E.; Quested, H.; Grigulis, K.; Lavorel, S.; Ansquer, P.; Castro, H.; Cruz, P.; et al. Leaf traits capture the effects of land use changes and climate on litter decomposability of grasslands across Europe. Ecology 2009, 90, 598–611. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Conti, G.; Díaz, S. Plant functional diversity and carbon storage—An empirical test in semi-arid forest ecosystems. J. Ecol. 2012, 101, 18–28. [Google Scholar] [CrossRef]
- Buzzard, V.; Michaletz, S.T.; Deng, Y.; He, Z.; Ning, D.; Shen, L.; Tu, Q.; Van Nostrand, J.D.; Voordeckers, J.W.; Wang, J.; et al. Continental scale structuring of forest and soil diversity via functional traits. Nat. Ecol. Evol. 2019, 3, 1298–1308. [Google Scholar] [CrossRef]
- Fang, C.; Moncrieff, J.B.; Gholz, H.L.; Clark, K.L. Soil CO2 efflux and its spatial variation in a Florida slash pine plantation. Plant Soil 1998, 205, 135–146. [Google Scholar] [CrossRef]
- Frank, A. Carbon dioxide fluxes over a grazed prairie and seeded pasture in the Northern Great Plains. Environ. Pollut. 2001, 116, 397–403. [Google Scholar] [CrossRef]
- Xu, X.; Shi, Z.; Li, D.; Zhou, X.; Sherry, R.A.; Luo, Y. Plant community structure regulates responses of prairie soil respiration to decadal experimental warming. Glob. Chang. Biol. 2015, 21, 3846–3853. [Google Scholar] [CrossRef]
- Wofsy, S.C.; Harriss, R.C.; Kaplan, W.A. Carbon dioxide in the atmosphere over the Amazon Basin. J. Geophys. Res. Earth Surf. 1988, 93, 1377–1387. [Google Scholar] [CrossRef]
- Yan, L.; Chen, S.; Huang, J.; Lin, G. Water regulated effects of photosynthetic substrate supply on soil respiration in a semiarid steppe. Glob. Chang. Biol. 2010, 17, 1990–2001. [Google Scholar] [CrossRef]
- Liu, W.; Zhang, Z.; Wan, S. Predominant role of water in regulating soil and microbial respiration and their responses to climate change in a semiarid grassland. Glob. Chang. Biol. 2009, 15, 184–195. [Google Scholar] [CrossRef]
- Hartman, K.; Tringe, S.G. Interactions between plants and soil shaping the root microbiome under abiotic stress. Biochem. J. 2019, 476, 2705–2724. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schwen, A.; Jeitler, E.; Böttcher, J. Spatial and temporal variability of soil gas diffusivity, its scaling and relevance for soil respiration under different tillage. Geoderma 2015, 259, 323–336. [Google Scholar] [CrossRef]
- Pangle, R.; Seiler, J. Influence of seedling roots, environmental factors and soil characteristics on soil CO2 efflux rates in a 2-year-old loblolly pine (Pinus taeda L.) plantation in the Virginia Piedmont. Environ. Pollut. 2001, 116, S85–S96. [Google Scholar] [CrossRef]
- Yang, X.-D.; Ali, A.; Xu, Y.-L.; Jiang, L.-M.; Lv, G.-H. Soil moisture and salinity as main drivers of soil respiration across natural xeromorphic vegetation and agricultural lands in an arid desert region. CATENA 2019, 177, 126–133. [Google Scholar] [CrossRef]
- Raich, J.W.; Potter, C.S. Global patterns of carbon dioxide emissions from soils. Glob. Biogeochem. Cycles 1995, 9, 23–36. [Google Scholar] [CrossRef] [Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Source of Variation | df | Sum Square | Mean Square | F Value | p Value |
|---|---|---|---|---|---|
| Plot type | 2 | 0.76 | 0.38 | 32.08 | <0.001 |
| Sampling scale | 1 | 0.03 | 0.03 | 2.87 | 0.09 |
| Plot type: Sampling scale | 2 | 0.03 | 0.01 | 1.17 | 0.31 |
| Residuals | 144 | 1.71 | 0.01 | - | - |
| Parameter | Plot 1 | Plot 2 | Plot 3 | |||
|---|---|---|---|---|---|---|
| 5 m × 5 m | 10 m × 10 m | 5 m × 5 m | 10 m × 10 m | 5 m × 5 m | 10 m × 10 m | |
| Soil respiration | 58.0 | 52.8 | 41.9 | 41.7 | 37.0 | 35.4 |
| Maximum plant height | 29.3 | 16.0 | 53.8 | 62.4 | 55.4 | 31.9 |
| Leaf length | 29.0 | 24.5 | 88.3 | 81.5 | 29.3 | 25.0 |
| Leaf width | 26.8 | 32.3 | 46.3 | 37.1 | 29.6 | 18.1 |
| Leaf thickness | 20.0 | 14.4 | 34.9 | 55.3 | 24.1 | 17.0 |
| Leaf dry matter content | 12.8 | 6.1 | 17.7 | 22.4 | 23.9 | 21.4 |
| Specific leaf area | 21.9 | 19.0 | 28.6 | 19.6 | 27.8 | 13.5 |
| Leaf carbon content | 11.9 | 5.4 | 13.3 | 13.7 | 17.8 | 11.6 |
| Leaf nitrogen content | 12.3 | 7.7 | 16.2 | 12.7 | 26.0 | 11.2 |
| Leaf phosphorus content | 15.1 | 10.0 | 21.1 | 14.7 | 24.9 | 14.0 |
| Soil organic carbon | 27.1 | 14.1 | 52.7 | 31.0 | 35.6 | 25.2 |
| Soil water content | 8.0 | 8.8 | 3.5 | 4.3 | 3.7 | 4.1 |
| Soil temperature | 28.3 | 25.7 | 39.0 | 25.6 | 33.7 | 23.4 |
| Soil available phosphorus | 4.5 | 1.6 | 2.5 | 1.5 | 2.8 | 1.4 |
| Soil salinity content | 66.8 | 33.3 | 40.1 | 22.2 | 26.9 | 23.1 |
| Soil pH | 31.8 | 20.0 | 18.1 | 15.9 | 29.6 | 11.5 |
| Soil total phosphorus | 19.5 | 9.3 | 28.7 | 32.7 | 46.1 | 22.3 |
| Total nitrogen content | 37.5 | 17.0 | 33.9 | 17.1 | 48.8 | 27.2 |
| Soil ammonium nitrogen | 31.4 | 25.9 | 28.0 | 16.1 | 49.8 | 20.6 |
| Soil nitrate nitrogen | 55.4 | 33.1 | 22.3 | 10.9 | 43.6 | 25.8 |
| Plot | Sampling Scale | Variogram Model Type | Proportion (C/(C0 + C)) | Range (m) | R2 |
|---|---|---|---|---|---|
| Plot 1 | 5 m × 5 m | Spherical | 0.92 | 11.65 | 0.72 |
| 10 m × 10 m | Spherical | 0.98 | 15.45 | 0.94 | |
| Plot 2 | 5 m × 5 m | Linear | 0.28 | 16.30 | 0.36 |
| 10 m × 10 m | Gaussian | 0.97 | 19.68 | 1.00 | |
| Plot 3 | 5 m × 5 m | Linear | <0.25 | 16.30 | 0.24 |
| 10 m × 10 m | Linear | <0.25 | 32.60 | 0.64 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, J.; He, X.; Ma, W.; Li, Z.; Chen, Y.; Lv, G. Spatial Scale Effects of Soil Respiration in Arid Desert Tugai Forest: Responses to Plant Functional Traits and Soil Abiotic Factors. Forests 2022, 13, 1001. https://0-doi-org.brum.beds.ac.uk/10.3390/f13071001
Wang J, He X, Ma W, Li Z, Chen Y, Lv G. Spatial Scale Effects of Soil Respiration in Arid Desert Tugai Forest: Responses to Plant Functional Traits and Soil Abiotic Factors. Forests. 2022; 13(7):1001. https://0-doi-org.brum.beds.ac.uk/10.3390/f13071001
Chicago/Turabian StyleWang, Jinlong, Xuemin He, Wen Ma, Zhoukang Li, Yudong Chen, and Guanghui Lv. 2022. "Spatial Scale Effects of Soil Respiration in Arid Desert Tugai Forest: Responses to Plant Functional Traits and Soil Abiotic Factors" Forests 13, no. 7: 1001. https://0-doi-org.brum.beds.ac.uk/10.3390/f13071001