
Effect of Different Presowing Treatments to Break Seed Dormancy and Seed Collection Methods on the Germination of Dracaena steudneri Schweinf. Ex Engl.


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
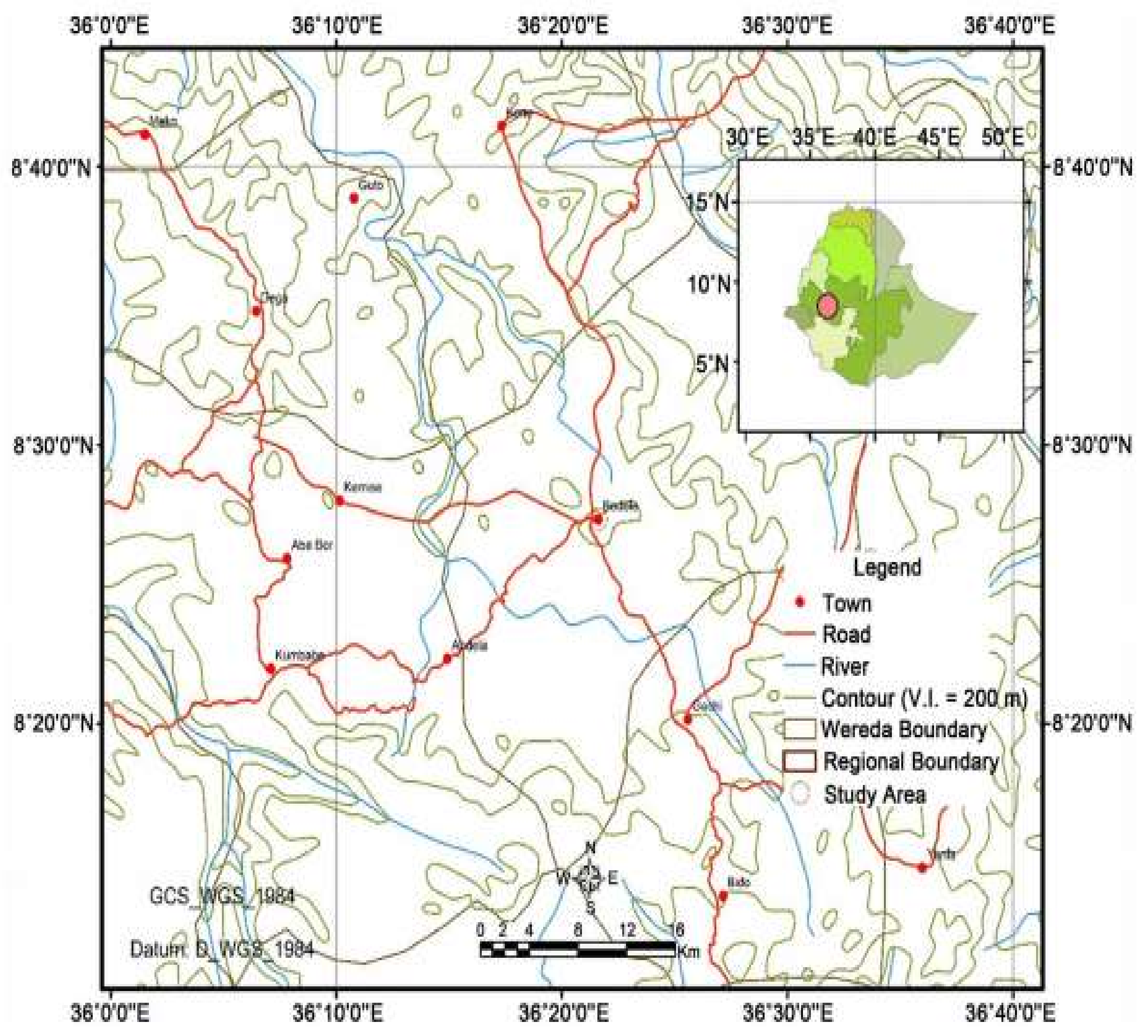
2.1. Seed Collection and Germination Experiment
2.2. Data Collection and Analysis
3. Results
3.1. Seed Germination in the Greenhouse
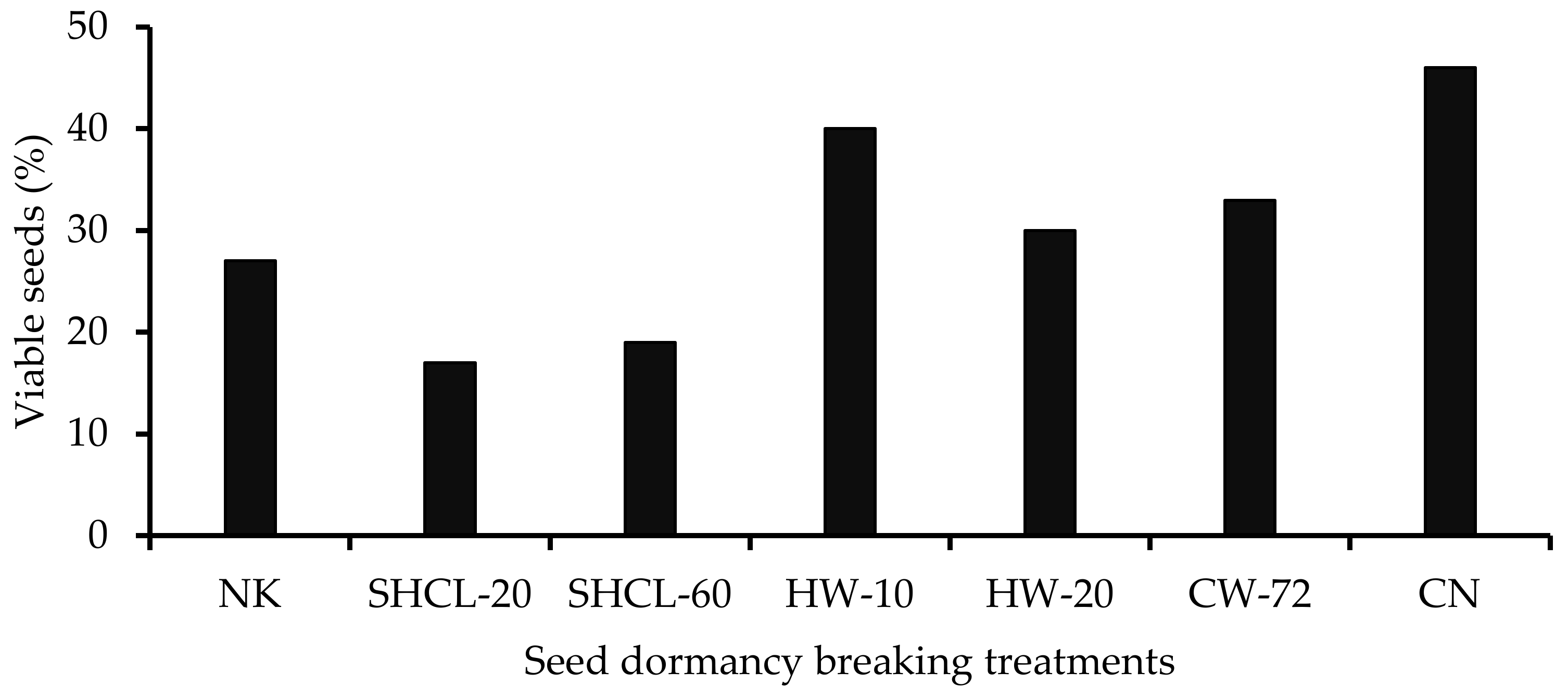
3.2. Seed Germination in the Laboratory
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- PL (The Plant List). Version 1.1. Published on the Internet. 2013. Available online: http://www.theplantlist.org/ (accessed on 1 January 2020).
- Damen, T.H.J.; van der Burg, W.J.; Wiland-Szymańska, J.; Sosef, M.S.M. Taxonomic novelties in African Dracaena (Dracaenaceae). Blumea 2018, 63, 31–53. [Google Scholar] [CrossRef]
- Govaerts, R.; Zonneveld, B.J.M.; Zona, S.A. World Checklist of Asparagaceae. Facilitated by the Royal Botanic Gardens, Kew. Published on the Internet. Available online: http://apps.kew.org/wcsp/ (accessed on 10 February 2017).
- APG (Angiosperm Phylogeny Group). An update of the Angiosperm Phylogeny Group classification for the orders and families of flowering plants: APG IV. Bot. J. Linn. Soc. 2016, 181, 1–20. [Google Scholar] [CrossRef] [Green Version]
- Chase, M.W.; Reveal, J.L.; Fay, M.F. A subfamilial classification for the expanded Asparagalean families Amaryllidaceae, Asparagaceae and Xanthorrhoeaceae. Bot. J. Linn. Soc. 2009, 161, 132–136. [Google Scholar] [CrossRef] [Green Version]
- Singh, H.P.; Dadlani, N.K. Current Status of Floriculture—National and International Scenario; Commercial Floriculture; Malhotra Publishing House: New Delhi, India, 2000. [Google Scholar]
- LEI; CBS. Land- en tuinbouwcijfers [Agricultural and Horticultural Figures]; Landbouw Economisch Instituut (LEI): The Hague, The Netherlands, 2012; p. 260. [Google Scholar]
- Staner, P.; Boutique, R. Matériaux pour l’étude des plantes medicinales indigènes du Congo Belge. In Memoire de l’Institute Royale College de Belge; Institut Royal Colonial Belge Section des Sciences Naturelles et Médicales: Bruxelles, Belgium, 1937. [Google Scholar]
- Sheridan, M. Tanzanian ritual perimetrics and African landscapes: The case of Dracaena. Int. J. Afr. Hist. Stud. 2008, 41, 491–521. [Google Scholar]
- Bekele-Tesemma, A.; Beánie, A.; Tengnas, B. Useful Trees and Shrubs for Ethiopia: Identification, Propagation and Management for 17 Agro-Climatic Zones; Regional Land Management Unit: Nairobi, Kenya, 2007. [Google Scholar]
- Awas, T.; Demissew, S. Ethnobotanical study of medicinal plants in Kafficho people, southwestern Ethiopia. In Proceedings of the 16th International Conference of Ethiopian Studies; Ege, S., Aspen, H., Tefera, B., Bekele, S., Eds.; Trondheim University: Trondheim, Norway, 2009. [Google Scholar]
- Etana, B. Ethnobotanical Study of Traditional Medicinal Plants of Goma Wereda, Jima Zone of Oromia Region, Ethiopia. Master’s Thesis, Addis Abeba University, Addis Abeba, Ethiopia, 2010; p. 102. [Google Scholar]
- Moshi, M.J.; Otien, D.F.; Weisheit, A. Ethnomedicine of the Kagera Region, north western Tanzania. Part 3: Plants used in traditional medicine in Kikuku village, Muleba District. J. Ethnobiol. Ethnomed. 2012, 8, 1–11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kadima, N.J.; Marhegeko, A.B.; Kasali, F.M.; Mugaruka, N.J. Medicinal Plants Used in Alternative Medicine to Treat Cancer in Bukavu. Eur. J. Med. Plants 2016, 12, 1–13. [Google Scholar] [CrossRef]
- Adia, M.M.; Anywar, G.; Byamukama, R.; Kamatenesi-Mugisha, M.; Sekagya, Y.; Kakudidi, E.K.; Kiremire, B.T. Medicinal plants used in malaria treatment by Prometra herbalists in Uganda. J. EthnoPharmacol. 2014, 155, 580–588. [Google Scholar] [CrossRef]
- Alelign, A.; Teketay, D.; Yemshaw, Y.; Edwards, S. Diversity and status of regeneration of woody plants on the peninsula of Zegie, northwestern Ethiopia. Trop. Ecol. 2007, 48, 37–49. [Google Scholar]
- Alem, S.; Woldemariam, T. A comparative assessment on regeneration status of indigenous woody plants in Eucalyptus grandis plantation and adjacent natural forest. J. For. Resour. 2009, 20, 31–36. [Google Scholar] [CrossRef]
- Assefa, A.; Demissew, S.; Woldu, Z. Floristic composition, structure and regeneration status of Masha forest, south-west Ethiopia. Afr. J. Ecol. 2013, 52, 151–162. [Google Scholar] [CrossRef]
- Nune, S. Flora Biodiversity Assessment in Bonga, Boginda and Mankira Forest, Kafa, Ethiopia. PPP Project, Addis Abeba. 2008. Available online: https://dokumen.tips/documents/flora-biodiversity-assessment-in-bonga-boginda-and-biodiversity-assessment.html (accessed on 15 January 2020).
- Nonogaki, H. Seed Biology Updates: Highlights and New Discoveries in Seed Dormancy and Germination Research. Front. Plant Sci. 2017, 8, 524. [Google Scholar] [CrossRef] [Green Version]
- Nguyen, T.P.; Keizer, P.; van Eeuwijk, F.; Smeekens, S.; Bentsink, L. Natural Variation for Seed Longevity and Seed Dormancy are Negatively Correlated in Arabidopsis. Plant Physiol. 2012, 160, 2083–2092. [Google Scholar] [CrossRef] [Green Version]
- Baskin, J.M.; Baskin, C.C. A classification system for seed dormancy. Seed Sci. Res. 2004, 14, 1–16. [Google Scholar] [CrossRef] [Green Version]
- Benech-Arnold, R.I.; Sanches, R.A.; Forcella, F.; Kruk, B.C.; Ghersa, C.M. Environmental control of dormancy in weed seed bank in soil. Field Crops Res. 2000, 67, 105–122. [Google Scholar] [CrossRef]
- Yuningsih, A.F.V.; Wahyuni, S. Effective Methods for Dormancy Breaking of 15 New-Improved Rice Varieties to Enhance the Validity of Germination Test. In Proceedings of the International Seminar on Promoting Local Resources for Food and Health, Bengkulu, Indonesia, 12–13 October 2015. [Google Scholar]
- Hartmann, H.T.; Kester, D.E.; Davies, F., Jr.; Geneve, R.L. Plant Propagation Principles and Practices, 8th ed.; Prentice Hall Inc.: Hoboken, NJ, USA, 2010; 928p. [Google Scholar]
- Sideman, B. Starting Plants Indoors from Seed: Fact sheet. In UNH Cooperative Extension Programs; State Office, Taylor Hall: Durham, UK, 2017. [Google Scholar]
- Kaur, A.; Singh, A.; Monga, R. Seed Germination Enhancement through Breaking Seed Dormancy: A Review in Tropical and Temperate Tree Species. Int. J. Curr. Microbiol. Appl. Sci. 2020, 9, 1673–1688. [Google Scholar] [CrossRef]
- Graeber, K.; Nakabayashi, K.; Miatton, E.; Leubner-Metzger, G.; Soppe, W.J. Molecular mechanisms of seed dormancy. Plant Cell Environ. 2012, 35, 1769–1786. [Google Scholar] [CrossRef]
- Tang, Y.; Zhang, K.; Zhang, Y.; Tao, J. Dormancy-Breaking and Germination Requirements for Seeds of Sorbus alnifolia (Siebold & Zucc.) K.Koch (Rosaceae), a Mesic Forest Tree with High Ornamental Potential. Forests 2019, 10, 319. [Google Scholar] [CrossRef] [Green Version]
- Iralu, V.; Upadhaya, K. Seed dormancy, germination and seedling characteristics of Elaeocarpus prunifolius Wall. ex Müll. Berol.: A threatened tree species of north-eastern India. N. Z. J. For. Sci. 2018, 48, 1–10. [Google Scholar] [CrossRef]
- Di-Tommaso, A.; Nurse, R.E. Impact of sodium hypochlorite concentration and exposure period on germination and radicle elongation of three annual weed species. Seed Sci. Technol. 2004, 32, 377–391. [Google Scholar] [CrossRef] [Green Version]
- Krishnapillay, B.; Tompsett, P.B. Seed handling. In A Review of Taxonomy, Ecology and Silviculture Dipterocarps; Appana, S., Turnbull, J.M., Eds.; Center for International Forestry Research Bogor: Bogor, Indonesia, 1998. [Google Scholar]
- Adolt, R. Proposal of Principles of Dragon Trees Gene Resources Protection in Forests on Socotra and Canary Islands. Master’s Thesis, Mendel University of Agriculture and Forestry Brno, Brno, Czech Republic, 2001; 247. (In Czech). [Google Scholar]
- Bauerová, L.; Alem Munie, S.; Houšková, K.; Habrová, H. Germination of Dracaena cinnabari Balf.f. Seeds under Controlled Temperature Conditions. Forests 2020, 11, 521. [Google Scholar] [CrossRef]
- Yaman, F.; Kahrıman, F. Classification of viable/non-viable seeds of specialty maize genotypes using spectral and image data plus morphological features. J. Crop Improv. 2022, 36, 285–300. [Google Scholar] [CrossRef]
- Bradbeer, J.W. Seed Viability and Vigour. In: Seed Dormancy and Germination. In Tertiary Level Biology; Springer: Boston, MA, USA, 1988. [Google Scholar] [CrossRef]
- Putri, K.P.; Yuniarti, N.; Aminah, A.; Suita, E.; Sudrajat, D.J.; Syamsuwida, D. Seed handling of specific forest tree species: Recalcitrant and intermediate seed. IOP Conf. Ser. Earth Environ. Sci. 2020, 522, 012015. [Google Scholar] [CrossRef]
- Kheloufi, A.; Boukhecha, M.; Ouachi, A. Effect of pre-soaking, substrate and light availability on seed germination and seedling establishment of Dracaena draco (L.) L., a threatened tree species. Reforesta 2020, 9, 20–29. [Google Scholar] [CrossRef]
- Chan-Chin, D.; Govinden-Soulange, J. Germination profile of selected plants from Mauritius—Towards a conservation strategy. Seed Sci. Technol. 2015, 43, 536–540. [Google Scholar] [CrossRef]
- Dewir, Y.H.; Aldubai, A.A.; Al-Obeed, R.S.; El-Hendawy, S.; Seliem, M.K.; Al-Harbi, K.R. Micro-propagation to conserve the endangered Gabal Elba Dragon Tree (Dracaena ombet Heuglin ex Kotschy & Peyr). Hortscience 2019, 54, 162–166. [Google Scholar]
- Vinterhalter, D.; Vinterhalter, B. Micropropagation of Dracaena Species. In High-Tech and Micropropagation VI. Biotechnology in Agriculture and Forestry; Bajaj, Y.P.S., Ed.; Springer: Berlin/Heidelberg, Germany, 1997. [Google Scholar] [CrossRef]
- Attia, F.A.K.; El-Sallami, I.H.; Gad, M.M.; Abudl-Hafeez, E.Y. Factors Influencing Rootability of Dracaena Cuttings. Assiut J. Agric. Sci. 2015, 46, 71–79. [Google Scholar]
- Blanco, M.; Valverde, R.; Gomez, L. Micropropagation of Dracaena deremensis. Agron. Costarric. 2004, 28, 7–15. [Google Scholar]
- Jazib, A.; Hossain, M.T.; Raju, R.I. Clonal propagation of Dracaena fragrans cv. Victoria through tissue cult ure technology. Jahangirnagar Univ. J. Biol. Sci. 2019, 8, 1–11. [Google Scholar] [CrossRef]
- Aslam, J.; Mujib, A.; Sharma, M.P. In vitro micropropagation of Dracaena sanderiana Sander ex Mast: An important indoor ornamental plant. Saudi J. Biol. Sci. 2013, 20, 63–68. [Google Scholar] [CrossRef] [Green Version]
- Liu, J.; Deng, M.; Henny, R.J.; Chen, J.; Xie, J. Regeneration of Dracaena surculosa through indirect shoot organogenesis. HortScience 2010, 45, 1250–1254. [Google Scholar] [CrossRef] [Green Version]
- Badawy, E.M.; Habib, A.M.A.; El-Bana, A.; Yosry, G.M. Propagation of Dracaena fragrans plants by tissue culture technique. Arab. J. Biotechnol. 2005, 8, 329–342. [Google Scholar]









Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Munie, S.A.; Habrová, H.; Houšková, K.; Karas, L. Effect of Different Presowing Treatments to Break Seed Dormancy and Seed Collection Methods on the Germination of Dracaena steudneri Schweinf. Ex Engl. Forests 2022, 13, 1232. https://0-doi-org.brum.beds.ac.uk/10.3390/f13081232
Munie SA, Habrová H, Houšková K, Karas L. Effect of Different Presowing Treatments to Break Seed Dormancy and Seed Collection Methods on the Germination of Dracaena steudneri Schweinf. Ex Engl. Forests. 2022; 13(8):1232. https://0-doi-org.brum.beds.ac.uk/10.3390/f13081232
Chicago/Turabian StyleMunie, Shiferaw Alem, Hana Habrová, Kateřina Houšková, and Lukáš Karas. 2022. "Effect of Different Presowing Treatments to Break Seed Dormancy and Seed Collection Methods on the Germination of Dracaena steudneri Schweinf. Ex Engl." Forests 13, no. 8: 1232. https://0-doi-org.brum.beds.ac.uk/10.3390/f13081232