
The Climate-Growth Relationship between Picea smithiana (Wall.) Boiss. and Abies pindrow (Royle ex D.Don) Royle along the Latitudinal Gradient in Northern Pakistan

 ,
,  ,
,  and
and
Abstract
:1. Introduction
2. Methods
2.1. Study Site and Sample Collection
2.2. Tree-Ring Data and Chronology Development
2.3. Climatic Data and Statistical Analysis
3. Results
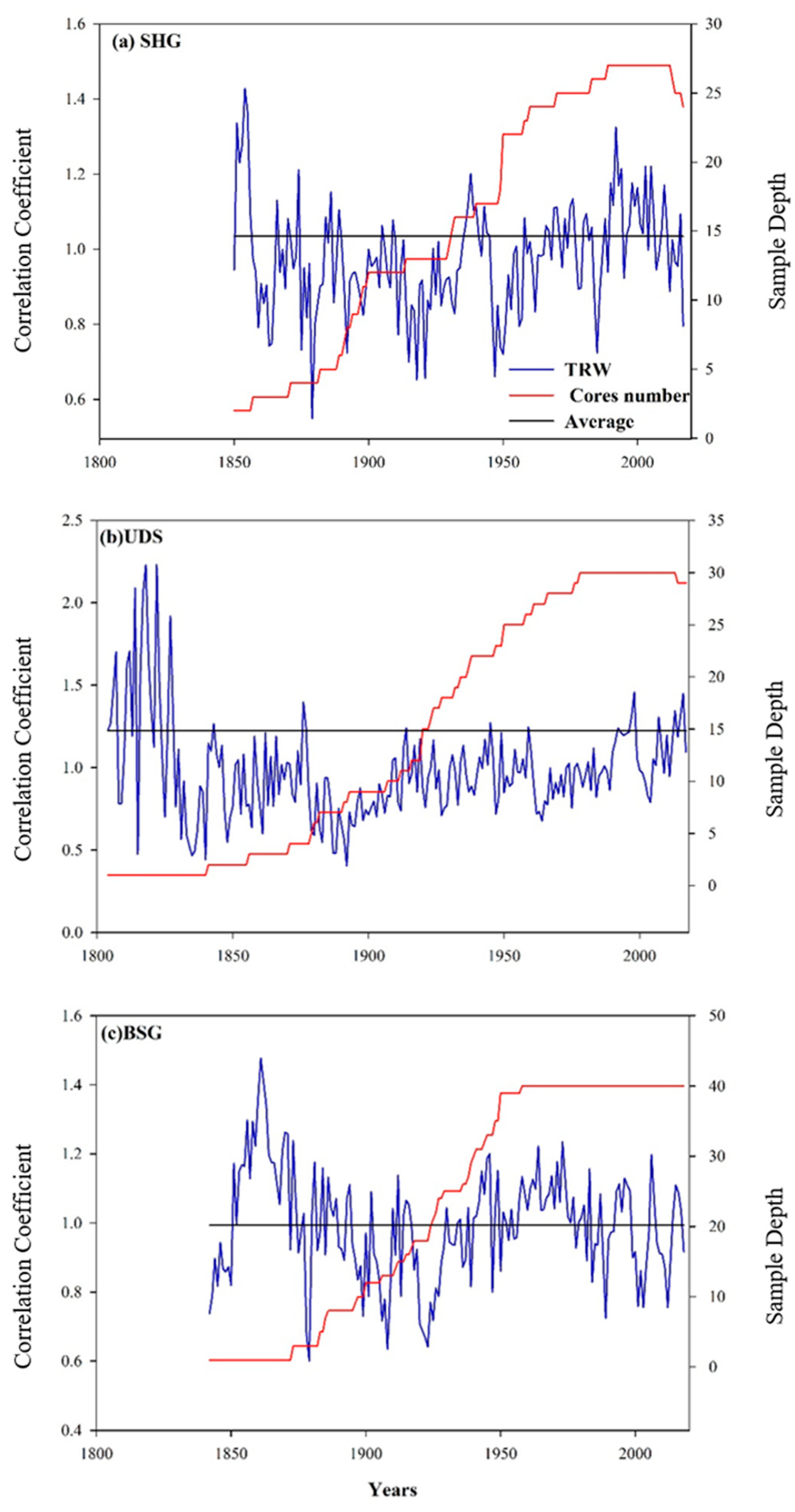
3.1. Tree-Ring Width Chronologies
3.2. Growth–Climate Relationship
3.3. Dendroclimatological Relationship before and after Rapid Warming
3.4. Dendroclimatological Relationship with Latitude
4. Discussion
4.1. Tree-Ring Chronology Characteristics
4.2. Climate Growth Relationship of Fir and Picea smithiana
4.3. Dendroclimatological Relationship with Latitude
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Jandl, R.; Spathelf, P.; Bolte, A.; Prescott, C.E. Forest adaptation to climate change—Is non-management an option? Ann. For. Sci. 2019, 76, 48. [Google Scholar] [CrossRef]
- Stefanidis, S.; Alexandridis, V. Precipitation and potential evapotranspiration temporal variability and their relationship in two forest ecosystems in greece. Hydrology 2021, 8, 160. [Google Scholar] [CrossRef]
- Zindros, A.; Radoglou, K.; Milios, E.; Kitikidou, K. Tree line shift in the Olympus mountain (Greece) and climate change. Forests 2020, 11, 985. [Google Scholar] [CrossRef]
- IPCC. Climate Change 2021: The physical science basis. In Contribution of Working Group I to the Sixth Assessment Report of the Intergovernmental Panel on Climate Change; Cambridge University Press: Cambridge, UK, 2021. [Google Scholar]
- Pretzsch, H.; Biber, P.; Schütze, G.; Uhl, E.; Rötzer, T. Forest stand growth dynamics in Central Europe have accelerated since 1870. Nat. Commun. 2014, 5, 4967. [Google Scholar] [CrossRef] [PubMed]
- Reed, C.C.; Ballantyne, A.P.; Cooper, L.A.; Sala, A. Limited evidence for CO2-related growth enhancement in northern Rocky Mountain lodgepole pine populations across climate gradients. Glob. Chang. Biol. 2018, 24, 3922–3937. [Google Scholar] [CrossRef] [PubMed]
- Rogers, B.M.; Jantz, P.; Goetz, S.J. Vulnerability of eastern US tree species to climate change. Glob. Chang. Biol. 2017, 23, 3302–3320. [Google Scholar] [CrossRef] [PubMed]
- Kirschbaum, M.U. Will changes in soil organic carbon act as a positive or negative feedback on global warming? Biogeochemistry 2000, 48, 21–51. [Google Scholar] [CrossRef]
- Boisvenue, C.; Running, S.W. Impacts of climate change on natural forest productivity–evidence since the middle of the 20th century. Glob. Chang. Biol. 2006, 12, 862–882. [Google Scholar] [CrossRef]
- Bukhari, S.S.B.; Bajwa, G.A. Development of National Response Strategy to Combat Impacts of Climate Change on Forest of Pakistan; Pakistan Forest Inistitute Peshawar: Peshawar, Pakistan, 2012. [Google Scholar]
- Camarero, J.J.; Fajardo, A. Poor acclimation to current drier climate of the long-lived tree species Fitzroyacupressoides in the temperate rainforest of southern Chile. Agric. For. Meteorol. 2017, 239, 141–150. [Google Scholar] [CrossRef]
- Liu, B.; Liang, E.; Liu, K.; Camarero, J.J. Species-and elevation-dependent growth responses to climate warming of mountain forests in the Qinling Mountains, central China. Forests 2018, 9, 248. [Google Scholar] [CrossRef]
- Davi, N.K.; D’Arrigo, R.; Jacoby, G.; Cook, E.; Anchukaitis, K.; Nachin, B.; Rao, M.; Leland, C. A long-term context (931–2005 ce) for rapid warming over Central Asia. Quat. Sci. Rev. 2015, 121, 89–97. [Google Scholar] [CrossRef]
- Shi, C.; Masson-Delmotte, V.; Daux, V.; Li, Z.; Carré, M.; Moore, J.C. Unprecedented recent warming rate and temperature variability over the east Tibetan Plateau inferred from Alpine treeline dendrochronology. Clim. Dyn. 2015, 45, 1367–1380. [Google Scholar] [CrossRef]
- Büntgen, U.; Myglan, V.S.; Ljungqvist, F.C.; McCormick, M.; Di Cosmo, N.; Sigl, M.; Kaplan, J.O. Cooling and societal change during the Late Antique Little Ice Age from 536 to around 660 AD. Nat. Geosci. 2016, 9, 231–236. [Google Scholar] [CrossRef]
- Yang, Y.; Guan, J.; Yin, J.; Shao, B.; Li, H. Urinary levels of bisphenol analogues in residents living near a manufacturing plant in south China. Chemosphere 2014, 112, 481–486. [Google Scholar] [CrossRef]
- Hadad, M.A.; González-Reyes, Á.; Roig, F.A.; Matskovsky, V.; Cherubini, P. Tree-ring-based hydroclimatic reconstruction for the northwest Argentine Patagonia since 1055 CE and its teleconnection to large-scale atmospheric circulation. Glob. Planet. Chang. 2021, 202, 103496. [Google Scholar] [CrossRef]
- Allen, K.J.; Freund, M.B.; Palmer, J.G.; Simkin, R.; Williams, L.; Brookhouse, M.; Cook, E.R.; Stewart, S.; Baker, P.J. Hydroclimate extremes in a north Australian drought reconstruction asymmetrically linked with Central Pacific Sea surface temperatures. Glob. Planet. Chang. 2020, 195, 103329. [Google Scholar] [CrossRef]
- Panthi, S.; Fan, Z.X.; van der Sleen, P.; Zuidema, P.A. Long-term physiological and growth responses of Himalayan fir to environmental change are mediated by mean climate. Glob. Chang. Biol. 2020, 26, 1778–1794. [Google Scholar] [CrossRef]
- Salick, J.; Ghimire, S.K.; Fang, Z.; Dema, S.; Konchar, K.M. Himalayan alpine vegetation, climate change and mitigation. J. Ethnobiol. 2014, 34, 276–293. [Google Scholar] [CrossRef]
- Schwab, N.; Kaczka, R.J.; Janecka, K.; Böhner, J.; Chaudhary, R.P.; Scholten, T.; Schickhoff, U. Climate change-induced shift of tree growth sensitivity at a central Himalayan treeline ecotone. Forests 2018, 9, 267. [Google Scholar] [CrossRef]
- Salzer, M.W.; Hughes, M.K.; Bunn, A.G.; Kipfmueller, K.F. Recent unprecedented tree-ring growth in bristlecone pine at the highest elevations and possible causes. Proc. Natl. Acad. Sci. USA 2009, 106, 20348–20353. [Google Scholar] [CrossRef]
- Qi, Z.; Liu, H.; Wu, X.; Hao, Q. Climate-driven speedup of alpine treeline forest growth in the Tianshan Mountains, northwestern China. Glob. Chang. Biol. 2015, 21, 816–826. [Google Scholar] [CrossRef] [PubMed]
- Liang, E.; Shao, X.; Xu, Y. Tree-ring evidence of recent abnormal warming on the southeast Tibetan Plateau. Theor. Appl. Climatol. 2009, 98, 9–18. [Google Scholar] [CrossRef]
- Bayramzadeh, V.; Zhu, H.; Lu, X.; Attarod, P.; Zhang, H.; Li, X.; Asad, F.; Liang, E. Temperature variability in northern Iran during the past 700 years. Sci. Bull. 2018, 63, 462–464. [Google Scholar] [CrossRef]
- Hartl-Meier, C.; Dittmar, C.; Zang, C.; Rothe, A. Mountain forest growth response to climate change in the Northern Limestone Alps. Trees 2014, 28, 819–829. [Google Scholar] [CrossRef]
- Zhang, X.; Liu, X.; Zhang, Q.; Zeng, X.; Xu, G.; Wu, G.; Wang, W. Species-specific tree growth and intrinsic water-use efficiency of Dahurian larch (Larix gmelinii) and Mongolian pine (Pinus sylvestris var. Mongolica) growing in a boreal permafrost region of the greater Hinggan Mountains, northeastern China. Agric. For. Meteorol. 2018, 248, 145–155. [Google Scholar] [CrossRef]
- Liang, E.; Dawadi, B.; Pederson, N.; Eckstein, D. Is the growth of birch at the upper timberline in the Himalayas limited by moisture or by temperature? Ecology 2014, 95, 2453–2465. [Google Scholar] [CrossRef]
- Liang, E.; Leuschner, C.; Dulamsuren, C.; Wagner, B.; Hauck, M. Global warming related tree growth decline and mortality on the north-eastern Tibetan plateau. Clim. Chang. 2016, 134, 163–176. [Google Scholar] [CrossRef]
- Weigel, R.; Muffler, L.; Klisz, M.; Kreyling, J.; van der Maaten-Theunissen, M.; Wilmking, M.; van der Maaten, E. Winter matters: Sensitivity to winter climate and cold events increases towards the cold distribution margin of European beech (Fagus sylvatica L.). J. Biogeogr. 2018, 45, 2779–2790. [Google Scholar] [CrossRef]
- Harvey, J.E.; Smiljanić, M.; Scharnweber, T.; Buras, A.; Cedro, A.; Cruz-García, R.; Drobyshev, I.; Janecka, K.; Jansons, Ā.; Kaczka, R.; et al. Tree growth influenced by warming winter climate and summer moisture availability in northern temperate forests. Glob. Chang. Biol. 2020, 26, 2505–2518. [Google Scholar] [CrossRef]
- Zeng, X.; Evans, M.N.; Liu, X.; Wang, W.; Xu, G.; Wu, G.; Zhang, L. Spatial patterns of precipitation-induced moisture availability and their effects on the divergence of conifer stem growth in the western and eastern parts of China’s semi-arid region. For. Ecol. Manag. 2019, 451, 117524. [Google Scholar] [CrossRef]
- Stajić, B.; Kazimirović, M.; Dukić, V.; Radaković, N. First Dendro-climatological Insight into Austrian Pine (Pinus nigra Arnold) Climate-Growth Relationship in Belgrade Area, Serbia. South-East Eur. 2020, 11, 127–134. [Google Scholar] [CrossRef]
- Manrique-Alba, À.; Beguería, S.; Molina, A.J.; González-Sanchis, M.; Tomàs-Burguera, M.; Del Campo, A.D.; Camarero, J.J. Long-term thinning effects on tree growth, drought response and water use efficiency at two Aleppo pine plantations in Spain. Sci. Total Environ. 2020, 728, 138536. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Cao, R.; Yin, J.; Tian, K.; Xiao, D.; Zhang, W.; Yin, D. Radial growth response of major conifers to climate change on Haba Snow Mountain, Southwestern China. Dendrochronologia 2020, 60, 125682. [Google Scholar] [CrossRef]
- Iqbal, J.; Ahmed, M.; Siddiqui, M.F.; Khan, A. Tree ring studies from some conifers and present condition of forest of Shangla district of Khyber Pukhtunkhwa Pakistan. Pak. J. Bot. 2020, 52, 653–662. [Google Scholar] [CrossRef]
- Bhandari, S.; Speer, J.H.; Khan, A.; Ahmed, M. Drought signal in the tree rings of three conifer species from Northern Pakistan. Dendrochronologia 2020, 63, 125742. [Google Scholar] [CrossRef]
- Thapa, U.K.; George, S.S.; Kharal, D.K.; Gaire, N.P. Tree growth across the Nepal Himalaya during the last four centuries. Prog. Phys. Geogr. 2017, 41, 478–495. [Google Scholar] [CrossRef]
- Spach. Histoire Naturelle des Vegetaux; V. 11; Labrairie Encyclopedique de Roret: Paris, France, 1841; Available online: https://www.biodiversitylibrary.org/page/31487119#page/434/mode/1up (accessed on 9 March 2019).
- Shrestha, K.B.; Hofgaard, A.; Vandvik, V. Tree-growth response to climatic variability in two climatically contrasting treeline ecotone areas, central Himalaya, Nepal. Can. J. For. Res. 2015, 45, 1643–1653. [Google Scholar] [CrossRef]
- Rayback, S.A.; Shrestha, K.B.; Hofgaard, A. Growth variable-specific moisture and temperature limitations in co-occurring alpine tree and shrub species, central Himalayas, Nepal. Dendrochronologia 2017, 44, 193–202. [Google Scholar] [CrossRef]
- Zhuang, L.; Axmacher, J.C.; Sang, W. Different radial growth responses to climate warming by two dominant tree species at their upper altitudinal limit on Changbai Mountain. J. For. Res. 2017, 28, 795–804. [Google Scholar] [CrossRef]
- Keenan, R.J.; Reams, G.A.; Achard, F.; de Freitas, J.V.; Grainger, A.; Lindquist, E. Dynamics of global forest area: Results from the FAO Global Forest Resources Assessment. For. Ecol. Manag. 2015, 352, 9–20. [Google Scholar] [CrossRef]
- Xu, H.J.; Wang, X.P.; Zhang, X.X. Decreased vegetation growth in response to summer drought in Central Asia from 2000 to 2012. Int. J. Appl. Earth Obs. Geoinf. 2016, 52, 390–402. [Google Scholar] [CrossRef]
- Sun, J.; Liu, Y. Age-independent climate-growth response of Chinese pine (Pinus tabulaeformis Carrière) in North China. Trees 2015, 29, 397–406. [Google Scholar] [CrossRef]
- Khan, N.; Ahmed, M.; Wahab, M. Dendroclimatic potential of Picea smithiana (Wall) Boiss, from Afghanistan. Pak. J. Bot. 2008, 40, 1063–1070. [Google Scholar]
- Laxton, S.C.; Smith, D.J. Dendrochronological reconstruction of snow avalanche activity in the Lahul Himalaya, Northern India. Nat. Hazards 2009, 49, 459–467. [Google Scholar] [CrossRef]
- Ahmed, M.; Wahab, M.; Khan, N.; Siddiqui, M.F.; Khan, M.U.; Hussain, S.T. Age and growth rates of some gymnosperms of Pakistan: A dendrochronological approach. Pak. J. Bot 2009, 41, 849–860. [Google Scholar]
- Ahmed, M.; Wahab, M.; Khan, N.A.S.R.U.L.L.A.H.; Zafar, M.U.; Palmer, J. Tree-ring chronologies from upper indus basin of karakorum range, Pakistan. Pak. J. Bot. 2010, 42, 295–308. [Google Scholar]
- Zafar, M.U.; Ahmed, M.; Farooq, M.A.; Akbar, M.; Hussain, A. Standardized tree ring chronologies of Picea smithiana from two new sites of Northern area Pakistan. World Appl. Sci. J. 2010, 11, 1531–1536. [Google Scholar]
- Thapa, U.K.; Shah, S.K.; Gaire, N.P.; Bhuju, D.R. Spring temperatures in the far-western Nepal Himalaya since AD 1640 reconstructed from Picea smithiana tree-ring widths. Clim. Dyn. 2015, 45, 2069–2081. [Google Scholar] [CrossRef]
- Seim, A.; Omurova, G.; Azisov, E.; Musuraliev, K.; Aliev, K.; Tulyaganov, T.; Nikolyai, L.; Botman, E.; Helle, G.; Dorado Liñan, I.; et al. Climate Change Increases Drought Stress of Juniper Trees in the Mountains of Central Asia. PLoS ONE 2016, 11, e0153888. [Google Scholar] [CrossRef]
- Solomina, A.; Maximova, A.; Cook, A. Picea Schrenkian ring width and density at the upper and lower tree limits in the Tien Shan Mountains (Kirgizstan Republic) as a source of paleoclimatic information. Geogr. Environ. Sustain. 2014, 7, 66–79. [Google Scholar] [CrossRef]
- Zhang, T.; Yuan, Y.; He, Q.; Wei, W.; Diushen, M.; Shang, H.; Zhang, R. Development of tree-ring width chronologies and tree-growth response to climate in the mountains surrounding the Issyk-Kul Lake, Central Asia. Dendrochronologia 2014, 32, 230–236. [Google Scholar] [CrossRef]
- Lyu, L.; Deng, X.; Zhang, Q.B. Elevation pattern in growth coherency on the southeastern Tibetan Plateau. PLoS ONE 2016, 11, e0163201. [Google Scholar] [CrossRef]
- Rauning, A. Combined view of various tree ring parameters from different habitats in Tibet for the reconstruction of seasonal aspects of Asian Monsoon variability. Palaeobotanist 2001, 50, 1–12. [Google Scholar]
- Fan, Z.; Bräuning, A.; Cao, K. Tree-ring based drought reconstruction in the central Hengduan Mountains region (China) since AD 1655. Int. J. Climatol. 2008, 28, 1879–1887. [Google Scholar] [CrossRef]
- Korner, C. Alpine Treelines; Springer: Basel, Switzerland, 2012. [Google Scholar]
- Fan, Z.X.; Bräuning, A.; Cao, K.F.; Zhu, S.D. Growth–climate responses of high elevation conifers in the central Hengduan Mountains, southwestern China. For. Ecol. Manag. 2009, 258, 306–313. [Google Scholar] [CrossRef]
- Latreille, A.; Davi, H.; Huard, F.; Pichot, C. Variability of the climate-radial growth relationship among Abies alba trees and populations along altitudinal gradients. For. Ecol. Manag. 2017, 396, 150–159. [Google Scholar] [CrossRef]
- Wang, W.; Jia, M.; Wang, G.; Zhu, W.; Mcdowell, N.G. Rapid warming forces contrasting growth trends of subalpine fir (Abies fabri) at higher- and lower-elevations in the eastern Tibetan Plateau. For. Ecol. Manag. 2017, 402, 135–144. [Google Scholar] [CrossRef]
- Kharal, D.K.; Thapa, U.K.; St. George, S.; Meilby, H.; Rayamajhi, S.; Bhuju, D.R. Tree-climate relations along an elevational transect in Manang Valley, central Nepal. Dendrochronologia 2017, 41, 57–64. [Google Scholar] [CrossRef]
- Panthi, S.; Bräuning, A.; Zhou, Z.K.; Fan, Z.X. Tree rings reveal recent intensified spring drought in the central Himalaya, Nepal. Glob. Planet. Chang. 2017, 157, 26–34. [Google Scholar] [CrossRef]
- Sohar, K.; Altman, J.; Lehečková, E.; Doležal, J. Growth–climate relationships of Himalayan conifers along elevational and latitudinal gradients. Int. J. Climatol. 2017, 37, 2593–2605. [Google Scholar] [CrossRef]
- Li, J.; Shi, J.; Zhang, D.D.; Yang, B.; Fang, K.; Yue, P.H. Moisture increase in response to high-altitude warming evidenced by tree-rings on the southeastern Tibetan Plateau. Clim. Dyn. 2017, 48, 649–660. [Google Scholar] [CrossRef]
- Lloyd, A.H.; Bunn, A.G.; Berner, L. A latitudinal gradient in tree growth response to climate warming in the Siberian taiga. Glob. Chang. Biol. 2011, 17, 1935–1945. [Google Scholar] [CrossRef]
- Ahmed, M.; Palmer, J.; Khan, N.; Wahab, M.; Fenwick, P.; Esper, J.; Cook, E. The dendroclimatic potential of conifers from northern Pakistan. Dendrochronologia 2011, 29, 77–88. [Google Scholar] [CrossRef]
- Ahmed, M.; Khan, N.; Wahab, M.; Zafar, U.; Palmer, J. Climate/growth correlations of tree species in the Indus basin of the Karakorum range, north Pakistan. IAWA J. 2012, 33, 51–61. [Google Scholar] [CrossRef]
- Ahmed, M.; Zafar, M.U.; Hussain, A.; Akbar, M.; Wahab, M.; Khan, N. Dendroclimatic and dendrohydrological response of two tree species from Gilgit valleys. Pak. J. Bot. 2013, 45, 987–992. [Google Scholar]
- Ahmed, M.; Zafar, M.U. The Status of Tree-Ring Analysis in Pakistan. FUUAST J. Biol. 2014, 4, 13–19. [Google Scholar]
- Zafar, M.U.; Ahmed, M.; Rao, M.P.; Buckley, B.M.; Khan, N.; Wahab, M.; Palmer, J. Karakorum temperature out of phase with hemispheric trends for the past five centuries. Clim. Dyn. 2015, 46, 1943–1952. [Google Scholar] [CrossRef]
- Iqbal, J.; Ahmed, M.; Siddiqui, M.F.; Khan, A.; Wahab, M. Age and radial growth analysis of conifer tree species from Shangla, Pakistan. Pak. J. Bot. 2017, 49, 69–72. [Google Scholar]
- Khan, A.; Ahmed, M.; Siddiqui, M.F.; Iqbal, J.; Gaire, N.P. Dendrochronological potential of Abies pindrow Royle from Indus Kohistan, Khyber Pakhtunkhwa (KPK) Pakistan. Pak. J. Bot. 2018, 50, 365–369. [Google Scholar]
- Ahmad, N.; Ashraf, M.I.; Malik, S.U.; Qadir, I.; Malik, N.A.; Khan, K. Impact of Climatic and Topographic Factors on Distribution of Sub-tropical and Moist Temperate Forests in Pakistan. Géomorphol. Relief Process. Environ. 2020, 26, 157–172. [Google Scholar] [CrossRef]
- Ali, A.; Ashraf, M.I.; Gulzar, S.; Akmal, M. Estimation of forest carbon stocks in temperate and subtropical mountain systems of Pakistan: Implications for REDD+ and climate change mitigation. Environ. Monit. Assess. 2020, 192, 198. [Google Scholar] [CrossRef] [PubMed]
- Holmes, R.L. Computer-assisted quality control in tree-ring dating and measurement. Tree-Ring Bull. 1983, 43, 69–78. [Google Scholar]
- Duan, J.; Wu, P.; Ma, Z.; Duan, Y. Unprecedented recent late-summer warm extremes recorded in tree-ring density on the Tibetan Plateau. Environ. Res. Lett. 2020, 15, 024006. [Google Scholar] [CrossRef]
- Dai, A.; Trenberth, K.E.; Qian, T. A global dataset of Palmer Drought Severity Index for 1870–2002: Relationship with soil moisture and effects of surface warming. J. Hydrometeorol. 2004, 5, 1117–1130. [Google Scholar] [CrossRef]
- Rocha, E.; Gunnarson, B.E.; Holzkämper, S. Reconstructing summer precipitation with mxd data from Pinus sylvestris growing in the Stockholm archipelago. Atmosphere 2020, 11, 790. [Google Scholar] [CrossRef]
- Khaleghi, M.R. Application of dendroclimatology in evaluation of climatic changes. J. For. Sci. 2018, 64, 139–147. [Google Scholar] [CrossRef]
- Shah, H.; Jehan, N.; Rehman, S.S.; Bukhari, S.S.B. Comparative Study of Climate Change and its Impact on Ring-Widths of Spruce (Picea smithiana) at Kalam and Kaghan Forest Divisions, Khyber Pakhtunkhwa, Pakistan. Sarhad J. Agric. 2019, 35, 788–797. [Google Scholar] [CrossRef]
- Thapa, U.K.; Shah, S.K.; Gaire, N.P. Influence of climate on radial growth of Abies pindrow in western Nepal Himalaya. Banko Janakari 2013, 23, 14–19. [Google Scholar] [CrossRef]
- Palmer, J.; Ahmed, M.; Khan, Z. Application of tree-ring research in Pakistan. FUUAST J. Biol. 2011, 1, 19–25. [Google Scholar]
- Asad, F.; Zhu, H.; Zhang, H.; Liang, E.; Muhammad, S.; Farhan, S.B.; Hussain, I.; Wazir, M.A.; Ahmed, M.; Esper, J. Are Karakoram temperatures out of phase compared to hemispheric trends? Clim. Dyn. 2016, 48, 3381–3390. [Google Scholar] [CrossRef]
- Borgaonkar, H.P.; Sikder, A.B.; Ram, S. High altitude forest sensitivity to the recent warming: A tree-ring analysis of conifers from Western Himalaya, India. Quat. Int. 2011, 236, 158–166. [Google Scholar] [CrossRef]
- Shah, S.K.; Pandey, U.; Mehrotra, N. Precipitation reconstruction for the Lidder Valley, Kashmir Himalaya using tree-rings of Cedrus deodara. Int. J. Climatol. 2018, 38, 758–773. [Google Scholar] [CrossRef]
- Borgaonkar, H.P.; Pant, G.B.; Kumar, K.R. Tree-ring chronologies from western Himalaya and their dendroclimatic potential. IAWA J. 1999, 20, 295–309. [Google Scholar] [CrossRef]
- Ram, S.; Borgaonkar, H.P. Growth response of conifer trees from high-altitude region of western Himalaya. Curr. Sci. 2013, 105, 225–231. [Google Scholar]
- Gaire, N.P.; Fan, Z.X.; Bräuning, A.; Panthi, S.; Rana, P.; Shrestha, A.; Bhuju, D.R. Abies spectabilis shows stable growth relations to temperature, but changing response to moisture conditions along an elevation gradient in the central Himalaya. Dendrochronologia 2020, 60, 125675. [Google Scholar] [CrossRef]
- Wahab, M. Population Dynamics and Dendrochronological Potential of Pine Tree Species from District Dir. Ph.D. Thesis, Federal Urdu University of Arts, Science and Technology, Karachi, Pakistan, 2011. [Google Scholar]
- Häusser, M.; Szymczak, S.; Garel, E.; Santoni, S.; Huneau, F.; Bräuning, A. Growth variability of two native pine species on Corsica as a function of elevation. Dendrochronologia 2019, 54, 49–55. [Google Scholar] [CrossRef]
- van der Maaten-Theunissen, M.; Kahle, H.P.; van der Maaten, E. Drought sensitivity of Norway spruce is higher than that of silver fir along an altitudinal gradient in southwestern Germany. Ann. For. Sci. 2013, 70, 185–193. [Google Scholar] [CrossRef]
- Sigdel, S.R.; Wang, Y.; Camarero, J.J.; Zhu, H.; Liang, E.; Peñuelas, J. Moisture-mediated responsiveness of treeline shifts to global warming in the Himalayas. Glob. Chang. Biol. 2018, 24, 5549–5559. [Google Scholar] [CrossRef]
- Liang, E.; Camarero, J.J. Threshold-dependent and non-linear associations between temperature and tree growth at and below the alpine treeline. Trees 2018, 32, 661–662. [Google Scholar] [CrossRef]
- Gaire, N.P.; Dhakal, Y.R.; Shah, S.K.; Fan, Z.X.; Bräuning, A.; Thapa, U.K.; Bhuju, D.R. Drought (scPDSI) reconstruction of trans-Himalayan region of central Himalaya using Pinus wallichiana tree-rings. Palaeogeogr. Palaeoclimatol. Palaeoecol. 2019, 514, 251–264. [Google Scholar] [CrossRef]
- Yadav, R.R.; Mishra, K.G.; Kotila, B.; Upreti, N. Premonsoon precipitation variability in Kumaon Himalaya, India over a perspective of ~300 years. Quat. Int. 2014, 325, 213–219. [Google Scholar] [CrossRef]
- Singh, S.P.; Singh, R.D.; Gumber, S.U.R.A.B.H.I.; Bhatt, S.P.A.R.S.H. Two principal precipitation regimes in Himalayas and their influence on tree distribution. Trop. Ecol. 2017, 58, 679–691. [Google Scholar]
- Ram, S. Tree growth–climate relationships of conifer trees and reconstruction of summer season Palmer Drought Severity Index (PDSI) at Pahalgam in Srinagar, India. Quat. Int. 2012, 254, 152–158. [Google Scholar] [CrossRef]
- Roibu, C.C.; Sfeclă, V.; Mursa, A.; Ionita, M.; Nagavciuc, V.; Chiriloaei, F.; Popa, I. The Climatic Response of Tree Ring Width Components of Ash (Fraxinus excelsior L.) and Common Oak (Quercus robur L.) from Eastern Europe. Forests 2020, 11, 600. [Google Scholar] [CrossRef]
- Rai, S.; Dawadi, B.; Wang, Y.; Lu, X.; Ru, H.; Sigdel, S.R. Growth response of Abies spectabilis to climate along an elevation gradient of the Manang valley in the central Himalayas. J. For. Res. 2019, 31, 2245–2254. [Google Scholar] [CrossRef]
- Lyu, L.; Zhang, Q.B.; Pellatt, M.G.; Büntgen, U.; Li, M.H.; Cherubini, P. Drought limitation on tree growth at the Northern Hemisphere’s highest tree line. Dendrochronologia 2019, 53, 40–44. [Google Scholar] [CrossRef]
- Rossi, S.; Deslauriers, A.; Anfodillo, T.; Carraro, V. Evidence of threshold temperatures for xylogenesis in conifers at high altitudes. Oecologia 2007, 152, 1–12. [Google Scholar] [CrossRef]
- Henttonen, H.M.; Mäkinen, H.; Nöjd, P. Seasonal dynamics of the radial increment of Scots pine and Norway spruce in the southern and middle boreal zones in Finland. Can. J. For. Res. 2009, 39, 606–618. [Google Scholar] [CrossRef]
- Lv, L.X.; Zhang, Q.B. Asynchronous recruitment history of Abies spectabilis along an altitudinal gradient in the Mt. Everest region. J. Plant Ecol. 2012, 5, 147–156. [Google Scholar] [CrossRef]
- Jyske, T.; Mäkinen, H.; Kalliokoski, T.; Nöjd, P. Intra-annual tracheid production of Norway spruce and Scots pine across a latitudinal gradient in Finland. Agric. For. Meteorol. 2014, 194, 241–254. [Google Scholar] [CrossRef]
- Mäkinen, H.; Nöjd, P.; Kahle, H.P.; Neumann, U.; Tveite, B.; Mielikäinen, K.; Spiecker, H. Radial growth variation of Norway spruce (Picea abies (L.) Karst.) across latitudinal and altitudinal gradients in central and northern Europe. For. Ecol. Manag. 2002, 171, 243–259. [Google Scholar] [CrossRef]
- Bosela, M.; Tumajer, J.; Cienciala, E.; Dobor, L.; Kulla, L.; Marčiš, P.; Šebeň, V. Climate warming induced synchronous growth decline in Norway spruce populations across biogeographical gradients since 2000. Sci. Total Environ. 2020, 752, 141794. [Google Scholar] [CrossRef] [PubMed]
- Čater, M.; Levanič, T. Beech and silver fir’s response along the Balkan’s latitudinal gradient. Sci. Rep. 2019, 9, 16269. [Google Scholar] [CrossRef] [PubMed]
- Gazol, A.; Ibáñez, R. Plant species composition in a temperate forest: Multi-scale patterns and determinants. Acta Oecol. 2010, 36, 634–644. [Google Scholar] [CrossRef]
- Diaci, J.; Rozenbergar, D.; Anic, I.; Mikac, S.; Saniga, M.; Kucbel, S.; Ballian, D. Structural dynamics and synchronous silver fir decline in mixed old-growth mountain forests in Eastern and Southeastern Europe. Forestry 2011, 84, 479–491. [Google Scholar] [CrossRef]
- Ficko, A.; Poljanec, A.; Boncina, A. Do changes in spatial distribution, structure and abundance of silver fir (Abies alba Mill.) indicate its decline? For. Ecol. Manag. 2011, 261, 844–854. [Google Scholar] [CrossRef]
- Castagneri, D.; Nola, P.; Motta, R.; Carrer, M. Summer climate variability over the last 250 years differently affected tree species radial growth in a mesic Fagus–Abies–Picea old-growth forest. For. Ecol. Manag. 2014, 320, 21–29. [Google Scholar] [CrossRef]
- Liang, P.; Wang, X.; Sun, H.; Fan, Y.; Wu, Y.; Lin, X.; Chang, J. Forest type and height are important in shaping the altitudinal change of radial growth response to climate change. Sci. Rep. 2019, 9, 1336. [Google Scholar] [CrossRef]
- Yasmeen, S.; Wang, X.; Zhao, H.; Zhu, L.; Yuan, D.; Li, Z.; Han, S. Contrasting climate-growth relationship between Larix gmelinii and Pinus sylvestris var. mongolica along a latitudinal gradient in Daxing’an Mountains, China. Dendrochronologia 2019, 58, 125645. [Google Scholar] [CrossRef]












{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sampling Site | Latitude | Longitude | Forest Type | Elevation Range (Meter) |
|---|---|---|---|---|
| Shogran (SHG) | 34°38′30″ N | 73°29′10″ E | Moist Temperate Forests | 1830 to 2950 |
| Upper Dir (UDS) | 35°17′30″ N | 72°02′30″ E | Dry Temperate Forests | 2950 to 3600 |
| Bahrain Swat (BSG) | 35°12′30″ N | 72°34′30″ E | Moist Temperate Forests | 1830 to 2950 |
| Astore Gilgit (NPKA) | 35°34′10″ N | 74°59′20″ E | Dry Temperate Forests | 2950 to 3600 |
| Sharan Kaghan (SHA) | 34°41′30″ N | 73°25′50″ E | Moist Temperate Forests | 2950 to 3600 |
| SHGSP | UDSSP | BSGSP | SHGF | UDSF | BSGF | NPKAF | SHAF | |
|---|---|---|---|---|---|---|---|---|
| SHGSP | 1 | 0.281 ** | 0.200 ** | 0.447 ** | 0.286 ** | −0.142 | 0.262 ** | 0.458 ** |
| UDSSP | 0.281 ** | 1 | 0.164 * | 0.164 * | 0.579 ** | 0.050 | 0.162 * | 0.400 ** |
| BSGSP | 0.200 ** | 0.164 * | 1 | 0.288 ** | 0.179 * | 0.114 | −0.068 | 0.021 |
| SHGF | 0.447 ** | 0.164 * | 0.288 ** | 1 | 0.337 ** | 0.002 | 0.169 * | 0.516 ** |
| UDSF | 0.286 ** | 0.579 ** | 0.179 * | 0.337 ** | 1 | 0.103 | 0.101 | 0.369** |
| BSGF | −0.142 | 0.050 | 0.114 | 0.002 | 0.103 | 1 | −0.117 | −0.167 * |
| NPKAF | 0.262 ** | 0.162 * | −0.068 | 0.169 * | 0.101 | −0.117 | 1 | 0.295 ** |
| SHAF | 0.458 ** | 0.400 ** | 0.021 | 0.516 ** | 0.369 ** | −0.167 * | 0.295 ** | 1 |
| Picea smithiana | Abies pindrow | |||||||
|---|---|---|---|---|---|---|---|---|
| SHG | UDS | BSG | SHG | UDS | BSG | NPKA | SHA | |
| TS | 1850–2017 | 1804–2017 | 1842–2018 | 1806–2017 | 1835–2017 | 1851–2018 | 1505–2005 | 1696–2018 |
| TN | 27(34) | 30(34) | 40(45) | 23(30) | 25(30) | 28(34) | 20(25) | 26(30) |
| MRW (mm) | 0.9746 | 0.9798 | 0.9931 | 0.9763 | 0.958 | 0.9755 | 0.987 | 0.9725 |
| MS | 0.1182 | 0.2132 | 0.1122 | 0.143 | 0.1645 | 0.1608 | 0.1449 | 0.1458 |
| SD | 0.1434 | 0.2993 | 0.1587 | 0.1848 | 0.1952 | 0.1883 | 0.1449 | 0.2981 |
| AC1 | 0.5165 | 0.4938 | 0.6 | 0.5696 | 0.4501 | 0.4929 | 0.4528 | 0.7962 |
| ISC | 0.449 | 0.559 | 0.494 | 0.547 | 0.495 | 0.524 | 0.608 | 0.481 |
| Common Interval Analysis for Period 1891 to 2018 | ||||||||
| Var | 24.85% | 34.66% | 19.28% | 27.88% | 29.21% | 25.45% | 33.64% | 36.01% |
| SNR | 5.405 | 8.416 | 5.101 | 6.285 | 6.134 | 5.701 | 7.238 | 8.732 |
| EPS | 0.844 | 0.894 | 0.836 | 0.863 | 0.86 | 0.851 | 0.879 | 0.897 |
| MCC | 0.197 | 0.277 | 0.116 | 0.222 | 0.235 | 0.186 | 0.266 | 0.294 |
| SHG | UDS | BSG | NPKA | SHA | |||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| Picea smithiana | 1950–1980 | 1981–2017 | 1950–2017 | 1981–2017 | 1950–1980 | 1981–2018 | 1950–1980 | 1981–2005 | 1950–1980 | 1981–2017 | |
| pWinter | 0.026 | −0.019 | 0.393 * | 0.316 | 0.051 | 0.362 * | |||||
| Spring | −0.007 | −0.013 | 0.269 | 0.388 * | −0.088 | 0.416 * | |||||
| Summer | −0.119 | 0.043 | 0.208 | 0.454 ** | −0.146 | 0.429 ** | |||||
| Autumn | −0.167 | −0.037 | 0.253 | 0.352 * | −0.232 | 0.306 | |||||
| Abies pindrow | pWinter | −0.128 | 0.325 * | 0.247 | 0.205 | 0.265 | 0.227 | −0.052 | −0.419 * | −0.466 ** | 0.012 |
| Spring | −0.124 | 0.310 | 0.153 | 0.017 | 0.131 | 0.239 | −0.032 | −0.488 * | 0.079 | 0.079 | |
| Summer | −0.224 | 0.372 * | 0.201 | 0.393 * | 0.186 | 0.391 * | −0.265 | −0.379 | −0.413 * | 0.143 | |
| Autumn | −0.300 | 0.332 * | 0.159 | 0.340 * | 0.102 | 0.449 ** | −0.273 | −0.444 * | −0.454 * | 0.042 | |
| SHG | UDS | BSG | NPKA | SHA | |||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| Picea smithiana | 1950–1980 | 1981–2017 | 1950–2017 | 1981–2017 | 1950–1980 | 1981–2018 | 1950–1980 | 1981–2005 | 1950–1980 | 1981–2017 | |
| pWinter | −0.099 | 0.246 | 0.401 * | −0.077 | 0.142 | 0.000 | |||||
| Spring | −0.140 | 0.015 | −0.087 | 0.437 ** | −0.104 | 0.406 * | |||||
| Summer | −0.075 | 0.040 | −0.016 | 0.200 | −0.379 * | 0.120 | |||||
| Autumn | −0.103 | 0.114 | 0.274 | −0.153 | −0.056 | −0.080 | |||||
| Abies pindrow | pWinter | −0.004 | 0.044 | 0.130 | −0.065 | 0.095 | −0.023 | 0.058 | −0.446 * | −0.149 | 0.168 |
| Spring | 0.052 | 0.282 | 0.123 | 0.415 * | 0.065 | 0.365 * | 0.323 | −0.275 | 0.168 | 0.186 | |
| Summer | −0.380 * | 0.145 | −0.161 | 0.302 | −0.312 | 0.262 | −0.321 | 0.208 | −0.049 | −0.084 | |
| Autumn | 0.083 | −0.032 | 0.136 | −0.045 | 0.350 | 0.113 | −0.134 | −0.173 | −0.131 | −0.019 | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ullah, H.; Wang, X.; Hussain, Q.; Khan, A.; Ahmad, N.; Ali, N.; Riaz, M.W.; Hussain, I. The Climate-Growth Relationship between Picea smithiana (Wall.) Boiss. and Abies pindrow (Royle ex D.Don) Royle along the Latitudinal Gradient in Northern Pakistan. Forests 2022, 13, 1315. https://0-doi-org.brum.beds.ac.uk/10.3390/f13081315
Ullah H, Wang X, Hussain Q, Khan A, Ahmad N, Ali N, Riaz MW, Hussain I. The Climate-Growth Relationship between Picea smithiana (Wall.) Boiss. and Abies pindrow (Royle ex D.Don) Royle along the Latitudinal Gradient in Northern Pakistan. Forests. 2022; 13(8):1315. https://0-doi-org.brum.beds.ac.uk/10.3390/f13081315
Chicago/Turabian StyleUllah, Habib, Xiaochun Wang, Quaid Hussain, Abdullah Khan, Naveed Ahmad, Nizar Ali, Muhammad Waheed Riaz, and Izhar Hussain. 2022. "The Climate-Growth Relationship between Picea smithiana (Wall.) Boiss. and Abies pindrow (Royle ex D.Don) Royle along the Latitudinal Gradient in Northern Pakistan" Forests 13, no. 8: 1315. https://0-doi-org.brum.beds.ac.uk/10.3390/f13081315