
Organic Mulching Increases Microbial Activity in Urban Forest Soil
by
 and
and
Xiaodan Sun
1,2,*, 
Yuqian Ye
2,
Jiahui Liao
2,
Andrey V. Soromotin
3,
Pavel V. Smirnov
3 and
Yakov Kuzyakov
4,5,6 1
Marine Ecology Research Center, First Institute of Oceanography, Ministry of Natural Resources, Qingdao 266061, China
2
College of Biology and the Environment, Nanjing Forestry University, Nanjing 210037, China
3
Research Institute of Ecology and Natural Resources Management, University of Tyumen, 625003 Tyumen, Russia
4
Department of Soil Science of Temperate Ecosystems, Department of Agricultural Soil Science, University of Goettingen, 37073 Göttingen, Germany
5
Agro-Technological Institute, Peoples Friendship University of Russia (RUDN University), 117198 Moscow, Russia
6
Institute of Environmental Sciences, Kazan Federal University, 420049 Kazan, Russia
*
Author to whom correspondence should be addressed.
Forests 2022, 13(9), 1352; https://0-doi-org.brum.beds.ac.uk/10.3390/f13091352
Submission received: 24 July 2022
/
Revised: 19 August 2022
/
Accepted: 22 August 2022
/
Published: 25 August 2022
(This article belongs to the Special Issue Soil Organic Matter and Nutrient Cycling in Forests)
Abstract
:Interactions between C accumulation, resulting from plant residue mulching, and the accelerated decomposition of litter and soil organic matter due to higher enzyme activities remain poorly understood, particularly in urban forests. Here, the activities of four enzymes associated with carbon (C) and nitrogen (N) cycling after organic mulching were investigated in the rhizosphere and bulk soil of a 15-year-old Ligustrum lucidum urban forest. The enzymatic activities in the rhizosphere were closely correlated to those in bulk soil, however, they were more strongly affected by organic mulching and exhibited faster C and nutrient cycling. Moreover, the activity of urease in the rhizosphere and peroxidase in the bulk soil in autumn, as well as invertase in the rhizosphere in spring, decreased. Microbial C and N were most responsible for the observed increase in invertase activity in spring, during which microorganisms exhibited rapid growth and the highest activities, whereas soil organic C regulated urease activity in winter. Additionally, the activity of dehydrogenase increased by up to 75%; the degree of increase was typically dependent on the mulching amount. Generally, invertase and dehydrogenase activity increased with the abundance of dissolved organic matter and microbial biomass, whereas peroxidase exhibited the opposite trend. Taken together, these findings indicate that organic mulching efficiently increases enzymatic activity, particularly within the rhizosphere, thus accelerating soil C and N cycling. Hence, mulching may represent an effective measure for urban forest management and soil conservation.
1. Introduction
Large population influxes to urban areas have caused the continuous expansion of cities [1,2]. Consequently, the physico-chemical and biological properties, as well as nutrient cycling, in urban soils have been considerably altered, causing soil degradation [3,4]. Accordingly, the development of conservation and restoration strategies is of great environmental importance [5,6]. Organic mulching, that is, the application of natural origin materials capable of natural decomposition, improves the soil environment and quality. For example, application of plant residues to the soil surface can improve soil water content and increase nutrient levels, thereby promoting plant growth [7]. As such, mulching has an important role in urban forest management and soil conservation.
Enzyme activity reflects the intensity and direction of substrate transformation and element cycling [8] and responds more rapidly to natural and anthropogenic factors than other soil variables [9,10]. Peroxidase, invertase, dehydrogenase, and urease are the main enzymes associated with carbon (C) and nitrogen (N) cycling. However, these enzymes are sensitive to environmental changes [11] and may, in turn, affect the coupling of C and N [12,13]. The mechanisms underlying changes in enzymatic activity following disturbances are complex due to the myriad interactions among soil components.
One important factor that impacts enzyme activity in ecosystems is nutrient availability [14]. For instance, organic mulch addition alters soil microbial communities and enzymatic activity by acting as a source of nutrients and C. Furthermore, mulch affects nutrient uptake by plants, as well as the release of root exudates, consequently modifying the soil environment [15]. Meanwhile, analysis of the effect elicited by organic mulching on the enzymes within the rhizosphere is necessary to fully elucidate the mechanisms associated with plant–soil interactions [15,16]. The response of enzyme activity to mulching is expected to differ across ecosystems [17]. Currently, most studies on mulching have focused on agricultural lands [18,19], whereas a dearth of data is available for forest or urban ecosystems.
In this study, we employed a common evergreen tree species in urban green spaces, a 15-year-old Ligustrum lucidum plantation, to investigate the effects of organic mulching on C and N cycle-related enzymatic activities in the rhizosphere and bulk soil. The primary goal of this study was to address the following questions: (i) How are enzymatic activities impacted by organic mulching? (ii) Are these responses the same within the rhizosphere and bulk soil? (iii) What are the main factors that control the enzymatic activity? We hypothesized that organic mulching increases enzymatic activity due to C and N input, which accelerates organic matter decomposition and nutrient mineralization. Furthermore, these effects were expected increase with the mulching amount. Enzymatic activity after mulching was found to be higher in the rhizosphere than the bulk soil due to activation of microorganisms by root exudates. Thus, microbial C (MBC) and microbial N (MBN) significantly regulate enzymatic activities and mulching may prove effective as an urban forest management and soil conservation measure.
2. Materials and Methods
2.1. Site Description and Experimental Design
This study was conducted in a 15-year-old Ligustrum lucidum W. T. Aiton (broad-leaf privet; family, Oleaceae) plantation in Xiaolingwei, Xuanwu District, Nanjing City, China (32°02′37″–32°02′39″ N, 118°49′41″–118°49′43″ E). According to the historical records, the site was previously covered with 50–60 cm of soil after the demolition of buildings. The tree spacing of this plantation was >2 m, and the canopy density was ~85%. The average diameter at chest height was 10.9 cm, and the average tree height was 7.5 m. The soil’s basic physico-chemical properties are shown in Table 1.
Thirty-two plots were established in total within the study area by the end of November 2017; four adjacent trees were randomly selected as one plot. In each plot, we applied three treatments (35, 70, or 140 kg of mulch per tree) along with a control without mulch. Mulch was carefully mounded around each tree as uniformly as possible to a height of 0, 5, 10, or 20 cm above the ground (hereafter referred to as Control, Mulch-5, Mulch-10, and Mulch-20, respectively). Organic mulch was positioned around the trunk to a radius of 80 cm to ensure all the experimental units were handled in the same manner (i.e., 175, 350, 700 Mg ha−1), allowing for a buffer (>0.5 m) between trees. The organic mulch comprised urban garden litter after a semi-decomposition process (Shanghai Moqi Garden Co., Ltd., Shanghai, China); its basic physico-chemical properties are shown in Table 2.
2.2. Field Sampling
Soil samples were collected after 6, 9, 12, and 15 months of mulching (i.e., May, August, and November 2018 and February 2019, respectively, in spring, summer, autumn, and winter). At each monitoring time point, five experimental plots (i.e., n = 5) were randomly selected for each treatment, and each experimental plot was used only once during the entire experiment. Soil profiles were dug 50 cm away from the tree trunk, and each profile was divided into two depths (0–20 and 20–40 cm below the mulch layer). Soil was removed from each depth; rhizosphere soil was collected by gently shaking off the soil adhering to the roots, while the remaining soil was considered bulk soil. Soil samples were placed in self-sealing bags, transported to the laboratory immediately for analysis, and stored in a refrigerator at −4 °C before measurements.
2.3. Measurement Methods
After air-drying, the soil samples were sifted using 2-mm mesh sieves, and the enzymatic activities in rhizosphere and bulk soil were examined according to the methods described by Tan et al. [20]. Briefly, invertase (EC3.2.1.26) activity was measured based on the amount of glucose reduction under the catalysis of invertase for 24 h at 37 °C. Urease (EC3.5.1.5) activity was assayed using the indophenol blue colorimetric method, using 10% urea solution as the substrate; the soil mixture was incubated at 37 °C for 24 h. Dehydrogenase (EC1.1.1.1) activity was determined based on the reduction of triphenyl-tetrazolium to triphenyl formazone. Peroxidase (EC1.11.1.7) activity was measured using the pyrogallol colorimetric method (incubation 30 °C for 2 h).
Soil physical and chemical properties, including temperature, pH, water content, soil organic C (SOC), total N (TN), dissolved C, dissolved N, ammonium, nitrate, total K, total K, available P, available K, microbial C (MBC), and microbial N (MBN), were examined according to the methods described in our previous study [21].
2.4. Statistical Analysis
All statistical analyses were conducted using R Studio Version 1.1.463-© 2022–2018 (R Studio, Inc., Boston, MA, USA) [22]. Linear mixed-effects models were run using the R package “lme4”. A redundancy analysis (RDA) was performed using the R package “vegan”, and regression analyses were performed for the indicators exhibiting strong correlations according to the RDA results. All figures were prepared using the “ggplot2” package in R. A p < 0.05 was considered statistically significant.
3. Results
3.1. Enzyme Activities in the Rhizosphere and Bulk Soil after Organic Mulching
Compared to organic mulching, season and soil depth had a greater effect on the activity of all four enzymes (Table S1). The activities of dehydrogenase and invertase were the highest in the spring, while peroxidase activity was highest in the autumn, and urease activity was highest in the winter (Figure 1 and Figure S1, Table S2). Moreover, invertase and peroxidase exhibited lower activities in the rhizosphere than in the bulk soil, whereas urease and dehydrogenase exhibited the opposite trend.
Generally, organic mulching had a greater effect on the enzymatic activities in the rhizosphere than those in the bulk soil. That is, in spring, the invertase activity in the topsoil (0–20 cm) rhizosphere under Mulch-10 was 14% lower than that in the control, whereas it was 14% higher in the subsoil (20–40 cm) rhizosphere under Mulch-5 (Figure 1). Moreover, peroxidase activity in the rhizosphere under Mulch-5 increased in spring and decreased in the bulk soil in autumn. Meanwhile, in autumn, dehydrogenase activity increased by up to 75%, whereas rhizosphere urease activity decreased by up to 35%. The effects of organic mulching on enzymatic activities generally increased with the mulching amount. However, organic mulching decreased the differences in enzymatic activity between the rhizosphere and bulk soil (Figure 2).
3.2. C and N Content in the Rhizosphere and Bulk Soil after Organic Mulching
Mulching increased the dissolved C, MBN, and dissolved N in the bulk soil, as well as MBC in rhizosphere soil, while decreasing nitrate content in the bulk soil (Figure 3 and Figure S2, Table S3). Mulch-5 exhibited greater effects on MBN and bulk soil nitrate, whereas Mulch-10 did not affect dissolved C, dissolved N, ammonium, or MBC in the bulk soil. Dissolved C content increased by 1–13% after organic mulching, while the rhizosphere MBN increased under Mulch-5 by 22 and 27% in the topsoil and subsoil, respectively. In general, the SOC, TN, dissolved N, nitrate, and MBN contents in the rhizosphere soil were higher than those in the bulk soil, whereas that of ammonium and MBC exhibited the opposite trend (Figure 4 and Figure S2).
3.3. Relationship between Enzyme Activity and Soil Properties
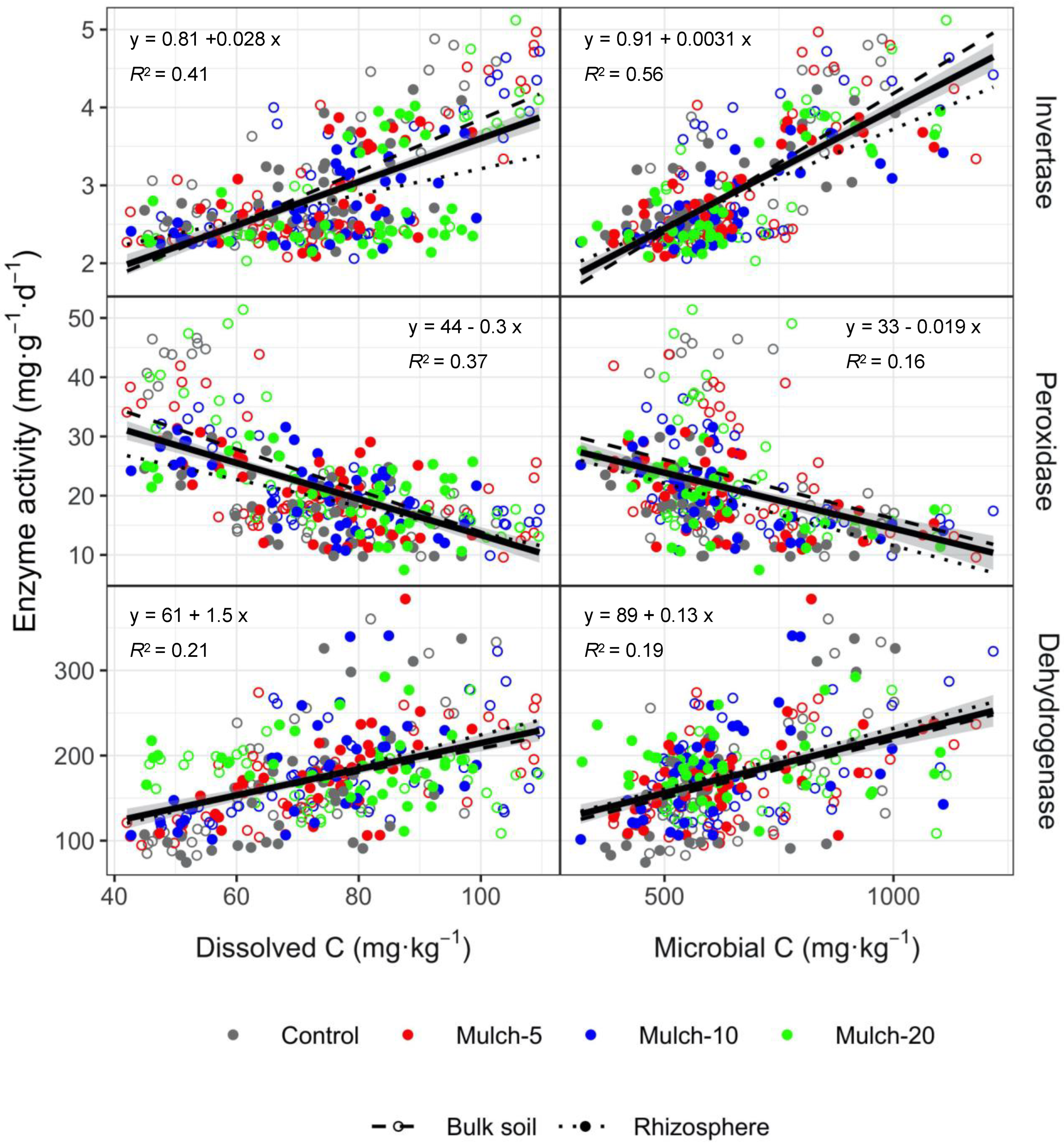
The activities of enzymes in the rhizosphere and bulk soils were closely related and exhibited a similar response to organic mulching (Figure 5 and Figure 6). More specifically, the activity of invertase, as well as the content of MBC and MBN, increased in spring. Meanwhile, peroxidase activity in autumn increased with nitrate and ammonium content, however, decreased with dissolved C and water content. Additionally, the SOC was determined to be the primary factor influencing urease activity in winter (Figure 5). Overall, the dissolved C and MBC were the most important factors causing invertase and dehydrogenase activity to increase and peroxidase activity to decrease (Figure 6).
4. Discussion
4.1. Response of Enzyme Activities in the Rhizosphere and Bulk Soil to Mulching
Organic mulching impacts C- and N-related enzyme activity in the rhizosphere and bulk soils, primarily in autumn and spring. These effects increased with the amount of mulching applied. The activity of invertase, urease, peroxidase, and dehydrogenase in the rhizosphere was closely related to that in the bulk soil, and exhibited similar responses to organic mulching. The activities of rhizosphere enzymes, however, were more sensitive to mulching due to more rapid C and N transformation (Figure 4). This phenomenon indicates enhanced plant–soil elemental exchange—the transfer of nutrients from soil to plant, and the deposition of C from plant to soil—implying that organic mulching promotes plant growth.
Invertase hydrolyzes sucrose to glucose and fructose [23,24]. In the current study, mulching was found to reduce invertase activity within the topsoil rhizosphere while increasing it in the subsoil during spring. Given that organic matter decomposition increases via microbial enzyme production [25], the slightly increased alkaline pH and decreased water content in spring [21] may have created the optimum conditions for invertase activity in the topsoil rhizosphere. Meanwhile, dissolved C content decreases with an increase in microbial biomass and C use in the rhizosphere [26], subsequently reducing invertase activity (Figure 6).
Urease mineralizes organic N to ammonium [24,27]. The urease activity continuously increased in bulk soil after mulching, along with available N (mineral N supply), N mineralization, and ammonium content (Figure 3 and Figure S2). However, the lower ammonium content within the rhizosphere with mulch decreased urease activity when compared with that without mulch. The decreased dissolved N content implies that mulch addition directly increased the available nutrients, which were rapidly utilized by microorganisms. Similar to a previous study [28], we found that SOC plays an important role in regulating enzyme activity. In particular, SOC increased urease activity in winter (Figure 5) following its accumulation in organic mulch. Hence mulching contributes to SOC accumulation and nutrient (N) cycling.
Peroxidase is directly related to the decomposition of refractory SOC components due to its involvement in lignin decomposition [29]. In our study, peroxidase activity decreased with dissolved C (Figure 6). When the abundance of lignin and other refractory substances increases, microorganisms secrete peroxidases [30]. Among the four mulching treatments, Mulch-5 exhibited the highest level of peroxidase activity in spring when compared to the control treatment, whereas Mulch-10 and Mulch-20 exhibited lower activity levels. We speculate that the small amount of mulch can more readily enter the soil with water in spring [31], where it effectively stimulates peroxidase activity. Meanwhile, in autumn, the MBC content began to decrease in bulk soil, thus reducing the MBC/MBN ratio. This was attributed to an increase in bacteria and decrease in fungi for 6 months after the application of organic mulching [32,33]. The average C/N ratio of bacteria is 4 whereas that of fungi is 10 [34]. A reduction in fungal abundance (decreased C/N ratio and MBC/MBN ratio) decreases peroxidase activity as peroxidase is secreted by fungi [35].
Dehydrogenase partially reflects the overall microbial enzyme activity in soil. Mulching increased the dehydrogenase activity in both the rhizosphere and bulk soil in autumn, thus accounting for the increased mineralization of organic substances from both plant root and litter decomposition in autumn. An additional N source provides sufficient N for soil microbes, thus alleviating N competition between microorganisms and plants [36,37] and increasing microbial activity [38,39]. As shown in the current study, dehydrogenase activity increased with dissolved C and MBC (Figure 6).
The observed enzymatic activity differed within the rhizosphere and bulk soils after mulching. That is, the enzyme activities in the rhizosphere exhibited a greater response to organic mulching than those in bulk soil. Indeed, the rhizosphere is directly affected by plant roots, and the organic matter inputted into the rhizosphere by root exudates is readily utilized by microbes [15,40,41]. Moreover, rhizosphere enzymes are directly derived from root secretions and exhibit a decreasing concentration gradient from the roots to soil [16]. However, the balance between root input and microbial decomposition, as well as the impact of other mechanisms associated with enzyme inactivation, might stabilize the activities of rhizosphere enzymes in the short-term [42].
Our results prove the second hypothesis, that is, that the effects induced by mulching on enzymatic activity increase with the amount of mulch applied; however, this trend was not observed for C and N content. This indicates that enzyme activity is more sensitive to mulching than the active and labile C and N fractions, which are readily transformed.
4.2. Factors Affecting Enzyme Activities
Enzyme activities are driven primarily by labile C [43,44]. Indeed, dissolved C and MBC were found to increase the activity of invertase and dehydrogenase, while decreasing that of peroxidase (Figure 6). Similar to previous studies, we found a strong correlation between enzymatic activity and the amount of dissolved organic C released from decomposing litter [44,45,46], as well as with the microbial PLFAs [47]. Season and soil depth—corresponding to the release of available C by roots—were more important factors contributing to a source of persistent C and N than mulching (Tables S1 and S4).
Season determines the soil temperature, water content, root exudation, and microbial activity, thereby controlling soil enzyme activities [48]. More specifically, temperature directly influences enzyme activity through substrate diffusion and by altering the kinetic characteristics of enzymes, whereas it exerts indirect effects by affecting microbial proliferation [49,50]. Generally, an increase in temperature enhances invertase activity associated with C and N; however, C enzymes are more sensitive to temperature than N enzymes [51,52], thus accounting for the greater temperature-related changes observed in invertase activity (Figure 5).
Considering the soil water content, enzyme activity is limited under low moisture conditions, particularly by decreasing substrate diffusion and offsetting the positive effects of high temperatures [40,53,54]. Affected by the distribution of organic C and microbial biomass in the soil profiles, enzyme activity decreases with soil depth [55,56,57]. Thus, in addition to mulching, the season and soil depth were determined to also represent important factors influencing enzymatic activity.
5. Conclusions
Overall, enzymatic activity was found to increase with an increase in the amount of applied organic mulch in an urban forest. Moreover, the rhizosphere invertase activity in spring, and urease activity in autumn decreased with a decrease in dissolved C and available N, respectively. Meanwhile, peroxidase activity increased with mulch addition in spring and decreased in autumn with a decrease in the microbial biomass C/N ratio. Additionally, dehydrogenase activity in autumn increased with dissolved C and MBC. Although the activity of these four enzymes were relatively similar in the rhizosphere and bulk soil, exhibiting similar responses to organic mulching, those in the rhizosphere were more sensitive to mulching due to more rapid C and nutrient cycling, as well as higher root activities. In the spring, increased C and N contents in microbial biomass were associated with an increase in invertase activity, whereas the SOC content determined urease activity in winter. Ammonium and nitrate were determined to be the most important factors contributing to peroxidase activity in autumn. Hence, the factors impacting enzymatic activity are controlled by the season and soil depth via changes in water content, temperature, and nutrient availability. Overall, organic mulch addition is an effective measure to accelerate the C and N cycles in soil and can be especially useful for the amendment of urban soil in parks and other green spaces.
Supplementary Materials
The following supporting information can be downloaded at: https://0-www-mdpi-com.brum.beds.ac.uk/article/10.3390/f13091352/s1, Figure S1: Effects of organic mulching on enzymatic activity in rhizosphere and bulk soil; Figure S2: Effects of organic mulching on C and N fractions in rhizosphere and bulk soil; Table S1: Analysis of variance of the effects of different seasons, soil layers and treatments on enzyme activities in rhizosphere and bulk soil; Table S2: Differences in the enzyme activities under mulching treatments in rhizosphere and bulk soil; Table S3: Differences in the C and N fractions under mulching treatments in rhizosphere and bulk soil; Table S4 Analysis of variance of the effects of different seasons, treatments and soil layers on carbon and nitrogen fractions in rhizosphere and bulk soil.
Author Contributions
Conceptualization, X.S. and Y.K.; methodology, Y.Y.; software, J.L.; validation, X.S. and Y.Y.; formal analysis, X.S.; investigation, X.S.; resources, X.S.; data curation, J.L.; writing—original draft preparation, X.S.; writing—review and editing, Soromotin, A.V.S., Smirnov, P.V.S. and Y.K.; visualization, J.L.; supervision, Y.K.; project administration, X.S. and Y.K.; funding acquisition, X.S. and Y.K. All authors have read and agreed to the published version of the manuscript.
Funding
This study was supported by the Qingdao Postdoctoral Foundation (QDBSH202109); the Program of Competitive Growth of Kazan Federal University, by the “RUDN University Strategic Academic Leadership Program”, the Russian Science Foundation project (19-77-30012), Project TerrArctic (89-DON), and Stability and Functions of Soil Carbon in Agroecosystems of Russia (CarboRus, 075-15-2021-610).
Data Availability Statement
Not available.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Wu, J.; Xiang, W.-N.; Zhao, J. Urban Ecology in China: Historical Developments and Future Directions. Landsc. Urban Plan. 2014, 125, 209–221. [Google Scholar] [CrossRef]
- Xu, M.; He, C.; Liu, Z.; Dou, Y. How Did Urban Land Expand in China between 1992 and 2015? A Multi-Scale Landscape Analysis. PLoS ONE 2016, 11, e0154839. [Google Scholar] [CrossRef]
- Vasenev, V.; Kuzyakov, Y. Urban Soils as Hot Spots of Anthropogenic Carbon Accumulation: Review of Stocks, Mechanisms and Driving Factors. Land Degrad. Dev. 2018, 29, 1607–1622. [Google Scholar] [CrossRef]
- Vogel, J.G.; He, D.; Jokela, E.J.; Hockaday, W.; Schuur, E.A.G. The Effect of Fertilization Levels and Genetic Deployment on the Isotopic Signature, Constituents, and Chemistry of Soil Organic Carbon in Managed Loblolly Pine (Pinus taeda L.) Forests. For. Ecol. Manag. 2015, 355, 91–100. [Google Scholar] [CrossRef]
- Foster, D.; Swanson, F.; Aber, J.; Burke, I.; Brokaw, N.; Tilman, D.; Knapp, A. The Importance of Land-Use Legacies to Ecology and Conservation. BioScience 2003, 53, 77–88. [Google Scholar] [CrossRef]
- Midgley, M.; Anderson, E.; Minor, E. Vacant Lot Plant Establishment Techniques Alter Urban Soil Ecosystem Services. Urban For. Urban Green. 2021, 61, 127096. [Google Scholar] [CrossRef]
- Luna, L.; Vignozzi, N.; Miralles, I.; Solé-Benet, A. Organic Amendments and Mulches Modify Soil Porosity and Infiltration in Semiarid Mine Soils. Land Degrad. Dev. 2018, 29, 1019–1030. [Google Scholar] [CrossRef]
- Piotrowska-Długosz, A. Significance of Enzymes and Their Application in Agriculture; Springer: Cham, Switzerland, 2019; pp. 277–308. ISBN 978-3-030-25022-5. [Google Scholar]
- Utobo, E.B.; Tewari, L. Soil Enzymes as Bioindicators of Soil Ecosystem Status. Appl. Ecol. Environ. Res. 2015, 13, 147–169. [Google Scholar] [CrossRef]
- Ovsepyan, L.; Kurganova, I.; de Gerenyu, V.L.; Kuzyakov, Y. Conversion of Cropland to Natural Vegetation Boosts Microbial and Enzyme Activities in Soil. Sci. Total Environ. 2020, 743, 140829. [Google Scholar] [CrossRef]
- Aponte, H.; Medina, J.; Butler, B.; Meier, S.; Cornejo, P.; Kuzyakov, Y. Soil Quality Indices for Metal(Loid) Contamination: An Enzymatic Perspective. Land Degrad. Dev. 2020, 31, 2700–2719. [Google Scholar] [CrossRef]
- Liu, M.; Xu, X.; Nannipieri, P.; Kuzyakov, Y.; Gunina, A. Diurnal Dynamics Can Modify Plant–Microbial Competition for N Uptake via C Allocation. Biol. Fertil. Soils 2021, 57, 949–958. [Google Scholar] [CrossRef]
- Li, Y.; Wang, C.; Gao, S.; Wang, P.; Qiu, J.; Shang, S. Impacts of Simulated Nitrogen Deposition on Soil Enzyme Activity in a Northern Temperate Forest Ecosystem Depend on the Form and Level of Added Nitrogen. Eur. J. Soil Biol. 2021, 103, 103287. [Google Scholar] [CrossRef]
- Wei, H.; Yan, W.; Quan, G.; Zhang, J.; Liang, K. Soil Microbial Carbon Utilization, Enzyme Activities and Nutrient Availability Responses to Bidens Pilosa and a Non-Invasive Congener under Different Irradiances. Sci. Rep. 2017, 7, 11309. [Google Scholar] [CrossRef]
- Sasse, J.; Martinoia, E.; Northen, T. Feed Your Friends: Do Plant Exudates Shape the Root Microbiome? Trends Plant Sci. 2018, 23, 25–41. [Google Scholar] [CrossRef]
- Kuzyakov, Y.; Razavi, B.S. Rhizosphere Size and Shape: Temporal Dynamics and Spatial Stationarity. Soil Biol. Biochem. 2019, 135, 343–360. [Google Scholar] [CrossRef]
- Jian, S.; Li, J.; Chen, J.; Wang, G.; Mayes, M.A.; Dzantor, K.E.; Hui, D.; Luo, Y. Soil Extracellular Enzyme Activities, Soil Carbon and Nitrogen Storage under Nitrogen Fertilization: A Meta-Analysis. Soil Biol. Biochem. 2016, 101, 32–43. [Google Scholar] [CrossRef]
- Kader, M.A.; Senge, M.; Mojid, M.A.; Ito, K. Recent Advances in Mulching Materials and Methods for Modifying Soil Environment. Soil Tillage Res. 2017, 168, 155–166. [Google Scholar] [CrossRef]
- Zhang, F.; Zhang, W.; Li, M.; Yang, Y.; Li, F.-M. Does Long-Term Plastic Film Mulching Really Decrease Sequestration of Organic Carbon in Soil in the Loess Plateau? Eur. J. Agron. 2017, 89, 53–60. [Google Scholar] [CrossRef]
- Tan, X.; Xie, B.; Wang, J.; He, W.; Wang, X.; Wei, G. County-Scale Spatial Distribution of Soil Enzyme Activities and Enzyme Activity Indices in Agricultural Land: Implications for Soil Quality Assessment. Sci. World J. 2014, 2014, 535768. [Google Scholar] [CrossRef]
- Sun, X.; Ye, Y.; Guan, Q.; Jones, D.L. Organic Mulching Masks Rhizosphere Effects on Carbon and Nitrogen Fractions and Enzyme Activities in Urban Greening Space. J. Soils Sediments 2021, 21, 1621–1632. [Google Scholar] [CrossRef]
- Team, R.C. R: A Language and Environment for Statistical Computing; R Development Core Team: Vienna, Austria, 2019. [Google Scholar]
- Yu, P.; Liu, S.; Han, K.; Guan, S.; Zhou, D. Conversion of Cropland to Forage Land and Grassland Increases Soil Labile Carbon and Enzyme Activities in Northeastern China. Agric. Ecosyst. Environ. 2017, 245, 83–91. [Google Scholar] [CrossRef]
- Wu, L.; Ma, H.; Zhao, Q.; Zhang, S.; Wei, W.; Ding, X. Changes in Soil Bacterial Community and Enzyme Activity under Five Years Straw Returning in Paddy Soil. Eur. J. Soil Biol. 2020, 100, 103215. [Google Scholar] [CrossRef]
- Gavrichkova, O.; Brykova, R.A.; Brugnoli, E.; Calfapietra, C.; Cheng, Z.; Kuzyakov, Y.; Liberati, D.; Moscatelli, M.C.; Pallozzi, E.; Vasenev, V.I. Secondary Soil Salinization in Urban Lawns: Microbial Functioning, Vegetation State, and Implications for Carbon Balance. Land Degrad. Dev. 2020, 31, 2591–2604. [Google Scholar] [CrossRef]
- Tang, H.; Xiao, X.; Li, C.; Shi, L.; Cheng, K.; Li, W.; Wen, L.; Xu, Y.; Wang, K. Microbial Carbon Source Utilization in Rice Rhizosphere Soil with Different Tillage Practice in a Double Cropping Rice Field. Sci. Rep. 2021, 11, 5048. [Google Scholar] [CrossRef]
- Liang, W.; Bin Wu, Z.; Cheng, S.P.; Zhou, Q.H.; Hu, H.Y. Roles of Substrate Microorganisms and Urease Activities in Wastewater Purification in a Constructed Wetland System. Ecol. Eng. 2003, 21, 191–195. [Google Scholar] [CrossRef]
- Jat, H.S.; Datta, A.; Choudhary, M.; Sharma, P.C.; Dixit, B.; Jat, M.L. Soil Enzymes Activity: Effect of Climate Smart Agriculture on Rhizosphere and Bulk Soil under Cereal Based Systems of North-West India. Eur. J. Soil Biol. 2021, 103, 103292. [Google Scholar] [CrossRef]
- Tian, L.; Shi, W. Soil Peroxidase Regulates Organic Matter Decomposition through Improving the Accessibility of Reducing Sugars and Amino Acids. Biol. Fertil. Soils 2014, 50, 785–794. [Google Scholar] [CrossRef]
- Ayuso-Fernández, I.; Rencoret, J.; Gutiérrez, A.; Ruiz-Dueñas, F.J.; Martínez, A.T. Peroxidase Evolution in White-Rot Fungi Follows Wood Lignin Evolution in Plants. Proc. Natl. Acad. Sci. USA 2019, 116, 17900–17905. [Google Scholar] [CrossRef]
- Sun, X.; Wang, G.; Ma, Q.; Liao, J.; Wang, D.; Guan, Q.; Jones, D.L. Organic Mulching Promotes Soil Organic Carbon Accumulation to Deep Soil Layer in an Urban Plantation Forest. For. Ecosyst. 2021, 8, 2. [Google Scholar] [CrossRef]
- Ma, Q.; Wu, L.; Wang, J.; Ma, J.; Zheng, N.; Hill, P.W.; Chadwick, D.R.; Jones, D.L. Fertilizer Regime Changes the Competitive Uptake of Organic Nitrogen by Wheat and Soil Microorganisms: An in-Situ Uptake Test Using 13C, 15N Labelling, and 13C-PLFA Analysis. Soil Biol. Biochem. 2018, 125, 319–327. [Google Scholar] [CrossRef]
- Song, D.; Chen, L.; Zhang, S.; Zheng, Q.; Ullah, S.; Zhou, W.; Wang, X. Combined Biochar and Nitrogen Fertilizer Change Soil Enzyme and Microbial Activities in a 2-Year Field Trial. Eur. J. Soil Biol. 2020, 99, 103212. [Google Scholar] [CrossRef]
- Cleveland, C.C.; Liptzin, D. C:N:P Stoichiometry in Soil: Is There a “Redfield Ratio” for the Microbial Biomass? Biogeochemistry 2007, 85, 235–252. [Google Scholar] [CrossRef]
- Hofrichter, M.; Ullrich, R.; Pecyna, M.J.; Liers, C.; Lundell, T. New and Classic Families of Secreted Fungal Heme Peroxidases. Appl. Microbiol. Biotechnol. 2010, 87, 871–897. [Google Scholar] [CrossRef] [PubMed]
- Jones, D.L.; Clode, P.L.; Kilburn, M.R.; Stockdale, E.A.; Murphy, D.V. Competition between Plant and Bacterial Cells at the Microscale Regulates the Dynamics of Nitrogen Acquisition in Wheat (Triticum Aestivum). New Phytol. 2013, 200, 796–807. [Google Scholar] [CrossRef]
- Kuzyakov, Y.; Xu, X. Competition between Roots and Microorganisms for Nitrogen: Mechanisms and Ecological Relevance. New Phytol. 2013, 198, 656–669. [Google Scholar] [CrossRef]
- Hodge, A.; Fitter, A.H. Microbial Mediation of Plant Competition and Community Structure. Funct. Ecol. 2013, 27, 865–875. [Google Scholar] [CrossRef]
- Wang, R.; Filley, T.R.; Xu, Z.; Wang, X.; Li, M.H.; Zhang, Y.; Luo, W.; Jiang, Y. Coupled Response of Soil Carbon and Nitrogen Pools and Enzyme Activities to Nitrogen and Water Addition in a Semi-Arid Grassland of Inner Mongolia. Plant Soil 2014, 381, 323–336. [Google Scholar] [CrossRef]
- Stock, S.C.; Köster, M.; Dippold, M.A.; Nájera, F.; Matus, F.; Merino, C.; Boy, J.; Spielvogel, S.; Gorbushina, A.; Kuzyakov, Y. Environmental Drivers and Stoichiometric Constraints on Enzyme Activities in Soils from Rhizosphere to Continental Scale. Geoderma 2019, 337, 973–982. [Google Scholar] [CrossRef]
- Yuan, H.; Zhu, Z.; Liu, S.; Ge, T.; Jing, H.; Li, B.; Liu, Q.; Lynn, T.M.; Wu, J.; Kuzyakov, Y. Microbial Utilization of Rice Root Exudates: 13C Labeling and PLFA Composition. Biol. Fertil. Soils 2016, 52, 615–627. [Google Scholar] [CrossRef]
- Ge, T.; Wei, X.; Razavi, B.S.; Zhu, Z.; Hu, Y.; Kuzyakov, Y.; Jones, D.L.; Wu, J. Stability and Dynamics of Enzyme Activity Patterns in the Rice Rhizosphere: Effects of Plant Growth and Temperature. Soil Biol. Biochem. 2017, 113, 108–115. [Google Scholar] [CrossRef] [Green Version]
- Kotroczó, Z.; Veres, Z.; Fekete, I.; Krakomperger, Z.; Tóth, J.A.; Lajtha, K.; Tóthmérész, B. Soil Enzyme Activity in Response to Long-Term Organic Matter Manipulation. Soil Biol. Biochem. 2014, 70, 237–243. [Google Scholar] [CrossRef]
- Wei, L.; Razavi, B.S.; Wang, W.; Zhu, Z.; Liu, S.; Wu, J.; Kuzyakov, Y.; Ge, T. Labile Carbon Matters More than Temperature for Enzyme Activity in Paddy Soil. Soil Biol. Biochem. 2019, 135, 134–143. [Google Scholar] [CrossRef]
- Błońska, E.; Piaszczyk, W.; Staszel, K.; Lasota, J. Enzymatic Activity of Soils and Soil Organic Matter Stabilization as an Effect of Components Released from the Decomposition of Litter. Appl. Soil Ecol. 2021, 157, 103723. [Google Scholar] [CrossRef]
- Liu, S.; Wang, J.; Pu, S.; Blagodatskaya, E.; Kuzyakov, Y.; Razavi, B.S. Impact of Manure on Soil Biochemical Properties: A Global Synthesis. Sci. Total Environ. 2020, 745, 141003. [Google Scholar] [CrossRef]
- Liu, Y.; Yang, F.; Yang, W.; Wu, F.; Xu, Z.; Liu, Y.; Zhang, L.; Yue, K.; Ni, X.; Lan, L.; et al. Effects of Naphthalene on Soil Fauna Abundance and Enzyme Activity in the Subalpine Forest of Western Sichuan, China. Sci. Rep. 2019, 9, 2849. [Google Scholar] [CrossRef]
- Machmuller, M.B.; Mohan, J.E.; Minucci, J.M.; Phillips, C.A.; Wurzburger, N. Season, but Not Experimental Warming, Affects the Activity and Temperature Sensitivity of Extracellular Enzymes. Biogeochemistry 2016, 131, 255–265. [Google Scholar] [CrossRef]
- Daniel, R.M.; Danson, M.J. Temperature and the Catalytic Activity of Enzymes: A Fresh Understanding. FEBS Lett. 2013, 587, 2738–2743. [Google Scholar] [CrossRef]
- Peterson, M.E.; Daniel, R.M.; Danson, M.J.; Eisenthal, R. The Dependence of Enzyme Activity on Temperature: Determination and Validation of Parameters. Biochem. J. 2007, 402, 331–337. [Google Scholar] [CrossRef]
- Wallenstein, M.D.; Mcmahon, S.K.; Schimel, J.P. Seasonal Variation in Enzyme Activities and Temperature Sensitivities in Arctic Tundra Soils. Glob. Chang. Biol. 2009, 15, 1631–1639. [Google Scholar] [CrossRef]
- Razavi, B.S.; Liu, S.; Kuzyakov, Y. Hot Experience for Cold-Adapted Microorganisms: Temperature Sensitivity of Soil Enzymes. Soil Biol. Biochem. 2017, 105, 236–243. [Google Scholar] [CrossRef]
- Steinweg, J.M.; Dukes, J.S.; Wallenstein, M.D. Modeling the Effects of Temperature and Moisture on Soil Enzyme Activity: Linking Laboratory Assays to Continuous Field Data. Soil Biol. Biochem. 2012, 55, 85–92. [Google Scholar] [CrossRef]
- Sanaullah, M.; Blagodatskaya, E.; Chabbi, A.; Rumpel, C.; Kuzyakov, Y. Drought Effects on Microbial Biomass and Enzyme Activities in the Rhizosphere of Grasses Depend on Plant Community Composition. Appl. Soil Ecol. 2011, 48, 38–44. [Google Scholar] [CrossRef]
- Loeppmann, S.; Blagodatskaya, E.; Pausch, J.; Kuzyakov, Y. Enzyme Properties down the Soil Profile—A Matter of Substrate Quality in Rhizosphere and Detritusphere. Soil Biol. Biochem. 2016, 103, 274–283. [Google Scholar] [CrossRef]
- Dick, R.P.; Kandeler, E. Enzymes in Soils. In Encyclopedia of Soils in the Environment; Elsevier: Amsterdam, The Netherlands, 2004; ISBN 9780080547954. [Google Scholar]
- Aponte, H.; Meli, P.; Butler, B.; Paolini, J.; Matus, F.; Merino, C.; Cornejo, P.; Kuzyakov, Y. Meta-Analysis of Heavy Metal Effects on Soil Enzyme Activities. Sci. Total Environ. 2020, 737, 139744. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Differences in enzymatic activities between mulching and control in the rhizosphere and bulk soil. Values represent means ± standard errors (n = 5). Treatments: Mulch-5, Mulch-10, and Mulch-20, corresponding to organic mulching to a depth of 5, 10 and 20 cm, respectively.
Figure 1.
Differences in enzymatic activities between mulching and control in the rhizosphere and bulk soil. Values represent means ± standard errors (n = 5). Treatments: Mulch-5, Mulch-10, and Mulch-20, corresponding to organic mulching to a depth of 5, 10 and 20 cm, respectively.

Figure 2.
Differences in enzyme activities between the rhizosphere and bulk soil based on organic mulching. Values represent mean ± standard error (n = 5). Values above the 0 line indicate higher enzymatic activity in the rhizosphere than in the bulk soil. Treatments: Mulch-5, Mulch-10, and Mulch-20, corresponding to organic mulching to a depth of 5, 10 and 20 cm, respectively.
Figure 2.
Differences in enzyme activities between the rhizosphere and bulk soil based on organic mulching. Values represent mean ± standard error (n = 5). Values above the 0 line indicate higher enzymatic activity in the rhizosphere than in the bulk soil. Treatments: Mulch-5, Mulch-10, and Mulch-20, corresponding to organic mulching to a depth of 5, 10 and 20 cm, respectively.

Figure 3.
Differences in the C and N fractions between mulching treatments and the control in the rhizosphere and bulk soil. Values represent mean ± standard error (n = 5). Values above the 0 line indicate C and N content higher in the rhizosphere than in the bulk soil.
Figure 3.
Differences in the C and N fractions between mulching treatments and the control in the rhizosphere and bulk soil. Values represent mean ± standard error (n = 5). Values above the 0 line indicate C and N content higher in the rhizosphere than in the bulk soil.


Figure 4.
Differences in the C and N fractions between the rhizosphere and bulk soil based on organic mulching. Values represent mean ± standard error (n = 5). Values above the 0 line indicate higher C and N fraction content in the rhizosphere than in the bulk soil.
Figure 4.
Differences in the C and N fractions between the rhizosphere and bulk soil based on organic mulching. Values represent mean ± standard error (n = 5). Values above the 0 line indicate higher C and N fraction content in the rhizosphere than in the bulk soil.


Figure 5.
Redundancy analysis (RDA) ordination plot of enzyme activities constrained by environmental variables that significantly explain the variation. I: invertase; U: urease; P: peroxidase; D: dehydrogenase; T: temperature; W: water content; BD: bulk density; SOC: soil organic C; TN: total N; DC: dissolved C; DN: dissolved N; AN: ammonium; NN: nitrate; MBC: microbial C; MBN: microbial N. “B” means “in bulk soil” and “R” means “in rhizosphere”. The RDA was performed based on original data (Figures S1 and S2). The curved arrow indicates the development of enzyme activities over the year.
Figure 5.
Redundancy analysis (RDA) ordination plot of enzyme activities constrained by environmental variables that significantly explain the variation. I: invertase; U: urease; P: peroxidase; D: dehydrogenase; T: temperature; W: water content; BD: bulk density; SOC: soil organic C; TN: total N; DC: dissolved C; DN: dissolved N; AN: ammonium; NN: nitrate; MBC: microbial C; MBN: microbial N. “B” means “in bulk soil” and “R” means “in rhizosphere”. The RDA was performed based on original data (Figures S1 and S2). The curved arrow indicates the development of enzyme activities over the year.

Figure 6.
Regression analysis for the effects of dissolved C and microbial C on enzyme activity. All regression lines are significant at p < 0.01. The black continuous line with shaded area shows the regression and 95% confidence interval for all points.
Figure 6.
Regression analysis for the effects of dissolved C and microbial C on enzyme activity. All regression lines are significant at p < 0.01. The black continuous line with shaded area shows the regression and 95% confidence interval for all points.


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
Soil physical and chemical properties.
| Soil Depth | pH | Total C (g·kg−1) | Total N (g·kg−1) | NH4+ (mg N·kg−1) | NO3− (mg N·kg−1) | Total P (g·kg−1) | Available P (mg·kg−1) | Total K (g·kg−1) | Available K (g·kg−1) |
|---|---|---|---|---|---|---|---|---|---|
| 0–20 cm | 7.29 ± 0.3 | 13.1 ± 0.6 | 1.58 ± 0.06 | 2.50 ± 0.1 | 1.71 ± 0.07 | 0.40 ± 0.01 | 29.0 ± 1.6 | 1.08 ± 0.04 | 0.14 ± 0.01 |
| 20–40 cm | 7.31 ± 0.3 | 11.2 ± 0.5 | 1.42 ± 0.05 | 1.43 ± 0.08 | 1.69 ± 0.07 | 0.32 ± 0.01 | 23.1 ± 1.2 | 0.95 ± 0.03 | 0.13 ± 0.01 |
Note: Values are mean ± standard error (n = 5).
Table 2.
Physical and chemical properties of the organic mulch from Shanghai Moqi Garden Co., Ltd.
| pH | Electrical Conductivity (mS·cm−1) | Organic Matter (g·kg−1) | Dry Density (Mg·m−3) | Wet Density (Mg·m−3) | Porosity (m3·m−3) | Total C (g·kg−1) | Total N (g·kg−1) | Total P (g·kg−1) | Total K (g·kg−1) |
|---|---|---|---|---|---|---|---|---|---|
| 6.40 | 1.35 | 902 | 0.14 | 0.79 | 318 | 599 | 23.8 | 4.30 | 19.5 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Sun, X.; Ye, Y.; Liao, J.; Soromotin, A.V.; Smirnov, P.V.; Kuzyakov, Y. Organic Mulching Increases Microbial Activity in Urban Forest Soil. Forests 2022, 13, 1352. https://0-doi-org.brum.beds.ac.uk/10.3390/f13091352
AMA Style
Sun X, Ye Y, Liao J, Soromotin AV, Smirnov PV, Kuzyakov Y. Organic Mulching Increases Microbial Activity in Urban Forest Soil. Forests. 2022; 13(9):1352. https://0-doi-org.brum.beds.ac.uk/10.3390/f13091352
Chicago/Turabian StyleSun, Xiaodan, Yuqian Ye, Jiahui Liao, Andrey V. Soromotin, Pavel V. Smirnov, and Yakov Kuzyakov. 2022. "Organic Mulching Increases Microbial Activity in Urban Forest Soil" Forests 13, no. 9: 1352. https://0-doi-org.brum.beds.ac.uk/10.3390/f13091352
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.