
Functional Diversity of Soil Microorganisms and Influencing Factors in Three Typical Water-Conservation Forests in Danjiangkou Reservoir Area
Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Area
2.2. Experimental Design and Sample Collection
2.3. Analysis of Soil Physicochemical Properties
2.4. Determination of Metabolic Characteristics of Soil Microbial Community
2.5. Data Analysis
3. Results
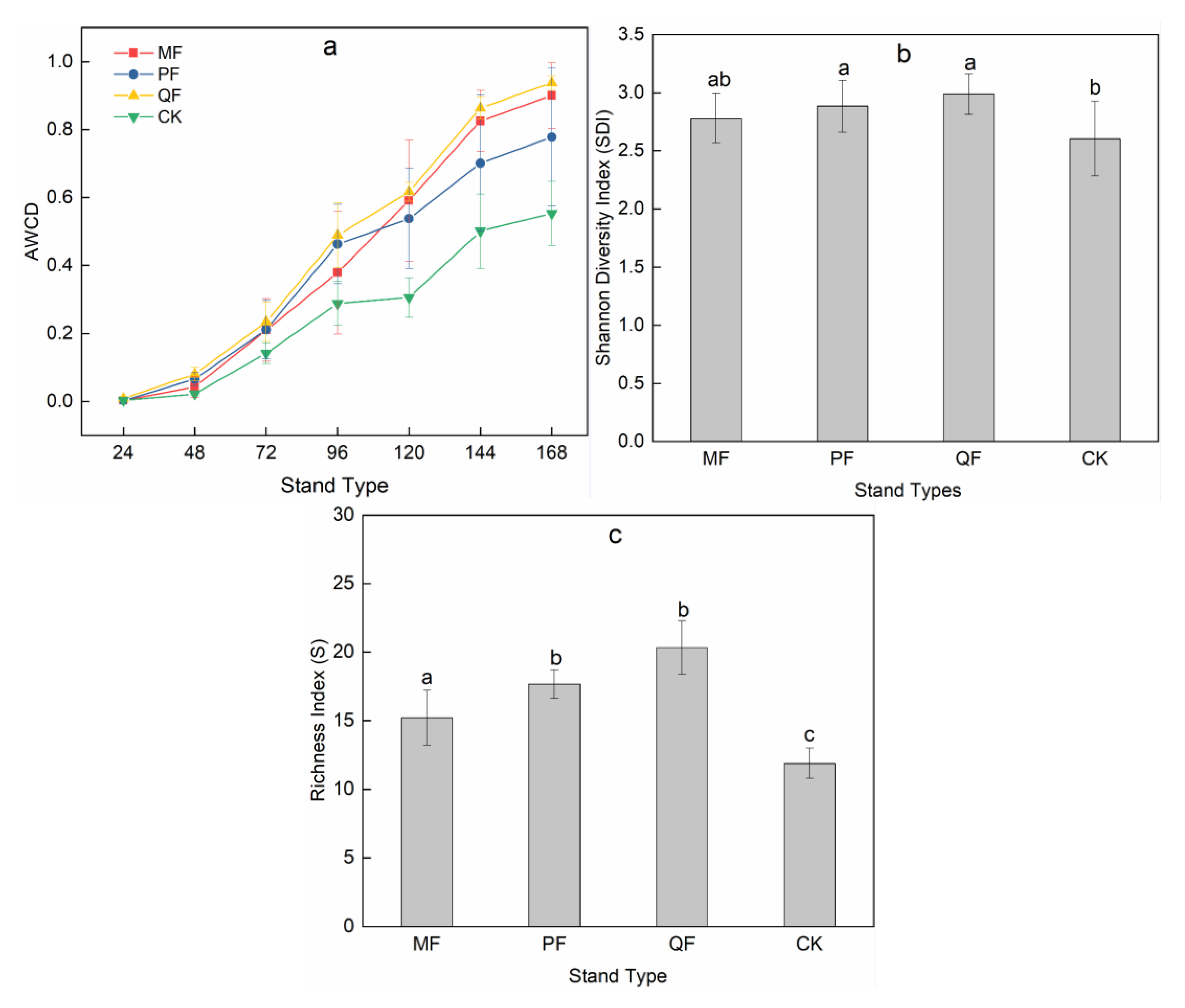
3.1. Plant (Community) Diversity
3.2. Soil Physicochemical Properties
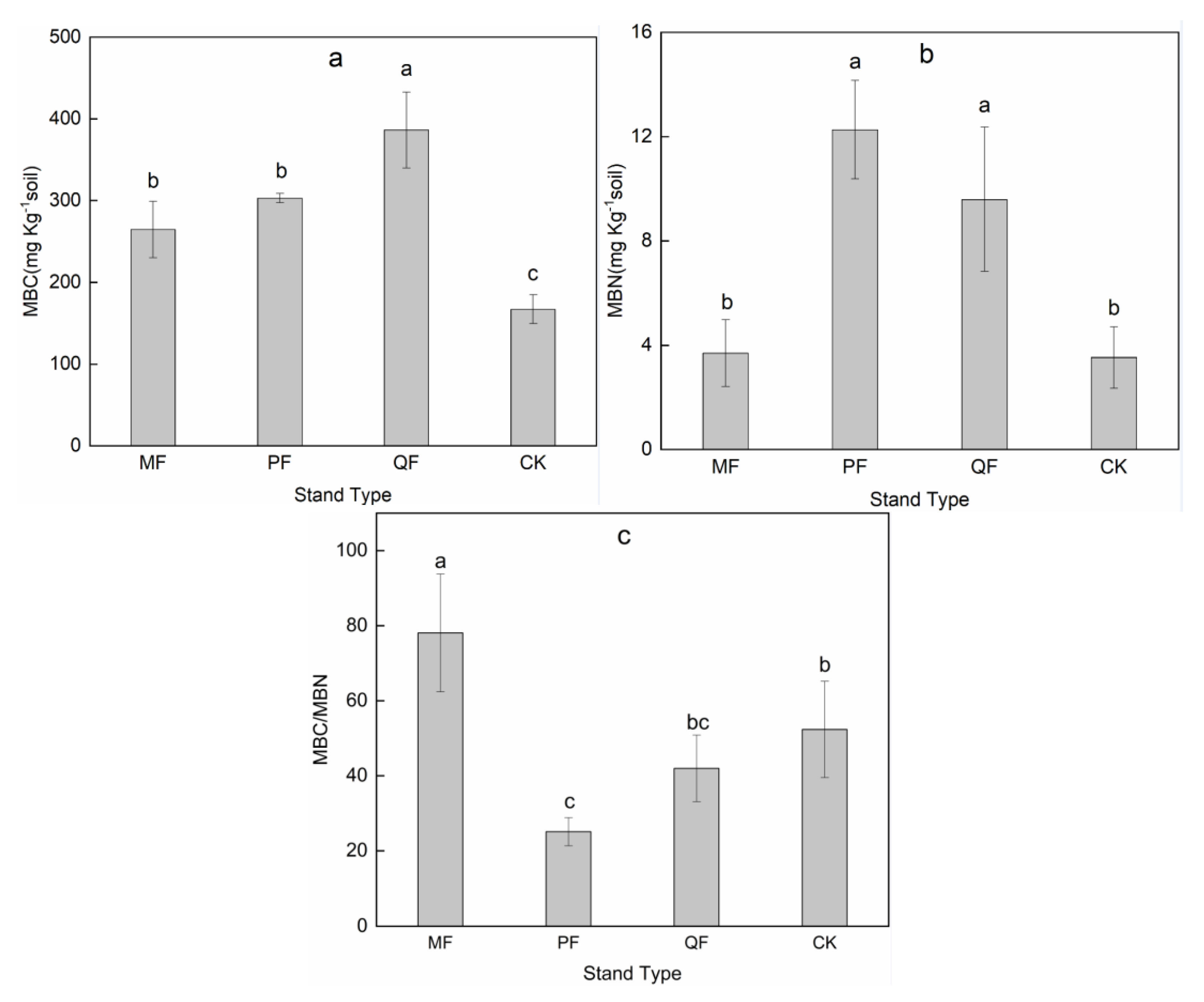
3.3. MBC, MBN and Microbial Biomass C/N Ratios
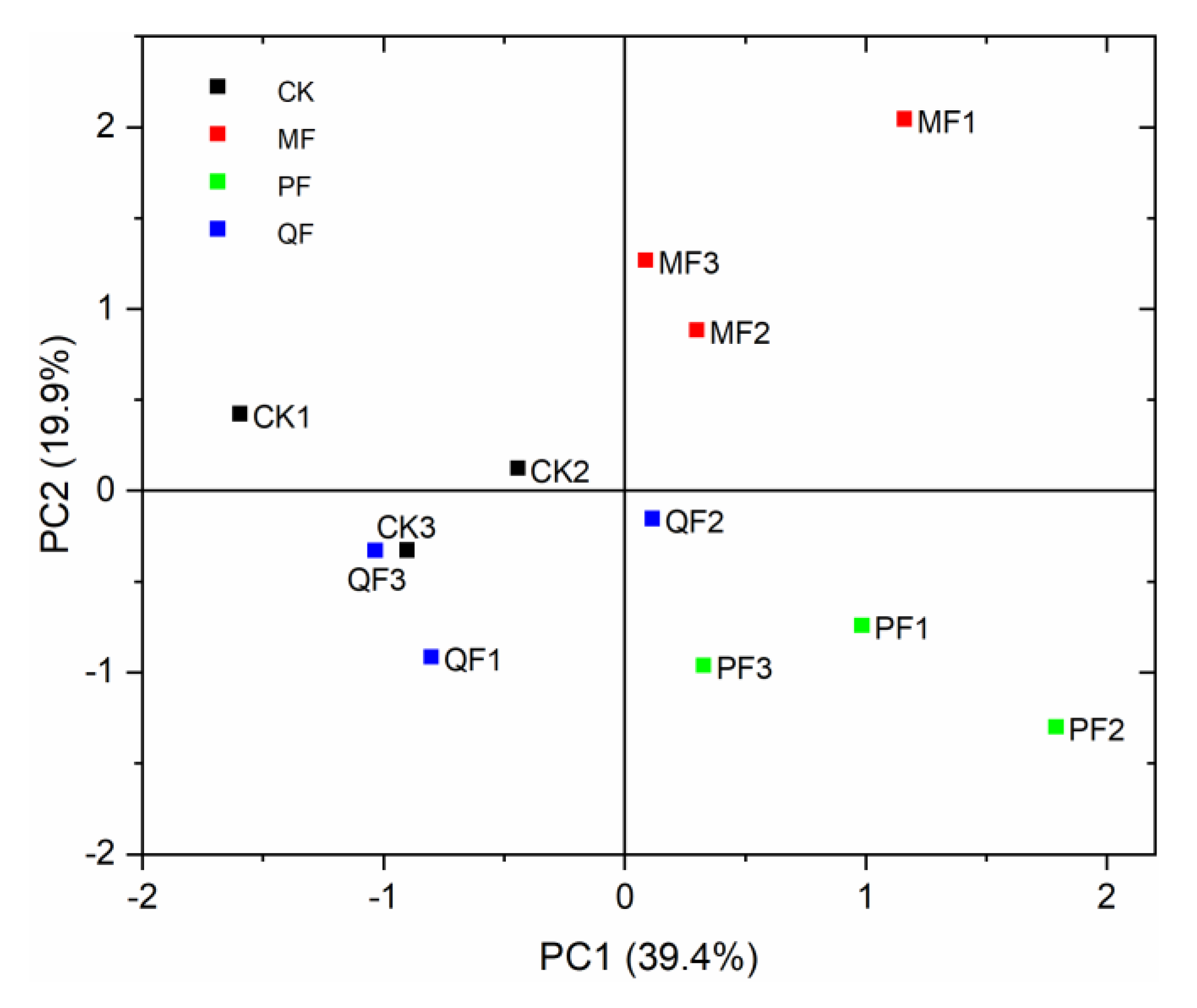
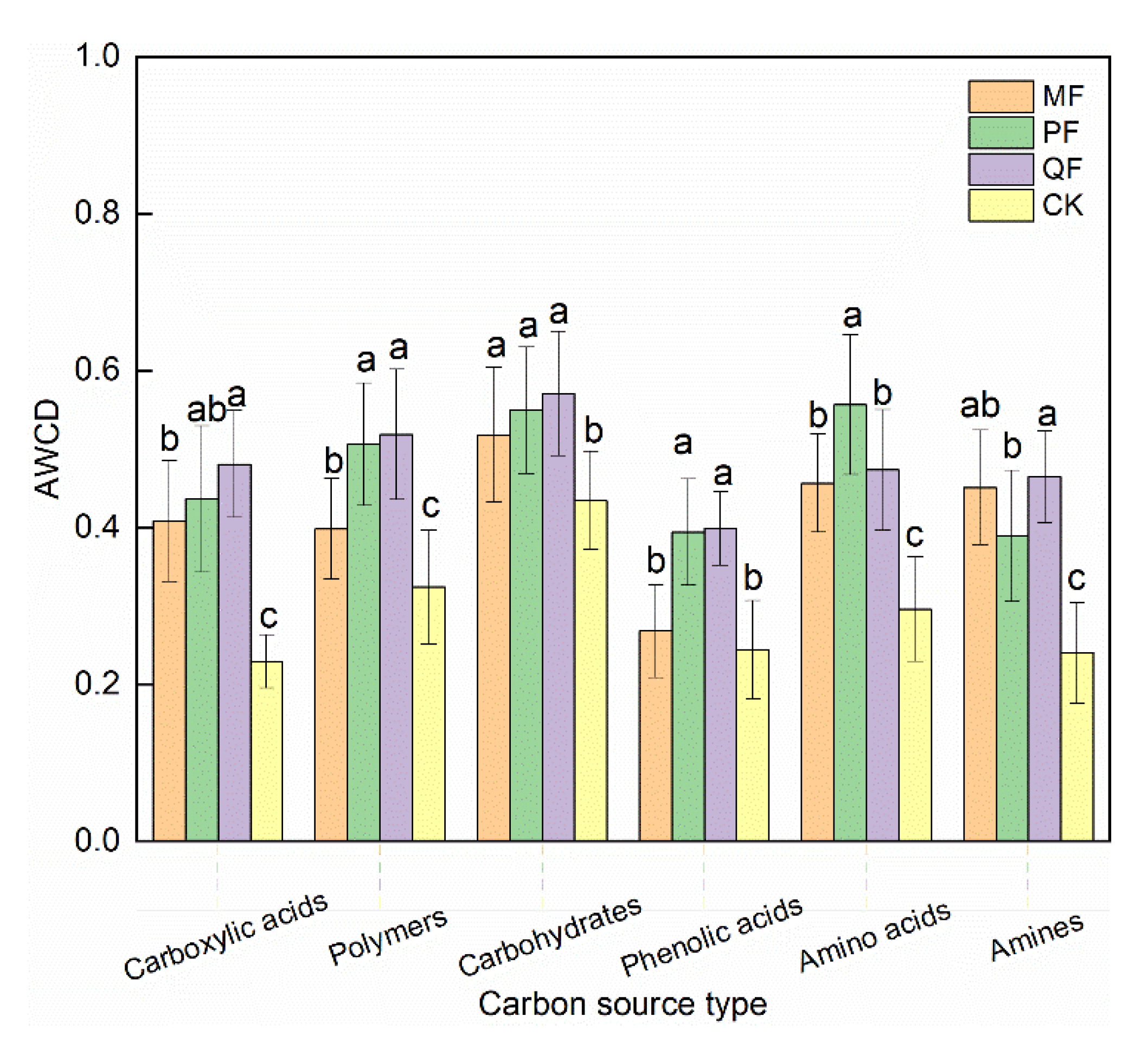
3.4. Functional Diversity of Soil Microorganisms
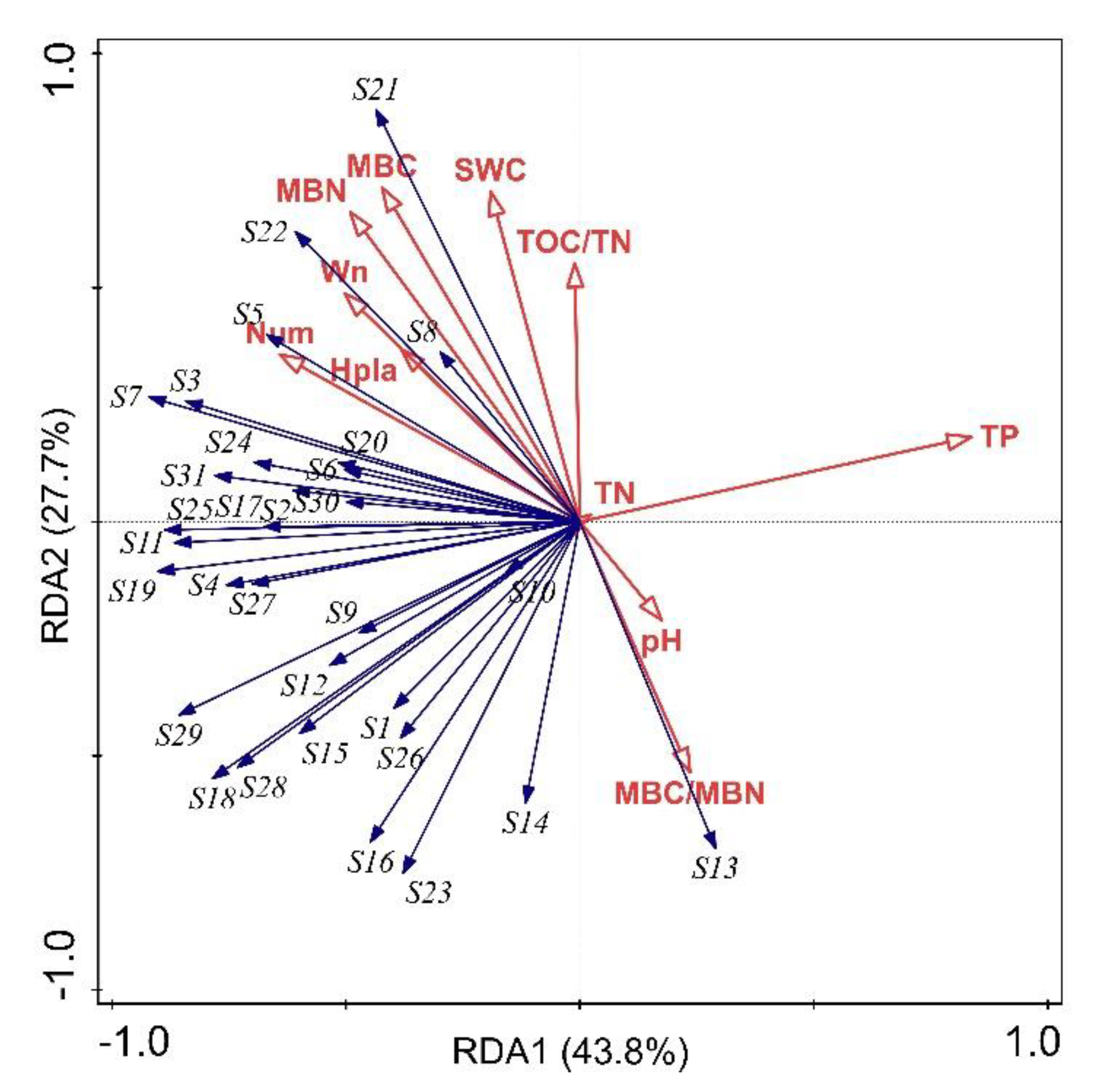
3.5. The Relationship between Soil Microbial Carbon Source Utilization and Environmental Factors
4. Discussion
4.1. Plant (Community) Diversity and Soil Physicochemical Properties
4.2. MBC, MBN, and Microbial Biomass C/N Rations
4.3. Functional Diversity of Soil Microorganisms
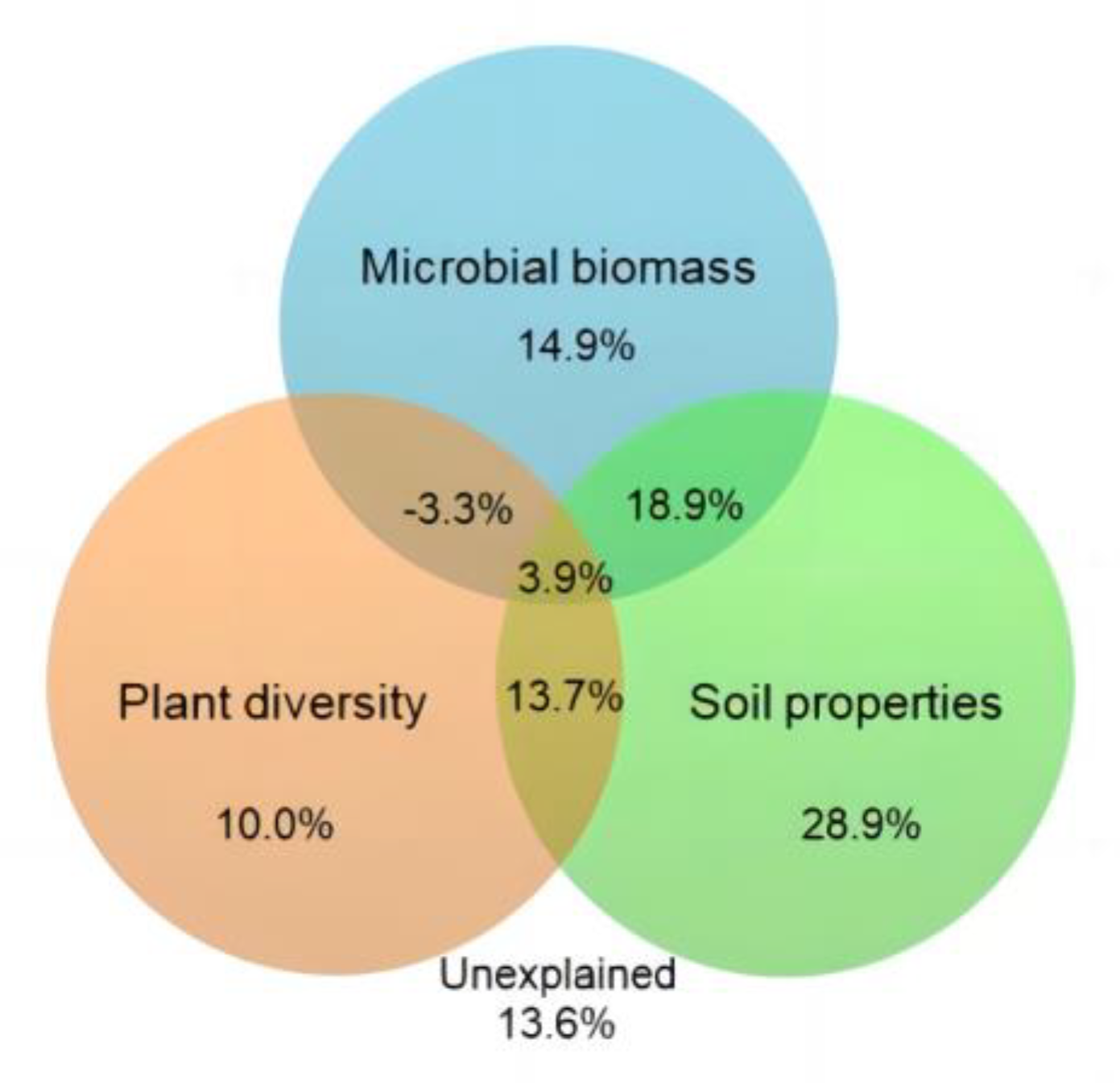
4.4. Relationship between Soil Microbial Carbon Source Utilization and Environmental Factors
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Gao, C.; Yu, X. Review on researchs of water conservation forests. J. Beijing For. Univ. 2000, 22, 78–82. (In Chinese) [Google Scholar]
- Gyssels, G.; Poesen, J.; Bochet, E.; Li, Y. Impact of plant roots on the resistance of soils to erosion by water: A review. Prog. Phys. Geogr. 2005, 29, 189–217. [Google Scholar] [CrossRef] [Green Version]
- Bian, F.Y.; Zhong, Z.K.; Zhang, X.P.; Li, Q.L.; Huang, Z.Y. Bamboo-based agroforestry changes phytoremediation efficiency by affecting soil properties in rhizosphere and non-rhizosphere in heavy metal-polluted soil (Cd/Zn/Cu). J. Soils Sediments 2022, 1–11. [Google Scholar] [CrossRef]
- Kong, F.X.; Sang, W.L.; Jiang, X.; Wang, L.S. Aluminum toxicity and tolerance in plants. Acta Ecol. Sin. 2000, 20, 855–862. [Google Scholar]
- Li, X.B.; Xia, L.L.; Yan, X.Y. Application of membrane inlet mass spectrometry to directly quantify denitrification in flooded rice paddy soil. Biol. Fert. Soils 2014, 50, 891–900. [Google Scholar] [CrossRef]
- Wu, Z.B.; Liang, W.; Qiu, D.R.; Zhou, Q.H.; Cheng, S.P.; Fu, G.P.; He, F. The Correlation between the substrate enzymatic activities and purification effect of wastewater in the integrated vertical constructed wetland. Acta Ecol. Sin. 2002, 22, 1012–1017. [Google Scholar]
- Smith, J.L.; Paul, E.A. The significance of soil microbial biomass estimations. In Soil Biochemistry; Bollag, G.M., Stotzkey, G., Eds.; Chemical Rubber Company Press: Boca Raton, FL, USA, 1990; Volume 6, pp. 357–396. [Google Scholar]
- Chen, F.L.; Zheng, H.; Zhang, K.; Ouyang, Z.Y.; Lan, J.; Li, H.L.; Qian, S. Changes in soil microbial community structure and metabolic activity following conversion from native Pinus massoniana plantations to exotic Eucalyptus plantations. For. Ecol. Manag. 2013, 291, 65–72. [Google Scholar] [CrossRef]
- Delgado-Baquerizo, M.; Maestre, F.T.; Reich, P.V.B.; Jeffries, T.C.; Gaitan, J.J.; Encinar, D.; Berdugo, M.; Campbell, C.D.; Singh, B.K. Microbial diversity drives multifunctionality in terrestrial ecosystems. Nat. Commun. 2016, 7, 10541. [Google Scholar] [CrossRef] [Green Version]
- Waring, B.G.; Averill, C.; Hawkes, C.V. Differences in fungal and bacterial physiology alter soil carbon and nitrogen cycling: Insights from meta-analysis and theoretical models. Ecol. Lett. 2013, 16, 887–894. [Google Scholar] [CrossRef] [PubMed]
- Bauhus, J.; Khanna, P.K. The significance of microbial biomass in forest soils. In Going Underground-Ecological Studies in Forest Soils; Rastin, N., Bauhus, J., Eds.; Research Signpost: Trivandrum, India, 1999; pp. 77–110. [Google Scholar]
- Bucher, A.E.; Lanyon, L.E. Evaluating soil management with microbial community-level physiological profiles. Appl. Soil Ecol. 2005, 29, 59–71. [Google Scholar] [CrossRef]
- Rogers, B.F.; Tate, R.L. Temporal analysis of the soil microbial community along a top sequence in Pineland soils. Soil Biol. Biochem. 2001, 33, 1389–1401. [Google Scholar] [CrossRef]
- Liu, Q.; Pang, Z.Q.; Yang, Z.L.; Nyumah, F.; Hu, C.H.; Lin, W.X.; Yuan, Z.N. Bio-fertilizer Affects Structural Dynamics, Function, and Network Patterns of the Sugarcane Rhizospheric Microbiota. Microb. Ecol. 2022, 84, 1195–1211. [Google Scholar] [CrossRef] [PubMed]
- Garcia-Pausas, J.; Paterson, E. Microbial community abundance and structure are determinants of soil organic matter mineralisation in the presence of labile carbon. Soil Biol. Biochem. 2011, 43, 1705–1713. [Google Scholar] [CrossRef]
- Degens, B.P.; Harris, J.A. Development of a physiological approach to measuring the catabolic diversity of soil microbial communities. Soil Biol. Biochem. 1997, 29, 1309–1320. [Google Scholar] [CrossRef]
- Chapman, S.J.; Campbell, C.D.; Artz, R.R. Assessing CLPPs using MicroRespTM-A comparison with biolog and multi-SIR. J. Soils Sediments 2007, 7, 406–410. [Google Scholar] [CrossRef]
- Lu, S.B.; Guo, X.M.; Rui, Y.C.; Zhou, X.Q.; Chen, C.R.; Xu, Z.H.; Niu, D.K. Utilization of carbon sources by the soil microbial communities of different forest types in subtropical Australia. Acta Ecol. Sin. 2012, 32, 2819–2826. [Google Scholar]
- Selmants, P.C.; Hart, S.C.; Boyle, S.I.; Stark, J.M. Red alder (Alnus rubra) alters community-level soil microbial function in conifer forests of the Pacific Northwest, USA. Soil Biol. Biochem. 2005, 37, 1860–1868. [Google Scholar] [CrossRef]
- Cheng, H.; Xu, W.N.; Luo, T.; Xia, L.; Xiang, H.Y.; Ma, P.F.; Xia, D. Microbial functional diversity as affected by different engineered eco-restoration methods at Xiangjiaba hydropower station. J. Environ. Eng. 2020, 146, 04019125. [Google Scholar] [CrossRef]
- Wang, Y.; Ouyang, Z.Y.; Zheng, H.; Wang, X.K.; Chen, F.L.; Zeng, J. Carbon metabolism of soil microbial communities of restored forests in Southern China. J. Soils Sediments 2011, 11, 789–799. [Google Scholar] [CrossRef]
- Huang, X.M.; Liu, S.R.; Wang, H.; Hu, Z.D.; Li, Z.G.; You, Y.M. Changes of soil microbial biomass carbon and community composition through mixing nitrogen fixing species with Eucalyptus urophylla in subtropical China. Soil Biol. Biochem. 2014, 73, 42–48. [Google Scholar] [CrossRef]
- Chen, X.L.; Wang, D.; Chen, X.; Wang, J.; Diao, J.J.; Zhang, J.Y.; Guan, Q.W. Soil microbial functional diversity and biomass as affected by different thinning intensities in a Chinese fir plantation. Appl. Soil Ecol. 2015, 92, 35–44. [Google Scholar] [CrossRef]
- Wang, X.; Gao, S.H.; Chen, J.Q.; Yao, Z.W.; Zhang, L.; Wu, H.L.; Shu, Q.; Zhang, X.D. Response of Functional Diversity of Soil Microbial Community to Forest Cutting and Regeneration Methodology in a Chinese Fir Plantation. Forests 2022, 13, 360. [Google Scholar] [CrossRef]
- Wang, Y.; Zong, N.; He, N.P.; Zhang, J.J.; Tian, J.; Li, L.T. Soil microbial functional diversity patterns and drivers along an elevation gradient on Qinghai-Tibet China. Acta Ecol. Sin. 2018, 38, 9. [Google Scholar]
- Xie, Y.; Guo, F.Y.; Chen, L.H.; Cao, B. Effects of Elevated CO2 Concentration on Soil Microbial Functional Diversity and Carbon Source Utilization Characteristics in the Root Zone of Lycium barbarum. Sci. Silv. Sin. 2018, 57, 163–172. [Google Scholar]
- Babur, E.; Dindaroglu, T.; Danish, S.; Häggblom, M.M.; Ozlu, E.; Gozukara, G.; Uslu, O.S. Spatial responses of soil carbon stocks, total nitrogen, and microbial indices to post-wildfire in the Mediterranean red pine forest. J. Environ. Manag. 2022, 320, 115939. [Google Scholar] [CrossRef]
- Zhang, G.S.; Yu, X.B.; Li, Y.; Liu, Y.; Zhang, H.; Jia, Y.F.; Xia, S.X. Effects of Burning on Carbon Utilization of Soil Microorganisms and Plant Growth of Carex brevicuspis Communities at Lake Poyang Wetlands, China. Wetlands 2019, 39, 1–15. [Google Scholar] [CrossRef]
- Zhu, P.; Chen, R.S.; Song, Y.X.; Liu, G.X.; Chen, T.; Zhang, W. Effects of land cover conversion on soil properties and soil microbial activity in an alpine meadow on the Tibetan Plateau. Environ. Earth Sci. 2015, 74, 4523–4533. [Google Scholar] [CrossRef]
- Wei, Y.C.; Ouyang, Z.Y.; Miao, H.; Zheng, H. Exotic Pinus carbaea causes soil quality to deteriorate on former abandoned land compared to an indigenous Podocarpus plantation in the tropical forest area of southern China. J. For. Res. 2009, 14, 221–228. [Google Scholar] [CrossRef]
- Burton, J.; Chen, C.R.; Xu, Z.H.; Ghadiri, H. Soil microbial biomass, activity and community composition in adjacent native and plantation forests of subtropical Australia. J. Soils Sediments 2010, 10, 1267–1277. [Google Scholar] [CrossRef]
- Ibell, P.T.; Xu, Z.H.; Blumfield, T.J. Effects of weed control and fertilization on soil carbon and nutrient pools in an exotic pine plantation of subtropical Australia. J. Soils Sediments 2010, 10, 1027–1038. [Google Scholar] [CrossRef]
- Ladygina, N.; Hedlund, K. Plant species influence microbial diversity and carbon allocation in the rhizosphere. Soil Biol. Biochem. 2010, 42, 162–168. [Google Scholar] [CrossRef]
- Ma, L.; Guo, C.; Lu, X.; Yuan, S.; Wang, R. Soil moisture and land use are major determinants of soil microbial community composition and biomass at a regional scale in northeastern China. Biogeosciences 2015, 12, 2585–2596. [Google Scholar] [CrossRef] [Green Version]
- Tian, Q.Y.; Liu, N.N.; Bai, W.M.; Li, L.H.; Chen, J.Q.; Reich, P.B.; Yu, Q.; Guo, D.L.; Smith, M.D.; Knapp, A.K.; et al. A novel soil manganese mechanism drives plant species loss with increased nitrogen deposition in a temperate steppe. Ecology 2016, 97, 65–74. [Google Scholar] [CrossRef] [PubMed]
- Tu, B.; Domene, X.; Yao, M.J.; Li, C.N.; Zhang, S.H.; Kou, Y.P.; Wang, Y.S.; Li, X.Z. Microbial diversity in Chinese temperate steppe: Unveiling the most influential environmental drivers. FEMS Microbiol. Ecol. 2017, 93, fix031. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Carney, K.M.; Matson, P.A. Plant communities, soil microorganisms, and soil carbon cycling: Does altering the world belowground matter to ecosystem functioning? Ecosystems 2005, 8, 928–940. [Google Scholar] [CrossRef]
- Wardle, D.A.; Lavelle, P. Linkages between soil biota, plant litter quality and decomposition. In Driven by Nature: Plant Litter Quality and Decomposition; Cadisch, G., Giller, K.E., Eds.; CAB International: Oxon, UK, 1997; pp. 107–124. [Google Scholar]
- Ozlu, E.; Arriaga, F.J.; Bilen, S.; Gozukara, G.; Babur, E. Carbon Footprint Management by Agricultural Practices. Biology 2022, 11, 1453. [Google Scholar] [CrossRef] [PubMed]
- Chen, H.; Guo, S.L.; Xu, C.Y.; Singh, V.P. Historical temporal trends of hydro-climatic variables and runoff response to climate variability and their relevance in water resource management in the Hanjiang basin. J. Hydrol. 2007, 344, 171–184. [Google Scholar] [CrossRef]
- Zhu, Y.P.; Zhang, H.P.; Chen, L.; Zhao, J.F. Influence of the south-north water diversion project and the mitigation projects on the water quality of Han River. Sci. Total Environ. 2008, 406, 57–68. [Google Scholar] [CrossRef]
- Ding, X.; Cheng, C.J.; Qi, L.H.; Zhang, J.; Lei, G.; Liu, X.Q. Evaluation of water conservation capacity of Pinus massoniana plantation with different densities in Hubei water source area of Danjiangkou reservoir area. Chin. J. Ecol. 2019, 38, 2291–2301. (In Chinese) [Google Scholar]
- Cheng, C.J.; Zhang, J.; Lei, G.; Ding, X.; Liu, X.Q.; Qi, L.H. Interception of N and P by the buffer zone of waterfront vegetation in Danjiangkou reservoir area of Hubei. Sci. Silvae Sin. 2020, 56, 12–20. (In Chinese) [Google Scholar]
- Bao, S.D. Soil Agrochemical Analysis, 3rd ed.; China Agriculture Press: Beijing, China, 2000. [Google Scholar]
- Brookes, P.C.; Landman, A.; Pruden, G.; Jenkinson, D.S. Chloroform fumigation and the release of soil nitrogen: A rapid direct extraction method to measure microbial biomass nitrogen in soil. Soil Biol. Biochem. 1985, 17, 837–842. [Google Scholar] [CrossRef]
- Joergensen, R.G.; Mueller, T. The fumigation-extraction method to estimate soil microbial biomass: Calibration of the kEN value. Soil Biol. Biochem. 1996, 28, 33–37. [Google Scholar] [CrossRef]
- Zabinski, C.A.; Gannon, J.E. Effects of recreational impacts on soil microbial communities. Environ. Manag. 1997, 21, 233–238. [Google Scholar] [CrossRef] [PubMed]
- Pengthamkeerati, P.; Motavalli, P.; Kremer, R. Soil microbial activity and functional diversity changed by compaction, poultry litter and cropping in a claypan soil. Appl. Soil Ecol. 2011, 48, 71–80. [Google Scholar] [CrossRef]
- Geijn, S.; Veen, J. Implications of increased carbon dioxide levels for carbon input and turnover in soils. Vegetatio 1993, 104/105, 283–292. [Google Scholar] [CrossRef]
- Stephan, A.; Meyer, A.H.; Schmid, B. Plant diversity affects culturable soil bacteria in experimental grassland communities. J. Ecol. 2000, 22, 988–998. [Google Scholar] [CrossRef]
- Haack, S.K.; Garchow, H.; Klug, M.J.; Forney, L.J. Analysis of factors affecting the accuracy, reproducibility, and interpretation of microbial community carbon source utilization patterns. Appl. Environ. Microbiol. 1995, 61, 1458–1468. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Grayston, S.J.; Campbell, C.D. Functional biodiversity of microbial communities in the rhizospheres of hybrid larch (Larix eurolepis) and Sitka spruce (Picea sitchensis). Tree Physiol. 1996, 16, 1031–1038. [Google Scholar] [CrossRef]
- Kim, K.Y.; Jordan, D.; Mcdonald, G.A. Enterobacter agglomerans, phosphate solubilizing bacteria, and microbial activity in soil: Effect of carbon sources. Soil Biol. Biochem. 1998, 30, 995–1003. [Google Scholar] [CrossRef]
- Wardle, D.A.; Nicholson, K.S. Synergistic effects of grassland plant species on soil microbial biomass and activity: Implications for ecosystem-level effects of enriched plant diversity. Funct. Ecol. 1996, 10, 410–416. [Google Scholar] [CrossRef]
- Marcin, C.; Klimek, B.; Azarbad, H.; Jazwa, M. Functional diversity of soil microbial communities under Scots pine, Norway spruce, silver birch and mixed boreal forests. Pedobiologia 2015, 58, 81–88. [Google Scholar]
- Hansson, K.; Olsson, B.A.; Olsson, M.; Johansson, U.; Kleja, D.B. Differences in soil properties in adjacent stands of Scots pine, Norway spruce and silver birch in SW Sweden. For. Ecol. Manag. 2011, 262, 522–530. [Google Scholar] [CrossRef]
- Paluch, J.; Gruba, P. Inter-crown versus under-crown area: Contribution of local configuration of trees to variation in topsoil morphology, pH and moisture in Abies alba Mill. forests. Eur. J. For. Res. 2012, 131, 857–870. [Google Scholar] [CrossRef] [Green Version]
- Galka, B.; Labaz, B.; Bogacz, A.; Bojko, O.; Kabala, C. Conversion of Norway spruce forests will reduce organic carbon pools in the mountain soils of SW Poland. Geoderma 2014, 213, 287–295. [Google Scholar] [CrossRef]
- Garau, G.; Morillas, L.; Roales, J.; Castaldi, P.; Mangia, N.P.; Spano, D.; Mereu, S. Effect of monospecific and mixed Mediterranean tree plantations on soil microbial community and biochemical functioning. Appl. Soil Ecol. 2019, 140, 78–88. [Google Scholar] [CrossRef]
- Gruba, P.; Mulder, J. Relationship between aluminum in soils and soil water in mineral horizons of a range of acid forest soils. Soil Sci. Soc. Am. J. 2008, 72, 1150–1157. [Google Scholar] [CrossRef]
- Mueller, K.E.; Eissenstat, D.M.; Hobbie, S.E.; Oleksyn, J.; Jagodzinski, A.M.; Reich, P.B.; Chadwick, O.A.; Chorover, J. Tree species effects on coupled cycles ofcarbon, nitrogen, and acidity in mineral soils at a common garden experiment. Biogeochemistry 2012, 111, 601–614. [Google Scholar] [CrossRef]
- Bolan, N.S.; Hedley, M.J.; White, R.E. Processes of soil acidification during nitrogen cycling with emphasis on legume based pastures. Plant Soil. 1991, 134, 53–63. [Google Scholar] [CrossRef]
- Shrestha, P.; Gautam, R.; Ashwath, N. Effects of agronomic treatments on functional diversity of soil microbial community and microbial activity in a revegetated coal mine spoil. Geoderma 2019, 338, 40–47. [Google Scholar] [CrossRef]
- Devi, N.B.; Yadava, P.S. Seasonal dynamics in soil microbial biomass C, N and P in a mixed-oak forest ecosystem of Manipur, North-east India. Appl. Soil Ecol. 2006, 31, 220–227. [Google Scholar] [CrossRef]
- Bolat, İ. The effect of thinning on microbial biomass C, N and basal respiration in black pine forest soils in Mudurnu. Turk. Eur. J. Oper. Res. 2014, 133, 131–139. [Google Scholar] [CrossRef]
- Zheng, H.; Ouyang, Z.Y.; Wang, X.K.; Fang, Z.G.; Zhao, T.Q.; Miao, H. Effects of regenerating forest cover on soil microbial communities: A case study in hilly red soil region, Southern China. For. Ecol. Manag. 2005, 217, 244–254. [Google Scholar] [CrossRef]
- Nayyar, A.; Hamel, C.; Lafond, G.; Gossen, B.D.; Hanson, K.; Germida, J. Soil microbial quality associated with yield reduction in continuous-pea. Appl. Soil Ecol. 2009, 43, 115–121. [Google Scholar] [CrossRef]
- Zak, D.R.; Holmes, W.E.; White, D.C.; Peacock, A.D.; Tilman, D. Plant diversity, soil microbial communities, and ecosystem function: Are there any links? Ecology 2003, 84, 2042–2050. [Google Scholar] [CrossRef] [Green Version]
- Broughton, L.C.; Gross, K.L. Patterns of diversity in plant and soil microbial communities along a productivity gradient in a Michigan old-field. Oecologia 2000, 125, 420–427. [Google Scholar] [CrossRef]
- Maly, S.; Korthals, G.W.; Van Dijk, C.; Van der Putten, W.H.; De Boer, W. Effect of vegetation manipulation of abandoned arable land on soil microbial properties. Biol. Fertil. 2000, 31, 121–127. [Google Scholar] [CrossRef]
- Jiang, Y.M.; Chen, C.R.; Xu, Z.H.; Liu, Y.Q. Effects of single and mixed species forest ecosystems on diversity and function of soil microbial community in subtropical China. J. Soils Sediments 2012, 12, 228–240. [Google Scholar] [CrossRef]
- Song, X.C.; Wang, H.L.; Qin, W.D.; Deng, X.J.; Tian, H.D.; Tan, Y.B.; Wang, S.N.; Cao, J.Z. Effects of stand type of artificial forests on soil microbial functional diversity. Chin. J. Appl. Ecol. 2019, 30, 841–848. (In Chinese) [Google Scholar]
- Zhou, J.; Kang, S.; Schadt, C.D.; Garten, C.T.J. Spatial scaling of functional gene diversity across various microbial taxa. Proc. Natl. Acad. Sci. USA 2008, 105, 7768–7773. [Google Scholar] [CrossRef] [Green Version]
- Liang, Y.; Nostrand, J.D.; Deng, Y.; He, Z.; Wu, L.; Xu, Z.; Li, G.; Zhou, J. Functional gene diversity of soil microbial communities from five oil-contaminated fields in China. ISME J. 2011, 5, 403–413. [Google Scholar] [CrossRef] [PubMed]
- Menyailo, O.V.; Hungate, B.A.; Zech, W. The effect of single tree species on soil microbial activities related to C and N cycling in the Siberian artificial afforestation experiment. Plant Soil. 2002, 242, 183–196. [Google Scholar] [CrossRef]
- Tripahti, B.M.; Kim, M.; Singh, D.; Lee-Cruz, L.; Lai-Hoe, A.; Ainuddin, A.N.; Go, R.; Rahim, R.A.; Husni, M.H.; Chun, J.; et al. Tropical soil bacterial communities in Malaysia: pH dominates in the equatorial tropics too. Microb. Ecol. 2012, 64, 474–484. [Google Scholar] [CrossRef]
- Chodak, M.; Gołebiewski, M.; Morawska-Płoskonka, J.; Kuduk, K.; Niklinska, M. Diversity of microorganisms from forest soils differently polluted with heavy metals. Appl. Soil Ecol. 2013, 64, 7–14. [Google Scholar] [CrossRef]
- Firestone, W. Microbial Community Utilization of Recalcitrant and Simple Carbon Compounds: Impact of Oak-Woodland Plant Communities. Oecologia 2004, 138, 275–284. [Google Scholar]
- Drenovsky, R.E.; Vo, D.; Graham, K.J.; Scow, K.M. Soil Water Content and Organic Carbon Availability Are Major Determinants of Soil Microbial Community Composition. Microb. Ecol. 2004, 48, 424–430. [Google Scholar] [CrossRef] [PubMed]
- Prevost-Boure, N.C.; Maron, P.A.; Ranjard, L.; Nowak, V.; Dufrene, E.; Damesin, C.; Soudani, K.; Lata, J.C. Seasonal dynamics of the bacterial community in forest soils under different quantities of leaf litter. Appl. Soil Ecol. 2011, 47, 14–23. [Google Scholar] [CrossRef]
- Ushio, M.; Kitayama, K.; Balser, T.C. Tree species effects on soil enzyme activities through effects on soil physicochemical and microbial properties in a tropical montane forest on Mt. Kinabalu, Borneo. Pedobiologia 2010, 53, 227–233. [Google Scholar] [CrossRef]
- Myers, R.T.; Zak, D.R.; White, D.C.; Peacock, A. Landscape-level patterns of microbial community composition and substrate use in upland forest ecosystems. Soil Sci. Soc. Am. J. 2001, 65, 359–367. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| MF | PF | QF | CK | |
|---|---|---|---|---|
| Height (m) | 10.8 ± 0.8 | 12.3 ± 0.5 | 11.1 ± 0.4 | — |
| DBH (m) | 12.4 ± 0.6 | 17.4 ± 0.1 | 9.8 ± 0.1 | — |
| Stand density (tree ha−1) | 1958 | 1328 | 1867 | — |
| Stand age (years) | 35 | 35 | 35 | — |
| MF | PF | QF | CK | |
|---|---|---|---|---|
| Num | 14.2 ± 4.5 a | 13.8 ± 2.6 a | 15.0 ± 5.0 a | 9.2 ± 1.7 b |
| H’plant | 1.60 ± 0.22 a | 1.50 ± 0.19 ab | 1.72 ± 0.42 a | 1.27 ± 0.25 b |
| Soil Properties | Unit | MF | PF | QF | CK |
|---|---|---|---|---|---|
| pH | 5.30 ± 0.22 b | 6.10 ± 0.45 a | 5.41 ± 0.11 b | 6.33 ± 0.14 a | |
| SWC | % | 13.10 ± 1.4 8b | 11.84 ± 0.52 b | 18.72 ± 1.78 a | 8.46 ± 2.17 c |
| TN | g kg−1 | 0.76 ± 0.11 a | 0.38 ± 0.05 b | 0.71 ± 0.05 a | 0.46 ± 0.05 b |
| TP | g kg−1 | 0.17 ± 0.03 c | 0.16 ± 0.01 c | 0.26 ± 0.02 b | 0.33 ± 0.01 a |
| TOC | g kg−1 | 4.58 ± 0.69 ab | 3.68 ± 1.08 b | 6.31 ± 1.05 a | 3.19 ± 0.82 b |
| TOC/TN | 6.09 ± 1.08 a | 9.71 ± 3.03 a | 8.93 ± 1.96 a | 7.06 ± 2.25 a | |
| NH4+-N | mg kg−1 | 1.32 ± 0.71 a | 0.85 ± 0.31 a | 0.80 ± 0.38 a | 0.77 ± 0.21 a |
| NO3--N | mg kg−1 | 1.02 ± 0.20 b | 1.98 ± 0.67 a | 1.08 ± 0.21 b | 0.58 ± 0.13 b |
| Wn | mg kg−1 | 47.33 ± 14.42 ab | 54.67 ± 17.33 a | 56.95 ± 11.95 a | 38.77 ± 3.61 b |
| Type of Carbon Source | Kind of Carbon Source | PC1 | PC2 |
|---|---|---|---|
| Amino acids | L-arginine | 0.614 | |
| L-asparagine | 0.816 | ||
| L-asparagine | 0.752 | ||
| L-serine | 0.724 | ||
| L-threonine | 0.707 | ||
| Glycyl-L-glutamic acid | 0.858 | ||
| Carbohydrates | D-cellobiose | ||
| α-D-lactose | 0.646 | −0.635 | |
| β-methyl-D-glucoside | |||
| D-xylose | |||
| i-erythritol | |||
| D-mannitol | 0.839 | ||
| α-D-glucose-1-phosphate | 0.727 | ||
| D,L-α-glycerol phosphate | |||
| N-acetyl-D-glucosamine | 0.809 | ||
| Carboxylic acids | D-glucosaminiczcid | 0.777 | |
| D-galactonic acid lactone | |||
| D-galacturonic acid | 0.770 | ||
| Pyruvic acid methyl ester | 0.646 | ||
| γ-hydroxybutyric acid | |||
| Itaconic acid | |||
| α-ketobutyric acid | |||
| D-malic acid | 0.653 | ||
| Amine | Phenylethylamine | 0.624 | |
| Putrescine | 0.805 | ||
| Polymers | 40 Tween 40 | 0.808 | |
| 80 Tween 80 | 0.702 | ||
| α-cyclodextrin | 0.747 | ||
| Glycogen | −0.745 | ||
| Phenolic acids | 2-hydroxybenzoic acid | 0.808 | |
| 4-hydroxybenzoic acid | 0.933 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yao, Z.; Zhang, X.; Wang, X.; Shu, Q.; Liu, X.; Wu, H.; Gao, S. Functional Diversity of Soil Microorganisms and Influencing Factors in Three Typical Water-Conservation Forests in Danjiangkou Reservoir Area. Forests 2023, 14, 67. https://0-doi-org.brum.beds.ac.uk/10.3390/f14010067
Yao Z, Zhang X, Wang X, Shu Q, Liu X, Wu H, Gao S. Functional Diversity of Soil Microorganisms and Influencing Factors in Three Typical Water-Conservation Forests in Danjiangkou Reservoir Area. Forests. 2023; 14(1):67. https://0-doi-org.brum.beds.ac.uk/10.3390/f14010067
Chicago/Turabian StyleYao, Zengwang, Xudong Zhang, Xu Wang, Qi Shu, Xinmiao Liu, Hailong Wu, and Shenghua Gao. 2023. "Functional Diversity of Soil Microorganisms and Influencing Factors in Three Typical Water-Conservation Forests in Danjiangkou Reservoir Area" Forests 14, no. 1: 67. https://0-doi-org.brum.beds.ac.uk/10.3390/f14010067