

Efficient Procedure for Induction Somatic Embryogenesis in Holm Oak: Roles of Explant Type, Auxin Type, and Exposure Duration to Auxin

Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Material
2.2. Somatic Embryogenesis Initiation
2.3. Statistical Analysis
3. Results
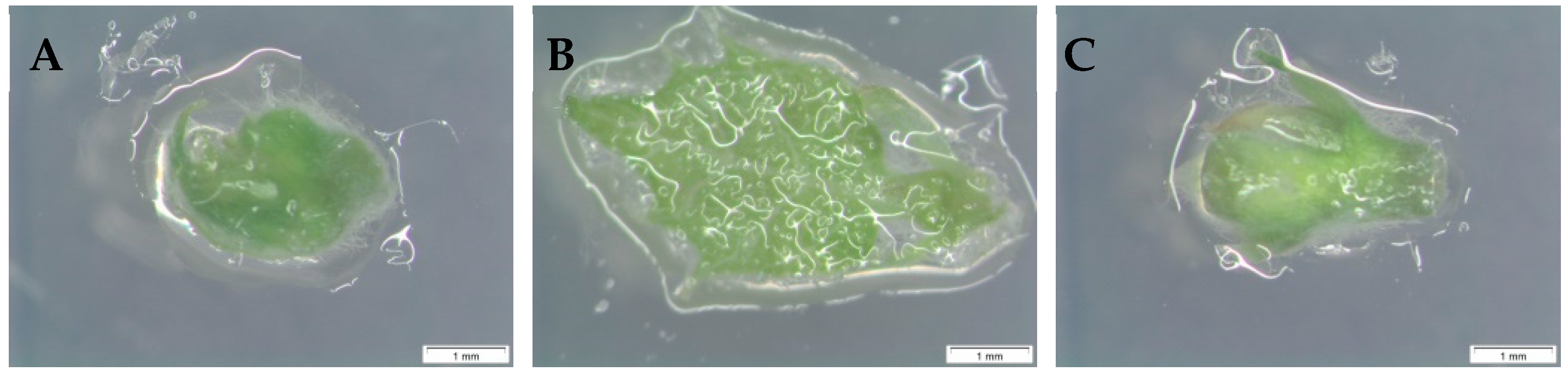
3.1. Embryogenic Response in Apex Explants
3.2. Embryogenic Response in Node Explants
3.3. Embryogenic Response in Leaf Explants
3.4. Overview of Results
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- De Rigo, D.; Cadullo, G. Quercus ilex in Europe: Distribution, habitat, usage and threats. In European Atlas of Forest Tree Species; Publication Office of the European Union: Luxembourg, 2016; pp. 152–153. [Google Scholar]
- Villar-Salvador, P.; Nicolás, J.L.; Heredia, N.; Uscola, M. Quercus ilex L. In Producción y Manejo de Semillas y Plantas Forestales. Tomo II. Organismo Autónomo Parques Nacionales; Pemán, J., Navarro-Cerrillo, R.M., Nicolás, J.L., Prada, M.A., Serrada, R., Eds.; Serie Forestal; Ministerio de Agricultura, Alimentación y Medio Ambiente: Madrid, Spain, 2013; pp. 226–250. [Google Scholar]
- Agricultura, Alimentación y Medio Ambiente en España. 2015. Available online: https://www.mapa.gob.es/es/ministerio/servicios/publicaciones/memoria_magrama_2015_completo_tcm30-83965.pdf (accessed on 12 January 2023).
- Oppermann, R.; Beaufoy, G.; Jones, G. High nature value farming in Europe. In 35 European Countries—Experiences and Perspectives; Verlag Regionalkultur: Ubstadt-Weiher, Germany, 2012; p. 544. [Google Scholar]
- Plieninger, T.; Hartel, T.; Martín-López, B.; Beaufoy, G.; Bergmeier, E.; Kirby, K.; Montero, M.J.; Moreno, G.; Oteros-Rozas, E.; Van Uytvanck, J. Wood-pastures of Europe: Geographic coverage, social-ecological values, conservation management, and policy implications. Biol. Conserv. 2015, 190, 70–79. [Google Scholar] [CrossRef]
- Roig, S.; San Miguel, A. ¿Cómo se mide el estado de conservación de la dehesa? Los pastos: Nuevos retos, nuevas oportunidades. In Proceedings of the Reunión Científica de la Sociedad Española para el Estudio de los Pastos, Badajoz, Spain, 8–11 April 2013. [Google Scholar]
- Plieninger, T.; Flinzberger, L.; Hetman, M.; Horstmannshoff, I.; Reinhard-Kolempas, M.; Topp, E.; Moreno, G.; Huntsinger, L. Dehesas as high nature value farming systems: A social-ecological synthesis of drivers, pressures, state, impacts, and responses. Ecol. Soc. 2021, 26, 23. [Google Scholar] [CrossRef]
- Martín-Delgado, L.-M.; Jiménez-Barrado, V.; Sánchez-Martín, J.-M. Sustainable Hunting as a Tourism Product in Dehesa Areas in Extremadura (Spain). Sustainability 2022, 14, 10288. [Google Scholar] [CrossRef]
- MITECO. Plan de Activación Socioeconómica del Sector Forestal. Available online: https://www.miteco.gob.es/es/biodiversidad/temas/politica-forestal/plan-pasfor/default.aspx (accessed on 12 January 2023).
- Corcobado, T.; Solla, A.; Madeira, M.A.; Moreno, G. Combined effects of soil properties and Phytophthora cinnamomi infections on Quercus ilex decline. Plant Soil 2013, 373, 403–413. [Google Scholar] [CrossRef]
- Camilo-Alves, C.S.P.; Clara, M.I.E.; Ribeiro, N.M.C.A. Decline of Mediterranean oak trees and its association with Phytophthora cinnamomi: A review. Eur. J. Forest Res. 2013, 132, 411–432. [Google Scholar] [CrossRef]
- Corredoira, E.; Merkle, S.A.; Martínez, M.T.; Toribio, M.; Canhoto, J.M.; Correia, S.I.; Ballester, A.; Vieitez, A.M. Non-Zygotic Embryogenesis in Hardwood Species. Crit. Rev. Plant Sci. 2019, 38, 29–97. [Google Scholar] [CrossRef]
- Corredoira, E.; Toribio, M.; Vieitez, A.M. Clonal propagation via somatic embryogenesis in Quercus spp. In Tree Biotechnology; Ramawhat, K.G., Mérillon, J.-M., Ahuja, M.R., Eds.; CRC Press: Boca Raton, FL, USA, 2014; pp. 262–302. [Google Scholar]
- Hernández, I.; Cuenca, B.; Carneros, E.; Alonso-Blázquez, N.; Ruiz, M.; Celestino, C.; Ocaña, L.; Alegre, J.; Toribio, M. Application of plant regeneration of selected cork oak trees by somatic embryogenesis to implement multivarietal forestry for cork production. Tree For. Sci. Biotech. 2011, 5, 19–26. [Google Scholar]
- Toribio, M.; Fernández, C.; Celestino, C.; Martínez, M.T.; San José, M.C.; Vieitez, A.M. Somatic embryogenesis in mature Quercus robur trees. Plant Cell Tissue Organ Cult. 2004, 76, 283–287. [Google Scholar] [CrossRef]
- Valladares, S.; Sánchez, C.; Martínez, M.T.; Ballester, A.; Vieitez, A.M. Plant regeneration through somatic embryogenesis from tissues of mature oak trees: True-to-type conformity of plantlets by RAPD analysis. Plant Cell Rep. 2006, 25, 879–886. [Google Scholar] [CrossRef]
- Corredoira, E.; San-José, M.C.; Vieitez, A.M. Induction of somatic embryogenesis from different explants of shoot cultures derived from young Quercus alba trees. Trees 2012, 26, 881–891. [Google Scholar] [CrossRef]
- Mauri, P.; Manzanera, J. Induction, maturation and germination of holm oak (Quercus ilex L.) somatic embryos. Plant Cell Tissue Organ Cult. 2003, 74, 229–235. [Google Scholar] [CrossRef]
- Mauri, P.V.; Manzanera, J.A. Protocol of somatic embryogenesis: Holm oak (Quercus ilex L.). In Protocol for Somatic Embryogenesis in Woody Plants; Jain, S.M., Gupta, P.K., Eds.; Springer: Dordrecht, The Netherlands, 2005; pp. 469–482. [Google Scholar]
- Blasco, M.; Barra, A.; Brisa, C.; Corredoira, E.; Segura, J.; Toribio, M.; Arrillaga, I. Somatic embryogenesis in holm oak male catkins. Plant Growth Regul. 2013, 71, 261–270. [Google Scholar] [CrossRef]
- Barra-Jiménez, A.; Blasco, M.; Ruiz-Galea, M.; Celestino, C.; Alegre, J.; Arrillaga, I.; Toribio, M. Cloning mature holm oak trees by somatic embryogenesis. Trees 2014, 28, 657–667. [Google Scholar] [CrossRef]
- Martínez, M.T.; José, M.C.S.; Vieitez, A.M.; Cernadas, M.J.; Ballester, A.; Corredoira, E. Propagation of mature Quercus ilex L. (holm oak) trees by somatic embryogenesis. Plant Cell Tissue Organ Cult. 2017, 131, 321–333. [Google Scholar] [CrossRef]
- Martínez, M.T.; Vieitez, F.J.; Solla, A.; Tapias, R.; Ramírez-Martín, N.; Corredoira, E. Vegetative Propagation of Phytophthora cinnamomi-Tolerant Holm Oak Genotypes by Axillary Budding and Somatic Embryogenesis. Forests 2020, 11, 841. [Google Scholar] [CrossRef]
- Martínez, M.T.; Arrillaga, I.; Sales, E.; Pérez-Oliver, M.A.; González-Mas, M.d.C.; Corredoira, E. Micropropagation, Characterization, and Conservation of Phytophthora cinnamomi-Tolerant Holm Oak Mature Trees. Forests 2021, 12, 1634. [Google Scholar] [CrossRef]
- Martínez, M.T.; San-José, M.C.; Arrillaga, I.; Cano, V.; Morcillo, M.; Cernadas, M.J.; Corredoira, E. Holm oak somatic embryogenesis: Current status and future perspectives. Front. Plant Sci. 2019, 10, 239. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gaj, M. Factors influencing somatic embryogenesis induction and plant regeneration with particular reference to Arabidopsis thaliana (L.) Heynh. Plant Growth Regul. 2004, 43, 27–47. [Google Scholar] [CrossRef]
- Fehér, A. Somatic embryogenesis—Stress-induced remodeling of plant cell fate. Biochim. Biophys. Acta Bioenerg. 2015, 1849, 385–402. [Google Scholar] [CrossRef]
- Fehér, A. The initiation phase of somatic embryogenesis: What we know and what we don’t. Acta Biol. Szeged. 2008, 52, 53–56. [Google Scholar]
- Su, Y.H.; Tang, L.P.; Zhao, X.Y.; Zhang, X.S. Plant cell totipotency: Insights into cellular reprogramming. J. Integr. Plant Biol. 2021, 63, 228–240. [Google Scholar] [CrossRef]
- Guan, Y.; Li, S.-G.; Fan, X.-F.; Su, Z.-H. Application of somatic embryogenesis in woody plants. Front. Plant Sci. 2016, 7, 938. [Google Scholar] [CrossRef] [Green Version]
- Wójcik, A.M.; Wójcikowska, B.; Gaj, M.D. Current Perspectives on the Auxin-Mediated Genetic Network that Controls the Induction of Somatic Embryogenesis in Plants. Int. J. Mol. Sci. 2020, 21, 1333. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nic-Can, G.I.; Loyola-Vargas, V.M. The Role of the Auxins During Somatic Embryogenesis. In Somatic Embryogenesis: Fundamental Aspects and Applications; Loyola-Vargas, V., Ochoa-Alejo, N., Eds.; Springer: Cham, Switzerland, 2016; pp. 171–182. [Google Scholar] [CrossRef]
- Elhiti, M.; Stasolla, C. Transduction of Signals during Somatic Embryogenesis. Plants 2022, 11, 178. [Google Scholar] [CrossRef]
- George, E.F.; Hall, M.A.; Klerk, G.J.D. Plant Growth Regulators I: Introduction; Auxins, their Analogues and Inhibitors. In Plant Propagation by Tissue Culture; George, E.F., Hall, M.A., Klerk, G.J.D., Eds.; Springer: Dordrecht, The Netherlands, 2008; pp. 175–204. [Google Scholar] [CrossRef]
- Vieitez, A.M.; Corredoira, E.; Martínez, M.T.; José, M.C.S.; Sánchez, C.; Valladares, S.; Vidal, N.; Ballester, A. Application of biotechnological tools to Quercus improvement. Eur. J. For. Res. 2012, 131, 519–539. [Google Scholar] [CrossRef] [Green Version]
- Merkle, S.A. Strategies for dealing with limitations of somatic embryogenesis in hardwood trees. Plant Tissue Cult. Biotech. 1995, 1, 112–121. [Google Scholar]
- Ballester, A.; Corredoira, E.; Vieitez, A.M. Limitations of somatic embryogenesis in hardwood trees. In Vegetative Propagation of Forest Trees; Park, Y.S., Bonga, J.M., Moon, H.K., Eds.; National Institute of Forest Science (NiFos): Seoul, Republic of Korea, 2016; pp. 56–74. [Google Scholar]
- Corredoira, E.; Ballester, A.; Ibarra, M.; Vieitez, A.M. Induction of somatic embryogenesis in leaf and shoot apex explants of shoot cultures derived from adult Eucalyptus globulus and Eucalyptus saligna × E. maidenii trees. Tree Physiol. 2015, 35, 663–667. [Google Scholar] [CrossRef]
- Maillot, P.; Deglène-Benbrahim, L.; Walter, B. Efficient somatic embryogenesis from meristematic explants in grapevine (Vitis vinifera L.) cv. Chardonnay: An improved protocol. Trees 2016, 30, 1377–1387. [Google Scholar] [CrossRef]
- Martínez, M.T.; Corredoira, E.; Vieitez, A.M.; Cernadas, M.J.; Montenegro, R.; Ballester, A.; Vieitez, F.J.; José, M.C.S. Micropropagation of mature Quercus ilex L. trees by axillary budding. Plant Cell Tissue Organ Cult. 2017, 131, 499–512. [Google Scholar] [CrossRef]
- Lloyd, G.; McCown, B. Commercially-feasible micropropagation of mountain laurel, Kalmia latifolia, by use of shoot-tip culture. Comb. Proc. Int. Plant Prop. Soc. 1980, 30, 421–427. [Google Scholar]
- Murashige, T.; Skoog, F. A revised medium for rapid growth and bioassays with tobacco tissue culture. Physiol. Plant 1962, 15, 473–497. [Google Scholar] [CrossRef]
- Bonga, J.M. Vegetative propagation in relation to juvenility, maturity and rejuvenation. In Tissue Culture in Forestry; Bonga, J.M., Durzan, D.J., Eds.; Martinus Nijhoff: Dordrecht, The Netherlands; Dr. W. Junk Publishers: Dordrecht, The Netherlands, 1982; pp. 387–412. [Google Scholar]
- Corredoira, E.; Martínez, M.T.; Cernadas, M.J.; San José, M.C. Application of Biotechnology in the Conservation of the Genus Castanea. Forests 2017, 8, 394. [Google Scholar] [CrossRef] [Green Version]
- Capriotti, L.; Limera, C.; Mezzetti, B.; Ricci, A.; Sabbadini, S. From induction to embryo proliferation: Improved somatic embryogenesis protocol in grapevine for Italian cultivars and hybrid Vitis rootstocks. Plant Cell Tissue Organ Cult. 2022, 151, 221–233. [Google Scholar] [CrossRef]
- Martins, J.; Correia, S.; Pinto, G.; Canhoto, J. Cloning adult trees of Arbutus unedo L. through somatic embryogenesis. Plant Cell Tissue Organ Cult. 2022, 150, 611–626. [Google Scholar] [CrossRef]
- Wendling, I.; Trueman, S.J.; Xavier, A. Maturation and related aspects in clonal forestry—Part II: Reinvigoration, rejuvenation and juvenility maintenance. New For. 2014, 45, 473–486. [Google Scholar] [CrossRef]
- Martínez, T.; Vidal, N.; Ballester, A.; Vieitez, A.M. Improved organogenic capacity of shoot cultures from mature pedunculate oak trees through somatic embryogenesis as rejuvenation technique. Trees 2012, 26, 321–330. [Google Scholar] [CrossRef]
- Capelo, A.M.; Silva, S.; Brito, G.; Santos, C. Somatic embryogenesis induction in leaves and petioles of a mature wild olive. Plant Cell Tissue Organ Cult. 2010, 103, 237–242. [Google Scholar] [CrossRef]
- Pedroso, M.C.; Pais, M.S. Direct embryo formation in leaves of Camellia japonica L. Plant Cell Rep. 1993, 12, 639–643. [Google Scholar] [CrossRef] [PubMed]
- Akula, A.; Dodd, W.A. Direct somatic embryogenesis in a selected tea clone, TRI-2025’(Camellia sinensis (L.) O. Kuntze) from nodal explants. Plant Cell Rep. 1998, 17, 804–809. [Google Scholar] [CrossRef]
- Geneye, R.L.; Kester, S.T. The initiation of somatic embryos and adventitious roots from developing zygotic embryo explants of Cercis canadensis L. cultured in vitro. Plant Cell Tissue Organ Cult. 1990, 22, 71–76. [Google Scholar] [CrossRef]
- Lad, B.L.; Jayasankar, S.; Pliego-Alfaro, F.; Moon, P.A.; Litz, R.E. Temporal effect of 2,4-D on induction of embryogenic nucellar cultures and somatic embryo development of ‘Carabao’ Mango. Cell. Dev. Biol. Plant 1997, 33, 253–257. [Google Scholar] [CrossRef]
- Iwase, A.; Mitsuda, N.; Koyama, T.; Hiratsu, K.; Kojima, M.; Arai, T.; Inoue, Y.; Seki, M.; Sakakibara, H.; Sugimoto, K.; et al. The AP2/ERF transcription factor WIND1 controls cell dedifferentiation in Arabidopsis. Curr. Biol. 2011, 21, 508–514. [Google Scholar] [CrossRef] [Green Version]
- Grzyb, M.; Kalandyk, A.; Waligórski, P.; Mikula, A. The content of endogenous hormones and sugars in the process of early somatic embryogenesis in the tree fern Cyathea delgadii Sternb. Plant Cell Tissue Organ Cult. 2017, 129, 387–397. [Google Scholar] [CrossRef] [Green Version]
- Mallón, R.; Martínez, M.T.; Corredoira, E.; Vieitez, A.M. The positive effect of arabinogalactan on induction of somatic embryogenesis in Quercus bicolor followed by embryo maturation and plant regeneration. Trees 2013, 27, 1285–1296. [Google Scholar] [CrossRef]
- San José, M.C.; Corredoira, E.; Martínez, M.T.; Vidal, N.; Valladares, S.; Mallón, R.; Vieitez, A.M. Shoot apex explants for induction of somatic embryogenesis in mature Quercus robur L. trees. Plant Cell Rep. 2010, 29, 661–671. [Google Scholar] [CrossRef] [PubMed]
- Zavattieri, M.A.; Frederico, A.M.; Lima, M.; Sabino, R.; Arnholdt-Schmitt, B. Induction of somatic embryogenesis as an example of stress-related plant reactions. Electron. J. Biotechnol. 2010, 13, 12–13. [Google Scholar] [CrossRef] [Green Version]
- Narváez, I.; Martín, C.; Jiménez-Díaz, R.M.; Mercado, J.A.; Pliego-Alfaro, F. Plant Regeneration via Somatic Embryogenesis in Mature Wild Olive Genotypes Resistant to the Defoliating Pathotype of Verticillium dahliae. Front. Plant Sci. 2019, 10, 1471. [Google Scholar] [CrossRef]
- Mazri, M.A.; Belkoura, I.; Meziani, R.; Mokhless, B.; Nour, S. Somatic embryogenesis from bud and leaf explants of date palm (Phoenix dactylifera L.) cv. Najda. 3 Biotech. 2017, 7, 58. [Google Scholar] [CrossRef] [Green Version]
- Rugkhla, A.; Jones, M.G.K. Somatic embryogenesis and plantlet formation in Santalum album and S. spicatum. J. Exp. Bot. 1998, 49, 563–571. [Google Scholar] [CrossRef]
- Bonga, J.M. Can explant choice help resolve recalcitrance problems in in vitro propagation, a problem still acute especially for adult conifers? Trees 2017, 31, 781–789. [Google Scholar] [CrossRef]
- Hernández, I.; Celestino, C.; Toribio, M. Vegetative propagation of Quercus suber L. by somatic embryogenesis: I. Factors affecting the induction in leaves from mature cork oak trees. Plant Cell Rep. 2003, 21, 759–764. [Google Scholar] [CrossRef] [PubMed]
- Loyola-Vargas, V.M.; Avilez-Montalvo, J.R.; Avilés-Montalvo, R.N.; Márquez-López, R.E.; Galaz-Ávalos, R.M.; Mellado-Mojica, E. Somatic Embryogenesis in Coffea spp. In Somatic Embryogenesis: Fundamental Aspects and Applications; Loyola-Vargas, V., Ochoa-Alejo, N., Eds.; Springer: Cham, Switzerland, 2016; pp. 241–266. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Treatment (mg/L) | Somatic Embryogenesis (%) | ||
|---|---|---|---|
| Q10-SE | Q3-SE | E00 | |
| IAA 4 + BA 0.5 | |||
| 2 wks | 15.0 ± 2.9 | 2.0 ± 1.9 | 12.0 ± 3.1 |
| 4 wks | 11.0 ± 3.6 | 7.0 ± 2.5 | 2.0 ± 1.3 |
| 8 wks | 12.0 ± 2.8 | 15.0 ± 5.0 | 1.0 ± 0.9 |
| NAA 4 + BA 0.5 | |||
| 2 wks | 3.0 ± 2.9 | 33.0 ± 4.3 | 0.0 ± 0.0 |
| 4 wks | 0.0 ± 0.0 | 20.0 ± 7.2 | 0.0 ± 0.0 |
| 8 wks | 0.0 ± 0.0 | 22.0 ± 4.2 | 0.0 ± 0.0 |
| IBA 3 + NAA 0.1 | |||
| 2 wks | 0.0 ± 0.0 | 12.0 ± 4.2 | 0.0 ± 0.0 |
| 8 wks | 0.0 ± 0.0 | 14.0 ± 2.9 | 0.0 ± 0.0 |
| ANOVA | |||
| Genotype (A) | p = 0.001 *** | ||
| Treatment (B) | p = 0.001 *** | ||
| Exposure duration (C) | p = 0.049 * | ||
| A × B | p = 0.001 *** | ||
| A × C | 0.111 ns | ||
| B × C | 0.495 ns | ||
| A × B × C | p = 0.001 *** | ||
| Treatment (mg/L) | Somatic Embryogenesis (%) | ||
|---|---|---|---|
| Q10-SE | Q3-SE | E00 | |
| IAA 4 + BA 0.5 | |||
| 2 wks | 37.0 ± 5.3 | 2.0 ± 1.9 | 4.0 ± 2.1 |
| 4 wks | 21.0 ± 5.6 | 2.0 ± 1.3 | 4.0 ± 2.1 |
| 8 wks | 3.0 ± 2.0 | 4.0 ± 1.6 | 3.0 ± 2.0 |
| NAA 4 + BA 0.5 | |||
| 2 wks | 0.0 ± 0.0 | 29.0 ± 5.0 | 1.0 ± 0.9 |
| 4 wks | 0.0 ± 0.0 | 4.0 ± 2.1 | 0.0 ± 0.0 |
| 8 wks | 0.0 ± 0.0 | 3.0 ± 1.5 | 0.0 ± 0.0 |
| IBA 3 + NAA 0.1 | |||
| 2 wks | 4.0 ± 2.9 | 24.0 ± 3.2 | 1.0 ± 0.0 |
| 8 wks | 0.0 ± 0.0 | 1.0 ± 0.95 | 0.0 ± 0.0 |
| ANOVA | |||
| Genotype (A) | p = 0.001 *** | ||
| Treatment (B) | p = 0.001 *** | ||
| Exposure duration (C) | p = 0.001 *** | ||
| A × B | p = 0.001 *** | ||
| A × C | p = 0.001 *** | ||
| B × C | 0.262 ns | ||
| A × B × C | p = 0.001 *** | ||
| Treatment (mg/L) | Somatic Embryogenesis (%) | ||
|---|---|---|---|
| Q10-SE | Q3-SE | E00 | |
| IAA 4 + BA 0.5 | |||
| 2 wks | 1.0 ± 0.9 | 0.0 ± 0.0 | 0.0 ± 0.0 |
| 8 wks | 0.0 ± 0.0 | 2.0 ± 1.3 | 0.0 ± 0.0 |
| NAA 4 + BA 0.5 | |||
| 2 wks | 0.0 ± 0.0 | 9.0 ± 3.0 | 0.0 ± 0.0 |
| 8 wks | 0.0 ± 0.0 | 1.0 ± 0.9 | 1.0 ± 0.95 |
| IBA 3 + NAA 0.1 | |||
| 2 wks | 0.0 ± 0.0 | 0.0 ± 0.0 | 0.0 ± 0.0 |
| 8 wks | 1.0 ± 0.9 | 1.0 ± 0.95 | 1.0 ± 0.9 |
| ANOVA | |||
| Genotype (A) | p = 0.003 ** | ||
| Treatment (B) | p = 0.019 * | ||
| Exposure duration (C) | 0.506 ns | ||
| A × B | p = 0.001 *** | ||
| A × C | 0.071 ns | ||
| B × C | p = 0.019 * | ||
| A × B × C | p = 0.001 *** | ||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Martínez, M.T.; Corredoira, E. Efficient Procedure for Induction Somatic Embryogenesis in Holm Oak: Roles of Explant Type, Auxin Type, and Exposure Duration to Auxin. Forests 2023, 14, 430. https://0-doi-org.brum.beds.ac.uk/10.3390/f14020430
Martínez MT, Corredoira E. Efficient Procedure for Induction Somatic Embryogenesis in Holm Oak: Roles of Explant Type, Auxin Type, and Exposure Duration to Auxin. Forests. 2023; 14(2):430. https://0-doi-org.brum.beds.ac.uk/10.3390/f14020430
Chicago/Turabian StyleMartínez, María Teresa, and Elena Corredoira. 2023. "Efficient Procedure for Induction Somatic Embryogenesis in Holm Oak: Roles of Explant Type, Auxin Type, and Exposure Duration to Auxin" Forests 14, no. 2: 430. https://0-doi-org.brum.beds.ac.uk/10.3390/f14020430