
Differences in the Responses of Tree-Ring Stable Carbon Isotopes of L. sibirica and P. schrenkiana to Climate in the Eastern Tianshan Mountains
 ,
,
Abstract
:1. Introduction
2. Data and Methods
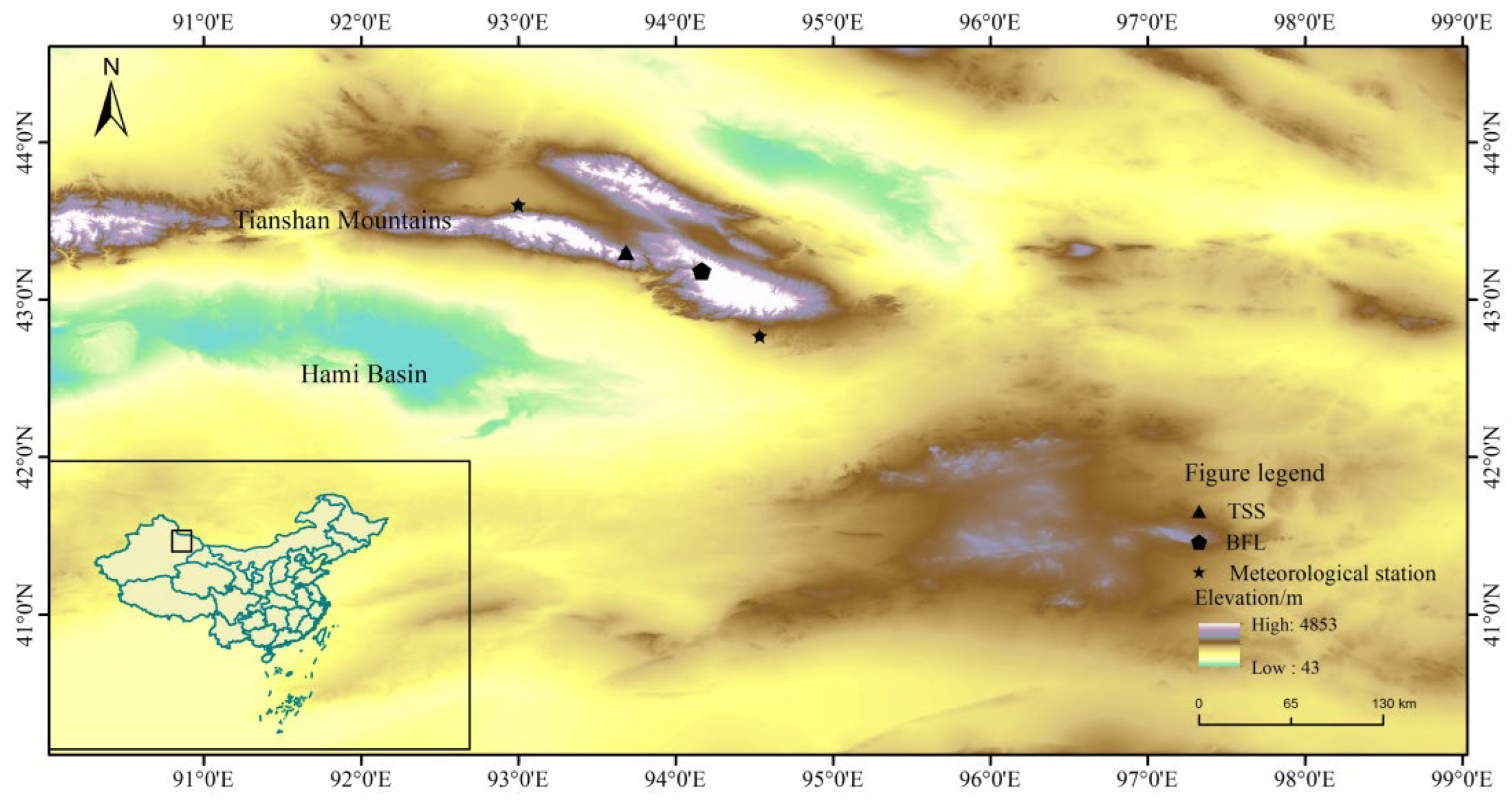
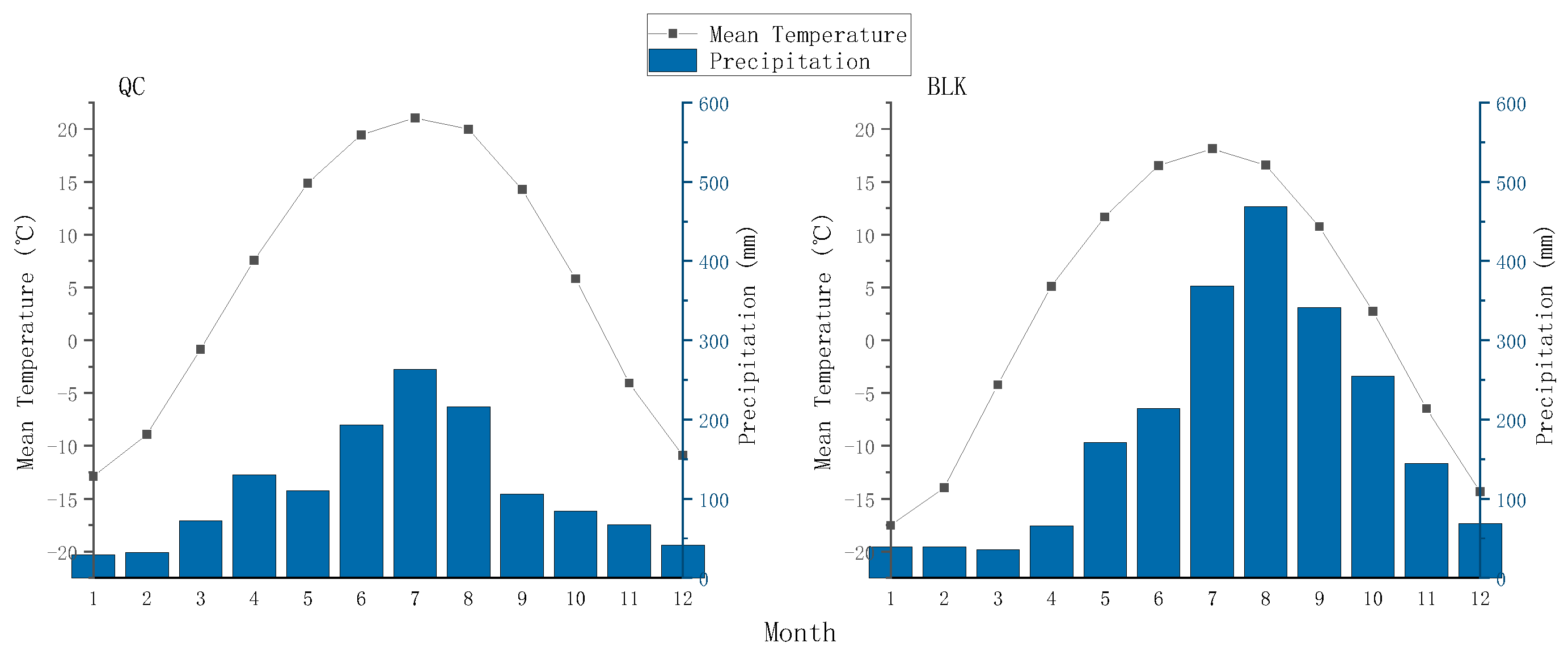
2.1. Overview of the Study Area and Sample Collection
2.2. Stable Carbon Isotope Chronologies Development
2.3. Meteorological Data
2.4. Analysis Methods
3. Results
3.1. Statistical Analysis of the Tree-Ring Stable Carbon Isotope Chronologies
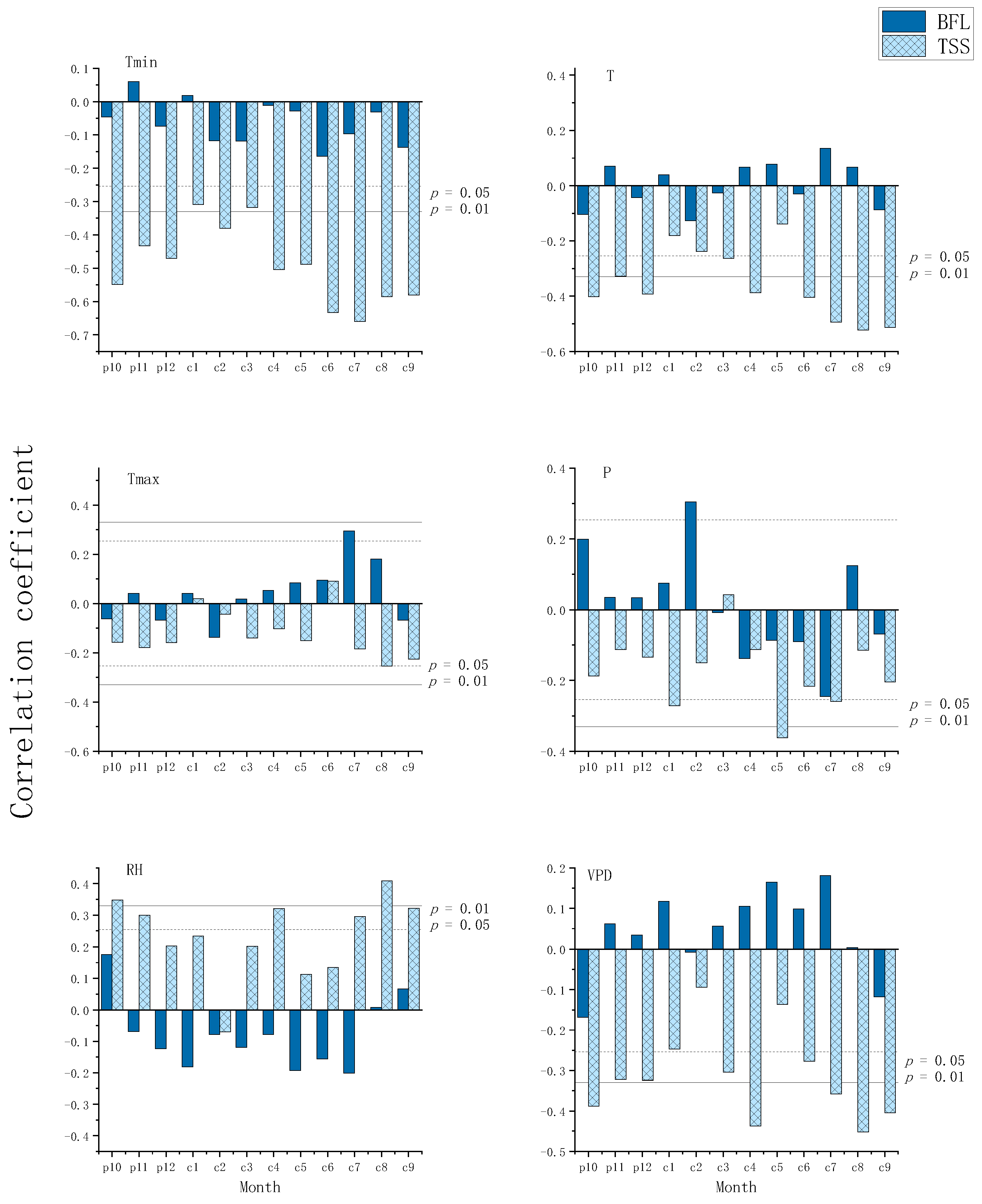
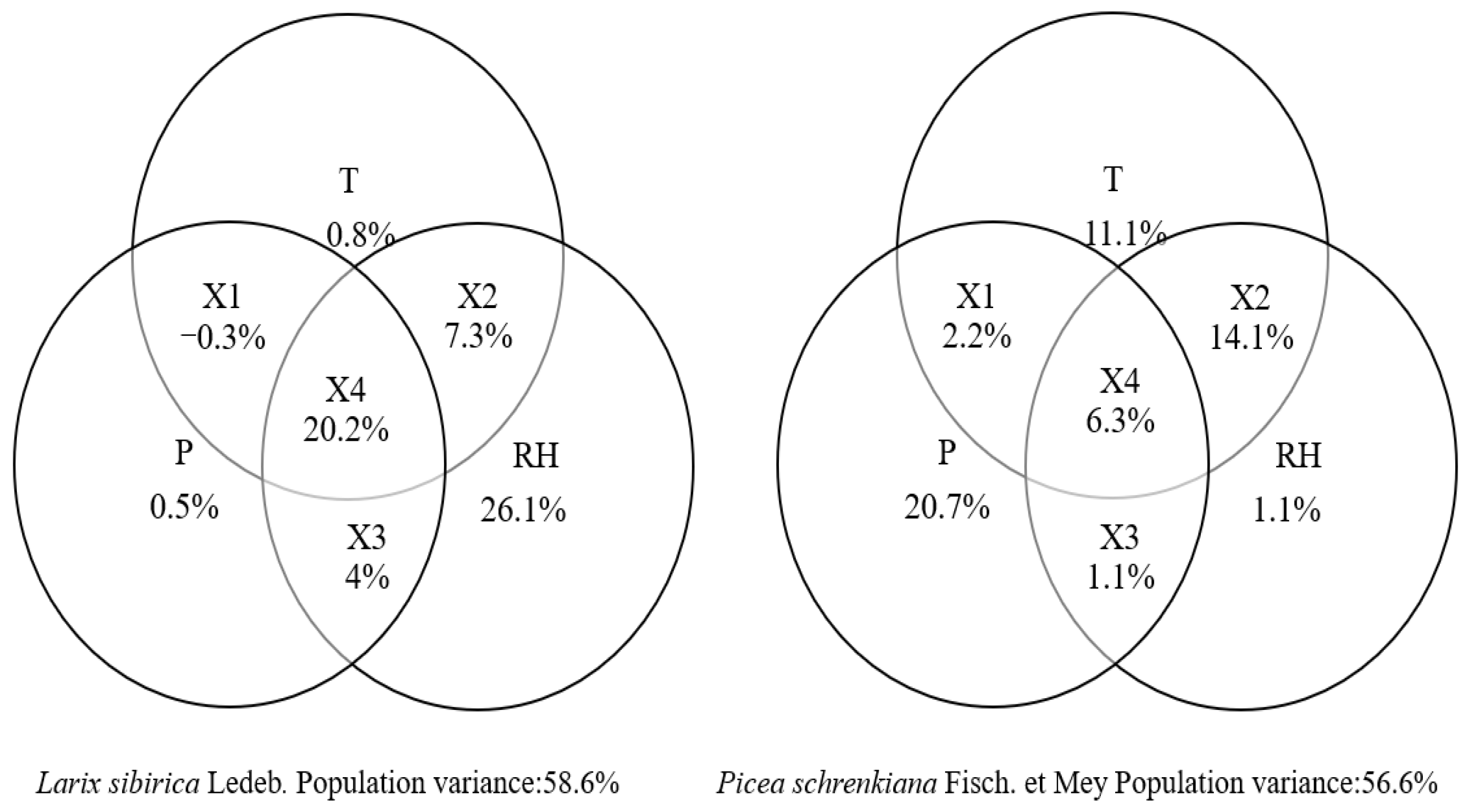
3.2. Response of Tree-Ring Stable Carbon Isotopes to Climate
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Robertson, I.; Switsur, V.R.; Carter, A.H.C.; Barker, A.C.; Waterhouse, J.S.; Briffa, K.R.; Jones, P.D. Signal strength and climate relationships in 13C/12C ratios of tree ring cellulose from oak in east England. J. Geophys. Res. Atmos. 1997, 102, 19507–19516. [Google Scholar] [CrossRef]
- Leavitt, S.W.; Lone, A. Seasonal stable-carbon isotope variability in tree-ring: Possible paleoenvironmental signals. Chem. Geol. Isot. Geosci. Sect. 1991, 87, 59–70. [Google Scholar] [CrossRef]
- Wilson, A.T.; Grinsted, M.J. 13C/12C in cellulose and lignin as palaeothermometers. Nature 1977, 265, 133–135. [Google Scholar] [CrossRef]
- Francey, R.J.; Farquhar, G.D. An explanation of 13C/12C variations in tree-rings. Nature 1982, 297, 28–31. [Google Scholar] [CrossRef]
- Saurer, M.; Siegenthaler, U.; Schweingruber, F. The climate-carbon isotope relationship in tree-ring and the significance of site conditions. Tellus B 1995, 47, 320–330. [Google Scholar] [CrossRef]
- Hemming, D.I.; Switsur, V.R.; Waterhouse, J.S.; Heaton, T.H.E.; Carter, A.H.C. Climate variation and the stable carbon isotope composition of tree ring Cellulose: An intercomparison of Quercus robur, Fagus sylvatica and Pinus silvestris. Tellus B Chem. Phys. Meteorol. 1998, 50, 25–33. [Google Scholar] [CrossRef]
- Saurer, M.; Siegenthaler, U. 13C/12C isotope ratios in trees are sensitive to relative humidity. Dendrochronologia 1989, 7, 9–13. [Google Scholar]
- Barber, V.A.; Juday, G.P.; Finney, B.P.; Wilmking, M. Reconstruction of Summer Temperatures in Interior Alaska from Tree-Ring Proxies: Evidence for Changing Synoptic Climate Regimes. Clim. Chang. 2004, 63, 91–120. [Google Scholar] [CrossRef]
- Hafner, P.; Levanič, T. Stable carbon isotopes in Norway Spruce (Picea abies (L.) Karst.) tree-ring at two sites in Slovenia. Zb. Gozdarstva Lesar. 2009, 88, 43–52. [Google Scholar]
- Pawelczyk, S.; Pazdur, A.; Halas, S. Stable carbon isotopic composition of tree-ring from a Pine tree from Augustów Wilderness, Poland, as a temperature and local environment conditions indicator. Isot. Environ. Health Stud. 2004, 40, 145–154. [Google Scholar] [CrossRef]
- Kirdyanov, A.V.; Treydte, K.S.; Nikolaev, A.; Helle, G.; Schleser, G.H. Climate signals in tree-ring width, density and δ13C from larches in eastern Siberia (Russia). Chem. Geol. 2008, 252, 31–41. [Google Scholar] [CrossRef]
- Sonninen, E.; Jungner, H. Stable carbon isotope in tree-rings of a Scots alpine pine (Pinus sylvestris L.) from northern Finland. Paläoklimaforschung 1995, 15, 121–128. [Google Scholar]
- Sheu, D.D.; Kou, P.; Chiu, C.H.; Chen, M.J. Variability of tree-ring δ13C in Taiwan fir: Growth effect and response to May–October temperatures. Geochim. Cosmochim. Acta 1996, 60, 171–177. [Google Scholar] [CrossRef]
- Saurer, M.; Borella, S.; Schweingruber, F.; Siegwolf, R. Stable carbon isotopes in tree-ring of beech: Climatic versus site-related influences. Trees 1997, 11, 291–297. [Google Scholar] [CrossRef]
- Liu, Y.; Wu, X.D.; Leavitt, S.W.; Hughes, M.K. Stable carbon isotope in tree-ring from Huangling, China and climatic Variation. Sci. China Ser. D Earth Sci. 1996, 39, 152–160. [Google Scholar]
- Liu, X.H.; Shao, X.M.; Wang, L.L.; Zhao, L.J.; Wu, P.; Chen, T.; Qin, D.H.; Ren, J.W. Climatic significance of the stable carbon isotope composition of tree-ring cellulose: Comparison of Chinese hemlock (Tsuga chinensis Pritz) and alpine pine (Pinus densata Mast) in a temperate-moist region of China. Sci. China Ser. D Earth Sci. 2007, 50, 1076–1085. [Google Scholar] [CrossRef]
- Wang, W.Z.; Liu, X.H.; Shao, X.M.; Leavitt, S.; Xu, G.B.; An, W.L.; Qin, D.H. A 200 years temperature record from tree ring δ13C at the Qaidam Basin of the Tibetan Plateau after identifying the optimum method to correct for changing atmospheric CO2 and δ13C. J. Geophys. Res. 2011, 116, G04022. [Google Scholar] [CrossRef]
- Liu, X.H.; Qin, D.H.; Shao, X.M.; Chen, T.; Ren, J.W. Tibetan Himalayan fir annual rings δ13C and climatic significance. Sci. Bull. 2003, 48, 1209–1213, (In Chinese, with English Abstract). [Google Scholar]
- Liu, X.H.; Qin, D.H.; Shao, X.M.; Ren, J.W.; Wang, Y. Stable Carbon Isotope of Abies spectabibis from Nyingchi County of Tibet Autonomous Region and Its Response to Climate Change. J. Glaciol. Geocryol. 2002, 24, 574–578, (In Chinese, with English Abstract). [Google Scholar]
- Liu, Y.; Wu, X.D.; Leavitt, S.W.; Hughes, M.K. Huangling tree rings stable C isotope and climate change. Chin. Sci. 1996, 26, 125–130, (In Chinese, with English Abstract). [Google Scholar]
- Zhang, R.; Ermenbaev, B.; Zhang, H.; Shang, H.; Zhang, T.; Yu, S.; Chontoev, D.T.; Satylkanov, R.; Qin, L. Natural discharge changes of the Naryn River over the past 265 years and their climatic drivers. Clim. Dyn. 2020, 55, 1269–1281. [Google Scholar] [CrossRef]
- Zhang, R.; Ermenbaev, B.; Zhang, T.; Ali, M.; Qin, L.; Satylkanov, R. The Radial Growth of Schrenk Spruce (Picea schrenkiana Fisch. et Mey.) Records the Hydroclimatic Changes in the Chu River Basin over the Past 175 Years. Forests 2019, 10, 223. [Google Scholar] [CrossRef]
- Zhang, R.; Shang, H.; Yu, S.; He, Q.; Yuan, Y.; Bolatov, K.; Mambetov, B.T. Tree-ring-based precipitation reconstruction in southern Kazakhstan, reveals drought variability since A.D. 1770. Int. J. Climatol. 2017, 37, 741–750. [Google Scholar] [CrossRef]
- Zhang, R.; Yuan, Y.; Gou, X.; He, Q.; Shang, H.; Zhang, T.; Chen, F.; Ermenbaev, B.; Yu, S.; Qin, L.; et al. Tree-ring-based moisture variability in western Tianshan Mountains since A.D. 1882 and its possible driving mechanism. Agric. For. Meteorol. 2016, 218–219, 267–276. [Google Scholar] [CrossRef]
- Zhang, R.; Qin, L.; Shang, H.; Yu, S.; Gou, X.; Mambetov, B.T.; Bolatov, K.; Zheng, W.; Ainur, U.; Bolatova, A. Climatic change in southern Kazakhstan since 1850 C.E. inferred from tree rings. Int. J. Biometeorol. 2020, 64, 841–851. [Google Scholar] [CrossRef] [PubMed]
- Zhang, R.; Wei, W.; Shang, H.; Yu, S.; Gou, X.; Qin, L.; Bolatov, K.; Mambetov, B.T. A tree ring-based record of annual mass balance changes for the TS.Tuyuksuyskiy Glacier and its linkages to climate change in the Tianshan Mountains. Quat. Sci. Rev. 2019, 205, 10–21. [Google Scholar] [CrossRef]
- Qin, L.; Shang, H.M.; Zhang, T.W.; Liu, W.P.; Zhang, R.B. Response comparison of the tree-ring δ13C to climate on the southern and northern slopes of Tianshan Mountains. Acta Ecol. Sin. 2021, 41, 5713–5724, (In Chinese, with English Abstract). [Google Scholar]
- Qin, L.; Yuan, Y.J.; Yu, S.L.; Shang, H.M.; Chen, F.; Fan, Z.A.; Zhang, T.W.; Tong, Z. Tree-ring based monthly mean relative humidity reconstruction for the Eastern Tianshan Mountains during the last 443 years. J. Earth Environ. 2012, 3, 908–914, (In Chinese, with English Abstract). [Google Scholar]
- Shang, H.M.; Wei, W.S.; Yuan, Y.J.; Yu, S.L.; Zhang, R.B.; Chen, X.J. Environmental Response of Tree-ring Width and Stable Carbon Isotope at North Slope of Eastern Tianshan Mountains. Desert Oasis Meteorol. 2010, 4, 6–10, (In Chinese, with English Abstract). [Google Scholar]
- Zhang, R.B.; Yuan, Y.J.; Wei, W.S.; Shang, H.M.; Yu, S.L.; Zhang, T.W.; Chen, F.; Fan, Z.A.; Qin, L. Response of Stable Carbon Isotope of Larix sibirica Ledeb. Tree-rings to Climate Change. Arid Zone Res. 2012, 29, 328–334, (In Chinese, with English Abstract). [Google Scholar]
- Peng, Z.B.; Li, X.J.; Zhang, R.B.; Qin, L.; Zhang, H.L.; Chen, Y.P.; Liu, R. The response of tree-ring chronologies of Schrenk spruce (Picea schrenkiana Fisch. et Mey.) to climate change at high- and low- elevations of the Eastern Tianshan Mountains, Xinjiang, using different detrending methods. Acta Ecol. Sin. 2019, 39, 1595–1604, (In Chinese, with English Abstract). [Google Scholar]
- Holmes, R.L. Computer-assisted quality control in tree-ring dating and measurement. Tree-Ring Bull. 1983, 43, 69–78. [Google Scholar]
- Cook, E.R. A Time Series Analysis Approach to Tree Ring Standardization; The University of Arizona: Tucson, AZ, USA, 1985. [Google Scholar]
- Porté, A.; Loustau, D. Seasonal and interannual variations in carbon isotope discrimination in a maritime pine (Pinus pinaster) stand assessed from the isotopic composition of cellulose in annual rings. Tree Physiol. 2001, 21, 861–868. [Google Scholar] [CrossRef]
- Gagen, M.; McCarroll, D.; Loader, N.; Robertson, I.; Jalkanen, R.; Anchukaitis, K. Exorcising the ‘segment length curse’: Summer temperature reconstruction since AD 1640 using non-detrended stable carbon isotope ratios from pine trees in northern Finland. Holocene 2007, 17, 435–446. [Google Scholar] [CrossRef]
- Brendel, O.; Iannetta, P.; Stewart, D. A rapid and simple method to isolate pure alpha-cellulose. Phytochem. Anal. 2000, 11, 7–10. [Google Scholar] [CrossRef]
- Crampton, E.W.; Maynard, L.A. The Relation of Cellulose and Lignin Content to the Nutritive Value of Animal Feeds. J. Nutr. 1938, 15, 383–395. [Google Scholar] [CrossRef]
- Evans, M.N.; Reichert, B.K.; Kaplan, A.; Anchukaitis, K.J.; Vaganov, E.A.; Hughes, M.K.; Cane, M.A. A forward modeling approach to paleoclimatic interpretation of tree-ring data. J. Geophys. Res. 2006, 111, G03008. [Google Scholar] [CrossRef]
- Coplen, T.B. Discontinuance of SMOW and PDB. Nature 1995, 375, 285. [Google Scholar] [CrossRef]
- Leavitt, S.W.; Lara, A. South American tree-ring show declining μ13C trend. Tellus B Chem. Phys. Meteorol. 1994, 46, 152–157. [Google Scholar] [CrossRef]
- Rubino, M.; Etheridge, D.M.; Thornton, D.P.; Howden, R.; Allison, C.E.; Francey, R.J.; Langenfelds, R.L.; Steele, L.P.; Trudinger, C.M.; Spencer, D.A.; et al. Revised records of atmospheric trace gases CO2, CH4, N2O, and δ13C-CO2 over the last 2000 years from Law Dome, Antarctica. Earth Syst. Sci. Data 2019, 11, 473–492. [Google Scholar] [CrossRef]
- Cernusak, L.A.; Ubierna, N. Carbon Isotope Effects in Relation to CO2 Assimilation by Tree Canopies. In Stable Isotopes in Tree Rings; Springer International Publishing: Cham, Switzerland, 2022; pp. 291–310. [Google Scholar] [CrossRef]
- Campbell, G.S.; Norman, J.M. An Introduction to Environmental Biophysics, 2nd ed.; Springer: New York, NY, USA, 1998. [Google Scholar] [CrossRef]
- Speer, J.H. Fundamentals of Tree-Ring Research; The University of Arizona Press: Tucson, AZ, USA, 2010; p. 333. [Google Scholar]
- McCarroll, D.; Loader, N.J. Stable isotopes in tree rings. Quat. Sci. Rev. 2004, 23, 771–801. [Google Scholar] [CrossRef]
- McCarroll, D.; Pawellek, F. Stable carbon isotope ratios of Pinus sylvestris from northern Finland and the potential for extracting a climate signal from long Fennoscandian chronologies. Holocene 2001, 11, 517–526. [Google Scholar] [CrossRef]
- Treydte, K.S.; Frank, D.C.; Saurer, M.; Helle, G.; Schleser, G.H.; Esper, J. Impact of climate and CO2 on a millennium-long tree-ring carbon isotope record. Geochim. Cosmochim. Acta 2009, 73, 4635–4647. [Google Scholar] [CrossRef]
- Jin, X.; Xu, Q.; Liu, S.; Jiang, C.Q. Relationships of stable carbon isotope of Abies faxoniana tree-rings to climate in sub-alpine forest in Western Sichuan. Acta Ecol. Sin. 2014, 34, 1831–1840, (In Chinese, with English Abstract). [Google Scholar]
- Yu, G.R.; Wang, Q.F. Physiological ecology of photosynthesis, transpiration, and water use in plants. J. Nucl. Agric. 2010, 24, 579, (In Chinese, with English Abstract). [Google Scholar]
- Zhang, R.B.; Yuan, Y.J.; Gou, X.H.; Zhang, T.W.; Zou, C.; Ji, C.R.; Fan, Z.A.; Qin, L.; Shang, H.M.; Li, X.J. Intra-annual radial growth of Schrenk spruce (Picea schrenkiana Fisch. et Mey) and its response to climate on the northern slopes of the Tianshan Mountains. Dendrochronologia 2016, 40, 36–42. [Google Scholar] [CrossRef]
- Schleser, G.H.; Frielingsdorf, J.; Blair, A. Carbon isotope behaviour in wood and cellulose during artifcial aging. Chem. Geol. 1999, 158, 121–130. [Google Scholar] [CrossRef]
- Xu, G.B.; Liu, X.H.; Qin, D.H.; Chen, T.; Sun, W.Z.; An, W.L.; Wang, W.Z.; Wu, G.J.; Zeng, X.M.; Ren, J.W. Drought history inferred from tree ring δ13C and δ18O in the central Tianshan Mountains of China and linkage with the North Atlantic Oscillation. Theor. Appl. Climatol. 2014, 116, 385–401. [Google Scholar] [CrossRef]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sampling Sites | Code Name | Latitude/N | Longitude/E | Elevation/m | Aspect | Slope (°) | Canopy Closure | |
|---|---|---|---|---|---|---|---|---|
| L. sibirica | Banfang Gorge | BFL | 43°11′ | 94°10′ | 2900–2950 | N | 10–15 | 0.3 |
| P. schrenkiana | Tianshan Temple | TSS | 43°18′ | 93°41′ | 2620 | NE | 30 | 0.5 |
| Codename | Year | Maximum | Minimum | Mean | Variance | Standard Deviation | Coefficient of Variation |
|---|---|---|---|---|---|---|---|
| BFL | 1850–2017 | −20.287‰ | −23.619‰ | −21.822‰ | 0.541 | 0.735 | −0.0337 |
| TSS | 1850–2017 | −20.581‰ | −24.603‰ | −21.912‰ | 0.701 | 0.837 | −0.0382 |
| p7c6 | p8c7 | p9c8 | p10c9 | c5c8 | c5c9 | c6c7 | c6c8 | c6c9 | c6c10 | c7c8 | |
|---|---|---|---|---|---|---|---|---|---|---|---|
| T | −0.035 | 0.021 | 0.061 | 0.250 | 0.521 ** | 0.493 ** | 0.411 * | 0.529 ** | 0.418 * | 0.298 | 0.497 ** |
| Tmax | 0.140 | 0.198 | 0.247 | 0.302 | 0.49 ** | 0.477 ** | 0.383 * | 0.526 ** | 0.439 ** | 0.36 * | 0.495 ** |
| P | −0.435 ** | −0.581 ** | −0.589 ** | −0.406 * | −0.483 ** | −0.494 ** | −0.375 * | −0.442 ** | −0.462 ** | −0.483 ** | −0.445 ** |
| RH | −0.621 ** | −0.663 ** | −0.671 ** | −0.681 ** | −0.739 ** | −0.759 ** | −0.717 ** | −0.724 ** | −0.741 ** | −0.715 ** | −0.636 ** |
| VPD | 0.562 ** | 0.655 ** | 0.711 ** | 0.704 ** | 0.699 ** | 0.727 ** | 0.664 ** | 0.707 ** | 0.717 ** | 0.707 ** | 0.623 ** |
| p7c6 | P8c7 | p9c8 | p10c9 | c5c8 | c5c9 | c6c7 | c6c8 | c6c9 | c6c10 | c7c8 | |
|---|---|---|---|---|---|---|---|---|---|---|---|
| T | −0.544 ** | −0.526 ** | −0.514 ** | −0.491 ** | −0.436 ** | −0.474 ** | −0.477 ** | −0.492 ** | −0.513 ** | −0.52 ** | −0.459 ** |
| Tmin | −0.638 ** | −0.634 ** | −0.627 ** | −0.635 ** | −0.656 ** | −0.667 ** | −0.678 ** | −0.671 ** | −0.677 ** | −0.692 ** | −0.653 ** |
| P | −0.422 ** | −0.531 ** | −0.55 ** | −0.5 ** | −0.432 ** | −0.48 ** | −0.378 ** | −0.377 ** | −0.424 ** | −0.452 ** | −0.267 * |
| RH | 0.474 ** | 0.437 ** | 0.41 ** | 0.338 ** | 0.324 * | 0.357 ** | 0.26 * | 0.351 ** | 0.376 ** | 0.391 ** | 0.383 ** |
| VPD | −0.597 ** | −0.536 ** | −0.488 ** | −0.453 ** | −0.403 ** | −0.432 ** | −0.373 ** | −0.434 ** | −0.456 ** | −0.466 ** | −0.431 ** |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jia, Y.; Lv, G.; Guligena, H.; Qin, L.; Peng, Z.; Abudureheman, R.; Zhang, R. Differences in the Responses of Tree-Ring Stable Carbon Isotopes of L. sibirica and P. schrenkiana to Climate in the Eastern Tianshan Mountains. Forests 2023, 14, 1032. https://0-doi-org.brum.beds.ac.uk/10.3390/f14051032
Jia Y, Lv G, Guligena H, Qin L, Peng Z, Abudureheman R, Zhang R. Differences in the Responses of Tree-Ring Stable Carbon Isotopes of L. sibirica and P. schrenkiana to Climate in the Eastern Tianshan Mountains. Forests. 2023; 14(5):1032. https://0-doi-org.brum.beds.ac.uk/10.3390/f14051032
Chicago/Turabian StyleJia, Yi, Guanghui Lv, Hailili Guligena, Li Qin, Zhengbing Peng, Ruxianguli Abudureheman, and Ruibo Zhang. 2023. "Differences in the Responses of Tree-Ring Stable Carbon Isotopes of L. sibirica and P. schrenkiana to Climate in the Eastern Tianshan Mountains" Forests 14, no. 5: 1032. https://0-doi-org.brum.beds.ac.uk/10.3390/f14051032