
Identification of Candidate Genes Involved in Bud Growth in Pinus pinaster through Knowledge Transfer from Arabidopsis thaliana Models

 , , , and
, , , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Selection of Candidate Genes in Arabidopsis thaliana and Identification of Homologue Sequences in Pinus pinaster
2.1.1. Selection of Candidate Genes in Arabidopsis thaliana
2.1.2. Identification of Homologue Sequences in Pinus pinaster, Sequence Search, and Comparison of Functional Domains
2.2. Expression Analyses
2.2.1. Plant Material
2.2.2. RNA Isolation and cDNA Synthesis
2.2.3. Quantitative Real-Time PCR (RT-qPCR)
2.2.4. Data Analysis
3. Results
3.1. Selection of Candidate Genes in Arabidopsis thaliana and Identification of Homologue Sequences in Pinus pinaster
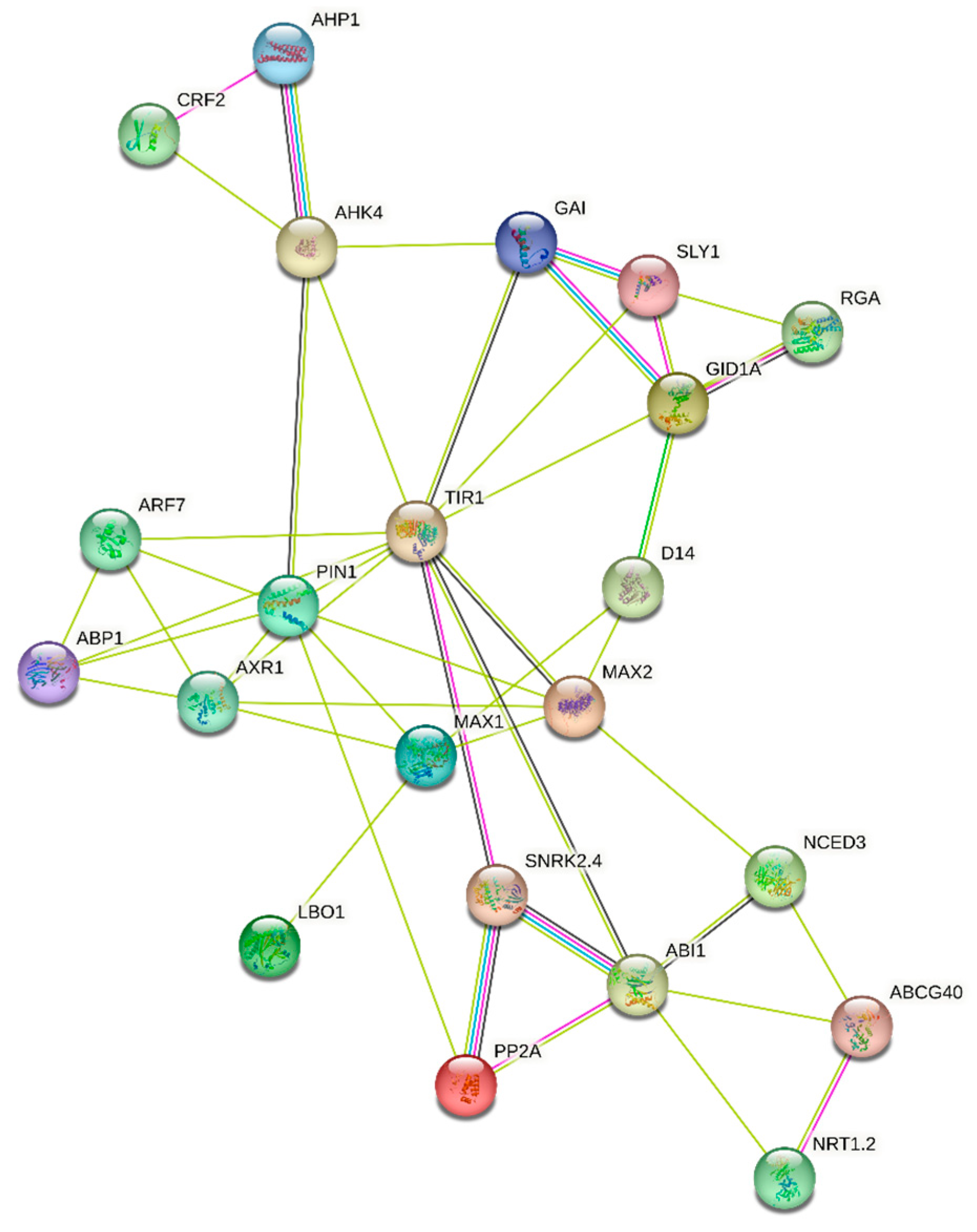
3.2. Bioinformatic Analysis of Homologues in Pinus pinaster and Their Functional Domains
3.3. Expression Analyses
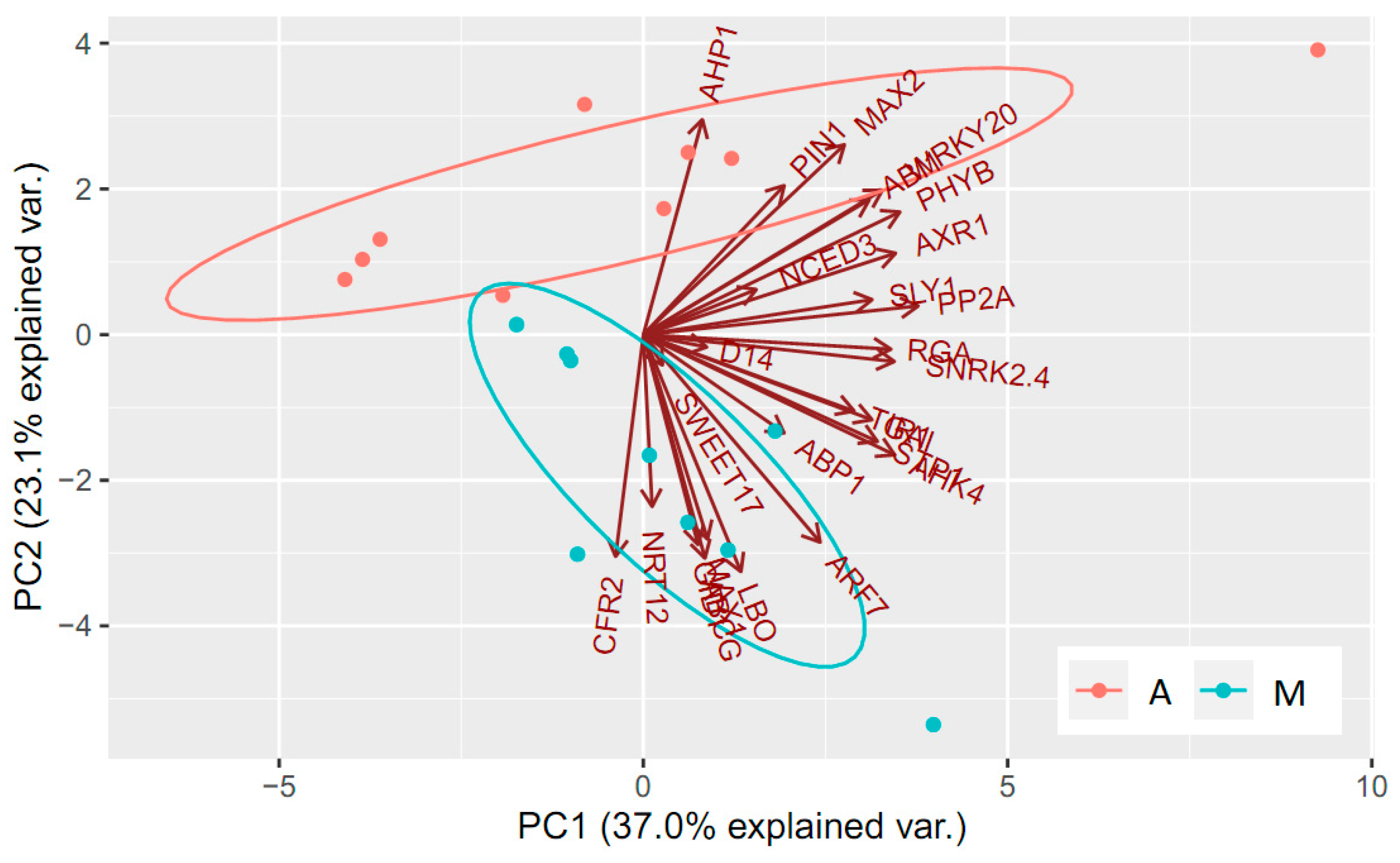
3.4. Multivariate Analyses of Gene Expression Data
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Vennetier, M.; Girard, F.; Taugourdeau, O.; Cailleret, M.; Caraglio, Y.; Sabatier, S.-A.; Ouarmim, S.; Didier, C.; Thabeet, A.; Vennetier, M.; et al. Climate Change Impact on Tree Architectural Development and Leaf Area. In Climate Change—Realities, Impacts Over Ice Cap, Sea Level and Risks; InTech: London, UK, 2013. [Google Scholar] [CrossRef]
- Anfang, M.; Shani, E. Transport Mechanisms of Plant Hormones. Curr. Opin. Plant Biol. 2021, 63, 102055. [Google Scholar] [CrossRef] [PubMed]
- Mason, M.G.; Ross, J.J.; Babst, B.A.; Wienclaw, B.N.; Beveridge, C.A. Sugar Demand, Not Auxin, Is the Initial Regulator of Apical Dominance. Proc. Natl. Acad. Sci. USA 2014, 111, 6092–6097. [Google Scholar] [CrossRef]
- Rameau, C.; Bertheloot, J.; Leduc, N.; Andrieu, B.; Foucher, F.; Sakr, S. Multiple Pathways Regulate Shoot Branching. Front. Plant Sci. 2015, 5, 741. [Google Scholar] [CrossRef]
- Emenecker, R.J.; Strader, L.C. Auxin-Abscisic Acid Interactions in Plant Growth and Development. Biomolecules 2020, 10, 281. [Google Scholar] [CrossRef] [PubMed]
- Weijers, D.; Wagner, D. Transcriptional Responses to the Auxin Hormone. Annu. Rev. Plant Biol. 2016, 67, 539–574. [Google Scholar] [CrossRef]
- Cherenkov, P.; Novikova, D.; Omelyanchuk, N.; Levitsky, V.; Grosse, I.; Weijers, D.; Mironova, V. Diversity of Cis-Regulatory Elements Associated with Auxin Response in Arabidopsis thaliana. J. Exp. Bot. 2018, 69, 329–339. [Google Scholar] [CrossRef]
- Enders, T.A.; Strader, L.C. Auxin Activity: Past, Present, and Future. Am. J. Bot. 2015, 102, 180–196. [Google Scholar] [CrossRef]
- Skalický, V.; Kubeš, M.; Napier, R.; Novák, O. Auxins and Cytokinins—The Role of Subcellular Organization on Homeostasis. Int. J. Mol. Sci. 2018, 19, 3115. [Google Scholar] [CrossRef]
- Alvarez, J.M.; Bueno, N.; Cuesta, C.; Feito, I.; Ordás, R.J. Hormonal and Gene Dynamics in de Novo Shoot Meristem Formation during Adventitious Caulogenesis in Cotyledons of Pinus pinea. Plant Cell Rep. 2020, 39, 527–541. [Google Scholar] [CrossRef]
- López-Hinojosa, M.; de María, N.; Guevara, M.A.; Vélez, M.D.; Cabezas, J.A.; Díaz, L.M.; Mancha, J.A.; Pizarro, A.; Manjarrez, L.F.; Collada, C.; et al. Rootstock Effects on Scion Gene Expression in Maritime Pine. Sci. Rep. 2021, 11, 11582. [Google Scholar] [CrossRef]
- Valledor, L.; Guerrero, S.; García-Campa, L.; Meijón, M. Proteometabolomic Characterization of Apical Bud Maturation in Pinus pinaster. Tree Physiol. 2021, 41, 508–521. [Google Scholar] [CrossRef]
- De Miguel, M.; Sánchez-Gómez, D.; Cervera, M.T.; Aranda, I. Functional and Genetic Characterization of Gas Exchange and Intrinsic Water Use Efficiency in a Full-Sib Family of Pinus pinaster Ait. in Response to Drought. Tree Physiol. 2012, 32, 94–103. [Google Scholar] [CrossRef] [PubMed]
- Alía, R.; Martín, S. Technical Guidelines for Genetic Conservation and Use for Maritime Pine (Pinus pinaster Ait.); EUFORGEN: Rome, Italy, 2003. [Google Scholar]
- Bueno, N.; Cuesta, C.; Centeno, M.L.; Ordás, R.J.; Alvarez, J.M. In Vitro Plant Regeneration in Conifers: The Role of WOX and KNOX Gene Families. Genes 2021, 12, 438. [Google Scholar] [CrossRef] [PubMed]
- Valledor, L.; Carbó, M.; Lamelas, L.; Escandón, M.; Colina, F.J.; Cañal, M.J.; Meijón, M. When the Tree Let Us See the Forest: Systems Biology and Natural Variation Studies in Forest Species. Prog. Bot. 2018, 81, 353–375. [Google Scholar] [CrossRef]
- Lee, Z.H.; Hirakawa, T.; Yamaguchi, N.; Ito, T. The Roles of Plant Hormones and Their Interactions with Regulatory Genes in Determining Meristem Activity. Int. J. Mol. Sci. 2019, 20, 4065. [Google Scholar] [CrossRef] [PubMed]
- Luo, Z.; Janssen, B.J.; Snowden, K.C. The Molecular and Genetic Regulation of Shoot Branching. Plant Physiol. 2021, 187, 1033–1044. [Google Scholar] [CrossRef]
- Altschul, S.F.; Madden, T.L.; Schäffer, A.A.; Zhang, J.; Zhang, Z.; Miller, W.; Lipman, D.J. Gapped BLAST and PSI-BLAST: A New Generation of Protein Database Search Programs. Nucleic Acids Res. 1997, 25, 3389–3402. [Google Scholar] [CrossRef]
- Thompson, J.D.; Higgins, D.G.; Gibson, T.J. CLUSTAL W: Improving the Sensitivity of Progressive Multiple Sequence Alignment through Sequence Weighting, Position-Specific Gap Penalties and Weight Matrix Choice. Nucleic Acids Res. 1994, 22, 4673–4680. [Google Scholar] [CrossRef]
- Udvardi, M.K.; Czechowski, T.; Scheible, W.R. Eleven Golden Rules of Quantitative RT-PCR. Plant Cell 2008, 20, 1736–1737. [Google Scholar] [CrossRef]
- Bustin, S.A.; Benes, V.; Garson, J.A.; Hellemans, J.; Huggett, J.; Kubista, M.; Mueller, R.; Nolan, T.; Pfaffl, M.W.; Shipley, G.L.; et al. The MIQE Guidelines: Minimum Information for Publication of Quantitative Real-Time PCR Experiments. Clin. Chem. 2009, 55, 611–622. [Google Scholar] [CrossRef]
- Álvarez, J.M.; Cortizo, M.; Ordás, R.J. Characterization of a Type-A Response Regulator Differentially Expressed during Adventitious Caulogenesis in Pinus pinaster. J. Plant Physiol. 2012, 169, 1807–1814. [Google Scholar] [CrossRef] [PubMed]
- Alonso, P.; Cortizo, M.; Canton, F.R.; Fernández, B.; Rodríguez, A.; Centeno, M.L.; Cánovas, F.M.; Ordás, R.J. Identification of Genes Differentially Expressed during Adventitious Shoot Induction in Pinus pinea Cotyledons by Subtractive Hybridization and Quantitative PCR. Tree Physiol. 2007, 27, 1721–1730. [Google Scholar] [CrossRef] [PubMed]
- Alvarez, J.M.; Cortizo, M.; Bueno, N.; Rodríguez, A.; Ordás, R.J. CLAVATA1-LIKE, a Leucine-Rich-Repeat Protein Receptor Kinase Gene Differentially Expressed during Adventitious Caulogenesis in Pinus pinaster and Pinus pinea. Plant Cell Tissue Organ. Cult. 2013, 112, 331–342. [Google Scholar] [CrossRef]
- Rozen, S.; Skaletsky, H. Primer3 on the WWW for General Users and for Biologist Programmers. Methods Mol. Biol. 2000, 132, 365–386. [Google Scholar] [CrossRef]
- Ritz, C.; Spiess, A.N. QpcR: An R Package for Sigmoidal Model Selection in Quantitative Real-Time Polymerase Chain Reaction Analysis. Bioinformatics 2008, 24, 1549–1551. [Google Scholar] [CrossRef]
- Gonzalez-Grandio, E.; Pajoro, A.; Franco-Zorrilla, J.M.; Tarancon, C.; Immink, R.G.H.; Cubas, P. Abscisic Acid Signaling Is Controlled by a BRANCHED1/HD-ZIP I Cascade in Arabidopsis Axillary Buds. Proc. Natl. Acad. Sci. USA 2017, 114, E245–E254. [Google Scholar] [CrossRef]
- Máthé, C.; M-Hamvas, M.; Freytag, C.; Garda, T. The Protein Phosphatase PP2A Plays Multiple Roles in Plant Development by Regulation of Vesicle Traffic-Facts and Questions. Int. J. Mol. Sci. 2021, 22, 975. [Google Scholar] [CrossRef]
- Qiu, Y.; Guan, S.C.; Wen, C.; Li, P.; Gao, Z.; Chen, X. Auxin and Cytokinin Coordinate the Dormancy and Outgrowth of Axillary Bud in Strawberry Runner. BMC Plant Biol. 2019, 19, 528. [Google Scholar] [CrossRef]
- Hallmark, H.T.; Rashotte, A.M. Review—Cytokinin Response Factors: Responding to More than Cytokinin. Plant Sci. 2019, 289, 110251. [Google Scholar] [CrossRef]
- Li, S.M.; Zheng, H.X.; Zhang, X.S.; Sui, N. Cytokinins as Central Regulators during Plant Growth and Stress Response. Plant Cell Rep. 2021, 40, 271–282. [Google Scholar] [CrossRef]
- Walker, C.H.; Siu-Ting, K.; Taylor, A.; O’Connell, M.J.; Bennett, T. Strigolactone Synthesis Is Ancestral in Land Plants, but Canonical Strigolactone Signalling Is a Flowering Plant Innovation. BMC Biol. 2019, 17, 70. [Google Scholar] [CrossRef] [PubMed]
- Waters, M.T.; Gutjahr, C.; Bennett, T.; Nelson, D.C. Strigolactone Signaling and Evolution. Annu. Rev. Plant Biol. 2017, 68, 291–322. [Google Scholar] [CrossRef] [PubMed]
- Yamada, K.; Kanai, M.; Osakabe, Y.; Ohiraki, H.; Shinozaki, K.; Yamaguchi-Shinozaki, K. Monosaccharide Absorption Activity of Arabidopsis Roots Depends on Expression Profiles of Transporter Genes under High Salinity Conditions. J. Biol. Chem. 2011, 286, 43577–43586. [Google Scholar] [CrossRef] [PubMed]
- Lemonnier, P.; Gaillard, C.; Veillet, F.; Verbeke, J.; Lemoine, R.; Coutos-Thévenot, P.; La Camera, S. Expression of Arabidopsis Sugar Transport Protein STP13 Differentially Affects Glucose Transport Activity and Basal Resistance to Botrytis cinerea. Plant Mol. Biol. 2014, 85, 473–484. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| Arabidopsis thaliana | Pinus pinaster | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Protein | ID | Length (aa) | ID | Length (aa) | E value | Identity % | Positive % (BLSM62) | |||
| Hormonal | Abscisic acid | Synthesis | NCED3 | AT3G14440 | 599 | PPI00015334 | 412 | 0.0 | 68.3 | 81.1 |
| Transport | ABCG40 | AT1G15210 | 1442 | PPI00033391 | 1457 | 0.0 | 63.1 | 78.4 | ||
| NRT1.2 | AT1G69850 | 585 | PPI00013776 | 598 | 0.0 | 48.5 | 67.3 | |||
| Signaling | SNRK2.4 | AT1G10940 | 371 | PPI00050983 | 455 | 0.0 | 80.1 | 90.1 | ||
| ABI1 | AT4G26080 | 434 | PPI00056219 | 594 | 1.0 × 10−111 | 49.9 | 65.9 | |||
| Auxins | Transport | PIN1 | AT1G73590 | 622 | PPI00011546 | 695 | 0.0 | 57.1 | 67.8 | |
| PP2A | AT1G69960 | 307 | PPI00008039 | 306 | 0.0 | 90.8 | 97.4 | |||
| Signaling | ABP1 | AT4G02980 | 198 | PPI00009705 | 160 | 2.0 × 10−53 | 49.9 | 65.9 | ||
| TIR1 | AT3G62980 | 594 | PPI00012072 | 574 | 0.0 | 65.4 | 79.3 | |||
| ARF7 | AT5G20730 | 1165 | PPI00041807 | 497 | 1.0 × 10−180 | 71.5 | 83.3 | |||
| AXR1 | AT1G05180 | 540 | PPI00013097 | 560 | 0.0 | 67.0 | 82.4 | |||
| Cytokinins | Signaling | AHK4 | AT2G01830 | 1080 | PPI00064460 | 1036 | 0.0 | 56.7 | 70.3 | |
| AHP1 | AT3G21510 | 154 | PPI00050577 | 156 | 4.0 × 10−62 | 57.6 | 80.1 | |||
| CRF2 | AT4G23750 | 343 | PPI00010365 | 270 | 3.0 × 10−31 | 43.1 | 56.9 | |||
| Strigolactones | Synthesis | LBO1 | AT3G21420 | 364 | PPI00010339 | 377 | 8.0 × 10−113 | 47.1 | 67.3 | |
| MAX1 | AT2G26170 | 522 | PPI00016810 | 421 | 3.0 × 10−134 | 53.4 | 69.3 | |||
| Signaling | D14 | AT3G03990 | 267 | PPI00017643 | 267 | 4.0 × 10−133 | 65.8 | 83.1 | ||
| MAX2 | AT2G42620 | 693 | PPI00014698 | 329 | 6.0 × 10−86 | 54.5 | 67.1 | |||
| Gibberellins | Signaling | GID1A | AT3G05120 | 345 | PPI00071627 | 357 | 1.0 × 10−162 | 66.6 | 78.2 | |
| GAI | AT1G14920 | 533 | PPI00014310 | 458 | 3.0 × 10−174 | 62.6 | 77.8 | |||
| RGA | AT2G01570 | 587 | PPI00011857 | 594 | 5.0 × 10−174 | 49.6 | 66.0 | |||
| SLY1 | AT4G24210 | 151 | PPI00016475 | 219 | 3.0 × 10−38 | 48.9 | 63.7 | |||
| Non-hormonal | Sugars | Transport | STP1 | AT1G11260 | 522 | PPI00012920 | 514 | 0.0 | 65.0 | 77.9 |
| SWEET17 | AT4G15920 | 241 | PPI00061954 | 280 | 8.0 × 10−75 | 52.9 | 71.7 | |||
| WRKY20 | AT4G26640 | 557 | PPI00011874 | 679 | 4.0 × 10−90 | 42.5 | 58.6 | |||
| Light | Signaling | PHYB | AT2G18790 | 1172 | PPI00062444 | 1139 | 0.0 | 67.3 | 81.5 | |
| Gene | Main Apical Bud | Main Whorl Bud | Secondary Apical Bud | |
|---|---|---|---|---|
| Abscisic acid | NCED3 | A ** | nd | nd |
| NRT1.2 | M * | nd | M ** | |
| Auxins | PIN1 | A * | nd | nd |
| PP2A | nd | M * | nd | |
| ARF7 | nd | nd | M * | |
| Cytokinins | AHK4 | nd | M * | nd |
| AHP1 | nd | A * | nd | |
| Strigolactones | MAX1 | nd | nd | M * |
| MAX2 | A ** | nd | nd | |
| Gibberellins | GID1 | M ** | nd | nd |
| Sugars | STP1 | nd | M * | M ** |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Alvarez, J.M.; Rodríguez, S.M.; Fuente-Maqueda, F.; Feito, I.; Ordás, R.J.; Cuesta, C. Identification of Candidate Genes Involved in Bud Growth in Pinus pinaster through Knowledge Transfer from Arabidopsis thaliana Models. Forests 2023, 14, 1765. https://0-doi-org.brum.beds.ac.uk/10.3390/f14091765
Alvarez JM, Rodríguez SM, Fuente-Maqueda F, Feito I, Ordás RJ, Cuesta C. Identification of Candidate Genes Involved in Bud Growth in Pinus pinaster through Knowledge Transfer from Arabidopsis thaliana Models. Forests. 2023; 14(9):1765. https://0-doi-org.brum.beds.ac.uk/10.3390/f14091765
Chicago/Turabian StyleAlvarez, José Manuel, Sonia María Rodríguez, Francisco Fuente-Maqueda, Isabel Feito, Ricardo Javier Ordás, and Candela Cuesta. 2023. "Identification of Candidate Genes Involved in Bud Growth in Pinus pinaster through Knowledge Transfer from Arabidopsis thaliana Models" Forests 14, no. 9: 1765. https://0-doi-org.brum.beds.ac.uk/10.3390/f14091765