
Assessment of Artificial Forest Restoration by Exploring the Microbial Community Structure and Function in a Reclaimed Coal Gob Pile in a Loess Hilly Area of Shanxi, China

Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Location and Soil Sampling
2.2. Analysis of Soil Physical and Chemical Properties
2.3. Soil DNA Extraction and Quantitative PCR Analysis of Bacterial 16S rRNA
2.4. Functional Genes Analysis
2.5. Statistical Analysis
3. Results
3.1. Soil Physico-Chemical and Enzyme Activity Properties
3.2. Soil Microbial Community Composition and Diversity
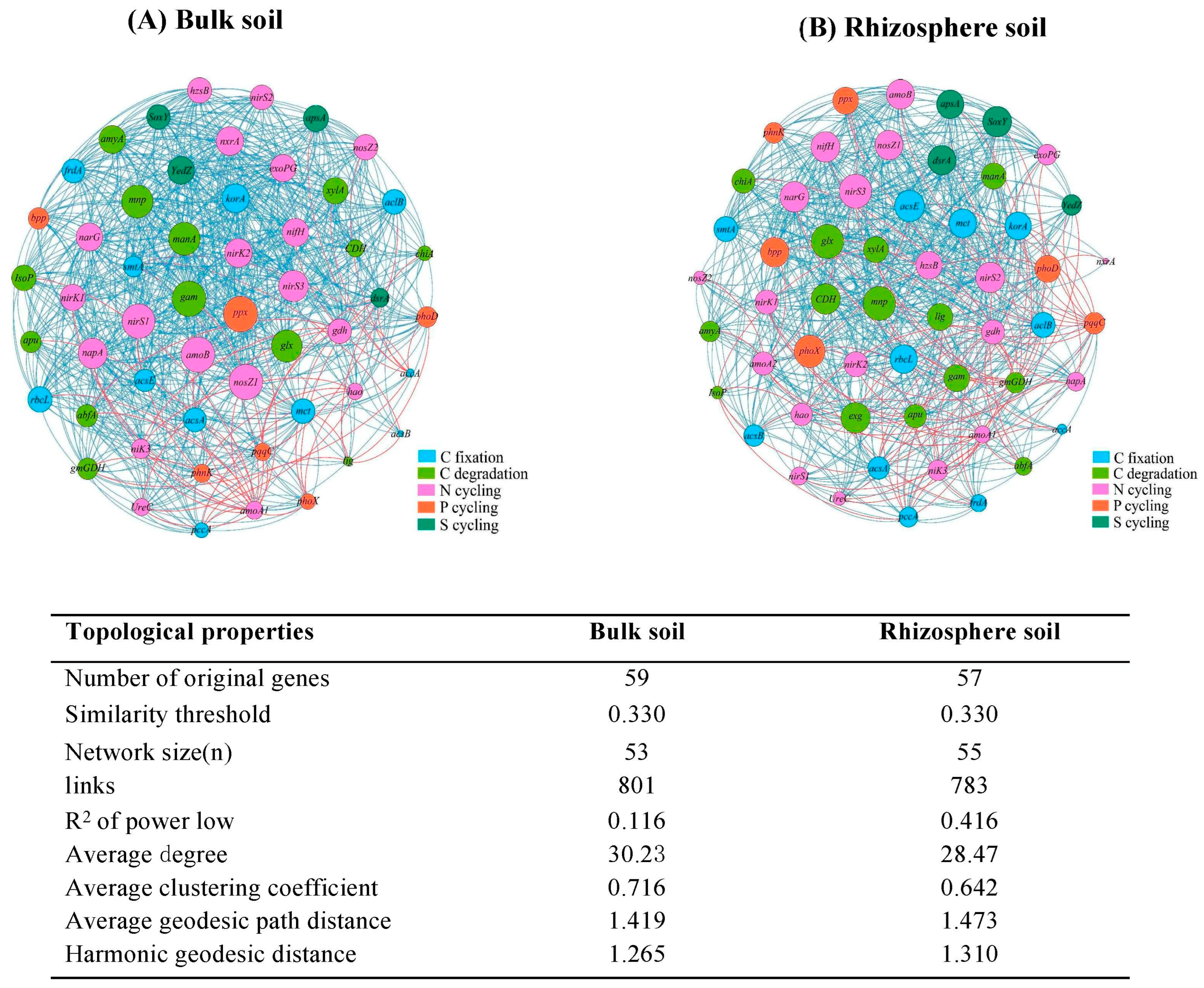
3.3. Soil C-, N-, P-, and S-Cycling Genes
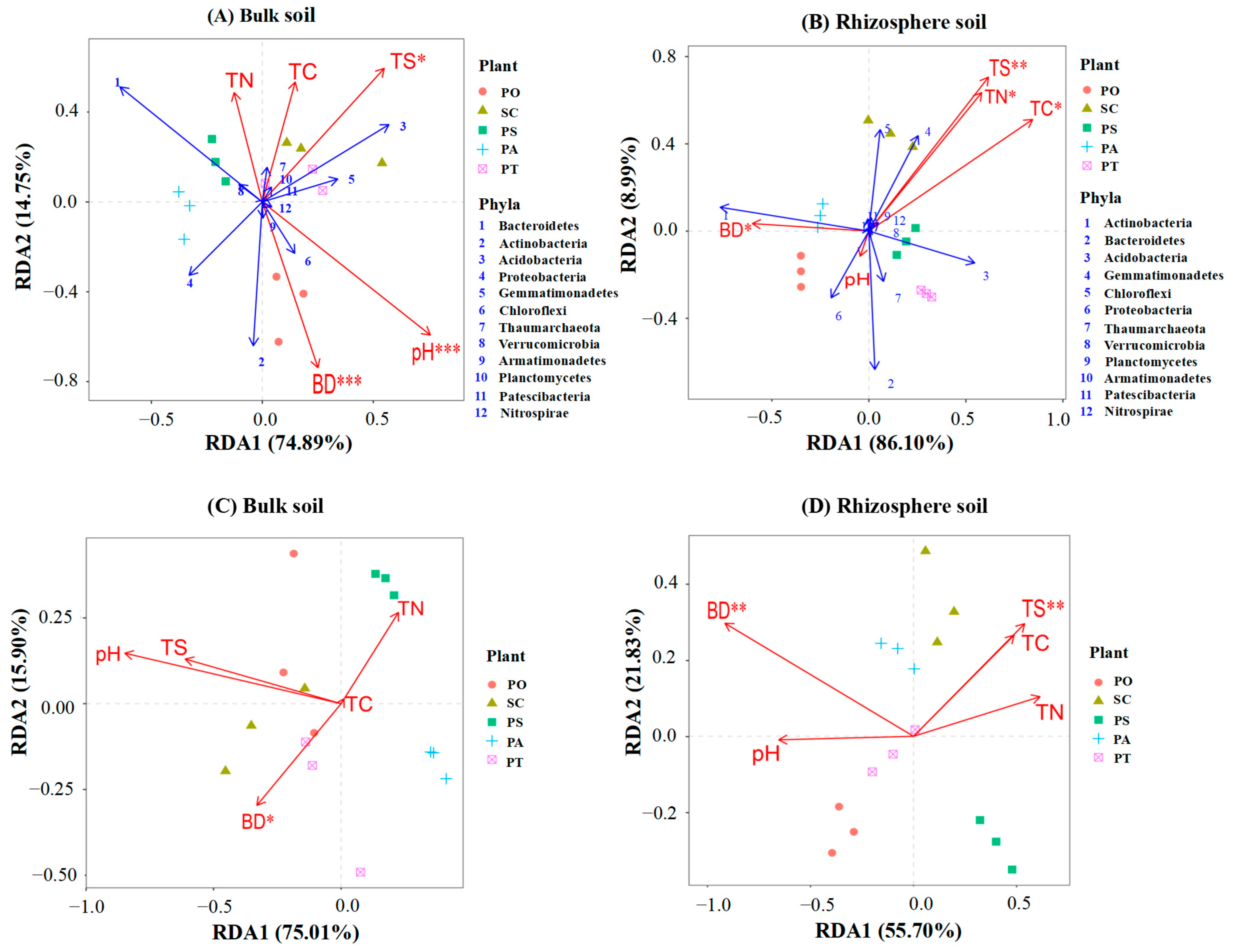
3.4. The Effect of Edaphic Factors on Microbial Community and Function
4. Discussion
4.1. Changes in Soil Physico-Chemical Properties
4.2. Changes in Microbial Community and Functional Characteristics
4.3. Relationships among Edaphic Factors and Microbial Community Function
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Bai, Z.K.; Fu, M.C.; Zhao, Z.Q. On soil environmental problems in mining area. Ecol. Environ. 2006, 15, 1122–1125. [Google Scholar]
- Topp, W.; Thelen, K.; Kappes, H. Soil dumping techniques and afforestation drive ground-dwelling beetle assemblages in a 25-year-old open-cast mining reclamation area. Ecol. Eng. 2010, 36, 751–756. [Google Scholar] [CrossRef]
- Zhao, D.; Hou, H.P.; Liu, H.Y.; Wang, C.; Ding, Z.Y.; Xiong, J.T. Microbial community structure and predictive functional analysis in reclaimed soil with different vegetation types: The example of the Xiaoyi mine waste dump in Shanxi. Land 2023, 12, 456. [Google Scholar] [CrossRef]
- Zhou, W.; Yang, K.; Bai, Z.K.; Cheng, H.X.; Liu, F. The development of topsoil properties under different reclaimed land uses in the Pingshuo opencast coalmine of Loess Plateau of China. Ecol. Eng. 2017, 100, 237–245. [Google Scholar] [CrossRef]
- Li, S.Q.; Liber, K. Influence of different revegetation choices on plant community and soil development nine years after initial planting on a reclaimed coal gob pile in the Shanxi mining area, China. Sci. Total Environ. 2018, 618, 1314–1323. [Google Scholar] [CrossRef]
- Liu, X.Y.; Zhou, W.; Bai, Z.K. Vegetation coverage change and stability in large open-pit coal mine dumps in China during 1990–2015. Ecol. Eng. 2016, 95, 447–451. [Google Scholar] [CrossRef]
- Zhang, Y.; Yang, J.Y.; Wu, H.L.; Shi, C.Q.; Zhang, C.L.; Li, D.X.; Feng, M.M. Dynamic changes in soil and vegetation during varying ecological recovery conditions of abandoned mines in Beijing. Ecol. Eng. 2014, 73, 676–683. [Google Scholar] [CrossRef]
- Li, S.Q.; Wu, D.M.; Zhang, J.T. Effects of vegetation and fertilization on weathered particles of coal gob in Shanxi mining areas, China. J. Hazard. Mater. 2005, 124, 209–216. [Google Scholar] [CrossRef]
- Querol, X.; Izquierdo, M.; Monfort, E.; Alvarez, E.; Font, O.; Moreno, T.; Alastuey, A.; Zhuang, X.; Lu, W.; Wang, Y. Environmental characterization of burnt coal gangue banks at Yangquan, Shanxi Province, China. Int. J. Coal Geol. 2008, 75, 93–104. [Google Scholar] [CrossRef]
- Tang, X.; Snowden, S.; McLellan, B.C.; Höök, M. Clean coal use in China: Challenges and policy implications. Energ. Policy 2015, 87, 517–523. [Google Scholar] [CrossRef]
- Gao, X.B.; Hu, Y.D.; Li, C.C.; Dai, C.; Li, L.; Ou, X.; Wang, Y.X. Evaluation of fluorine release from air deposited coal spoil piles: A case study at Yangquan city, northern China. Sci. Total Environ. 2016, 545–546, 1–10. [Google Scholar] [CrossRef]
- Biswas, A.; Hendry, M.J.; Essilfie-Dughan, J. Geochemistry of arsenic in low sulfidehigh carbonate coal waste rock, Elk Valley, British Columbia, Canada. Sci. Total Environ. 2017, 579, 396–408. [Google Scholar] [CrossRef]
- Tripathi, N.; Singh, R.S.; Hills, C.D. Soil carbon development in rejuvenated Indian coal mine spoil. Ecol. Eng. 2016, 90, 482–490. [Google Scholar] [CrossRef]
- Mukhopadhyay, S.; George, J.; Masto, R.E. Changes in polycyclic aromatic hydrocarbons (PAHs) and soil biological parameters in a revegetated coal mine spoil. Land Degrad. Dev. 2017, 28, 1047–1055. [Google Scholar] [CrossRef]
- Zhang, L.; Wang, J.; Bai, Z.; Lv, C. Effects of vegetation on runoff and soil erosion on reclaimed land in an opencast coal-mine dump in a loess area. Catena 2015, 128, 44–53. [Google Scholar] [CrossRef]
- Bradshaw, A.D. Restoration of mined lands-using natural processes. Ecol. Eng. 1997, 8, 255–269. [Google Scholar] [CrossRef]
- Dobson, A.P.; Bradshaw, A.D.; Baker, A.J.M. Hopes for the future: Restoration ecology and conservation biology. Science 1997, 277, 515–522. [Google Scholar] [CrossRef]
- Huang, D.; Liu, Q.S. Remote sensing monitoring and effect evaluation on ecological restoration of heidaigou coal mining area. In Proceedings of the International Conference on Remote Sensing, Environment and Transportation Engineering, RSETE 2013, Nanjing, China, 26–28 July 2013. [Google Scholar] [CrossRef]
- Dutta, R.K.; Agrawal, M. Effect of tree plantations on the soil characteristics and microbial activity of coal mine spoil land. Trop. ecol. 2002, 43, 315–324. [Google Scholar]
- Singh, R.S.; Tripathi, N.; Chaulya, S.K. Ecological study of revegetated coal mine spoil of an Indian dry tropical ecosystem along an age gradient. Biodegradation 2012, 23, 837–849. [Google Scholar] [CrossRef]
- Upadhyay, N.; Verma, S.; Singh, A.P.; Devi, S.; Vishwakarma, K.; Kumar, N.; Pandey, A.; Dubey, K.; Mishra, R.; Tripathi, D.K.; et al. Soil ecophysiological and microbiological indices of soil health: A study of coal mining site in Sonbhadra, Uttar Pradesh. J. Soil Sci. Plant Nut. 2016, 16, 778–800. [Google Scholar] [CrossRef]
- Ahirwal, J.; Maiti, S.K.; Reddy, M.S. Development of carbon, nitrogen and phosphate stocks of reclaimed coal mine soil within 8 years after forestation with Prosopis juliflora (Sw.) Dc. Catena 2017, 156, 42–50. [Google Scholar] [CrossRef]
- Wang, L.; Ji, B.; Hu, Y.H.; Liu, R.Q.; Sun, W. A review on in situ phytoremediation of mine tailings. Chemosphere 2017, 184, 594–600. [Google Scholar] [CrossRef] [PubMed]
- Fu, Y.; Lin, C.C.; Ma, J.J.; Zhu, T.C. Effects of plant types on physico-chemical properties of reclaimed mining soil in Inner Mongolia, China. Chin. Geogr. Sci. 2010, 20, 309–317. [Google Scholar] [CrossRef]
- Zhao, Z.; Shahrour, I.; Bai, Z.; Fan, W.; Feng, L.; Li, H. Soils development in opencast coal mine spoils reclaimed for 1–13 years in the West-Northern Loess Plateau of China. Eur. J. Soil Biol. 2013, 55, 40–46. [Google Scholar] [CrossRef]
- Nawaz, M.F.; Bourrie, G.; Trolard, F. Soil compaction impact and modelling. A review. Agron. Sustain. Dev. 2013, 33, 291–309. [Google Scholar] [CrossRef]
- Wali, M.K. Ecological succession and the rehabilitation of disturbed terrestrial ecosystems. Plant Soil 1999, 213, 195–220. [Google Scholar] [CrossRef]
- Yang, R.X.; Wang, J.M. The change law of RMSs characteristics in grassland opencast coal mine dump of China. Adv. Environ. Technol. 2013, 726, 4831–4837. [Google Scholar] [CrossRef]
- Ritter, J.B.; Gardner, T.W. Hydrologic evolution of drainage basins disturbed by surface mining, central Pennsylvania. Geol. Soc. Am. Bull. 1993, 105, 101–115. [Google Scholar] [CrossRef]
- Cejpek, J.; Kuraz, V.; Frouz, J. Hydrological properties of soils in reclaimed and unreclaimed sites after brown-coal mining. Pol. J. Environ. Stud. 2013, 22, 645–652. [Google Scholar]
- Courtney, R.; Mullen, G.; Harrington, T. An evaluation of revegetation successon bauxite residue. Restor. Ecol. 2009, 17, 350–358. [Google Scholar] [CrossRef]
- Kaschuk, G.; Alberton, O.; Hungria, M. Three decades of soil microbial biomass studies in Brazilian ecosystems: Lessons learned about soil quality and indications for improving sustainability. Soil Biol. Biochem. 2010, 42, 1–13. [Google Scholar] [CrossRef]
- Chen, J.; Mo, L.; Zhang, Z.C.; Nan, J.; Xu, D.L.; Chao, L.M.; Zhang, X.D.; Bao, Y.Y. Evaluation of the ecological restoration of a coal mine dump by exploring the characteristics of microbial communities. Appl. Soil Ecol. 2020, 147, 103430. [Google Scholar] [CrossRef]
- Helingerova, M.; Frouz, J.; Santruckova, H. Microbial activity in reclaimed and unreclaimed post-mining sites near Sokolov (Czech Republic). Ecol. Eng. 2010, 36, 768–776. [Google Scholar] [CrossRef]
- Claassens, S.; van Rensburg, P.J.; Liebenberg, D.; van Rensburg, L. A comparison of microbial community function and structure in rehabilitated asbestos and coal discard sites. Water Air Soil Poll. 2012, 223, 1091–1100. [Google Scholar] [CrossRef]
- Li, J.J.; Zhou, X.M.; Yan, J.X.; Li, H.J.; He, J.Z. Effects of regenerating vegetation on soil enzyme activity and microbial structure in reclaimed soils on a surface coal mine site. Appl. Soil Ecol. 2015, 87, 56–62. [Google Scholar] [CrossRef]
- Riley, D.; Barber, S.A. Bicarbonate accumulation and pH changes at the soybean (Glycine max (L.) Merr.) root-soil interface. Soil Sci. Soc. Am. J. 1969, 33, 905–908. [Google Scholar] [CrossRef]
- Riley, D.; Barber, S.A. Salt accumulation at the soybean (Glycine max (L.) Merr.) root-soil interface. Soil Sci. Soc. Am. J. 1970, 34, 154–155. [Google Scholar] [CrossRef]
- Liu, S.; Gao, Y.Y.; Lang, H.L.; Liu, Y.; Zhang, H. Effects of conventional tillage and no-tillage systems on maize (Zea mays L.) growth and yield, soil structure, and water in loess plateau of China: Field experiment and modeling studies. Land 2022, 11, 1881. [Google Scholar] [CrossRef]
- Mc Lean, E.O. Soil pH and lime requirement. In Methods of Soil Analysis. Part 2. Chemical and Microbiological Properties, 2nd ed.; Page, A.L., Miller, R.H., Keeney, D.R., Eds.; Agronomy Monograph No. 9; ASA: Madison, WI, USA; SSSA: Madison, WI, USA, 1982; pp. 199–224. [Google Scholar]
- Krämer, S.; Green, D.M. Acid and alkaline phosphatase dynamics and their relationship to soil microclimate in a semiarid woodland. Soil Biol. Biochem. 2000, 32, 179–188. [Google Scholar] [CrossRef]
- Ladd, J.N.; Butler, J.H.A. Short-term assays of soil proteolytic enzyme activities using proteins and dipeptide derivatives as substrates. Soil Biol. Biochem. 1972, 4, 19–30. [Google Scholar] [CrossRef]
- Nannipieri, P.; Giagnoni, L.; Renella, G.; Puglisi, E.; Ceccanti, B.; Masciandaro, G.; Fornasier, F.; Moscatelli, M.C.; Marinari, S. Soil enzymology: Classical and molecular approaches. Biol. Fert. Soils 2012, 48, 743–762. [Google Scholar] [CrossRef]
- Furtak, K.; Gawryjołek, K.; Gajda, A.M.; Gajązka, A. Effects of maize and winter wheat grown under different cultivation techniques on biological activity of soil. Plant Soil Environ. 2017, 63, 449–454. [Google Scholar] [CrossRef]
- Dennis, K.L.; Wang, Y.W.; Blatner, N.R.; Wang, S.Y.; Saadalla, A.; Trudeau, E.; Roers, A.; Weaver, C.T.; Lee, J.J.; Gilbert, J.A.; et al. Adenomatous polyps are driven by microbe-Instigated focal inflammation and are controlled by IL-10-producing T cells. Cancer Res. 2013, 73, 5905–5913. [Google Scholar] [CrossRef]
- Jia, L.X.; Haimeng Sun, H.M.; Zhou, Q.; Richeng Dai, R.C. Weizhong WuIntegrated evaluation for advanced removal of nitrate and phosphorus in novel PHBV/ZVI-based biofilters: Insight into functional genes and key enzymes. J. Clean. Prod. 2022, 349, 131199. [Google Scholar] [CrossRef]
- Chen, L.; Jiang, Y.; Liang, C.; Luo, Y.; Xu, Q.; Han, C.; Zhao, Q.; Sun, B. Competitive interaction with keystone taxa induced negative priming under biochar amendments. Microbiome 2019, 7, 77. [Google Scholar] [CrossRef] [PubMed]
- Kaiser, H.F. The application of electronic computers to factor analysis. Educ. Psychol. Meas. 1960, 20, 141–151. [Google Scholar] [CrossRef]
- Wander, M.M.; Bollero, G.A. Soil quality assessment of tillage impacts in Illinois. Soil Sci. Soc. Am. J. 1999, 63, 961–971. [Google Scholar] [CrossRef]
- Mukhopadhyay, S.; Maiti, S.K.; Masto, R.E. Development of mine soil quality index (MSQI) for evaluation of reclamation success: A chronosequence study. Ecol. Eng. 2014, 71, 10–20. [Google Scholar] [CrossRef]
- Ribbons, R.R.; Levy-Booth, D.J.; Masse, J.; Grayston, S.J.; McDonald, M.A.; Vesterdal, L.; Prescott, C.E. Linking microbial communities, functional genes and nitrogen-cycling processes in forest floors under four tree species. Soil Biol. Biochem. 2016, 103, 181–191. [Google Scholar] [CrossRef]
- Liu, X.Y.; Bai, Z.K.; Zhou, W.; Cao, Y.G.; Zhang, G.J. Changes in soil properties in the soil profile after mining and reclamation in an opencast coal mine on the Loess Plateau, China. Ecol. Eng. 2019, 98, 228–239. [Google Scholar] [CrossRef]
- Feng, Y.; Wang, J.M.; Bai, Z.K.; Reading, L. Effects of surface coal mining and land reclamation on soil properties: A review. Earth-Sci. Rev. 2019, 191, 12–25. [Google Scholar] [CrossRef]
- Shrestha, R.K.; Lal, R. Carbon and nitrogen pools in reclaimed land under forest and pastureecosystems in Ohio, USA. Geoderma 2010, 157, 196–205. [Google Scholar] [CrossRef]
- Frouz, J.; Elhottová, D.; Pižl, V.; Tajovský, K.; Šourková, M.; Picek, T.; Malý, S. The effect of litter quality and soil faunal composition on organic matter dynamics in postmining soil: A laboratory study. Appl. Soil Ecol. 2007, 37, 72–80. [Google Scholar] [CrossRef]
- Zipper, C.E.; Burger, J.A.; Barton, C.D.; Skousen, J.G. Rebuilding soils on mined land for native forests in Appalachia. Soil Sci. Soc. Am. J. 2013, 77, 337–349. [Google Scholar] [CrossRef]
- Fan, W.; Bai, Z.; Li, H.; Qiao, J.; Xu, J. Effects of different vegetation restoration patterns and reclamation years on microbes in reclaimed soil. Trans. Chin. Soc. Agric. Eng. 2011, 27, 330–336. (In Chinese) [Google Scholar]
- Deng, J.J.; Yin, Y.; Zhu, W.X.; Zhou, Y.B. Variations in soil bacterial community diversity and structures among different revegetation types in the baishilazi nature teserve. Front. Microbiol. 2018, 9, 2874. [Google Scholar] [CrossRef]
- Miyashita, N.T. Contrasting soil bacterial community structure between the phyla Acidobacteria and Proteobacteria in tropical southeast asian and temperate japanese forests. Genes Genet. Syst. 2015, 90, 61. [Google Scholar] [CrossRef]
- Hui, L.; Ye, D.D.; Wang, X.G.; Settles, M.L.; Wang, J.; Hao, Z.Q.; Zhou, L.S.; Dong, P.; Jiang, Y.; Ma, Z.S. Soil bacterial communities of different natural forest types in northeast china. Plant Soil 2014, 383, 203–216. [Google Scholar] [CrossRef]
- Fierer, N.; Bradford, M.A.; Jackson, R.B. Toward an ecological classification of soil bacteria. Ecology 2007, 88, 1354–1364. [Google Scholar] [CrossRef]
- Fazi, S.; Amalfitano, S.; Pernthaler, J.; Puddu, A. Bacterial communities associated with benthic organic matter in headwater stream microhabitats. Environ. Microbiol. 2005, 7, 1633–1640. [Google Scholar] [CrossRef]
- Thomson, B.C.; Tisserant, E.; Plassart, P.; Uroz, S.; Griffiths, R.I.; Hannula, S.E.; Buée, M.; Mougel, C.; Ranjard, L.; Van Veene, J.A.; et al. Soil conditions and land use intensification effects on soil microbial communities across a range of european field sites. Soil Biol. Biochem. 2015, 88, 403–413. [Google Scholar] [CrossRef]
- Hobbie, S.E.; Reich, P.B.; Oleksyn, J.; Ogdahl, M.; Zytkowiak, R.; Hale, C.; Karolewski, P. Tree species effects on decomposition and forest floor dynamics in a common garden. Ecology 2006, 87, 2288–2297. [Google Scholar] [CrossRef] [PubMed]
- Jesus, E.d.C.; Marsh, T.L.; Tiedje, J.M.; Moreira, F.M.d.S. Changes in land use alter the structure of bacterial communities in Western Amazon soils. ISME J. 2009, 3, 1004–1011. [Google Scholar] [CrossRef]
- Vesterdal, L.; Elberling, B.; Christiansen, J.R.; Callesen, I.; Schmidt, I.K. Soil respiration and rates of soil carbon turnover differ among six common European tree species. Forest Ecol. Manag. 2012, 264, 185–196. [Google Scholar] [CrossRef]
- Guo, A.N.; Zhao, Z.Q.; Zhang, P.F.; Yang, Q.; Li, Y.F.; Wang, G.Y. Linkage between soil nutrient and microbial characteristic in an opencast mine, China. Sci. Total Environ. 2019, 671, 905–913. [Google Scholar] [CrossRef]
- Yang, Y.; Gao, Y.; Wang, S.; Xu, D.; Yu, H.; Wu, L.; Lin, Q.; Hu, Y.; Li, X.; He, Z.; et al. The microbial gene diversity along an elevation gradient of the Tibetan grassland. ISME J. 2013, 8, 430–440. [Google Scholar] [CrossRef]
- Liu, Y.B.; Zhao, L.; Zengru Wang, Z.R.; Liu, L.C.; Zhang, P.; Sun, J.Y.; Wang, B.Y.; Song, G.; Li, X.R. Changes in functional gene structure and metabolic potential of the microbial community in biological soil crusts along a revegetation chronosequence in the Tengger Desert. Soil Biol. Biochem. 2018, 126, 40–48. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Soil | Tree Species | Bulk Density (g∙cm−3) | pH | Total N (g∙kg−1) | Total C (g∙kg−1) | Total S (g∙kg−1) |
|---|---|---|---|---|---|---|
| PO | 1.47 ± 0.02 a | 8.17 ± 0.02 a | 0.50 ± 0.18 b | 12.39 ± 2.73 d | 0.64 ± 0.04 c | |
| SC | 1.43 ± 0.03 b | 8.13 ± 0.02 ab | 1.38 ± 0.22 a | 29.91 ± 2.94 a | 1.82 ± 0.15 a | |
| Bulk soil | PS | 1.36 ± 0.01 c | 8.09 ± 0.01 c | 1.18 ± 0.02 a | 25.86 ± 0.44 b | 1.13 ± 0.05 b |
| PA | 1.42 ± 0.02 b | 8.08 ± 0.01 c | 0.80 ± 0.02 b | 17.53 ± 0.83 c | 0.33 ± 0.03 d | |
| PT | 1.41 ± 0.03 b | 8.12 ± 0.02 bc | 0.80 ± 0.02 b | 23.65 ± 0.63 b | 0.86 ± 0.09 c | |
| PO | 1.47 ± 0.02 a | 8.12 ± 0.01 a | 0.62 ± 0.20 b | 13.20 ± 2.20 d | 0.64 ± 0.03 c | |
| SC | 1.43 ± 0.03 b | 8.10 ± 0.02 a | 1.43 ± 0.20 a | 29.85 ± 3.20 a | 1.78 ± 0.10 a | |
| Rhizosphere soil | PS | 1.36 ± 0.01 c | 8.05 ± 0.01 b | 1.22 ± 0.03 a | 26.53 ± 1.23 a | 1.18 ± 0.04 b |
| PA | 1.42 ± 0.02 b | 8.05 ± 0.01 b | 0.88 ± 0.03 b | 17.80 ± 0.72 c | 0.35 ± 0.02 d | |
| PT | 1.41 ± 0.03 b | 8.10 ± 0.02 a | 0.90 ± 0.02 b | 23.56 ± 0.55 b | 0.85 ± 0.06 c | |
| Summary of ANOVA test | ||||||
| Tree species | *** | *** | *** | *** | *** | |
| Soil | ns | *** | *** | *** | *** | |
| Tree species × soil | ns | ns | *** | *** | *** | |
| Soil | Tree Species | PRO μmol∙(d∙g)−1 | URE mg∙(d∙g)−1 | DHA mL∙(h∙g)−1 | ALP μmol∙(d∙g)−1 |
|---|---|---|---|---|---|
| PO | 0.133 ± 0.019 d | 0.175 ± 0.024 d | 1.067 ± 0.146 b | 11.778 ± 1.257 b | |
| SC | 0.193 ± 0.014 c | 0.413 ± 0.0113 b | 0.658 ± 0.341 b | 10.618 ± 0.809 b | |
| Bulk soil | PS | 0.576 ± 0.036 a | 0.382 ± 0.011 bc | 0.667 ± 0.420 b | 20.796 ± 1.019 a |
| PA | 0.039 ± 0.007 e | 1.0565 ± 0.032 a | 1.958 ± 0.260 a | 13.208 ± 0.409 b | |
| PT | 0.406 ± 0.005 b | 0.335 ± 0.008 c | 1.183 ± 0.350 ab | 19.478 ± 1.247 a | |
| PO | 0.107 ± 0.001 c | 0.356 ± 0.0116 d | 1.283 ± 0.458 a | 7.115 ± 0.379 d | |
| SC | 0.051 ± 0.008 d | 0.429 ± 0.011 c | 1.250 ± 0.198 a | 2.454 ± 0.551 e | |
| Rhizosphere soil | PS | 0.175 ± 0.007 b | 0.753 ± 0.015 b | 1.425 ± 0.263 a | 11.070 ± 0.584 c |
| PA | 0.197 ± 0.006 b | 0.934 ± 0.029 a | 1.725 ± 0.152 a | 20.947 ± 1.322 a | |
| PT | 0.271 ± 0.039 a | 0.463 ± 0.008 c | 1.233 ± 0.628 a | 16.732 ± 1.313 b | |
| Summary of ANOVA test | |||||
| Tree species | *** | *** | ** | *** | |
| Soil | *** | *** | * | *** | |
| Tree species × soil | *** | *** | ns | *** | |
| Soil | Tree Species | Chao1 | Shannon | Operational Taxonomic Units (OTUs) |
|---|---|---|---|---|
| PO | 3895.5 ± 24.0 a | 9.49 ± 0.25 b | 3849.0 ± 19.97 b | |
| SC | 3565.1 ± 24.9 b | 9.57 ± 0.02 b | 3594.0 ± 16.82 c | |
| Bulk soil | PS | 3863.9 ± 36.1 a | 10.07 ± 0.28 a | 3939.3 ± 34.85 a |
| PA | 3092.6 ± 23.4 d | 9.25 ± 0.26 b | 3062.3 ± 13.32 e | |
| PT | 3359.3 ± 27.2 c | 9.30 ± 0.11 b | 3326.7 ± 14.29 b | |
| PO | 2893.9 ± 47.0 d | 8.72 ± 0.16 c | 2915.3 ± 22.90 e | |
| SC | 3422.1 ± 43.1 b | 9.34 ± 0.13 b | 3351.7 ± 25.38 c | |
| Rhizosphere soil | PS | 3624.2 ± 21.1 a | 9.89 ± 0.23 a | 3627.0 ± 19.70 a |
| PA | 3587.5 ± 18.2 a | 9.38 ± 0.14 b | 3559.3 ± 35.08 b | |
| PT | 3166.0 ± 28.3 c | 8.99 ± 0.22 c | 3191.0 ± 15.13 d | |
| Summary of ANOVA test | ||||
| Tree species | *** | *** | *** | |
| Soil | *** | *** | *** | |
| Tree species × soil | *** | *** | *** | |
| Soil | Method | Bacterial Community Structure | Functional Genes Structure |
|---|---|---|---|
| Bulk soil | MRPP | δ = 0.475 | δ = 0.115 |
| ANOSIM | R = 0.998 | R = 0.575 | |
| PERMANOVA | R2 = 0.775 | R2 = 0.660 | |
| Rhizosphere soil | MRPP | δ = 0.485 | δ = 0.163 |
| ANOSIM | R = 1.000 | R = 0.947 | |
| PERMANOVA | R2 = 0.788 | R2 = 0.758 |
| Soil | Tree Species | F1 | F2 | F3 | F4 | IFI |
|---|---|---|---|---|---|---|
| PO | −2.017 | −1.082 | −1.063 | −0.569 | −1.137 | |
| SC | 0.400 | −2.058 | 0.515 | 0.153 | −0.265 | |
| Bulk soil | PS | 2.974 | −0.504 | −1.968 | 0.918 | 0.579 |
| PA | −0.604 | 2.898 | 1.011 | 0.833 | 0.749 | |
| PT | 0.799 | −0.187 | −1.235 | −0.481 | −0.042 | |
| PO | −3.380 | −0.270 | −0.400 | 1.380 | −1.090 | |
| SC | 0.120 | 0.190 | −0.220 | −0.230 | 0.030 | |
| Rhizosphere soil | PS | 1.840 | 0.510 | 1.600 | −1.660 | 0.830 |
| PA | 0.580 | 2.550 | −0.170 | −0.370 | 0.730 | |
| PT | −0.550 | −1.910 | 1.750 | −0.570 | −0.390 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liu, S.; He, J.; Ning, Y.; Li, J.; Zhang, H.; Liu, Y. Assessment of Artificial Forest Restoration by Exploring the Microbial Community Structure and Function in a Reclaimed Coal Gob Pile in a Loess Hilly Area of Shanxi, China. Forests 2023, 14, 1888. https://0-doi-org.brum.beds.ac.uk/10.3390/f14091888
Liu S, He J, Ning Y, Li J, Zhang H, Liu Y. Assessment of Artificial Forest Restoration by Exploring the Microbial Community Structure and Function in a Reclaimed Coal Gob Pile in a Loess Hilly Area of Shanxi, China. Forests. 2023; 14(9):1888. https://0-doi-org.brum.beds.ac.uk/10.3390/f14091888
Chicago/Turabian StyleLiu, Shuang, Jiuping He, Yuewei Ning, Junjian Li, Hong Zhang, and Yong Liu. 2023. "Assessment of Artificial Forest Restoration by Exploring the Microbial Community Structure and Function in a Reclaimed Coal Gob Pile in a Loess Hilly Area of Shanxi, China" Forests 14, no. 9: 1888. https://0-doi-org.brum.beds.ac.uk/10.3390/f14091888