
Rapid Warming Exacerbates Winter Drought Stress in Trees at High-Altitude Areas in Northeast China
1
College of Life Sciences and Food Engineering, Inner Mongolia Minzu University, Tongliao 028000, China
2
College of Forestry, Shenyang Agricultural University, Shenyang 110866, China
*
Author to whom correspondence should be addressed.
Forests 2024, 15(3), 565; https://0-doi-org.brum.beds.ac.uk/10.3390/f15030565
Submission received: 20 February 2024
/
Revised: 18 March 2024
/
Accepted: 19 March 2024
/
Published: 20 March 2024
(This article belongs to the Special Issue Response of Tree Rings to Climate Change and Climate Extremes)
Abstract
:Against the background of global warming, trees in high-latitude and high-altitude areas are more sensitive to rapid warming. Revealing the response patterns of trees at different altitudes to rapid warming in typical alpine mountain environments can help to predict the future distribution of forests in the region and the associated changes in the timberline. This study used tree-ring width data (band sampling) from Dahurian larch (Larix gmelinii Rupr.) along an altitudinal gradient (970–1409 m) on Oakley Mountain to establish 10 chronologies and to analyze the growth–climate response of larch to warming and altitudinal changes along a gradient. The results showed that before rapid warming, larch growth was strongly influenced mainly by precipitation in July–August, while after warming, the growth was controlled mainly by precipitation (snowfall) in winter (October–April) and showed a significant positive correlation with the SPEI in winter and a negative correlation with temperature in February–April (early spring). This indicates that the rapid warming event led to a drastic change in the water heat balance during the pre-growth period of the trees in the study area, which shifted the tree growth from being restricted by drought in the growing season before warming to being restricted by winter drought (i.e., the lagged effect of snowfall in the pre-growing season) after warming and that the trees at low altitudes suffered from more severe winter drought. In the future, as the global climate warms further, the growth of mountain larch at low altitudes (below 1200 m) will continue to decrease, and the coupling of winter snowfall and seasonal (February–April) warming will increase the upper altitude limit at which winter drought occurs for larch in the study area; additionally, larch at higher altitudes will be able to cope with this ecological process better.
1. Introduction
The sixth report of the IPCC states that human activities have led to a sustained and rapid increase in global temperatures and a high frequency of multiple extreme events, with irreversible impacts on the development of human societies and the evolution of terrestrial ecosystems [1,2]. Forest ecosystems are important components of terrestrial ecosystems and provide resources and ecological services for human survival and development [3]. Rapid climate change will likely put enormous pressure on forest ecosystems in this century [4].
This rapid warming phenomenon is already having major impacts on forest ecosystems, including changes in tree growth and development and in forest composition and function, especially in some alpine and high-altitude areas [5,6,7,8]. Altitude is an important factor that indirectly affects tree growth in mountain habitats, and several studies have reported that rapid warming may negatively affect trees at lower altitudes but favor tree growth at higher altitudes [9,10]. Reduced water availability due to rapid warming in arid and semiarid zones inhibits tree growth and leads to tree mortality and forest decline [11,12]. In addition, several studies have emphasized the important role of seasonal temperatures at high altitudes and latitudes in determining tree growth [10,13].
Located in the northernmost part of China, the Great Xing’an Mountains is the largest and best-preserved original cold-temperate needle-leaved forest area in China and is also one of the regions experiencing the most drastic climate warming in China [8,13,14]. Numerous studies have shown that the forests of the Great Xing’an Mountains have been experiencing rapid warming and localized extreme drought events over the past half century [14,15,16], and associated dendroecological studies have shown that warming has resulted in significant differences in larch growth at different altitudes [17].
Mountain forests account for a large proportion of the total forest area in the study area, and the high-altitude areas referred to in previous studies on Dahurian larch along the altitudinal gradient are mainly flatlands in the high-altitude zone rather than mountains. This topographic difference greatly affects tree growth in the permafrost zone. For example, during summer in high-altitude flatlands, compared to mountainous terrain, where trees are well watered due to seasonal freezing and thawing of perennial permafrost, trees in mountainous environments are more susceptible to preseason drought constraints [18]. In this study, in order to explore the effect of winter drought on the radial growth of Dahurian larch at different altitudes in Oakley Mountain, a typical peak of the Great Xing’an Range against the background of rapid warming, we collected tree ring samples from 86 Dahurian larch species along an altitudinal gradient (970–1409 m), explored the relationship between growth and climatic factors by means of dendrochronology, and revealed the spatial and temporal variations of the distribution pattern of winter climate-driven tree growth under sudden temperature change. Our objectives were to (1) analyze the variability in the effects of dominant climate factors on the growth of Dahurian larch before and after abrupt temperature changes [19] in the context of global warming and (2) assess the spatial and temporal heterogeneity in the growth response of Dahurian larch to winter drought along an altitudinal gradient. We hypothesize that warming-induced winter drought stress is already a major limiting factor for the radial growth of low-elevation trees in alpine regions and that this stress is exacerbated by rapid warming.
2. Materials and Methods
2.1. Study Area
The study area is located on Oakley Mountain (51°50′ N, 122°02′ E, 1523 m above sea level) in the Great Xing’an Mountains in northeast China, which, as the highest peak in the northern section of the Great Xing’an Mountains, can better reflect the characteristics of the vertical distribution of vegetation in the forested area (Figure 1a). According to the actual survey results and literature records, the distribution of mountain vegetation exhibits significant vertical zonation along the altitudinal gradient [20,21]. The main coniferous or coniferous and broadleaved mixed forest belts (750–1350 m) and subalpine forest belts (1350–1500 m) are included, and the specific tree species from low to high are Larix gmelinii Rupr.—Betula platyphylla Sukaczev—Rhododendron simsii Planch., Larix gmelinii Rupr.—Betula platyphylla Sukaczev—Pinus pumila Pall., Larix gmelinii Rupr.—Betula ermanii Cham.—Pinus pumila Pall., Betula ermanii Cham.—Pinus pumila Pall.—Sabina davurica Pall. and Pinus pumila Pall. pure forests [22]. A variety of extremely drought-tolerant plants (Pinus pumila Pall., Sabina davurica Pall.) occur in the upper elevation coniferous forest belt, which is the main environmental characteristic of the region’s typical high-latitude, high-altitude, alpine, and arid environment. Dahurian larch is the main tree species of zonal vegetation in the northern part of the Great Xing’an Mountains, with a vertical distribution range of approximately 400–1350 m above sea level, and it is the forest species with the largest distribution area and widest range in the region [23,24]. In this study, only Dahurian larch within a continuous altitudinal gradient (970–1409 m) on Oakley Mountain were selected for growth–climate analysis to avoid the influence of microtopography and other factors.
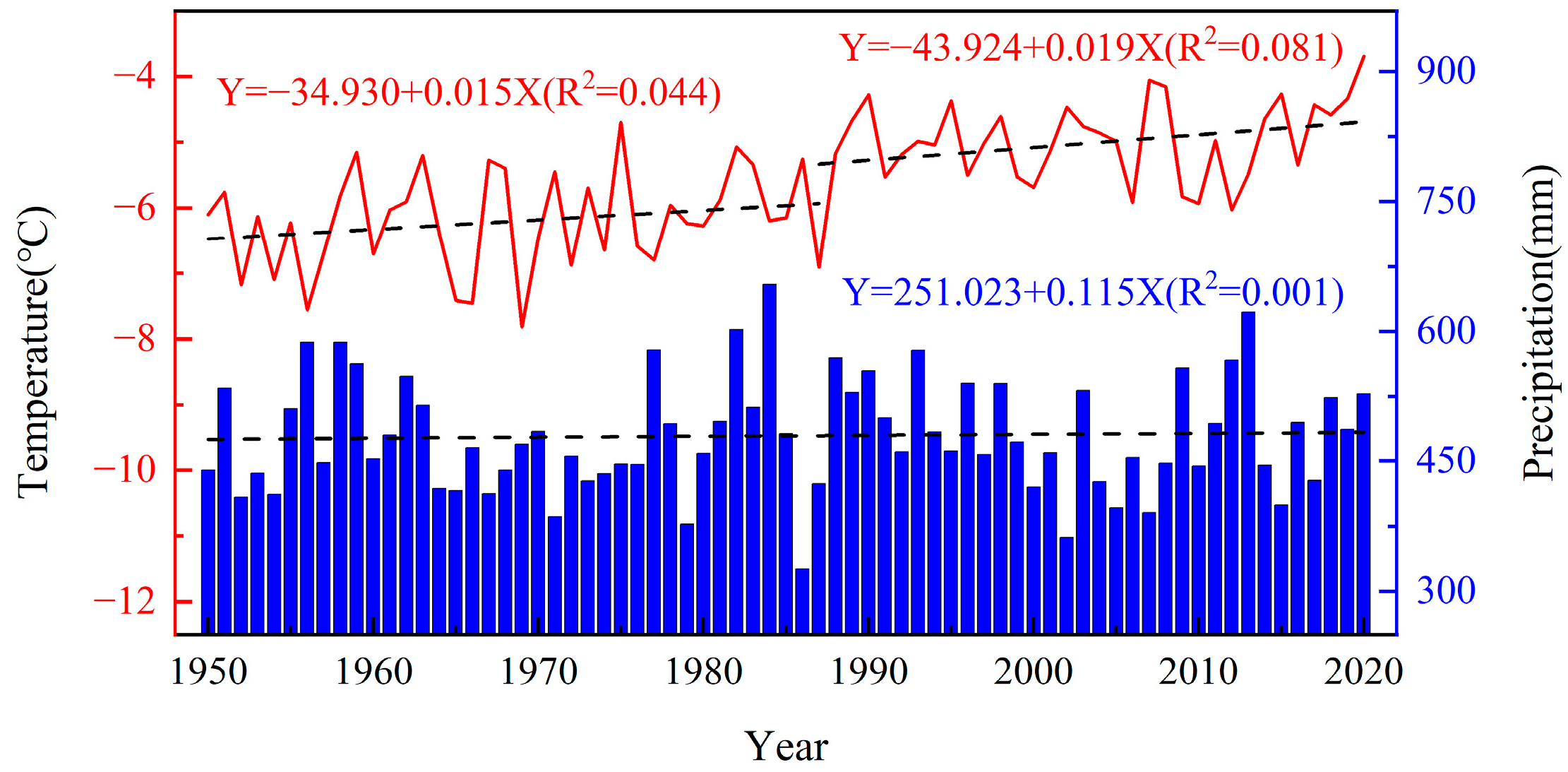
Oakley Mountain has a cold-temperate continental monsoon climate that is cold and dry in winter with a short, hot, and humid summer. The temperature difference between day and night is large, the average annual snow storage period on the summit of the mountain is 240 days, and the frost-free period is short. The mean annual temperature is −5.6 °C (Figure 1e), the average precipitation is 470 mm, and the highest average temperature occurs in July (16.2 °C), while the lowest average temperature occurs in January (−30.5 °C); the precipitation is mainly concentrated in the months of June–September, which accounts for 77% of the total amount of the year. There has been a significant warming trend in the study area since 1950 (Figure 2). The mean annual temperature showed a significant upward trend (0.29 °C·(10a)−1), with a stronger warming trend in winter (October–April) (0.34 °C·(10a)−1), and there was no significant change in annual precipitation.
2.2. Sample Collection and Chronology Development
In this study, we selected Dahurian larch trees, which are characterized by little anthropogenic disturbance and good growth (except near the tree line), for tree sampling along a continuously increasing elevational gradient (970–1409 m) on the southeastern slope of Oakley Mountain. Using an increment borer with an inner diameter of 5.15 mm, 1–4 cores were taken in the vertical direction from a height of approximately 1.3 m per tree. Samples within every 40 m along the elevational gradient were divided into one group during the sampling process, and 10 groups of tree ring samples were collected, totaling 174 sample cores from 86 trees. In this study, to ensure the consistency of the growth of the same group of trees within a certain elevation or slope direction (within 40 m), the sampling did not cross a large vertical or horizontal space. Therefore, the sample size of each chronology group was smaller than that of a conventional study. In addition, at an elevation of 1330 m above the Dahurian larch tree line height, there were fewer tree samples, so the sample data above the tree line height (1330–1409 m) were merged into one group (Table 1). The obtained cores were sealed in custom-made plastic tubes, and the number and sample information of each tree were recorded to facilitate the subsequent processing of the samples. The samples were brought back to the laboratory and fixed in a special wooden trough with white latex. After the samples were completely dried, they were sanded and polished with sandpaper of different meshes from coarse to fine, until the boundaries of the tree ring were clear. Cross-dating was carried out using a microscope, and the width of the tree ring was subsequently measured using the Velmex Tree Ring Analysis System (accuracy of 0.001 mm). The measurement results were verified using the COFECHA program to ensure the accuracy of the dating results [25]. The ARSTAN program was used to address the missing values (gaps) generated by the loss of sample breaks; the negative exponential function and the two-thirds step length spline function were used to detrend for the tree-ring width series and establish a standard chronology of 10 groups of Dahurian larch. We chose the STD chronology as a representative of the tree growth changes along the altitudinal gradient to analyze their climatic responses [26,27].
2.3. Climate Data
Climatic information was obtained from gridded meteorological data (CRU TS 4.06 gridded dataset, resolution 0.5° × 0.5°) using the KNMI Climate Explorer data sharing website (http://climexp.knmi.nl), with selected gridded coordinates ranging from 51.5° to 52.5° N and 121.5° to 122.5° E. The selected climatic data span the period of 1950–2020 and include the mean monthly temperature, mean monthly precipitation, actual vapor pressure, and standardized precipitation evapotranspiration index (SPEI). The relative humidity (RH) is used to analyze the effects of changing hydrothermal conditions on tree growth. In this study, RH data were calculated from actual and saturated vapor pressure data [28], and saturated water vapor pressure was calculated from grid point temperature data [29]. The relevant formulae are as follows:
where RH is the relative humidity, E is the saturated water vapor pressure, e is the actual water vapor pressure, and T is the air temperature.
2.4. Data Analysis
Mutation points in the time series of major climate factors were tested using the Mann–Kendall method [30]. Due to the strong low-frequency variation and autocorrelation of meteorological data series (e.g., temperature), to reduce the influence of trends in chronological and climatic variables on the results of the correlation analyses, Pearson’s correlation coefficients were calculated on first-order differences for both the chronology and the meteorological data. Indeed, this correlation coefficient represents the relationship between original tree growth and climate data in our study. The selected calculation periods are 1950–1986 and 1987–2020, respectively. The single-month meteorological factor was selected from the previous October to the current September and the winter meteorological factor was selected for the combination month of October–April (pOA). The regression analysis was conducted between the correlations of growth and winter climate and altitude, and validated by covariance analysis. The correlation analyses and charting were performed using the packages ‘stats’, ‘ggbiplot’ and ‘ggplot2’ of R4.3.0 software [31], the charting and study area mapping were performed using Origin 2022 and Arcmap v10.2 software.
3. Results
3.1. Winter Climatic Characteristics
Since 1950, the study area has experienced a significant winter warming trend with a rate of 0.34 °C·(10a)−1 and no significant trend in winter precipitation (snow) (Figure 3). The mutation point test analysis of the mean annual temperatures for the years of 1950–2020 showed that there was an abrupt change in the temperature before and after 1987, and the t test results also indicated that there was a significant difference of the temperature data in approximately 1987 (p < 0.05). After the sudden temperature change in 1987, the mean winter temperatures (October–April) for the two periods (1950–1986 and 1987–2020) were −18.27 °C and −16.88 °C, respectively, while the warming during this period was 1.39 °C. The values of the extreme low temperatures around the time of warming were −20.85 °C in 1966 and −19.47 °C in 2001. The mean winter precipitation in the two periods was basically the same (71.59 mm and 72.77 mm), but the precipitation extremes were lower and more frequent in the last 20 years, with the minimum precipitation decreasing from 48.57 mm in 1953 to 40.95 mm in 2008. The overall trend of the winter SPEI is consistent with that of winter precipitation, which has fluctuated greatly over the last 20 years, indicating that rapid warming has been followed by more drastic winter hydrothermal changes in the study area.
3.2. Radial Growth Characteristics
The chronology spans from 141 to 276 years, with the longest dating back to 1745 (Table 1). The main purpose of this study was to explore the processes and patterns of Dahurian larch growth in response to climatic factors under rapid warming; therefore, chronological data from 1950 to 2020 matched with valid meteorological record data were used for the relevant analyses. The mean sensitivity (MS) of the chronology varied between 0.181 and 0.343, the standard deviation varied between 0.223 and 0.507, and the mean signal-to-noise ratio (SNR) of the chronologies was 11.876, indicating that the Dahurian larch chronology contained more climate signals. Samples of KLD10 were located near the Dahurian larch tree line, with smaller sample sizes and younger trees not reaching the public interval of chronological statistics; thus, they had lower overall representative (EPS) value. In the remaining nine chronologies, EPS values were above 0.85. Overall, the chronologies used in this paper met the criteria for conducting dendrochronological statistical analyses of trees.
In this study, we analyzed the radial growth characteristics and segmentation trends of Dahurian larch before and after rapid warming (Figure 4). Eight chronologies along the altitudinal gradient exhibited a slow upward trend with large interannual fluctuations throughout the study period (1950–2020). After suffering a sudden temperature change in the 1980s, the growth of Dahurian larch at low altitudes (below 1200 m) increased rapidly for a short period of time and then showed a decreasing trend (Figure 4), whereas the growth of trees at high altitudes showed no significant changes during this time period. In addition, before the rapid warming in 1987, for the two low altitude areas, the chronologies showed an increasing trend, and for the middle altitude and most high-altitude areas, the chronologies showed a slow decreasing trend; after 1987, approximately 60% of all the chronologies showed a decreasing trend, with the decreasing trend in the low altitude areas being more pronounced and the interannual fluctuations more drastic.
3.3. Growth–Climate Relationships
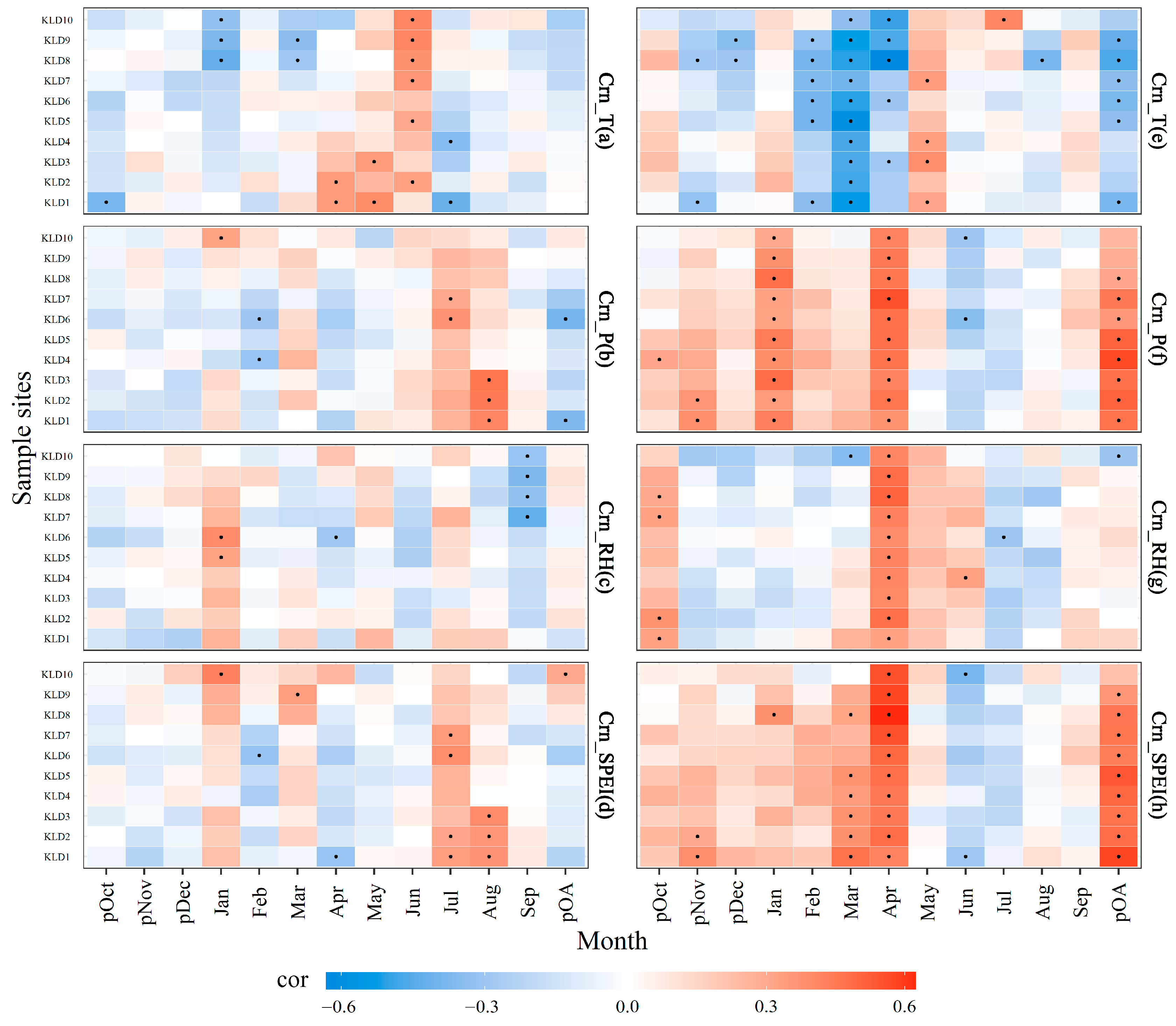
The results of correlation analysis revealed significant differences in the effects of climate factors on the radial growth of Dahurian larch in the study area before and after rapid warming. Before rapid warming, the growth was mainly positively correlated with the mean temperature in April–June, the correlation with temperature in April changed from positive to negative with increasing elevation, and the negative correlation was dominant in the other months (Figure 5a). After rapid warming, the growth was significantly positively correlated with the mean temperature in May and significantly negatively correlated with the mean temperature from February to April, while the correlation with temperature in April changed from negative to significantly negative with increasing elevation (Figure 5e). Before rapid warming (Figure 5b), the growth was mainly positively correlated with summer (July–August) precipitation (especially at low altitudes) and weakly correlated with winter precipitation (snow). After rapid warming, the growth was mainly positively correlated with winter (October–April) precipitation and reached a significant positive correlation with precipitation in January and April. On the contrary, the higher precipitation in summer was detrimental to growth compared with that in the prewarming period (Figure 5f).
Before rapid warming, the RH had no significant effect on the growth during the growing season (Figure 5c). After rapid warming, the growth was mainly negatively correlated with the RH in November–February and reached a significant negative correlation at high altitudes. Additionally, the correlation between growth and RH in March changed from a positive correlation to a negative correlation as the altitude increased. The growth is mainly positively correlated with RH from April to June and reached a significant positive correlation in April (Figure 5g). Before rapid warming, the correlation between growth and SPEI in winter (October–April) shifted from negative to positive with increasing altitude (Figure 5d); after rapid warming, the correlation between growth and SPEI in winter (October–April) was positive (Figure 5h) and became significantly positive in March–April.
Before rapid warming, the winter climate had no significant effect on Dahurian larch growth, and water deficit was the main factor limiting the growth of the trees in summer (July–August), whereas higher temperatures and less snowfall in winter after warming resulted in Dahurian larch growth being significantly affected by winter drought.
3.4. Altitudinal Divergence of Growth–Winter Drought Associations
We conducted covariance analysis and the results showed significant differences in the relationship between growth and climate along the altitudinal gradients before and after rapid warming; the p-values are all less than 0.05. Before rapid warming, the growth of Dahurian larch was predominantly insignificantly negatively correlated with winter (October–April) temperatures and the negative correlation increased with altitude (R2 = 0.930, p < 0.001; Figure 6a). Also before rapid warming, growth and winter precipitation were predominantly negatively correlated and tended to weaken significantly with altitude (R2 = 0.261, p < 0.05); after rapid warming, growth and winter precipitation shifted to a positive correlation across the entire altitudinal gradient, and the correlation tended to increase highly significantly with decreasing altitude (R2 = 0.572, p < 0.0001; Figure 6b). Before and after rapid warming, the correlations between winter RH and Dahurian larch growth showed opposite results across the altitudinal gradient; neither reached significance, but there was an altitudinal trend in these correlations (Figure 6c). Before rapid warming, the correlation between growth and the SPEI shifted from a negative correlation to a positive correlation with increasing altitude (R2 = 0.453, p < 0.0001; Figure 6d); after warming, the correlation became positive across the entire altitudinal gradient, and there was a highly significant altitudinal trend (R2 = 0.763, p < 0.0001). Taken together, these results suggest that under the influence of rapid warming, the radial growth of Dahurian larch is increasingly affected by winter drought in the study area, which was particularly obvious before and after warming at low altitudes.
4. Discussion
4.1. Temporal–Spatial Differences in Radial Growth
In the context of rapid warming, frequent and extreme climatic events can directly or indirectly trigger forest decline and lead to changes in its distribution area [10,32]. Numerous studies have shown that at high latitudes, which are sensitive to warming, there is a general increase in tree growth, while tree growth at low altitudes tends to decrease due to drought limitations [11,17,33]. This finding does not contradict our hypothesis that tree growth in high-altitude mountain habitats is subject to drought stress because the high and low altitudes, as referred to in this study, are relative, and in fact those areas greater than 900 m in the Great Xing’an Mountains are considered high-altitude environments. Since the 1980s, the trees at different altitudes in the study area have exhibited spatial and temporal growth trends (Figure 4), with the growth occurring at low altitudes showing a trend of increasing and then decreasing, while the growth at high altitudes (above 1200 m) has shown a slight decreasing trend. This suggests that low-altitude trees benefited from adequate water heating in the short term during the rapid warming process, whereas at high altitudes, Dahurian larch approached the tree line where low temperatures acted as a major limiting factor, and this rapid warming did not completely alter their long-term acclimatization to a state of water heat equilibrium. In conclusion, the sample sizes at each altitude in this study, although small, still represented the significant variability in tree growth across the altitudinal gradient.
4.2. Temporal–Spatial Differences in Growth–Climate Associations
Before and after rapid warming, the correlations between Dahurian larch growth and major climatic factors changed significantly, especially between precipitation and temperature and between the SPEI in winter (October–April) (Figure 7). This indicates that the hydrothermal conditions of the tree habitats in the study area were strongly affected by rapid warming, which caused a shift in the previously stable growth–climate relationship. Before warming, the winter climate had no significant effect on the growth of Dahurian larch, while summer drought was the main limiting factor for their growth at low altitudes, which decreased with increasing altitude. After warming, Dahurian larch growth was negatively correlated with winter temperature (Figure 7e) and significantly positively correlated with winter precipitation (especially at low altitudes; Figure 7f), indicating that abrupt warming gradually changed Dahurian larch growth at mountainous low altitudes from drought limitation in the growing season in the past to drought limitation in the pre-growing season. This shift is primarily driven by the amount and timing of winter snow storage. For example, less snowpack in winter and earlier snowmelt in spring can lead to insufficient soil available moisture as trees enter the pre-growth period, which is indirectly reflected in the impact of winter drought stress on the radial growth of Dahurian larch in the following year [34,35]. The average winter temperature in the study area reaches −25 °C, and trees are dormant [36]. Thus, the lag effect of winter moisture (snowpack) on trees is primarily related to changes in temperature the following spring.
Before warming, temperature promoted the growth of Dahurian larch in April–June (Figure 5a), which was the period during which the physiological activities of the trees gradually recovered (leaf spread in early June) in the study area. The relatively high temperatures and slower rate of snowmelt in April–May replenish effective soil moisture, and it has been documented that larch has a high percentage of fine roots in shallow organic soils [37,38]. It mainly absorbs shallow soil moisture early in the growing season, and snowmelt replenishes the surface soil water content. [39]. Together, the water retention of the humus layer and the higher temperatures in June triggered rapid activity in the cambium (especially at high altitudes, Figure 5a), promoting tree growth in the current year [40]. After rapid warming, the February–April temperatures significantly inhibited Dahurian larch growth (Figure 5e). February–April is the most significant warming period in the study area, and excessive temperatures and early snowmelt depletion during this period can result in insufficient available soil moisture during the early part of the growing season, causing tree growth to suffer from early spring physiological drought stress [6,16,41]. In particular, the inhibitory effect of temperature on Dahurian larch growth in April gradually diminished with decreasing altitude, which was attributed to the gradual increase in temperature in April, when the soil warmed more quickly at low altitudes than at high altitudes, and when soil conditions unfavorable to the tree root system lessened.
The heterogeneity of increasing warming and decreasing humidity leads to severe water deficit and inhibits tree growth. Before warming, the growth was positively correlated with precipitation and the SPEI in July and August (Figure 5b,d) and was more significant at low altitudes. After warming, the growth was positively correlated with winter precipitation and the winter SPEI (Figure 5f,h), while it was positively correlated with relative humidity only in April–June (Figure 5g). This suggests that increased evapotranspiration of water during the pre-growing season further exacerbates winter drought stress caused by soil moisture deficit. In addition, the location of the sampling environment on the southeastern leeward slopes, coupled with the rapid warming process during the winter season, exacerbated the effects of winter (early spring) drought on the growth of trees at low altitudes [42,43]. However, this drought effect gradually weakened with altitude (Figure 7f,h) because water evapotranspiration decreases rapidly with the increase in altitude, and trees are less susceptible to water stress at high altitudes than at low altitudes [9]. This variability was also confirmed by the negative correlation between Dahurian larch growth at high altitudes and relative humidity from November to March.
4.3. Future Winter Warming-Drying Trends and Dahurian larch Distribution
Major changes in temperature and precipitation patterns occur under global warming [2]. The study area is located at a high latitude and high altitude and its forest ecosystems are highly sensitive to climate change [24,44,45]. During the past 70 years, dramatic climate change in the study area has led to a shift in the correlation between tree growth and climate, with winter precipitation (snowfall) becoming the main limiting factor for Dahurian larch growth. The drought effect of rapid climate warming on tree growth in the study area increased across the altitudinal gradient, but the magnitude of the effect tended to decrease significantly with altitude (Figure 6); i.e., trees in high-altitude environments were more hydrothermally stable in response to this rapid warming (Figure 7). Compared to those at low altitudes, the chronologies at high altitudes experienced no significant change in radial growth and were less affected by warming, which differed from the results of other studies that showed an increase in the growth of larch at high altitudes [14,16,17]. This is, first and foremost, because the present study site is a high-altitude mountainous environment (unlike the high-altitude flatland tundra) where tree growth is limited by a combination of heat and water; it is also because some of the samples in the high-altitude area are close to the timberline (tree line), and their microhabitats have a greater impact on the trees (e.g., infertile soils and greater wind speeds, etc.). In summary, the sensitivity of tree growth to drought will increase under continued warming in the future, with trees at low altitudes facing more severe challenges and trees at high altitudes being better able to balance the relationship between water and heat and cope with short-term warming events.
5. Conclusions
Our study was designed to explore the growth trends of Dahurian larch and climate response patterns with altitude under rapid warming conditions. In the 1980s, rapid warming led to an increase and then a decrease in the growth of Dahurian larch at low altitudes but had a smaller effect on growth at high altitudes (above 1200 m). The response of larch forests to climate has shifted from drought limitations in the growing season before warming to winter drought (the lag effects of snowfall before the next growing season) limitations after warming, especially for low-altitude trees. As the climate continues to warm, the coupled effects of winter snowpack and rapid warming will drive an increase in the upper altitude limit at which winter drought occurs in larch on the studied altitudinal gradient, which are currently better able to cope with this ecological change in higher-altitude environments. In conclusion, rapid warming increases the risk of forest decline at the lower altitude, and in the future forest management and strategy formulation, we should focus on the growth dynamics of dominant conifer species at low and medium altitudes.
Author Contributions
R.Z. and X.B. conceived and designed the study; R.Z. contributed to data measurement and wrote the manuscript; Field work was completed by X.B., Z.C. and R.Z.; X.T., H.Z. and H.L. contributed to investigation, writing and data analyses. All authors have read and agreed to the published version of the manuscript.
Funding
This study was funded by the National Natural Science Foundation of China (grant number 32001136), and partly supported by the Doctoral Research Start-up Funds for the Inner Mongolia Minzu University (KYQD19072).
Data Availability Statement
The raw/processed data required to reproduce these findings cannot be shared at this time as the data also form part of an ongoing study.
Acknowledgments
We would like to thank the Laboratory of Dendrochronology, College of Forestry, Shenyang Agricultural University, for providing the experimental space, Zhenju Chen, Junxia Li, and Yuting Jin for helping with my experiments.
Conflicts of Interest
The authors declare that they have no known competing financial interests or personal relationships that could have appeared to influence the work reported in this paper.
References
- IPCC. Climate Change 2021: The Physical Science Basis. Contribution of Working Group I to the Sixth Assessment Report of the Intergovernmental Panel on Climate Change; Cambridge Press: Cambridge, UK, 2021. [Google Scholar]
- IPCC. Climate Change 2022: Impacts, Adaptation, and Vulnerability. Contribution of Working Group II to the Sixth Assessment Report of the Intergovernmental Panel on Climate Change; Cambridge Press: Cambridge, UK, 2022. [Google Scholar]
- Anderegg, W.R.L.; Trugman, A.T.; Badgley, G.; Anderson, C.M.; Bartuska, A.; Ciais, P.; Cullenward, D.; Field, C.B.; Freeman, J.; Goetz, S.J.; et al. Climate-driven risks to the climate mitigation potential of forests. Science 2020, 368, eaaz7005. [Google Scholar] [CrossRef]
- Anderegg, W.R.L.; Wu, C.; Acil, N.; Carvalhais, N.; Pugh, T.A.M.; Sadler, J.P.; Seidl, R. A climate risk analysis of Earth’s forests in the 21st century. Science 2022, 377, 1099–1103. [Google Scholar] [CrossRef]
- Du, H.; Liu, J.; Li, M.H.; Buntgen, U.; Yang, Y.; Wang, L.; Wu, Z.; He, H.S. Warming-induced upward migration of the alpine treeline in the Changbai Mountains, northeast China. Glob. Chang. Biol. 2018, 24, 1256–1266. [Google Scholar] [CrossRef] [PubMed]
- Gaire, N.P.; Fan, Z.X.; Bräuning, A.; Panthi, S.; Rana, P.; Shrestha, A.; Bhuju, D.R. Abies spectabilis shows stable growth relations to temperature, but changing response to moisture conditions along an elevation gradient in the central Himalaya. Dendrochronologia 2020, 60, 125675. [Google Scholar] [CrossRef]
- Shi, H.; Zhou, Q.; Xie, F.; He, N.; He, R.; Zhang, K.; Zhang, Q.; Dang, H. Disparity in elevational shifts of upper species limits in response to recent climate warming in the Qinling Mountains, North-central China. Sci. Total Environ. 2020, 706, 135718. [Google Scholar] [CrossRef]
- Zheng, L.; Gaire, N.P.; Shi, P.; Schöb, C. High-altitude tree growth responses to climate change across the Hindu Kush Himalaya. J. Plant Ecol. 2021, 14, 829–842. [Google Scholar] [CrossRef]
- Yuan, S.; Jiang, Y.; Zhao, Z.; Cui, M.; Shi, D.; Wang, S.; Kang, M. Different trends and divergent responses to climate factors in the radial growth of Abies georgei along elevations in the central Hengduan Mountains. Dendrochronologia 2023, 80, 126114. [Google Scholar] [CrossRef]
- Panthi, S.; Fan, Z.X.; van der Sleen, P.; Zuidema, P.A. Long-term physiological and growth responses of Himalayan fir to environmental change are mediated by mean climate. Glob. Chang. Biol. 2020, 26, 1778–1794. [Google Scholar] [CrossRef]
- Jiao, L.; Jiang, Y.; Zhang, W.T.; Wang, M.C.; Zhang, L.N.; Zhao, S.D. Divergent responses to climate factors in the radial growth of Larix sibirica in the eastern Tianshan Mountains, northwest China. Trees 2015, 29, 1673–1686. [Google Scholar] [CrossRef]
- Lyu, L.; Zhang, Q.B.; Pellatt, M.G.; Büntgen, U.; Li, M.H.; Cherubini, P. Drought limitation on tree growth at the Northern Hemisphere’s highest tree line. Dendrochronologia 2019, 53, 40–47. [Google Scholar] [CrossRef]
- Gao, J.; Liu, Y. Climate warming and land use change in Heilongjiang Province, Northeast China. Appl. Geogr. 2011, 31, 476–482. [Google Scholar] [CrossRef]
- Yasmeen, S.; Wang, X.; Zhao, H.; Zhu, L.; Yuan, D.; Li, Z.; Zhang, Y.; Ahmad, S.; Han, S. Contrasting climate-growth relationship between Larix gmelinii and Pinus sylvestris var. mongolica along a latitudinal gradient in Daxing’an Mountains, China. Dendrochronologia 2019, 58, 125645. [Google Scholar] [CrossRef]
- Zhang, X.; Liu, X.; Zhang, Q.; Zeng, X.; Xu, G.; Wu, G.; Wang, W. Species-specific tree growth and intrinsic water-use efficiency of Dahurian larch (Larix gmelinii) and Mongolian pine (Pinus sylvestris var. mongolica) growing in a boreal permafrost region of the Greater Hinggan Mountains, Northeastern China. Agric. For. Meteorol. 2018, 248, 145–155. [Google Scholar] [CrossRef]
- Zhang, X.; Bai, X.; Hou, M.; Chen, Z.; Manzanedo, R.D. Warmer Winter Ground Temperatures Trigger Rapid Growth of Dahurian Larch in the Permafrost Forests of Northeast China. J. Geophys. Res. Biogeosci. 2019, 124, 1088–1097. [Google Scholar] [CrossRef]
- Bai, X.P.; Zhang, X.L.; Li, J.X.; Duan, X.Y.; Jin, Y.T.; Chen, Z.J. Altitudinal disparity in growth of Dahurian larch (Larix gmelinii Rupr.) in response to recent climate change in northeast China. Sci. Total Environ. 2019, 670, 466–477. [Google Scholar] [CrossRef]
- Li, Y.; Liu, H.; Zhu, X.; Yue, Y.; Xue, J.; Shi, L. How permafrost degradation threatens boreal forest growth on its southern margin? Sci. Total Environ. 2021, 762, 143154. [Google Scholar] [CrossRef]
- Pithan, F.; Mauritsen, T. Arctic amplification dominated by temperature feedbacks in contemporary climate models. Nat. Geosci. 2014, 7, 181–184. [Google Scholar] [CrossRef]
- Gao, M.; Meng, F. On the delineation of forest vegetation zones in the Oakleigh Pile Mountains. Inn. Mong. For. Investig. Des. 1999, 4, 141–143. [Google Scholar]
- Zhang, L. The Research on the East Slope of Middle Region of Daxing’an Mountains Vertical Zone. Master’s Thesis, Northeast Normal University, Changchun, China, 2007. [Google Scholar]
- Huang, X.; Zhang, X.; Yuan, L. Vegetation Types and Vertical Distribution Patterns of Oakley Mountain. High. Educ. Forum. 2009. Available online: https://www.docin.com/p-236712927.html (accessed on 20 February 2024).
- Yang, Z.; Zhou, G.; Yin, X.; Jia, B. Geographic distribution of Larix gmelinii natural forest in China and its climatic suitability. Chin. J. Ecol. 2014, 33, 1429–1436. [Google Scholar] [CrossRef]
- Zhang, H.; Zhang, S.; Song, W.; Tigabu, M.; Fu, M.; Xue, H.; Sun, A.; Zhao, M.; Cai, K.; Li, Y.; et al. Climate response of radial growth and early selection of Larix olgensis at four trials in northeast China. Dendrochronologia 2022, 73, 125955. [Google Scholar] [CrossRef]
- Holmes, R.L. Computer-Assisted Quality Control in Tree-Ring Dating and Measurement. Tree-Ring Bull. 1983, 43, 51–67. [Google Scholar]
- Cook, E.R. A Time Series Approach to Tree-Ring Standardization. Ph.D. Thesis, The University of Arizona, Tucson, AZ, USA, 1985. [Google Scholar]
- Holmes, R.L.; Cook, E.R. Guide for Computer Program ARSTAN; Laboratory of Tree-Ring Research, University of Arizona: Tucson, AZ, USA, 1994. [Google Scholar]
- Peixoto, J.; Oort, A.H. The Climatology of Relative Humidity in the Atmosphere. J. Clim. 1996, 9, 3443–3463. [Google Scholar] [CrossRef]
- Liu, J.; Guo, H.; Li, Y.; Liu, H.; Wu, B. Fundamentals of Physical Quantity Calculation in Weather Analysis and Forecasting; China Meteorological Press: Beijing, China, 2005. [Google Scholar]
- Kendall, M.G. Rank Correlation Methods. Biometrika 1957, 44, 146–163. [Google Scholar] [CrossRef]
- Team R Core. R: A language and environment for statistical computing. MSOR Connect. 2014, 1, 12–21. [Google Scholar]
- Hepting, G.H. Climate and Forest diseases. Annu. Rev. Phytopathol. 1963, 60, 133–149. [Google Scholar] [CrossRef]
- Huang, R.; Zhu, H.; Liu, X.; Liang, E.; Grießinger, J.; Wu, G.; Li, X.; Bräuning, A. Does increasing intrinsic water use efficiency (iWUE) stimulate tree growth at natural alpine timberline on the southeastern Tibetan Plateau? Glob. Planet. Chang. 2017, 148, 217–226. [Google Scholar] [CrossRef]
- Su, J.; Gou, X.; Deng, Y.; Zhang, R.; Liu, W.; Zhang, F.; Lu, M.; Chen, Y.; Zheng, W. Tree growth response of Fokienia hodginsii to recent climate warming and drought in southwest China. Int. J. Biometeorol. 2017, 61, 2085–2096. [Google Scholar] [CrossRef] [PubMed]
- Bogachev, M.I.; Gafurov, A.M.; Iskandirov, P.Y.; Kaplun, D.I.; Kayumov, A.R.; Lyanova, A.I.; Pyko, N.S.; Pyko, S.A.; Safonova, A.N.; Sinitca, A.M.; et al. Reversal in the drought stress response of the Scots pine forest ecosystem: Local soil water regime as a key to improving climate change resilience. Heliyon 2023, 9, e21574. [Google Scholar] [CrossRef]
- Freeland, R.O. Apparent Photosynthesis in Some Conifers During Winter. Plant Physiol. 1944, 19, 179–185. [Google Scholar] [CrossRef]
- Iversen, C.M.; Childs, J.; Norby, R.J.; Ontl, T.A.; Kolka, R.K.; Brice, D.J.; McFarlane, K.J.; Hanson, P.J. Fine-root growth in a forested bog is seasonally dynamic, but shallowly distributed in nutrient-poor peat. Plant Soil 2017, 424, 123–143. [Google Scholar] [CrossRef]
- Zhang, Y.P.; Jiang, Y.; Wang, B.; Jiao, L.; Wang, M.c. Seasonal water use by Larix principis-rupprechtii in an alpine habitat. For. Ecol. Manag. 2018, 409, 47–55. [Google Scholar] [CrossRef]
- Sugimoto, A.; Naito, D.; Yanagisawa, N.; Ichiyanagi, K.; Kurita, N.; Kubota, J.; Kotake, T.; Ohata, T.; Maximov, T.C.; Fedorov, A.N. Characteristics of soil moisture in permafrost observed in East Siberian taiga with stable isotopes of water. Hydrol. Process. 2002, 17, 1073–1092. [Google Scholar] [CrossRef]
- Pompa García, M.; Cerano Paredes, J.; Fulé, P.Z. Variation in radial growth of Pinus cooperi in response to climatic signals across an elevational gradient. Dendrochronologia 2013, 31, 198–204. [Google Scholar] [CrossRef]
- Ren, P.; Rossi, S.; Camarero, J.J.; Ellison, A.M.; Liang, E.; Penuelas, J. Critical temperature and precipitation thresholds for the onset of xylogenesis of Juniperus przewalskii in a semi-arid area of the north-eastern Tibetan Plateau. Ann. Bot. 2018, 121, 617–624. [Google Scholar] [CrossRef] [PubMed]
- Qiao, J.; Sun, Y. Effects of altitude and slope on the climate–radial growth relationships of Larix olgensis A. Henry in the southern Lesser Khingan Mountains, Northeast China. Ecol. Process. 2022, 11, 46. [Google Scholar] [CrossRef]
- Li, J.; Li, S.; Liu, C.; Guo, D.; Zhang, Q. Response of Chinese pine regeneration density to forest gap and slope aspect in northern China: A meta-analysis. Sci. Total Environ. 2023, 873, 162428. [Google Scholar] [CrossRef] [PubMed]
- Gaire, N.P.; Zaw, Z.; Bräuning, A.; Grießinger, J.; Sharma, B.; Rana, P.; Bhandari, S.; Basnet, S.; Fan, Z.-X. The impact of warming climate on Himalayan silver fir growth along an elevation gradient in the Mt. Everest region. Agric. For. Meteorol. 2023, 339, 109575. [Google Scholar] [CrossRef]
- Zhang, W.; Gou, X.; Zhang, F.; Liu, W.; Zhang, Y.; Gao, L. Divergent responses of Qinghai spruce (Picea crassifolia) to recent warming along elevational gradients in the central Qilian Mountains, Northwest China. J. Geogr. Sci. 2023, 33, 151–168. [Google Scholar] [CrossRef]
Figure 1.
Geographical locations of the samples in the study area (a–d) and climatic characteristics of the monthly values for 1950–2020 (e).
Figure 1.
Geographical locations of the samples in the study area (a–d) and climatic characteristics of the monthly values for 1950–2020 (e).

Figure 2.
Interannual variation of the mean annual temperature (line) and annual precipitation (bar) in the study area from 1950 to 2020. Dotted lines represent the trend of the corresponding series.
Figure 2.
Interannual variation of the mean annual temperature (line) and annual precipitation (bar) in the study area from 1950 to 2020. Dotted lines represent the trend of the corresponding series.

Figure 3.
Winter climate trends in the study area (1950–2020, red line). The boxplots show the distributional differences in the values of the corresponding climate factors before and after rapid warming. (a) Temperature; (b) precipitation; (c) standardized precipitation evapotranspiration index (SPEI). The red shaded areas indicate 95% confidence interval.
Figure 3.
Winter climate trends in the study area (1950–2020, red line). The boxplots show the distributional differences in the values of the corresponding climate factors before and after rapid warming. (a) Temperature; (b) precipitation; (c) standardized precipitation evapotranspiration index (SPEI). The red shaded areas indicate 95% confidence interval.

Figure 4.
Changes in the tree ring width index along the altitudinal gradient from 1950 to 2020. The blue line represents the growth trend from 1950 to 1986, the red line represents the growth trend from 1987 to 2020, and gray represents the standard deviation. The altitude of the sampling site (m above sea level) is given at the top right of each subplot.
Figure 4.
Changes in the tree ring width index along the altitudinal gradient from 1950 to 2020. The blue line represents the growth trend from 1950 to 1986, the red line represents the growth trend from 1987 to 2020, and gray represents the standard deviation. The altitude of the sampling site (m above sea level) is given at the top right of each subplot.

Figure 5.
Correlations between chronologies and climatic factors at different altitudes for the periods of 1950–1986 (a–d) and 1987–2020 (e–h). t, p, RH, and SPEI represent the temperature, precipitation, relative humidity, and standardized evapotranspiration index (SPEI), respectively; lowercase p represents the previous year; pOA represents the period from October of the previous year to April of the current year; blue indicates a negative correlation, and red indicates a positive correlation; black dots represent correlation coefficients reaching the 95% confidence level; and vertical coordinate sampling points are arranged from bottom to top according to the increase in elevation.
Figure 5.
Correlations between chronologies and climatic factors at different altitudes for the periods of 1950–1986 (a–d) and 1987–2020 (e–h). t, p, RH, and SPEI represent the temperature, precipitation, relative humidity, and standardized evapotranspiration index (SPEI), respectively; lowercase p represents the previous year; pOA represents the period from October of the previous year to April of the current year; blue indicates a negative correlation, and red indicates a positive correlation; black dots represent correlation coefficients reaching the 95% confidence level; and vertical coordinate sampling points are arranged from bottom to top according to the increase in elevation.

Figure 6.
Change in correlations, between growth and winter T (a), P (b), RH (c), and SPEI (d), with altitude for the periods of 1950–1986 (blue) and 1987–2020 (red), respectively. The shaded areas indicate 95% confidence interval.
Figure 6.
Change in correlations, between growth and winter T (a), P (b), RH (c), and SPEI (d), with altitude for the periods of 1950–1986 (blue) and 1987–2020 (red), respectively. The shaded areas indicate 95% confidence interval.

Figure 7.
The correlation-scales between growth and climate along the altitudinal gradient. (a,e), (b,f), (c,g), (d,h) represent the winter T, P, RH, and SPEI, respectively. (a–d) (1950–1986) and (e–h) (1987–2020) represent before and after warming, respectively. Correlations range from −0.6 (blue) to 0.6 (red).
Figure 7.
The correlation-scales between growth and climate along the altitudinal gradient. (a,e), (b,f), (c,g), (d,h) represent the winter T, P, RH, and SPEI, respectively. (a–d) (1950–1986) and (e–h) (1987–2020) represent before and after warming, respectively. Correlations range from −0.6 (blue) to 0.6 (red).


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
Statistical characteristics of the tree ring width chronologies.
| Altitudinal Classes | Site ID | Time Span | Trees (Cores) | MS | SD | EPS | SNR |
|---|---|---|---|---|---|---|---|
| 970–1009 | KLD1 | 1880–2021 | 7 (14) | 0.267 | 0.507 | 0.970 | 31.907 |
| 1010–1049 | KLD2 | 1849–2021 | 13 (26) | 0.181 | 0.312 | 0.934 | 14.205 |
| 1050–1089 | KLD3 | 1849–2021 | 8 (16) | 0.231 | 0.416 | 0.859 | 6.112 |
| 1090–1129 | KLD4 | 1845–2021 | 13 (26) | 0.194 | 0.366 | 0.920 | 11.491 |
| 1130–1169 | KLD5 | 1873–2021 | 8 (16) | 0.343 | 0.223 | 0.851 | 5.695 |
| 1170–1209 | KLD6 | 1880–2021 | 5 (11) | 0.252 | 0.377 | 0.914 | 10.644 |
| 1210–1249 | KLD7 | 1794–2021 | 10 (20) | 0.258 | 0.439 | 0.925 | 12.346 |
| 1250–1289 | KLD8 | 1745–2021 | 12 (24) | 0.220 | 0.282 | 0.928 | 12.795 |
| 1290–1329 | KLD9 | 1811–2021 | 8 (16) | 0.251 | 0.314 | 0.920 | 11.484 |
| 1330–1409 | KLD10 | 1769–2021 | 3 (5) | 0.292 | 0.270 | 0.675 | 2.073 |
MS: mean sensitivity; SD: standard deviation; EPS: expressed population signal; SNR: signal-to-noise ratio.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Zhang, R.; Bai, X.; Tian, X.; Chen, Z.; Zhang, H.; Liu, H. Rapid Warming Exacerbates Winter Drought Stress in Trees at High-Altitude Areas in Northeast China. Forests 2024, 15, 565. https://0-doi-org.brum.beds.ac.uk/10.3390/f15030565
AMA Style
Zhang R, Bai X, Tian X, Chen Z, Zhang H, Liu H. Rapid Warming Exacerbates Winter Drought Stress in Trees at High-Altitude Areas in Northeast China. Forests. 2024; 15(3):565. https://0-doi-org.brum.beds.ac.uk/10.3390/f15030565
Chicago/Turabian StyleZhang, Rui, Xueping Bai, Xun Tian, Zhenju Chen, Haiyue Zhang, and Haiting Liu. 2024. "Rapid Warming Exacerbates Winter Drought Stress in Trees at High-Altitude Areas in Northeast China" Forests 15, no. 3: 565. https://0-doi-org.brum.beds.ac.uk/10.3390/f15030565
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.