
Quantifying the Impact of Different Ways to Delimit Study Areas on the Assessment of Species Diversity of an Urban Forest


Abstract
:1. Introduction
2. Materials and Methods
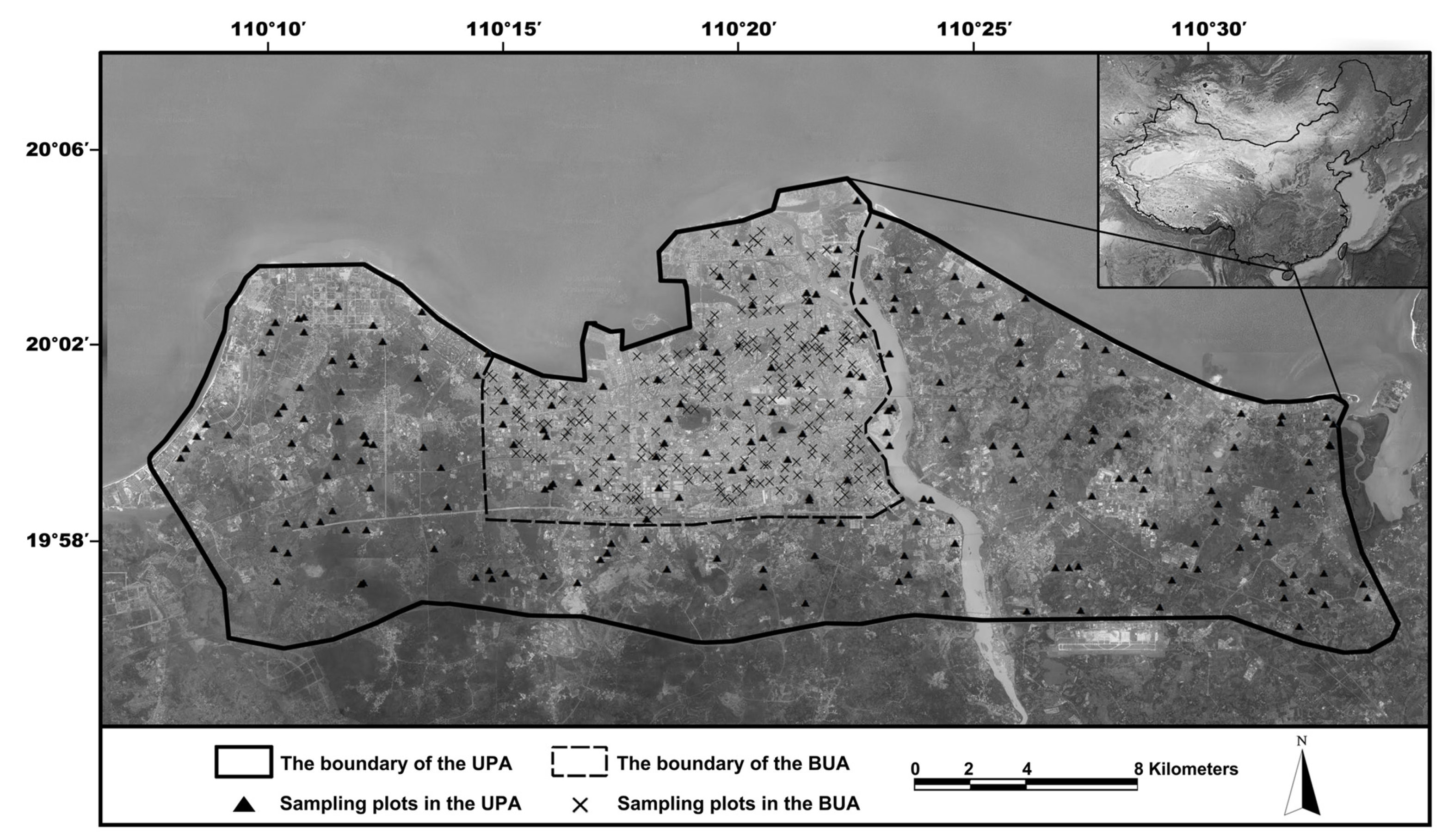
2.1. Study Area
2.2. Field Surveys

2.3. Data Analysis
3. Results
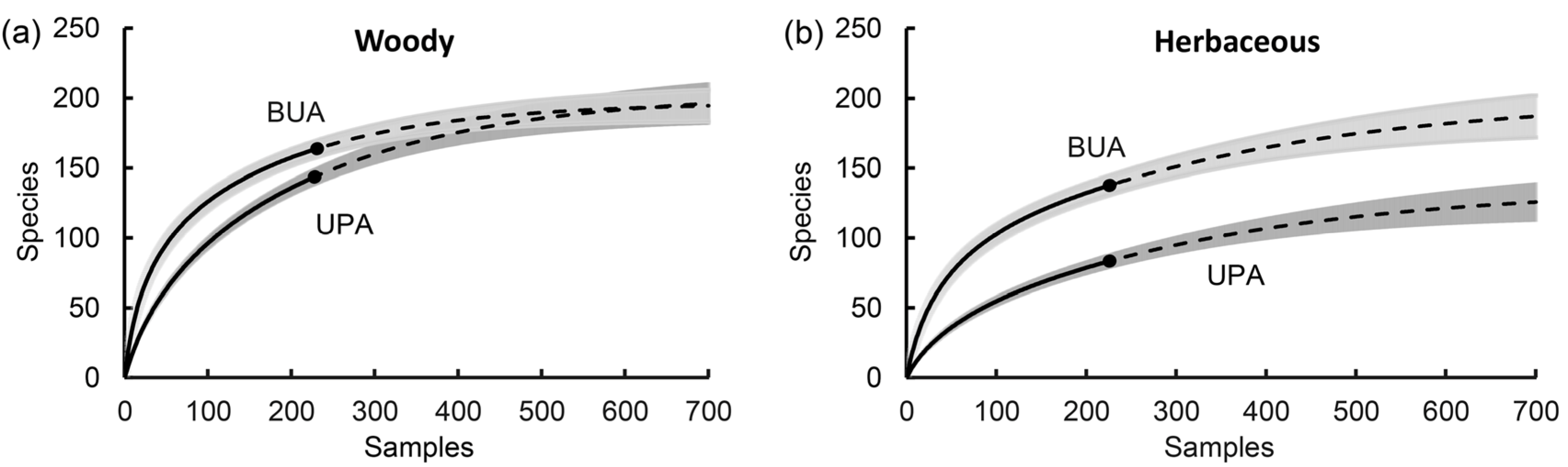
3.1. Species Richness in the Two Areas


{kind=link}
{kind=link}
{kind=link}
| Land Use | Description |
|---|---|
| Agricultural land | All lands used primarily for the production of food and fiber and associated structures |
| Commercial, institutional or industrial lands | Areas that contain structures predominantly used for sale of products and services and areas occupied by light or heavy industry |
| Public green spaces | Green spaces that are maintained by a government agency, such as parks and golf courses |
| Residential areas | Densely populated urban zones containing single or multiple dwelling units |
| Transportation areas | Areas that contain transportation routes and facilities |
| Transitional areas | Lands for which future land use has not been realized, e.g., areas that are under construction for unknown use, vacant lands, abandoned agricultural land |
| Woodlands | Areas predominately covered by woody vegetation |
| Land Use a | All Plants | Woody | Herbaceous | ||||||
|---|---|---|---|---|---|---|---|---|---|
| UPA | BUA | Combined | UPA | BUA | Combined | UPA | BUA | Combined | |
| AGR | 58 | 0 | 58 | 21 | 0 | 21 | 37 | 0 | 37 |
| CII | 29 | 195 | 201 | 22 | 108 | 112 | 7 | 87 | 89 |
| PGS | 36 | 154 | 162 | 27 | 91 | 98 | 9 | 63 | 64 |
| RES | 75 | 199 | 222 | 53 | 117 | 131 | 22 | 82 | 91 |
| TRA | 59 | 104 | 122 | 40 | 54 | 69 | 19 | 50 | 53 |
| TRS | 78 | 73 | 121 | 45 | 21 | 55 | 33 | 52 | 66 |
| WOO | 103 | 0 | 103 | 72 | 0 | 72 | 31 | 0 | 31 |
| All land uses | 228 | 303 | 393 | 144 | 164 | 224 | 84 | 139 | 169 |
| Categories | All Plants | Woody Plants | Herbaceous Plants |
|---|---|---|---|
| Entire study area | p < 0.001 | p < 0.001 | p < 0.001 |
| Origin | |||
| Native species | p < 0.001 | p < 0.001 | p < 0.001 |
| Exotic species | p < 0.001 | p < 0.001 | p < 0.001 |
| Land use a | |||
| CII | p < 0.001 | p < 0.001 | p < 0.001 |
| PGS | p = 0.020 | p = 0.122 | p = 0.004 |
| RES | p < 0.001 | p < 0.001 | p < 0.001 |
| TRA | p = 0.001 | p = 0.049 | p = 0.008 |
| TRS | p = 0.001 | p = 0.962 | p < 0.001 |

| Rarefaction | Extrapolation | Sampling Units Prediction | |||||
|---|---|---|---|---|---|---|---|
| t | SE | t* | SE | ℊ | |||
| (a) BUA-Woody, Sobs = 164, T = 232 | |||||||
| 1 | 5.37 | 0.60 | 0 | 164.00 | 5.23 | 0.86 | 32.15 |
| 50 | 95.73 | 4.88 | 100 | 178.14 | 6.24 | 0.90 | 92.36 |
| 100 | 126.20 | 4.99 | 200 | 186.20 | 7.77 | 0.94 | 184.09 |
| 150 | 144.56 | 4.99 | 300 | 190.79 | 9.28 | 0.98 | 383.90 |
| 200 | 157.51 | 5.08 | 400 | 193.41 | 10.49 | ||
| 232 | 164.00 | 5.23 | 468 | 194.51 | 11.12 | ||
| (b) UPA-Woody, Sobs = 144, T = 229 | |||||||
| 1 | 2.19 | 0.26 | 0 | 144.00 | 6.50 | 0.86 | 170.73 |
| 50 | 63.58 | 4.12 | 100 | 165.17 | 7.99 | 0.90 | 248.46 |
| 100 | 96.38 | 5.08 | 200 | 178.84 | 10.03 | 0.94 | 367.18 |
| 150 | 118.96 | 5.66 | 300 | 187.67 | 12.19 | 0.98 | 628.43 |
| 200 | 135.90 | 6.17 | 400 | 193.36 | 14.13 | ||
| 229 | 144.00 | 6.50 | 471 | 196.13 | 15.31 | ||
| (c) BUA-Herbaceous, Sobs = 139, T = 232 | |||||||
| 1 | 3.52 | 0.46 | 0 | 139.00 | 6.55 | 0.86 | 249.74 |
| 50 | 75.43 | 4.92 | 100 | 155.95 | 7.78 | 0.90 | 356.27 |
| 100 | 102.98 | 5.61 | 200 | 168.26 | 9.57 | 0.94 | 519.00 |
| 150 | 119.86 | 5.94 | 300 | 177.20 | 11.65 | 0.98 | 877.46 |
| 200 | 132.31 | 6.28 | 400 | 183.70 | 13.75 | ||
| 232 | 139.00 | 6.55 | 468 | 187.07 | 15.11 | ||
| (d) UPA-Herbaceous, Sobs = 84, T = 229 | |||||||
| 1 | 1.38 | 0.27 | 0 | 84.00 | 5.70 | 0.86 | 313.05 |
| 50 | 36.2 | 3.38 | 100 | 98.86 | 7.08 | 0.90 | 417.00 |
| 100 | 54.86 | 4.26 | 200 | 109.57 | 8.87 | 0.94 | 576.07 |
| 150 | 68.12 | 4.83 | 300 | 117.28 | 10.87 | 0.98 | 928.85 |
| 200 | 78.69 | 5.37 | 400 | 122.82 | 12.85 | ||
| 229 | 84.00 | 5.70 | 471 | 125.79 | 14.16 | ||
3.2. Difference in Species Composition
| Area | Species | Origin a | Life Form | Freq. |
|---|---|---|---|---|
| UPA | Melia azedarach L. | N | Tree | 30 |
| Casuarina equisetifolia L. | E | Tree | 27 | |
| Pterocarpus indicus Willd. | E | Tree | 21 | |
| Lantana camara L. | E | Shrub | 19 | |
| Ficus hispida L.f. | N | Shrub | 18 | |
| Cocos nucifera L. | N | Tree | 18 | |
| Eucalyptus robusta Sm. | E | Tree | 13 | |
| Carica papaya L. | E | Tree | 12 | |
| Pandanus tectorius Parkinson ex Du Roi | N | Shrub | 12 | |
| Psidium guajava L. | E | Tree | 11 | |
| BUA | Pterocarpus indicus Willd. | E | Tree | 78 |
| Ficus microcarpa “GoldenLeaves” | E | Shrub | 68 | |
| Ixora chinensis Lam. | E | Shrub | 66 | |
| Cocos nucifera L. | N | Tree | 52 | |
| Hibiscus rosa-sinensis L. | E | Shrub | 44 | |
| Ficus benjamina L. | N | Tree | 39 | |
| Ficus microcarpa L.f. | N | Tree | 36 | |
| Duranta repens “Variegata” | E | Shrub | 34 | |
| Plumeria rubra “Acutifolia” | E | Tree | 33 | |
| Roystonea regia (Kunth) O.F.Cook | E | Tree | 32 |
| Categories | All Plants | Woody Plants | Herbaceous Plants |
|---|---|---|---|
| Entire study area | 0.395 | 0.417 | 0.357 |
| Origin | |||
| Native species | 0.540 | 0.464 | 0.463 |
| Exotic species | 0.252 | 0.229 | 0.289 |
| Land use a | |||
| CII | 0.207 | 0.182 | 0.286 |
| PGS | 0.222 | 0.259 | 0.111 |
| RES | 0.307 | 0.264 | 0.409 |
| TRA | 0.305 | 0.375 | 0.158 |
| TRS | 0.589 | 0.476 | 0.424 |
4. Discussion
4.1. Influences of the Delimitations of Study Areas on Quantifying Species Richness
4.2. Influences of the Delimitations of Study Areas on Comparing Species Compositions
4.3. Implications for Urban Forestry Studies
5. Conclusions
Supplementary Materials
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Department of Economic and Social Affairs. World Urbanization Prospects: The 2014 Revision, Highlights (ST/ESA/SER.A/352); the United Nations: New York, NY, USA, 2014. [Google Scholar]
- Seto, K.C.; Güneralp, B.; Hutyra, L.R. Global forecasts of urban expansion to 2030 and direct impacts on biodiversity and carbon pools. Proc. Nat. Acad. Sci. USA 2012, 109, 16083–16088. [Google Scholar] [CrossRef] [PubMed]
- Dearborn, D.C.; Kark, S. Motivations for conserving urban biodiversity. Conserv. Biol. 2010, 24, 432–440. [Google Scholar] [CrossRef] [PubMed]
- Alvey, A.A. Promoting and preserving biodiversity in the urban forest. Urban For. Urban Green. 2006, 5, 195–201. [Google Scholar] [CrossRef]
- Goddard, M.A.; Dougill, A.J.; Benton, T.G. Scaling up from gardens: Biodiversity conservation in urban environments. Trends Ecol. Evol. 2010, 25, 90–98. [Google Scholar] [CrossRef] [PubMed]
- Miller, R.W. Urban Forestry: Planning and Managing Urban Greenspaces; Prentice Hall: Upper Saddle River, NJ, USA, 1988. [Google Scholar]
- Sanders, R.A.; Stevens, J.C. Urban forest of Dayton, Ohio: A preliminary assessment. Urban Ecol. 1984, 8, 91–98. [Google Scholar] [CrossRef]
- Pauleit, S.; Jones, N.; Garcia-Martin, G.; Garcia-Valdecantos, J.L.; Rivière, L.M.; Vidal-Beaudet, L.; Bodson, M.; Randrup, T.B. Tree establishment practice in towns and cities—Results from a European survey. Urban For. Urban Green. 2002, 1, 83–96. [Google Scholar] [CrossRef]
- Nowak, D.J. Urban biodiversity and climate change. In Urban Biodiversity and Design; Müller, N., Werner, P., Kelcy, J.G., Eds.; Wiley-Blacwell: Hoboken, NJ, USA, 2010; Volume 5, pp. 101–117. [Google Scholar]
- Nowak, D.J. Contrasting natural regeneration and tree planting in fourteen North American cities. Urban For. Urban Green. 2012, 11, 374–382. [Google Scholar] [CrossRef]
- Yang, J.; McBride, J.; Zhou, J.; Sun, Z. The urban forest in Beijing and its role in air pollution reduction. Urban For. Urban Green. 2005, 3, 65–78. [Google Scholar] [CrossRef]
- Kendal, D.; Williams, N.S.; Williams, K.J. A cultivated environment: Exploring the global distribution of plants in gardens, parks and streetscapes. Urban Ecosyst. 2012, 15, 637–652. [Google Scholar] [CrossRef]
- McKinney, M.L. Effects of urbanization on species richness: A review of plants and animals. Urban Ecosyst. 2008, 11, 161–176. [Google Scholar] [CrossRef]
- Pyšek, P. Factors affecting the diversity of flora and vegetation in central European settlements. Vegetatio 1993, 106, 89–100. [Google Scholar] [CrossRef]
- Nowak, D.J.; Walton, J.T.; Stevens, J.C.; Crane, D.E.; Hoehn, R.E. Effect of plot and sample size on timing and precision of urban forest assessments. Arboric. Urban For. 2008, 34, 386–390. [Google Scholar]
- Alvarez, I.A.; del Nero Velasco, G.; Barbin, H.S.; Lima, A.; do Couto, H.T.Z. Comparison of two sampling methods for estimating urban tree density. J. Arboric. 2005, 31, 209. [Google Scholar]
- Tait, C.J.; Daniels, C.B.; Hill, R.S. Changes in species assemblages within the Adelaide Metropolitan Area, Australia, 1836–2002. Ecol. Appl. 2005, 15, 346–359. [Google Scholar] [CrossRef]
- McIntyre, N.E.; Knowles-Yánez, K.; Hope, D. Urban ecology as an interdisciplinary field: Differences in the use of “urban” between the social and natural sciences. Urban Ecosyst. 2000, 4, 5–24. [Google Scholar] [CrossRef]
- Wu, J. Urban ecology and sustainability: The state-of-the-science and future directions. Landsc. Urban Plan. 2014, 125, 209–221. [Google Scholar] [CrossRef]
- Hainan Bureau of Statistics. Hainan Statistics Yearbook 2014; China Statistics Press: Beijing, China, 2014.
- Xing, F.; Zhou, J.; Wang, F.; Zeng, Q.; Yi, Q.; Liu, D. Inventory of Plant Species Diversity of Hainan; Huazhong University of Science & Technology Press: Wuhan, China, 2012. [Google Scholar]
- Kendal, D.; Williams, N.S.G.; Williams, K.J.H. Drivers of diversity and tree cover in gardens, parks and streetscapes in an Australian city. Urban For. Urban Green. 2012, 11, 257–265. [Google Scholar] [CrossRef]
- Milović, M.; Mitić, B. The urban flora of the city of Zadar (Dalmatia, Croatia). Nat. Croat. 2012, 21, 65–100. [Google Scholar]
- Wang, G.; Zuo, J.; Li, X.; Liu, Y.; Yu, J.; Shao, H.; Li, Y. Low plant diversity and floristic homogenization in fast-urbanizing towns in Shandong Peninsular, China: Effects of urban greening at regional scale for ecological engineering. Ecol. Eng. 2014, 64, 179–185. [Google Scholar] [CrossRef]
- Zhao, J.; Ouyang, Z.; Zheng, H.; Zhou, W.; Wang, X.; Xu, W.; Ni, Y. Plant species composition in green spaces within the built-up areas of Beijing, China. Plant. Ecol. 2010, 209, 189–204. [Google Scholar] [CrossRef]
- Guangdong Institute of Botany. Flora of Hainan; Science Press: Beijing, China, 1974. [Google Scholar]
- South China Botanical Garden. Flora of Guangdong; Guangdong Science and Technology Press: Guangzhou, China, 2009. [Google Scholar]
- Ricotta, C.; Pavoine, S.; Bacaro, G.; Acosta, A.T. Functional rarefaction for species abundance data. Methods Ecol. Evol. 2012, 3, 519–525. [Google Scholar] [CrossRef]
- Colwell, R.K.; Chao, A.; Gotelli, N.J.; Lin, S.Y.; Mao, C.X.; Chazdon, R.L.; Longino, J.T. Models and estimators linking individual-based and sample-based rarefaction, extrapolation and comparison of assemblages. J. Plant. Ecol. 2012, 5, 3–21. [Google Scholar] [CrossRef]
- EstimateS, version 9.1.0. Robert K. Colwell: Storrs, CT, USA, 2014. Available online: http://purl.oclc.org/estimates (accessed on 11 May 2015).
- Baselga, A. Partitioning the turnover and nestedness components of beta diversity. Glob. Ecol. Biogeogr. 2010, 19, 134–143. [Google Scholar] [CrossRef]
- Hill, J.L.; Curran, P.J.; Foody, G.M. The effect of sampling on the species-area curve. Glob. Ecol. Biogeogr. Lett. 1994, 97–106. [Google Scholar] [CrossRef]
- Hope, D.; Gries, C.; Zhu, W.; Fagan, W.F.; Redman, C.L.; Grimm, N.B.; Nelson, A.L.; Martin, C.; Kinzig, A. Socioeconomics drive urban plant diversity. Proc. Nat. Acad. Sci. USA 2003, 100, 8788–8792. [Google Scholar] [CrossRef] [PubMed]
- Kowarik, I.; Lippe, M.; Cierjacks, A. Prevalence of alien versus native species of woody plants in Berlin differs between habitats and at different scales. Preslia 2013, 85, 113–132. [Google Scholar]
- Walker, J.S.; Grimm, N.B.; Briggs, J.M.; Gries, C.; Dugan, L. Effects of urbanization on plant species diversity in central Arizona. Front. Ecol. Environ. 2009, 7, 465–470. [Google Scholar] [CrossRef]
- Karp, D.S.; Rominger, A.J.; Zook, J.; Ranganathan, J.; Ehrlich, P.R.; Daily, G.C. Intensive agriculture erodes β-diversity at large scales. Ecol. Lett. 2012, 15, 963–970. [Google Scholar] [CrossRef] [PubMed]
- Jim, C.Y.; Chen, W.Y. Pattern and divergence of tree communities in Taipei’s main urban green spaces. Landsc. Urban Plan. 2008, 84, 312–323. [Google Scholar] [CrossRef]
- Hobbs, E.R. Species richness of urban forest patches and implications for urban landscape diversity. Landsc. Ecol. 1988, 1, 141–152. [Google Scholar] [CrossRef]
- Trentanovi, G.; Lippe, M.; Sitzia, T.; Ziechmann, U.; Kowarik, I.; Cierjacks, A. Biotic homogenization at the community scale: Disentangling the roles of urbanization and plant invasion. Divers. Distrib. 2013, 19, 738–748. [Google Scholar] [CrossRef]
- Williams, N.S.G.; Schwartz, M.W.; Vesk, P.A.; McCarthy, M.A.; Hahs, A.K.; Clemants, S.E.; Corlett, R.T.; Duncan, R.P.; Norton, B.A.; Thompson, K.; et al. A conceptual framework for predicting the effects of urban environments on floras. J. Ecol. 2009, 97, 4–9. [Google Scholar] [CrossRef]
- Doody, B.J.; Sullivan, J.J.; Meurk, C.D.; Stewart, G.H.; Perkins, H.C. Urban realities: The contribution of residential gardens to the conservation of urban forest remnants. Biodivers. Conserv. 2010, 19, 1385–1400. [Google Scholar] [CrossRef]
- Deutschewitz, K.; Lausch, A.; Kühn, I.; Klotz, S. Native and alien plant species richness in relation to spatial heterogeneity on a regional scale in Germany. Global Ecol. Biogeogr. 2003, 12, 299–311. [Google Scholar] [CrossRef]
- Ricotta, C.; Godefroid, S.; Rocchini, D. Patterns of native and exotic species richness in the urban flora of Brussels: Rejecting the “rich get richer” model. Biol. Invasions 2010, 12, 233–240. [Google Scholar] [CrossRef]
- Bourne, K.S.; Conway, T.M. The influence of land use type and municipal context on urban tree species diversity. Urban Ecosyst. 2013, 17, 329–348. [Google Scholar] [CrossRef]
- Lamichhane, D.; Thapa, H.B. Participatory urban forestry in Nepal: Gaps and ways forward. Urban For. Urban Green. 2012, 11, 105–111. [Google Scholar] [CrossRef]
- Yang, J.; Zhou, J.; Ke, Y.; Xiao, J. Assessing the structure and stability of street trees in Lhasa, China. Urban For. Urban Green. 2012, 11, 432–438. [Google Scholar] [CrossRef]
- Francis, R.A.; Chadwick, M.A. What makes a species synurbic? Appl. Geogr. 2012, 32, 514–521. [Google Scholar] [CrossRef]
- Duncan, R.P.; Clemants, S.E.; Corlett, R.T.; Hahs, A.K.; McCarthy, M.A.; McDonnell, M.J.; Schwartz, M.W.; Thompson, K.; Vesk, P.A.; Williams, N.S.G. Plant traits and extinction in urban areas: A meta-analysis of 11 cities. Global Ecol. Biogeogr. 2011, 20, 509–519. [Google Scholar] [CrossRef]
- Kunick, W. Woody vegetation in settlements. Landscape Urban Plan. 1987, 14, 57–78. [Google Scholar] [CrossRef]
- Sitzia, T.; Campagnaro, T.; Weir, R.G. Novel woodland patches in a small historical Mediterranean city: Padova, northern Italy. Urban Ecosyst. 2015. [Google Scholar] [CrossRef]
- Yang, J.; Huang, C.; Zhang, Z.; Wang, L. The temporal trend of urban green coverage in major Chinese cities between 1990 and 2010. Urban For. Urban Green. 2014, 13, 19–27. [Google Scholar] [CrossRef]
- Firbank, L.G.; Petit, S.; Smart, S.; Blain, A.; Fuller, R.J. Assessing the impacts of agricultural intensification on biodiversity: A British perspective. Phil. Trans. Roy. Soc. B Biol. Sci. 2008, 363, 777–787. [Google Scholar] [CrossRef] [PubMed]
- Gotelli, N.J.; Colwell, R.K. Quantifying biodiversity: Procedures and pitfalls in the measurement and comparison of species richness. Ecol. Lett. 2001, 4, 379–391. [Google Scholar] [CrossRef]
- Strayer, D.L.; Power, M.E.; Fagan, W.F.; Pickett, S.T.; Belnap, J. A classification of ecological boundaries. BioScience 2003, 53, 723–729. [Google Scholar] [CrossRef]
- USDA Forest Service. i-Tree Eco User’s Manual. Available online: http://www.itreetools.org/resources/manuals/i-Tree%20Eco%20Users%20Manual.pdf (accessed on 31 January 2015).
- Godefroid, S.; Koedam, N. How important are large vs. small forest remnants for the conservation of the woodland flora in an urban context? Global Ecol. Biogeogr. 2003, 12, 287–298. [Google Scholar] [CrossRef]
© 2016 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons by Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
He, R.; Yang, J.; Song, X. Quantifying the Impact of Different Ways to Delimit Study Areas on the Assessment of Species Diversity of an Urban Forest. Forests 2016, 7, 42. https://0-doi-org.brum.beds.ac.uk/10.3390/f7020042
He R, Yang J, Song X. Quantifying the Impact of Different Ways to Delimit Study Areas on the Assessment of Species Diversity of an Urban Forest. Forests. 2016; 7(2):42. https://0-doi-org.brum.beds.ac.uk/10.3390/f7020042
Chicago/Turabian StyleHe, Rongxiao, Jun Yang, and Xiqiang Song. 2016. "Quantifying the Impact of Different Ways to Delimit Study Areas on the Assessment of Species Diversity of an Urban Forest" Forests 7, no. 2: 42. https://0-doi-org.brum.beds.ac.uk/10.3390/f7020042