
Changes of Heart Rate and Lipid Composition in Mytilus Edulis and Modiolus Modiolus Caused by Crude Oil Pollution and Low Salinity Effects


Abstract
:1. Introduction
2. Materials and Methods
2.1. Mollusc Sampling, Maintenance, and Acclimation
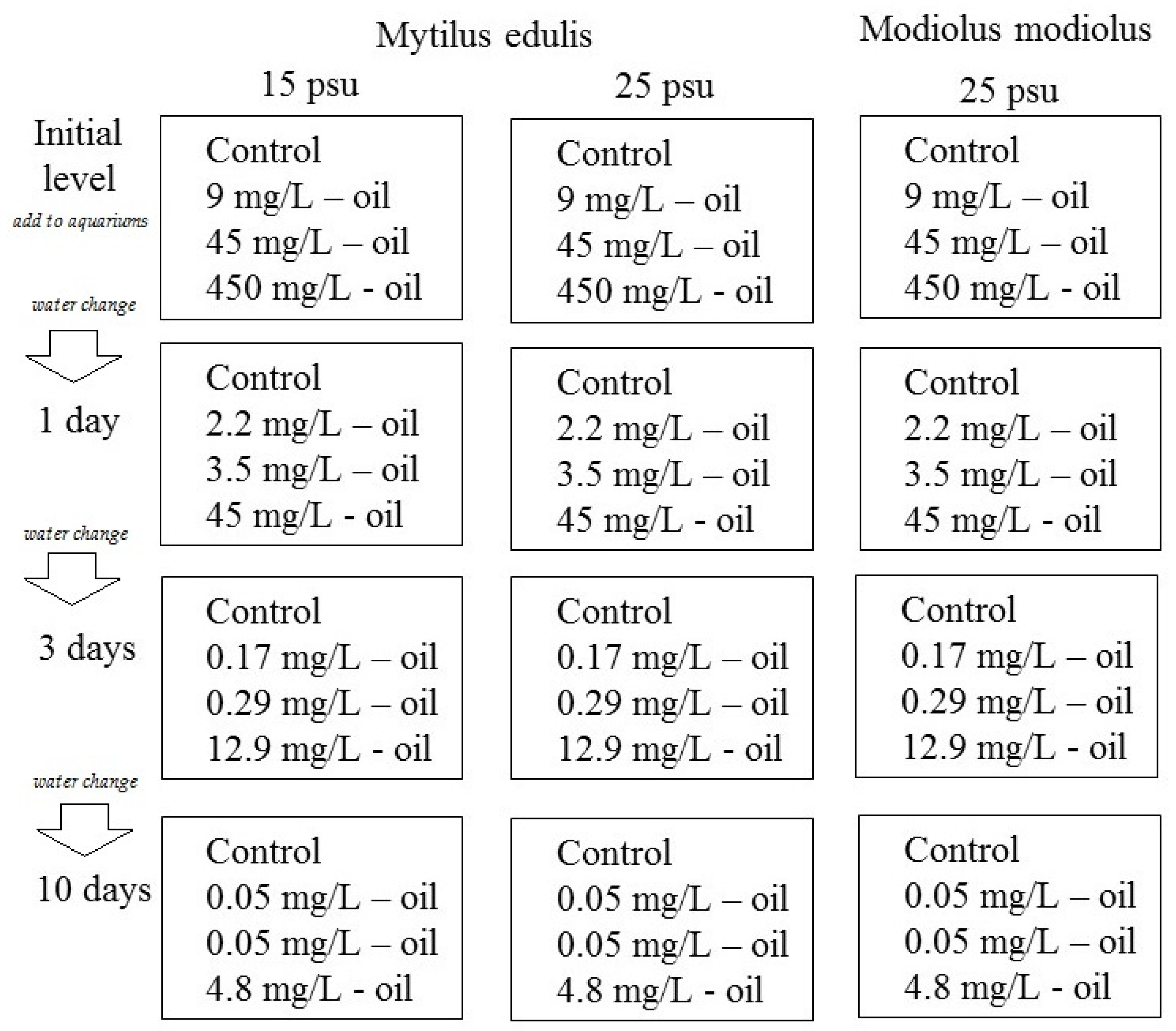
2.2. Crude Oil Exposure Experiment
2.3. Heart Rate Monitoring and Data Processing
2.4. Biochemical Analyses
2.5. Statistical Analysis
3. Results
3.1. Total Hydrocarbon Content (THC) in Aquarium Water
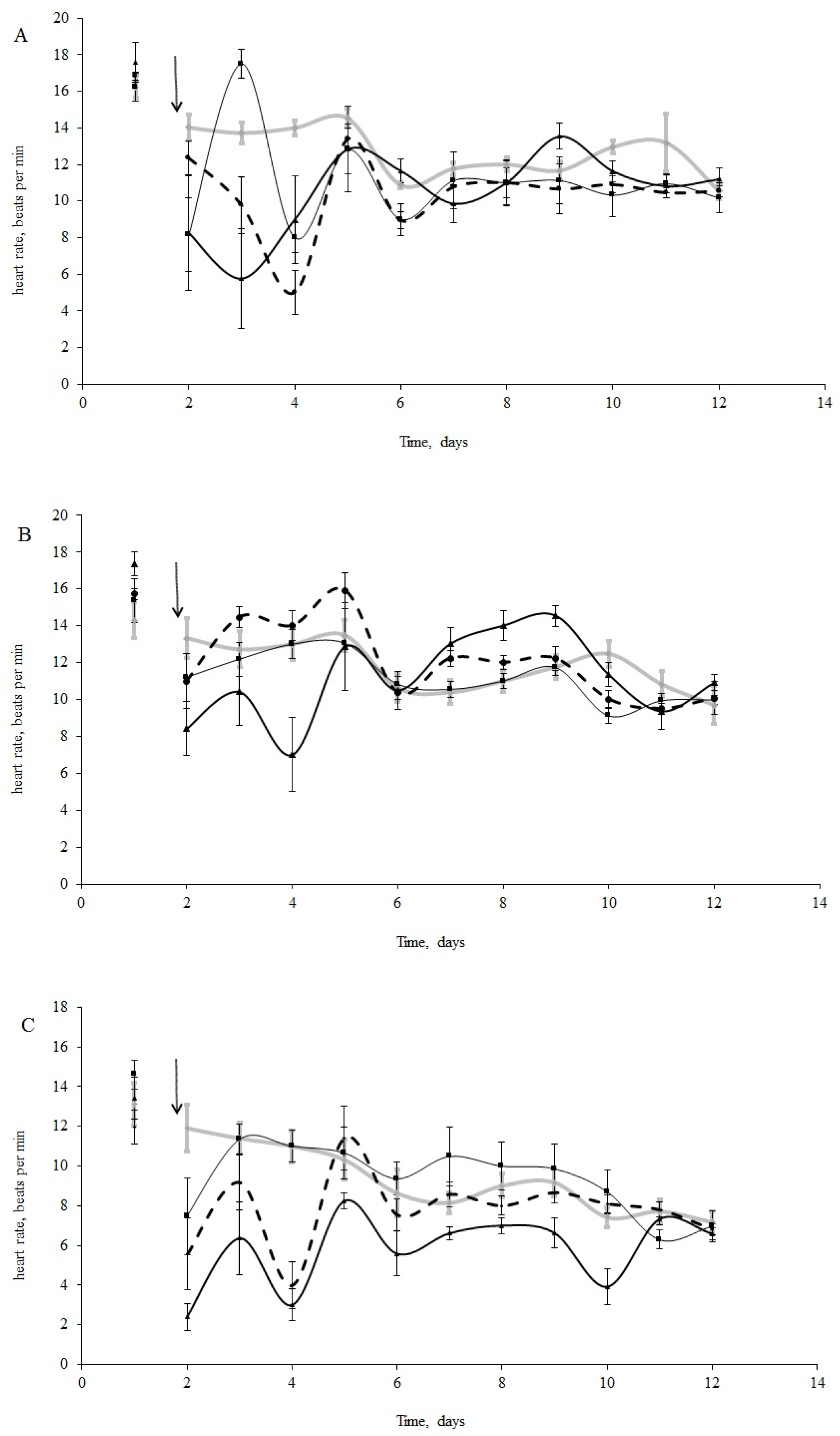
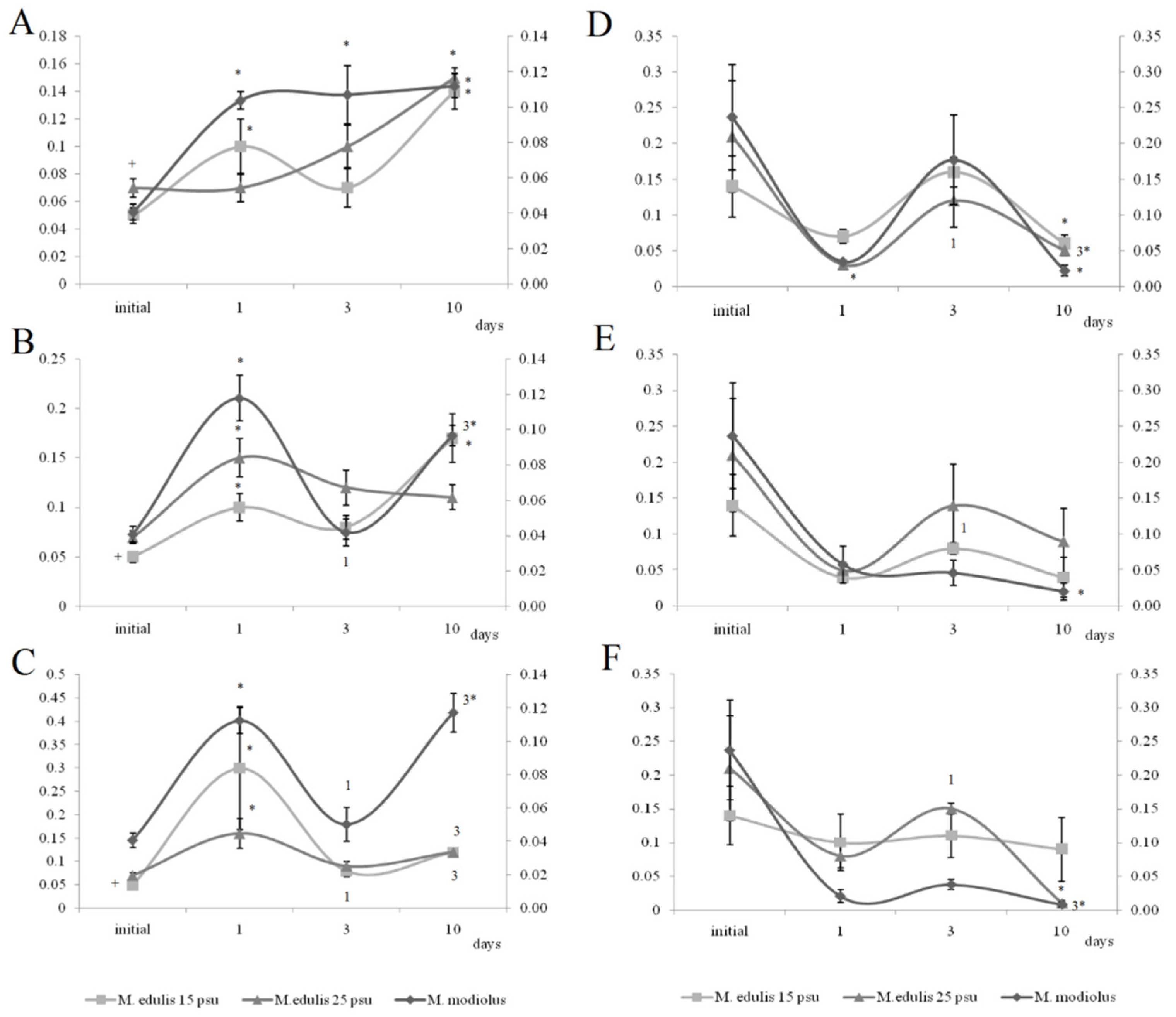
3.2. Physiological Indices
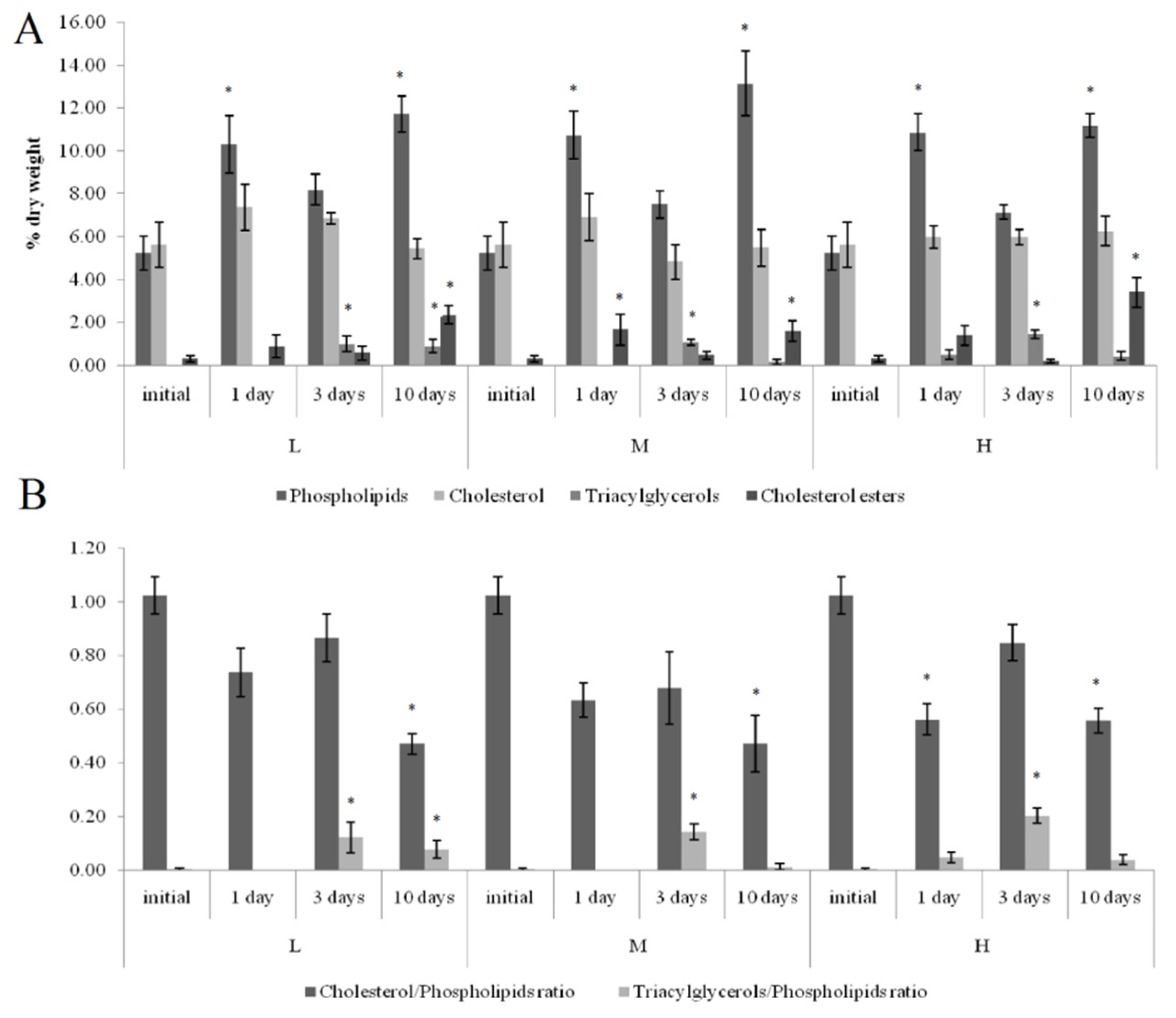
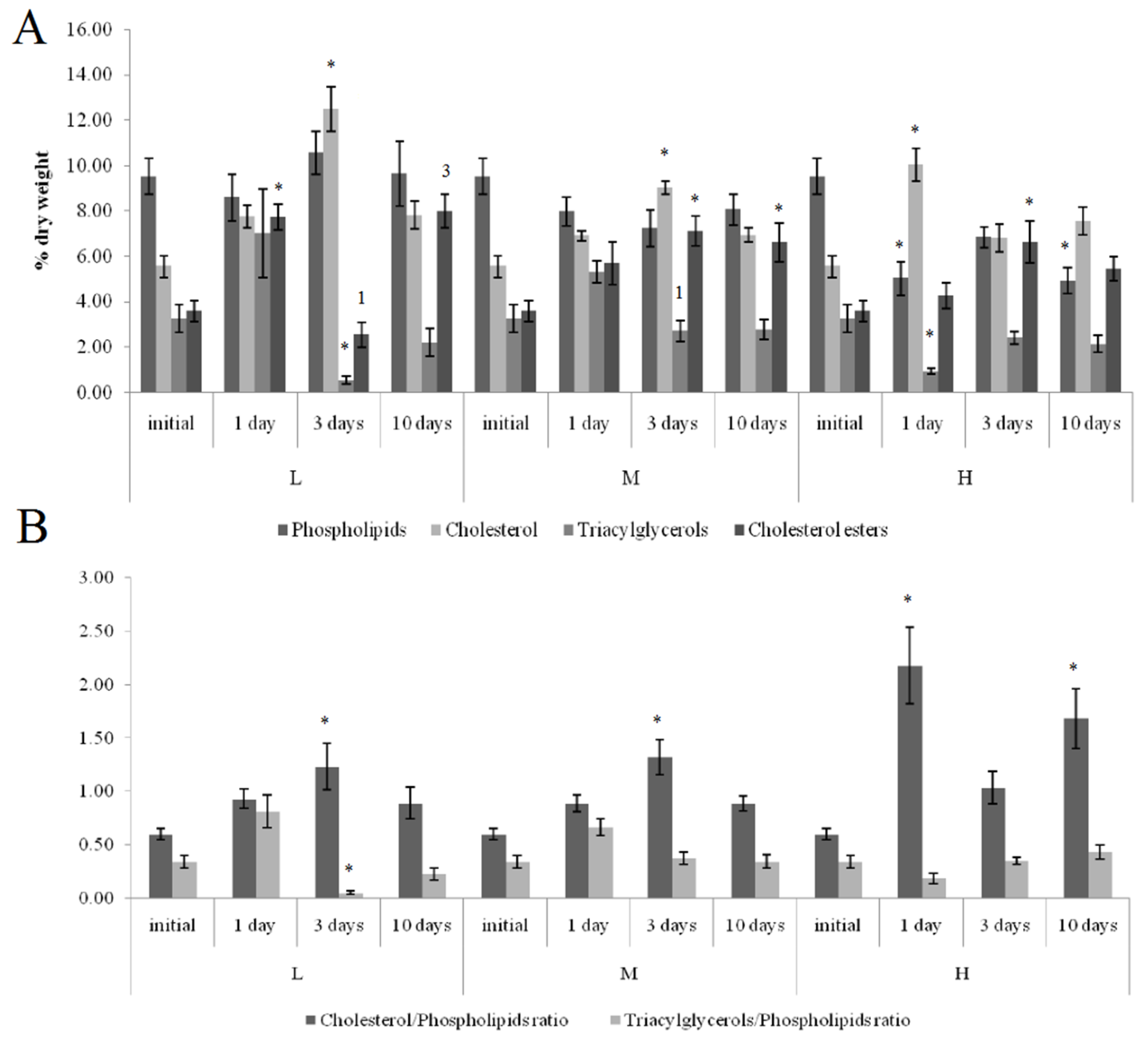
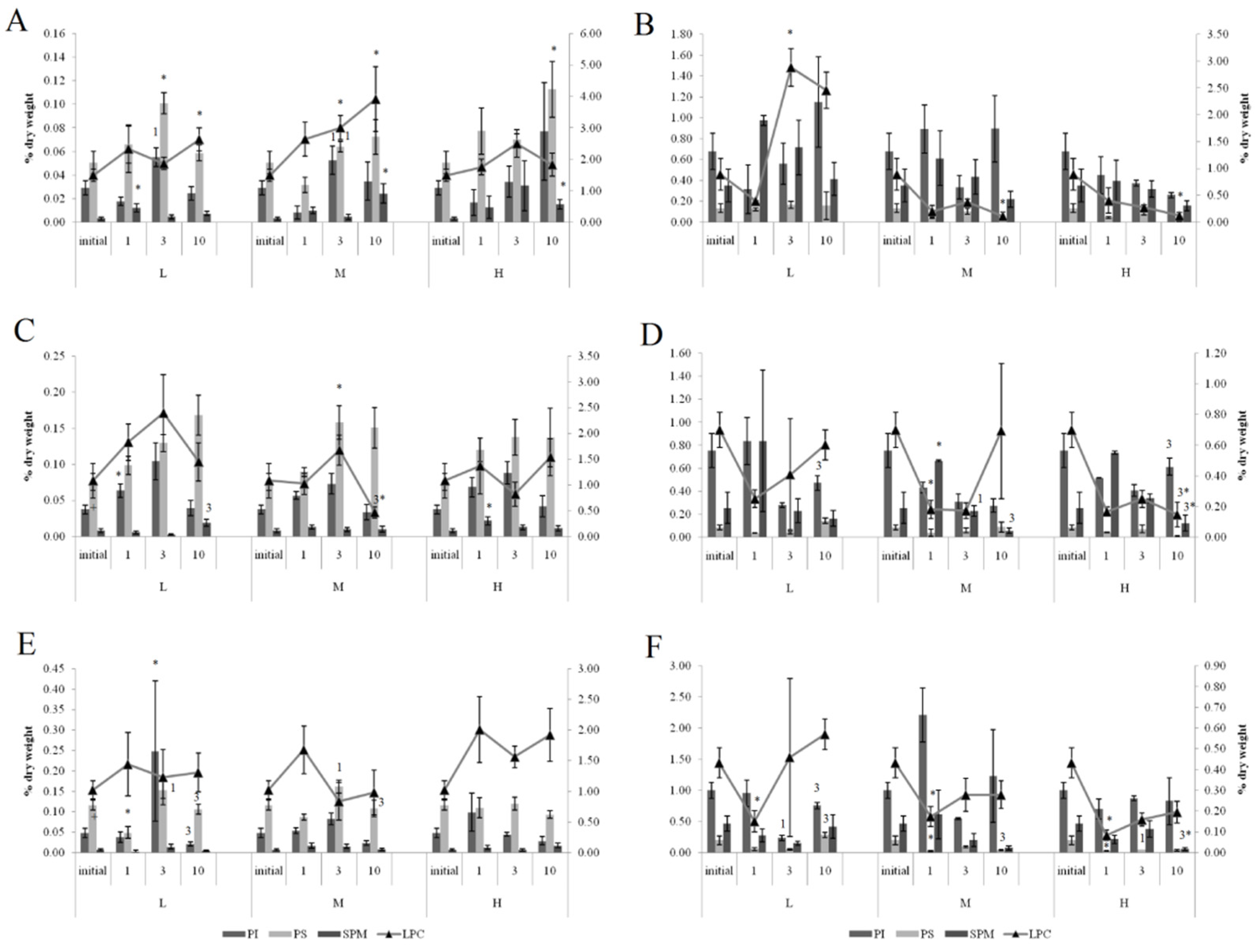
3.3. Lipid Composition
3.3.1. Effects of Crude Oil Pollution on Horse Mussels
3.3.2. Effects of Oil Pollution on Blue Mussels (Normal Salinity)
3.3.3. Effects of Low Salinity and Oil Pollution on Blue Mussels
4. Discussion
4.1. Heart Rates
4.2. Lipid Composition
4.3. Effect of Low Salinity and Oil Pollution on Lipid Composition in Blue Mussels
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Noyes, P.D.; McElwee, M.K.; Miller, H.D.; Clark, B.W.; Van Tiem, L.A.; Walcott, K.C.; Erwin, K.N.; Levin, E.D. The toxicology of climate change: Environmental contaminants in a warming world. Environ. Int. 2009, 35, 971–986. [Google Scholar] [CrossRef]
- Arctic Monitoring and Assessment Programme (AMAP). AMAP Assessment 2009: Persistent Organic Pollutants in the Arctic; Arctic Monitoring and Assessment Programme (AMAP): Oslo, Norway, 2010. [Google Scholar]
- Anwar, N.; Richardson, C.; Seed, R. Age determination, growth rate and population structure of the horse mussel Modiolus modiolus. J. Mar. Biol. Assoc. U. K. 1990, 70, 441–457. [Google Scholar] [CrossRef]
- Brooks, S.J.; Farmen, E. The Distribution of the Mussel Mytilus Species along the Norwegian Coast. J. Shellfish Res. 2013, 32, 265–270. [Google Scholar] [CrossRef] [Green Version]
- Menge, A.B. Top-down and bottom-up community regulation in marine rocky intertidal habitats. J. Exp. Mar. Biol. Ecol. 2000, 250, 257–289. [Google Scholar] [CrossRef]
- Marshall, D.J.; McQuaid, C.D. Seasonal and diel variations of in situ heart rate of the intertidal limpet Siphonaria oculus Kr. (Pulmonata). J. Exp. Mar. Biol. Ecol. 1994, 179, 1–9. [Google Scholar] [CrossRef]
- Santini, G.; Williams, G.A.; Chelazzi, G. Assessment of factors affecting heart rate of the limpet Patella vulgata on the natural shore. Mar. Biol. 2000, 137, 291–296. [Google Scholar] [CrossRef]
- Marchan, S.; Davies, M.S.; Fleming, S.; Jones, H.D. Effect of copper and zinc on the heart rate of the limpet Patella vulgata L. Comp. Biochem. Physiol. B 1999, 123, 89–93. [Google Scholar] [CrossRef]
- Bloxham, M.J.; Worsfold, P.J.; Depledge, M.H. Integrated biological and chemical monitoring: In Situ physiological responses of freshwater crayfish to fluctuations in environmental ammonia concentrations. Ecotoxicology 1999, 8, 225–237. [Google Scholar] [CrossRef]
- Endo, S.; Escher, B.I.; Goss, K.-U. Capacities of Membrane Lipids to Accumulate Neutral Organic Chemicals. Environ. Sci. Technol. 2011, 45, 5912–5921. [Google Scholar] [CrossRef]
- Camus, L.; Birkely, S.; Jones, M.; Børseth, J.; Grøsvik, B.; Gulliksen, B.; Lønne, O.; Regoli, F.; Depledge, M. Biomarker responses and PAH uptake in Mya truncata following exposure to oil-contaminated sediment in an Arctic fjord (Svalbard). Sci. Total Environ. 2003, 308, 221–234. [Google Scholar] [CrossRef]
- Reich, T.; DePew, M.; Narks, G.; Singer, M.; Wan, J. Effect of polychlorinated biphenyls (PCBs) on phospholipid membrane fluidity. J. Environ. Sci. Health Environ. Sci. Eng. 1981, 16, 65–71. [Google Scholar] [CrossRef]
- Dzogbefia, V.P.; Kling, D.; Gamble, W. Polychlorinated biphenyls: In Vivo and in vitro modifications of phospholipid and glyceride biosynthesis. J. Environ. Pathol. Toxicol. 1978, 1, 841–856. [Google Scholar] [PubMed]
- Capuzzo, J.M.; Leavitt, D.F. Lipid composition of the digestive glands of Mytlus edulis and Carcinus maenas in response to pollutant gradients. Mar. Ecol. Prog. Ser. 1988, 46, 139–145. [Google Scholar] [CrossRef]
- Tedengren, M.; Kautsky, N. Comparative study of the physiology and its probable effect on size in Blue Mussels (Mytilus edulis L.) from the North Sea and the Northern Baltic Proper. Ophelia 1986, 25, 147–155. [Google Scholar] [CrossRef]
- Lehtonen, K.; Leiniö, S.; Schneider, R.; Leivuori, M. Biomarkers of pollution effects in the bivalves Mytilus edulis and Macoma balthica collected from the southern coast of Finland (Baltic Sea). Mar. Ecol. Prog. Ser. 2006, 322, 155–168. [Google Scholar] [CrossRef] [Green Version]
- Naumov, A.D. Bivalve mulluscs of the White Sea. The Experience of Ecology-Phaunisticalanalyze; ZIN: St. Peterburg, Russia, 2006. (In Russian) [Google Scholar]
- Depledge, M.; Andersen, B. A computer-aided physiological monitoring system for continuous, long-term recording of cardiac activity in selected invertebrates. Comp. Biochem. Physiol. A 1990, 96, 473–477. [Google Scholar] [CrossRef]
- De Pirro, M.; Santini, G.; Chelazzi, G. Cardiac responses to salinity variations in two differently zoned Mediterranean limpets. J. Comp. Physiol. B 1999, 169, 501–506. [Google Scholar] [CrossRef]
- Folch, J.; Lees, M.; Sloan-Stanley, G.H. A simple method for the isolation and purification of total lipids animal tissue (for brain, liver and muscle). J. Biol. Chem. 1957, 226, 497–509. [Google Scholar] [CrossRef]
- Sidorov, V.S.; Lizenko, E.I.; Bolgova, O.M.; Nefedova, Z.A. Fish Lipids. 1. Analysis Technique; Karelian Branch of the USSR Academy of Science: Petrozavodsk, Russia, 1972; pp. 150–162. (In Russian) [Google Scholar]
- Engelbrecht, F.M.; Mari, F.; Anderson, J.T. Cholesterol Determination in serum. A rapid direction method. S. Afr. Med. J. 1974, 48, 250–256. [Google Scholar]
- Arduini, A.; Peschechera, A.; Dottori, S.; Sciarroni, A.F.; Serafini, F.; Calvani, M. High performance liquid chromatography of long-chain acylcarnitine and phospholipids in fatty acid turnover studies. J. Lip. Res. 1996, 37, 684–689. [Google Scholar] [CrossRef]
- Fokina, N.; Bakhmet, I.; Shklyarevich, G.; Nemova, N. Effect of seawater desalination and oil pollution on the lipid composition of blue mussels Mytilus edulis L. from the White Sea. Ecotoxicol. Environ. Saf. 2014, 110, 103–109. [Google Scholar] [CrossRef]
- Fokina, N.; Bakhmet, I.; Nemova, N. Cooperative effect of crude oil and low salinity on the digestive glands lipid composition of the White Sea blue mussels Mytilus edulis. Proc. Zool. Inst. RAS 2016, 320, 357–366. [Google Scholar] [CrossRef] [Green Version]
- Navarro, J.M.; Thompson, R.J. Physiological energetics of the horse mussel Modiolus modiolus in a cold ocean environment. Mar. Ecol. Prog. Ser. 1996, 138, 135–148. [Google Scholar] [CrossRef] [Green Version]
- Bamber, S.D.; Depledge, M.H. Evaluation of changes in the adaptive physiology of shore crabs (Carcinus maenas) as an indicator of pollution in estuarine environments. Mar. Biol. 1997, 129, 667–672. [Google Scholar] [CrossRef]
- Kuwasawa, K.; Hill, R. Evidence for cholinergic inhibitory and serotonergic excitatory neuromuscular transmission in the heart of the bivalve Mercenaria mercenaria. J. Exp. Biol. 1997, 200, 2123–2135. [Google Scholar] [CrossRef] [PubMed]
- Bakhmet, I.N.; Fokina, N.N.; Nefedova, Z.A.; Nemova, N.N. Physiological-biochemical properties of blue mussel Mytilus edulis adaptation to oil contamination. Environ. Monit. Assess. 2009, 155, 581–591. [Google Scholar] [CrossRef] [PubMed]
- Berger, V.J.; Kharazova, A.D. Mechanisms of salinity adaptations in marine mollusks. Hydrobiology 1997, 355, 115–126. [Google Scholar] [CrossRef]
- McDowell, J.E.; Lancaster, B.A.; Leavitt, D.F.; Rantamaki, P.; Ripley, B. The effects of lipophilic organic contaminants on reproductive physiology and disease processes in marine bivalve molluscs. Limnol. Oceanogr. 1999, 44, 903–909. [Google Scholar] [CrossRef]
- Nechev, J.; Stefanov, K.; Popov, S. Effect of cobalt ions on lipid and sterol metabolism in the marine invertebrates Mytilus galloprovincialis and Actinia equine. Comp. Biochem. Physiol. A 2006, 144, 112–118. [Google Scholar] [CrossRef]
- Hochachka, P.M.; Somero, G.N. Biochemical Adaptation; Princeton University Press: Oxford, UK, 2002. [Google Scholar]
- Cappello, T.; Pereira, P.; Maisano, M.; Mauceri, A.; Pacheco, M.; Fasulo, S. Advances in understanding the mechanisms of mercury toxicity in wild golden grey mullet (Liza aurata) by 1H NMR-based metabolomics. Environ. Pollut. 2016, 219, 139–148. [Google Scholar] [CrossRef]
- Moore, M.N.; Viarengo, A.; Donkin, P.; Hawkins, A.J. Autophagic and lysosomal reactions to stress in the hepatopancreas of blue mussels. Aquat. Toxicol. 2007, 84, 80–91. [Google Scholar] [CrossRef] [PubMed]
- Vance, D.E.; Vance, J.E. (Eds.) Biochemistry of Lipids, Lipoproteins and Membranes, 4th ed.; Elsevier: Amsterdam, The Netherlands, 2002. [Google Scholar]
- Scheek, S.; Brown, M.S.; Goldstein, J.L. Sphingomyelin depletion in cultured cells blocks proteolysis of sterol regulatory element binding proteins at site 1. Proc. Natl. Acad. Sci. USA 1997, 94, 11179–11183. [Google Scholar] [CrossRef] [PubMed] [Green Version]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Days | Low | Medium | High |
|---|---|---|---|
| 0–Initial (nominal) | 9 | 45 | 450 |
| 1 day (measured) | 2.20 ± 0.64 | 3.50 ± 0.17 | 45.00 ± 3.80 |
| 2 days (measured) | 0.80 ± 0.10 | 0.97 ± 0.04 | 18.60 ± 1.31 |
| 3 days (measured) | 0.17 ± 0.04 | 0.29 ± 0.05 | 12.90 ± 2.03 |
| 10 days (measured) | 0.05 ± 0.01 | 0.05 ± 0.01 | 4.80 ± 0.93 |
| Effects | 1 Day | 2 Days | 3 Days | 4 Days | 5 Days | 6 Days |
|---|---|---|---|---|---|---|
| Oil concentration | 0.001 | 0.001 | 0.001 | 0.059 | 0.014 | 0.070 |
| Species | 0.001 | 0.060 | 0.001 | 0.001 | 0.001 | 0.001 |
| Interaction | 0.040 | 0.036 | 0.004 | 0.382 | 0.058 | 0.094 |
| Effects | 1 Day | 2 Days | 3 Days | 4 Days |
|---|---|---|---|---|
| Oil concentration | 0.006 | 0.001 | 0.001 | 0.527 |
| Salinity | 0.922 | 0.351 | 0.001 | 0.647 |
| Interaction | 0.768 | 0.006 | 0.001 | 0.224 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bakhmet, I.; Fokina, N.; Ruokolainen, T. Changes of Heart Rate and Lipid Composition in Mytilus Edulis and Modiolus Modiolus Caused by Crude Oil Pollution and Low Salinity Effects. J. Xenobiot. 2021, 11, 46-60. https://0-doi-org.brum.beds.ac.uk/10.3390/jox11020004
Bakhmet I, Fokina N, Ruokolainen T. Changes of Heart Rate and Lipid Composition in Mytilus Edulis and Modiolus Modiolus Caused by Crude Oil Pollution and Low Salinity Effects. Journal of Xenobiotics. 2021; 11(2):46-60. https://0-doi-org.brum.beds.ac.uk/10.3390/jox11020004
Chicago/Turabian StyleBakhmet, Igor, Natalia Fokina, and Tatiana Ruokolainen. 2021. "Changes of Heart Rate and Lipid Composition in Mytilus Edulis and Modiolus Modiolus Caused by Crude Oil Pollution and Low Salinity Effects" Journal of Xenobiotics 11, no. 2: 46-60. https://0-doi-org.brum.beds.ac.uk/10.3390/jox11020004