1. Introduction
Shortage of fresh water along with the changes in global environmental conditions have created a problem for the agricultural researchers to fulfill the food demand for the increasing world population with a target to fulfill this demand till 2050 [
1,
2]. Global environmental changes have altered the ongoing rainfall patterns all over the world, resulting in drought stress in some areas and flooding in others [
3]. Under such conditions, the demand for food is increasing due to the deleterious effects of drought on crop production [
4,
5]. Different ways or techniques are being employed to handle this problem and improve and conserve plants. These include improved agronomic practices with water conservation, genetic characterization, developing and selection of drought tolerant genotypes with better production as well as the exogenous use of organic and inorganic chemicals through different modes [
6,
7,
8,
9,
10]. Under soil water deficit conditions, the perturbations in plant water relations is the foremost effect along with reduced uptake of nutrients and disturbed cellular ion homeostasis, and this situation becomes more severe when the soil is already deficit to specific micro or macro nutrients [
11]. Among different essential micro-nutrients, iron (Fe) is an important one due to its multiple roles in plant cellular metabolic activities linked to enhanced plant growth, metabolism, and morphology [
12]. It is important for maintaining the function and structure of chloroplast involved in chlorophyll biosynthesis [
13]. It also plays a crucial role as a prosthetic group of many enzymes, including peroxidases, cytochromes and catalases. Young and Ajami [
14] reported its active role in synthesis of cellular protein. As an important component of enzymes and protein, it also plays an active role in many biological processes, including photosynthesis and nitrogen fixation, as well as synthesis of metallo-protein that need Fe to perform their proper functioning [
15]. As a part of Fe-S centers, it also plays an important role in the electron transport chain [
16].
The presence of inadequate concentration of Fe in plants results in reduced growth and yellowing of leaves [
13]. Studies reveal that Fe deficiency in rooting medium results in reduced nutritional quality and seed yield [
17]. Iron is required in an optimal concentration in plants for better growth because it helps plants in performing their biochemical and physiological functions in a proper way. The excessive concentration of Fe plants in ionic form is potentially toxic and results in excessive formation of reactive oxygen species, resulting in lipid peroxidation by damaging the cellular membranes [
13,
18]. Iron also has a very low solubility and bioavailability under low soil osmotic potential [
19,
20], resulting in restricted plant productivity and low quality. To fulfill the Fe deficiency, different inorganic fertilizers are being used in soil or foliar spray. These include mainly the use of FeSO
4, FeCl as foliar spray or soil application, and the use of FeSO
4 is most common, but FeSO
4 use has some drawbacks in crop plants [
21]. As a chemical in nature, its use is not eco-friendly [
22]. It is well reported that chemical fertilizers such as FeSO
4 have raised issues including the significant risks that may happen for animal, humans and plants, as well as for the ecosystem’s health [
23,
24]. Generally, used fertilizers are simple chemicals and readily available in ionic form that undergo various harmful reactions in soil solution as well as in cells after absorbing, resulting in low uptake and disturbed metabolism [
24,
25,
26]. These include the complex formation, volatilization, precipitation, fixation and leaching that reduce the efficiency of chemical fertilizers [
24,
27]. It has been found that Fe availability in plants is also reduced due to the different cellular oxidation reactions [
21].
To minimize the adversities of such effects, the interest in the use of eco-friendly biofertilizers is increasing, including the micro-nutrient chelated amino acids [
21]. In chelation, the uptake and availability of the micro-nutrients increase with no or less chances of oxidation of the nutrients [
24]. The key feature that makes the use of aminochelate fertilizers as eco-friendly ones with higher efficiency is the stability of the chelated bond between micronutrient cations and amino acids in nutrient solution as compared with its use as a chemical fertilizer [
21,
24,
28].
Additionally, in the case of chelation with an organic molecule, both parts of the chelation play a significant role in different cellular metabolic activities [
29]. For example, along with the beneficial effects of micro-nutrients, the chelated amino acid metabolism also plays a key role in improving stress tolerance of plants [
7,
29]. Among different amino acids, glutamate plays multiple roles in growth improvement and other metabolic activities both under normal and stressful conditions. Studies revealed its role in the synthesis of lysine, arginine and orthenine as a precursor as well as its indirect role in various other metabolic activities [
30,
31]. Dinu et al. [
32] reported that glutamate has an important role in osmotic adjustment of the stomatal guard cells as it controls the opening and closing of the stomata. The endogenous excessive accumulation of the glutamate under normal or stressed conditions or through its exogenous application, confirms its role as a signaling molecule in ameliorating the deleterious effects of multiple abiotic stresses [
31]. For example, the exogenous application of glutamate on
Brassica compestris reduced the negative impacts of salt and osmotic stress [
33]. Ramos et al. [
34] reported an endogenous increase in glutamate of
Glycine max L. grown under water deficiency, resulting in the increased accumulation of many other amino acids along with its own accumulation. In another study conducted by Sadak et al. [
35], it was found that exogenous application of glutamate as a foliar spray on faba bean reduced the deleterious impacts of salinity.
Ajwain is an aromatic herb belonging to the family umbelliferae (apiaceae). Its seed has a high amount of essential minerals, flavone (7.1%), protein (15.4%), carbohydrates (38.9%), moisture (8.9%) and fiber (11.9%) [
36]. The essential oil extracted from seeds of ajwain is being used in minor quantities in perfumery, food flavoring as preservatives and, most extensively, in folk medicines [
36], especially for remedies of stomach disorders. Dry and hot fruit fomentation is externally applied on the chest for curing asthma, and a crushed fruit paste is applied for colic pains [
37]. In many developing countries, this crop is ignored and mainly grown in dry and rainfed areas with no availability of water for irrigation, where it is facing the problem of yield losses. Moreover, in Pakistan, the ajwain crop faces the problem of a hot and dry environment during the onset of reproduction and harvesting growth stages.
Till now, no reports are available about the use of Fe-Glu for improving the plant performance and plant sensitivity to limited water supply as well as to overcome the plant Fe deficiency under water stressed conditions. So, in view of the information available, it was hypothesized that Fe-chelated glutamate might improve the plant performance and water stress tolerance, and will be helpful to overcome the plant Fe deficiency in ajwain plants grown under water stress and non-stressed conditions in comparison with FeSO4 due to its foliar application at vegetative stage. The aim of the study was to explore the comparative roles of different levels of Fe-Glu and FeSO4 to overcome the plant iron deficiency along with the induction of plant performance in view of different morphological attributes, biomass production, physio-biochemical parameters, oxidative defense mechanism, nutrient acquisition and seed yield. Furthermore, the present study aimed to find out the most effective level of Fe required for better seed yield.
2. Materials and Methods
Experimental site was a research area located in the Botanical Garden at Government College University Faisalabad (latitude 30°30′ N, longitude 73°10′ E and altitude 213 m). Water stress was maintained as the number of irrigations. Experiment was conducted in a factorial arrangement. Experimental area was divided into two main plots; each is specific to water stress level (normal irrigation and water stress with irrigation numbers). Each main plot was then divided into eight subplots corresponding to specific treatments. The different treatments used were No Spray (NS), Water Spray (WS), FeSO
4 (0.25%), FeSO
4 (0.5%), Fe-Glu (0.25%), and Fe-Glu (0.5%). There were four rows of equal size in each subplot corresponding to the specific treatment. In each plot, one row was specified for NS, and the other one for WS, while the remaining two lines were specified to 0.25% level of FeSO
4 or Fe-Glu. The same arrangement was followed for 0.5% level of FeSO
4 or Fe-Glu. In each main plot out of eight subplots, four were allocated to 0.25% level and the other four for 0.5% level. The subplots in each main plot were distributed randomly to minimize the error due to edaphic factors. The same arrangement was followed for the other main plot. The complete layout plan of the experiment is given in
Table S1 as a
Supplementary Material.
2.1. Soil Analysis
The soil of the experimental site was sandy loam having organic matter (1.15%) and available total N (0.73%) and P (8.6 ppm), with a saturation percentage of 34%. The average pH and EC of the soil was 7.8 and 2.53 ds.m
−1, respectively. The soil solution has the HCO
3− (4.93 meqL
−1), soluble CO
32− (traces), SO
4−2 (1.98 meq L
−1), Cl
− (8.52 meq L
−1), Ca
2++Mg
2+ (14.3 meq L
−1), Fe (0.041 meq L
−1), Na (2.98 meq L
−1) and SAR (0.086 meq L
−1). The soil’s physical and chemical properties were assayed following Davis and Freitas [
38].
2.2. Application of Water Stress and Foliar Spray of Different Treatments
During growing season (October/November 2018), the soil was irrigated well with canal water before seed sowing for field preparation. Healthy seeds of the ajwain (
Trachyspermum ammi L.) locally known as Desi Ajwain were collected from the market and checked for germination before sowing. Then, the soil was ploughed well for seed sowing after 15 days of irrigation. Sowing of seeds was done at ½ inch depth on 1st November 2018. The first irrigation was applied after 10 days of sowing. This irrigation was not included in water stress schedule. Second irrigation was applied after 15 days of first irrigation, to all the plots including the selected ones for water stress treatment. Later, the non-stressed plots were irrigated as per water requirement till maturity, but the plants selected for water stress treatment were watered twice, i.e., at fully vegetative and flowering stages only. The irrigated water was tested in terms of quality, having BOD (1.50 L
−1), SAR (0.85), pH (7.6), EC (0.37 dS m
−1), acidity (86 mg L
−1), alkalinity (18 mg L
−1), HCO
3 + CO
3 (1.7 mg L
−1), SO
4 (0.9 meq L
−1), PO
4 (0.114 mg L
−1), and NO
3 (0.95 mg L
−1). Different minerals in irrigation water were K (4.29 mg L
−1), P (3.11 mg L
−1), N (12 mg L
−1), As (1 ppb), Zn (0.04 mg L
−1), Cu (0.15 mg L
−1), Na (0.34 mg L
−1), Ca (1.62 meq L
−1), Mg (1.18 meq L
−1), Cl (53.9 meq L
−1) and Fe (0.11 mg L
−1). The plants were thinned in a row to maintain the distance of 9 inches among plants, after 10 days of 1st irrigation. However, row to row distance was kept 18 inches at the time of seed sowing. After three weeks of 2nd irrigation, different treatments were applied as foliar spray. Different concentrations of Fe-Glu and FeSO
4 were prepared in water and before foliar spray, 0.1% twin-20 was mixed in solution as surfactant. The FeSO
4 and glutamate were of analytical grade purchased from Merck, Germany. The plants were sprayed with Fe-Glu and FeSO
4 by using the concentrations of 0.25% and 0.5% of each. The method given by Leu [
39] was followed for preparation of Fe-Glu. Briefly, 260 g of FeSO
4.7H
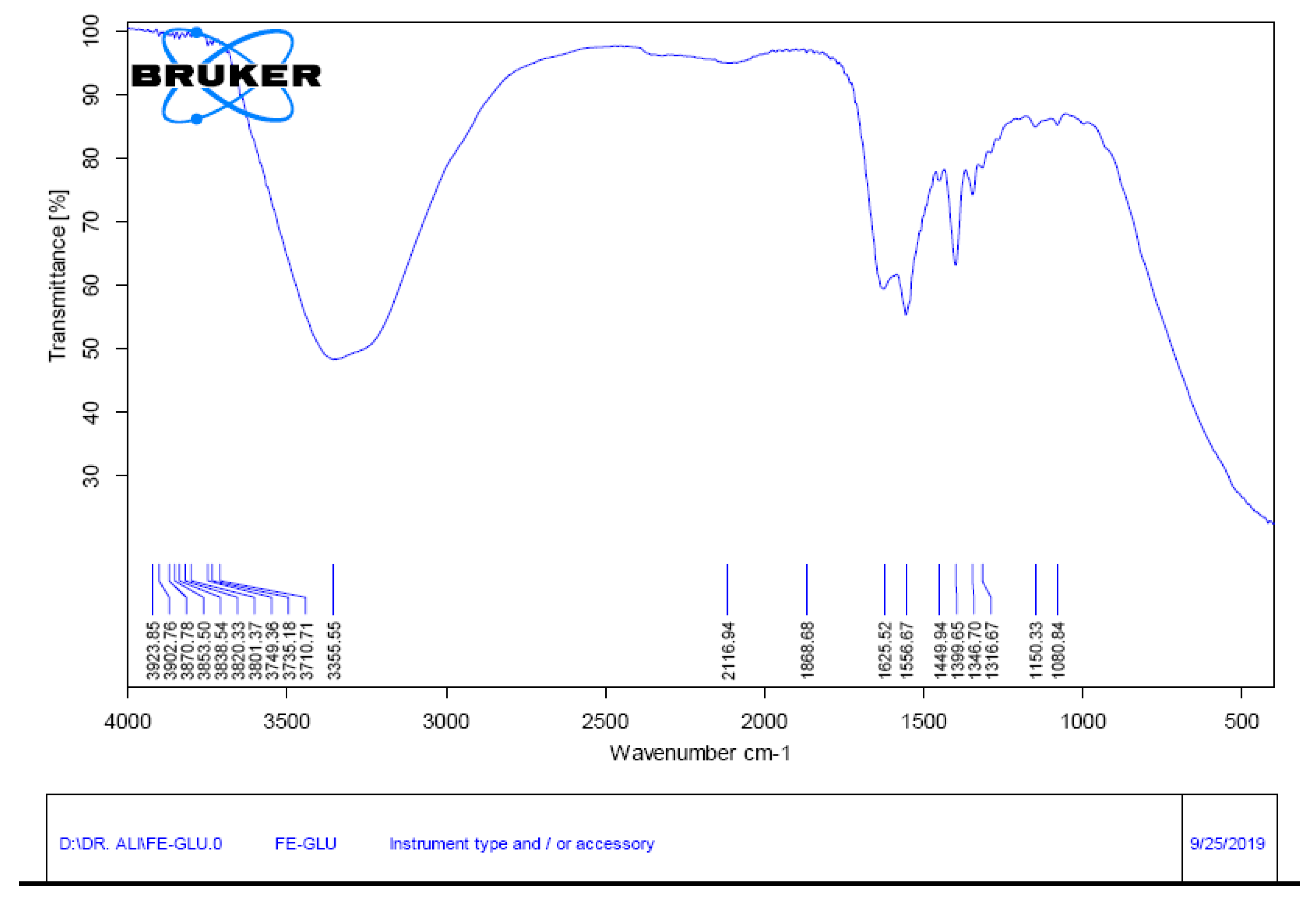
2O were dissolved in water (150 mL) and 146.12 g (equimolar quantity) of L-glutamate were added. The chelation was confirmed by fourier transform infrared spectroscopy (FTIR) analysis for Fe-Glu (
Figure 1) and FeSO
4.7H
2O [
40]. Foliar application of solutions was done in the evening for the maximum absorption by leaves. To avoid the rain in applied water stress, the whole experimental area was covered with a polythene sheet that was used only when there were the chances of rain. Data were collected for different attributes after three weeks of foliar application of different treatments. Two plants were selected from each row (treatment) in each plot (a total number of eight plants against each treatment) for measuring the shoot and root fresh masses and lengths of shoots and roots. These plant samples were then kept in an electric oven for 48 h at 65 °C to obtain their dry masses. The oven-dried root and shoot samples were then used to measure the mineral nutrients.
2.3. Estimation of Leaf Relative Water Content (LRWC)
Fully developed leaf (3rd from top) from each plant was used for measuring the LRWC. The leaves were excised using scissors and their fresh weights were measured. Leaves were then dipped in distilled water after tagging with permanent marker for 6 h. The leaves were then taken out from water, adsorbed the excess water on their surface and their turgid weights were measured. Then, dry weights of the leaves were measured after oven-drying at 70 °C for 48 h. Then, the following equation was used for measuring LRWC:
2.4. Estimation of Leaf Relative Membrane Permeability (LRMP)
The method described by Yang et al. [
41] was followed to find out the LRMP. The known amount (0.5 g) of excised leaf was cut into small pieces (approximately 1 cm) and put in test tubes having 20 mL of de-ionized dH
2O. After vortexing well for 5 s, the EC of the assayed material was measured and termed as EC0. The test tubes containing leaf material were then kept at 4 °C for 24 h and the EC1 was measured. These test tubes containing leaf material were then autoclaved for 30 min at 120 °C and assayed the EC2. The LRMP was measured using the equation as follows:
2.5. Estimation of Leaf Chlorophyll (Chl.) and Carotenoid (Car.) Contents
For the estimation of leaf Chl.
a,
b, total Chl. (T. Chl.) and Chl.
a/b, the method described by Arnon [
42] was followed. The content of Car. was estimated following Kirk and Allen [
43]. The extraction of the pigments was done using 80% acetone. Briefly, fresh leaf material (0.1 g) was chopped and put in 10 mL 80% acetone for overnight at 4 °C and the absorbance of the extract was read at 663, 645 and 480 nm using spectrophotometer (Hitachi U-2001, Tokyo, Japan). The quantities were computed using the specific formulas:
ΔA = absorbance at respective wavelength
v = volume of the extract (mL)
w = weight of the fresh leaf tissue (g).
2.6. Estimation of Leaf Total Phenolic Content (TPC)
The method ascribed by Julkenen-Titto [
44] was followed for the estimation of TPC in fresh leaves. After homogenization of fresh leaf material (0.05 g) in 5 mL acetone (80%), the homogenate was centrifuged at 10,000×
g for 10 min. Then, 100 µL of the obtained supernatant was reacted with Folin–Ciocalteau’s phenol reagent (1 mL). Two mL of dH
2O were added to the triturate along with 20% Na
2CO
3 (5.0 mL) solution. The final volume of the resultant mixture was raised to 10 mL with dH
2O. After mixing vigorously, the OD of the triturate was read at 750 nm using a UV-Visible spectrophotometer (IRMECO U2020) (GmbH, Geesthacht, Germany).
2.7. Estimation of Leaf Total Anthocyanin (T. antho) Content
For the estimation of leaf anthocyanin content, fresh leaf material (50 mg) was homogenized thoroughly in 1000 μL of acidic MeOH (1% HCl,
w/
v) in an ice bath. The homogenate was centrifuged at (14,000 rpm) for 5 min at room temperature. The OD of the supernatant was read at 657 and 530 nm. The equation given below was used for the quantification of T. antho content:
where A657 and A530 is the absorption at specific wavelengths and M is the fresh leaf mass used for the extraction (g).
2.8. Determination of Leaf Ascorbic Acid (AsA) Content
The method ascribed by Mukherjee and Choudhuri [
45] was followed for the estimation of AsA in fresh leaf. Fresh leaf (0.25 g) was ground well in 10 mL trichloroacetic acid (TCA) solution (6%). After centrifugation at 10,000×
g for 10 min, the supernatant was reacted with 2% acidic dinitrophenyl hydrazine (2 mL) solution. Then, 10% thiourea (one drop) was added to the triturate. The resulted mixture was boiled for 20 min in a water bath. The mixture was then cooled at 0 °C and 80% H
2SO
4 (5 mL) was added in the mixture. The OD of the finally prepared colored material was read at 530 nm. The standard curve was prepared from a range of standard solutions (50–300 ppm) by using pure AsA to estimate the concentration in the leaf samples.
2.9. Estimation of Leaf Antioxidant Enzyme Activities and Total Soluble Protein (TSP)
2.9.1. Extraction of Antioxidant Enzymes and Total Soluble Proteins (TSP)
For the extraction of antioxidant enzymes and TSP, fresh leaf material (0.5 g) was ground in 10 mL chilled (50 mM phosphate buffer (pH 7.8). The mixture was then centrifuged at 10,000× g for 20 min at 4 °C. The supernatant so obtained was then used for the estimation of total soluble proteins (TSP) and antioxidative enzymes activities.
2.9.2. Estimation of TSP
TSP in the buffer extracts was estimated following the method of Bradford [
46]. The absorbance of the triturate was measured at 595 nm and the quantities of the TSP in samples were computed using a series of protein standards (200–1400 mg/kg) prepared from analytical grade bovine serum albumin (BSA).
2.9.3. Estimation of Superoxide Dismutase (SOD) Activity
The method ascribed by Giannopolitis and Ries [
47] was employed for the estimation of SOD activity that was measured based on the photochemical reduction inhibition principle of nitroblue tetrazolium (NBT) in light. The reduction inhibition of NBT was measured at 560 nm. Briefly, the prepared mixture for the study was consisted of 1.3 μM riboflavin, 13 mM methionine, 50 μM NBT, 75 nM EDTA, 50 μL enzyme extract and 50 mM phosphate buffer (pH 7.8). The mixture was placed in a box, coated internally with aluminum foil under a fluorescent light (20 W) for 15 min. Then, the absorbance of the reaction mixture was read using a UV-visible spectrophotometer at 560 nm. Along with the reaction mixture prepared using the sample, a reaction mixture without sample was also used as a blank. The SOD activity in samples was estimated as unit/mg protein using the content of TSP.
2.9.4. Determination of Peroxidase (POD) Activity
The method ascribed by Chance and Maehly [
48] was followed to assay the POD activity in the leaf buffer extract. The basic mechanism behind the estimation of POD activity was based on the oxidation of guaiacol used in reaction mixture. The reaction mixture was comprised 40 mM H
2O
2, 20 mM guaiacol, 50 mM phosphate buffer and 100 μL leaf phosphate buffer extract. The guaiacol was added last in the reaction solution to start the reaction. The absorbance of the reaction mixture was read with the interval of 20 s, at 470 nm by setting spectrophotometer in a time scan manner. The change of 0.01 absorbance unit min
−1 mg
−1 of protein was taken as unit POD activity. The final activity was measured as units/mg protein.
2.9.5. Determination of Catalase (CAT) and Ascorbate Peroxidase (APX) Activity
The method ascribed by Chance and Maehly [
48] was employed for the estimation of CAT activity in leaf buffer extract. The decrease in the absorbance of the mixture was taken as disappearance of H
2O
2 as a basic phenomenon behind the estimation of CAT activity. The reaction mixture comprised 1.9 mL H
2O
2, 1 mL dH
2O and 100 µL of extract. The APX activity was estimated using the method given by Asada and Takahashi [
49]. The activities of these enzymes were estimated by the decrease in absorbance at 240 and 290 nm, respectively, and expressed as units mg
−1 protein.
2.10. Estimation of Total Free Amino Acid (FAA) Content
The method ascribed by Hamilton and Van-slyke [
50] was followed for the estimation of FAA. The same buffer extract as used for the estimation of TSP and enzyme activities was used for the estimation of FAA. The reaction mixture was comprised 1 mL of 2% ninhydrin, 1 mL of 1% pyridine and 1 mL of the extract (buffer extract). The mixture was then heated in a water bath at 95 °C for 30 min. After cooling, the final volume was made 50 mL and the absorbance was read at 570 nm.
2.11. Determination of Leaf Reducing Sugars (RS)
Leaf RS were estimated following the method ascribed by Wood and Bhat [
51]. Fresh leaf sample (0.5 g) was extracted using 80% methanol (5 mL) and centrifuged at 10,000×
g. In a test tube, 4 mL of the 3, 5-dinitrosalicylic acid reagent (DNS) (prepared by dissolving 1 g DNS in 20 mL of 2M NaOH and mixed with 30 g sodium potassium tartrate and diluted to 100 mL using dH
2O) were added to 1 mL of the sugar solution, and the mixture was then kept in a water bath at 95 °C for 5 min. Then, the samples were kept in chilled water to rapidly cool the solution and to incubate the resulting material at 25 °C, and the absorbance was then read at 540 nm.
2.12. Estimation of Total Soluble Sugars (TSS)
The same leaf extract as used for the estimation of reducing sugars was also used for the determination of TSS. The reaction mixture was comprised 3 mL of antheron reagent and 0.1 mL of the extract. The mixture was then put in water bath for 10 min at 95 °C. The mixture was then cooled in chilled water. The mixture was then incubated for 30 min at room temperature and the absorbance was read at 625 nm. The final concentration of TSS in samples was estimated using the curve prepared from pure standards.
2.13. Estimation of Leaf Malondialdehyde (MDA) Content
Content of MDA was measured in terms of lipid peroxidation using the method given by Cakmak and Horst [
52]. Trichloroacetic acid (TCA) method was used for the estimation of MDA content. One g leaf material freshly taken was ground in 10 mL of TCA (10% solution prepared in dH
2O). The supernatant (0.5 mL) as obtained from the homogenized material was mixed with 2 mL of 0.5% thiobarbituric acid (TBA), prepared in 20% TCA. Test tubes having the triturate were kept at 95 °C for 50 min, and then cooled immediately in chilled water. After centrifugation (10,000×
g) of mixture for 10 min, the absorbance of colored part was read at 600 and 532 nm. The content of MDA was calculated using the formula:
Absorption coefficient for the calculation of MDA is 156 mmol−1 cm−1.
2.14. Estimation of Leaf H2O2 Content
Leaf H
2O
2 content was determined following the method of Velikova et al. [
53]. The fresh leaf sample (0.5 g) was ground well in 10 mL TCA (6%). To 0.1 mL of centrifuged TCA extract, 1 mL of l M KI solution was added. The absorbance of resultant mixture was read at 390 nm.
2.15. Estimation of Different Amino Acids
2.15.1. Glutamate (Glu) Determination
L-glutamate concentrations in phosphate buffer extracted samples were assayed following Beutler and Michal [
54]. The basic mechanism was the formation of 2-oxoglutarate by the oxidative deamination in the presence of glutamate dehydrogenase and NAD. Then, the formation of iodonitrotetrazolium chloride (INT) in the presence of diaphorase takes place by oxidative process to a formazan in the presence of NADH. Then the absorbance of INT was read at 492 nm. The samples were deproteinized using 20% PCA. The reaction mixture was comprised 3.5 mL sample, 500 μL triethanolamine (57 mM), diaphorase (0.14 i.u./mL), NAD (0.38 mM), INT (0.068 mM), potassium phosphate (14 mM, pH 8.6) and GDH (14 i.u./mL). The absorbance of mixture was read at 492 nm against a reagent blank using spectrophotometer.
2.15.2. Estimation of Leaf Glycine Betaine (GB) Content
The estimation of GB was ascribed following Grieve and Grattan [
55]. Briefly, 1 g dry leaf material was extracted with 10 mL dH
2O by shaking overnight, followed by the filtration. The reaction mixture was prepared by the addition of 1 mL of the extract and 1 mL of 2 N HCl. Then, this prepared mixture (0.5 mL) was mixed with KI
3 solution (0.2 mL). Then, to the ice-cooled mixture, dichloromethane (20 mL) and ice-cooled H
2O (2 mL) were added. Then, the mixture was shook well with a continuous passing of air. The absorbance of lower phase was read at 365 nm and the concentration of GB was calculated using a curve prepared from known standards.
2.15.3. Estimation of Leaf Proline (Pro) Content
The method ascribed by Bates et al. [
56] was followed for the estimation of proline. Briefly, 0.1 g of leaf (fresh material) was crushed in 5 mL of sulfosalicylic acid (3%). After filtration, 100 µL of the extract was mixed well with 20 mL of 6 M phosphoric acid (2 mL each). Then, the mixture was reacted with glacial acetic acid (2 mL of each) and acidic ninhydrin heated the mixture in a water bath for 1 h at 95 °C. After cooling well, 1 mL of toluene was mixed with reaction mixture and the optical density of colored phase was read at 520 nm. Proline concentration was measured following the equation:
2.15.4. Estimation of Lysine (Lys) and Methionine (Meth) Contents
Leaf samples (0.5 g) were well grinded in a mortar containing 0.1% HCl. The resulted solution was then mixed with 3 mL of 50% glycerol, 1 mL phosphate buffer and 1 mL ninhydrin in order to extract lysine. It was then placed in boiling water (100 °C). The absorption was read at 570 nm. Methionine was also extracted by adding NaOH (5 N), glycine dihydrate (50%), sodium nitroferricyanide dihydrate (0.1%) and HCl (1:1) to the resulted solution as mentioned above and the absorption was read at 510 nm [
57].
2.16. Estimation of Oxidized Glutathione (GSH) and Reduced Glutathione (GSSG)
The contents of oxidized (GSSG) and reduced (GSH) glutathione in ajwain leaves were assayed following Griffith [
58]. Briefly, fresh leaf material (250 mg) was ground well in 0.1 M HCl (2 mL) containing EDTA (1 mM). Then, the supernatant was obtained by the centrifugation of extract at 12,000×
g for 15 min at 4 °C. The reaction mixture was prepared by adding 200 μL phosphate buffer (125 μM) having 6.3 mM EDTA (pH 7.5), 100 μL DTNB (6.0 mM), 200 μL extract and 500 μL NADPH (0.3 mM). The absorbance of mixture was read at 412 nm.
2.17. Determination of Yield Attributes
At maturity, different yield attributes such as 100 seed weight, number of umbels (NOU), and seed yield per plant were estimated manually. At physiological maturity, 10 plants from each replicate were selected for the estimation of yield attributes. Number of umbels were counted from each plant and separated from the plant. The seeds were then separated by hands by slight rubbing. Then, 100 seeds were separated and weighed. Seed yield per plant was estimated by weighing the obtained seeds from each selected plant.
2.18. Determination of Nutrients in Roots and Shoots
Powdered samples were digested using sulphuric acid as ascribed by Wolf [
59]. Contents of different cations like Ca
+, K
+, Mg
2+ and Fe
2+ were determined by AAS (Hitachi, Model 7JO-8024, Tokyo, Japan). Phosphorus content was determined spectrophotometrically using Barton’s reagent. The content of (N) was determined using the method as ascribed by Bremner and Keeney [
60] from the digested material.
2.19. Statistical Analysis
Data recorded for varying studied parameters were analyzed statistically using Co-STAT version 6.3 (developed by Cohort Software Berkley, CA, USA) to find out significant difference among the treatments. To find out the significant difference among mean values of different treatments both under stressed and non-stressed conditions, least significance difference (LSD) test was employed at 5% level of significance. Correlations and PCA analysis of the studied parameters were performed using the XLSTAT software and the significance among the generated values of each attribute was found using the Spearman’s correlation table.
3. Results
Water stress adversely affected different morphological and growth attributes such as root length (RL), shoot length (SL), shoot fresh weight (SFW), shoot dry weight (SDW), root fresh weight (RFW), and root dry weight (RDW) of ajwain plants (
Figure 2;
Table 1). Exogenously-applied Fe-Glu and FeSO
4 as foliar spray was found effective in improving all studied morphological and growth attributes. More improvement in these morphological and growth attributes was due to foliar spray with both levels of Fe-Glu in comparison with FeSO
4. Furthermore, both levels of Fe-Glu were found equally effective in enhancing plant performance and reducing the deleterious effects of water stress on studied growth attributes. Similarly, both levels of FeSO
4 were also found equally effective but the effectiveness was less than Fe-Glu.
A significant reduction due to imposition of water stress was recorded in number of umbels (NOU), 100 seed weight and seed yield per plant of ajwain as presented in
Table 1. Foliar spray of both levels of Fe-Glu and FeSO
4 were found helpful in enhancing plant performance under water stress and non-stressed conditions. They further reduced the negative impact of water shortage on these yield parameters of ajwain plants but both the levels of Fe-Glu were found more effective. More increase regarding yield per plant under water stress was found due to foliar spray with Fe-Glu (26.17 and 36.40% increase at 0.25 and 0.50% levels, respectively) in comparison with FeSO
4 (5.32 and 4.91% increase at 0.25 and 0.50% levels, respectively). Similarly, under non-stressed conditions, this increase in seed yield was more in plants sprayed with Fe-Glu (20.61 and 18.62% increase at 0.25 and 0.50% levels, respectively) as compared with plants supplied with FeSO
4 (6.76 and 8.78% increase at 0.25 and 0.50% levels, respectively). Both levels of Fe-Glu were found more effective in comparison with FeSO
4 in reducing the deleterious effects of water deficiency on these yield attributes of ajwain plants (
Table 1).
Leaf Chl.
a, Chl.
b, Car. and total Chl. contents of ajwain plants increased significantly when grown under water deficit conditions. Exogenous application of only Fe-Glu as foliar spray at both levels significantly improved the leaf Chl.
a,
b and total Chl. content, both under stressed and non-stressed conditions, but in the case of leaf Chl.
a, both levels of FeSO
4 were also found effective. However, in the case of leaf Car. foliar spray, both levels of FeSO
4 and Fe-Glu significantly improved the leaf Car. both under stressed and non-stressed conditions. Under water-stressed conditions, both levels of FeSO
4 or Fe-Glu were found equally effective but in the case of non-stressed conditions, this increase in leaf Car. was slightly more higher of Fe-Glu. Moreover, a non-significant effect of water stress was found on leaf Chl.
a/b. Foliar spray of both levels of FeSO
4 and Fe-Glu significantly improved the leaf Chl.
a/b but this increasing effect was found only in non-stressed plants and the higher level of FeSO
4 was more effective, but under water deficit conditions, a non-significant decrease in leaf Chl.
a/
b was found due to foliar spray of Fe-Glu or FeSO
4 (
Table 2).
Imposition of water stress significantly increased the leaf T. antho, TSS and RS contents of ajwain plants. Exogenous application of FeSO
4 and Fe-Glu as foliar spray further significantly improved the contents of T. antho, TSS and RS, both under stressed and non-stressed conditions. However, foliar spray of Fe-Glu was found comparatively more effective (
Table 2).
Imposition of water stress significantly improved the contents of leaf Asp, Glu, Meth, GB and Pro in leaf of ajwain plants but this increase was not found in the case of Lys content, which remained unaffected. Foliar applied at both levels of only the Fe-Glu further improved the accumulation of Glu, Meth, GB and Pro, except to that of leaf Meth and Lys. In the case of Asp, foliar spray of both of the FeSO
4 and Fe-Asp were found effective in increasing the content but slightly more was due to higher level of Fe-Asp. In the case of Lys, more increase was found in plants sprayed with higher level of FeSO
4. Furthermore, in the case of leaf GB content, slightly more accumulation was found due to foliar spray of lower level (0.25%) of Fe-Glu (
Table 3).
Uptake of N, P, K and Fe in root and shoot of ajwain pants decreased significantly due to imposition of water stress (
Table 4). Foliar-applied Fe-Glu and FeSO
4 significantly reduced the deleterious effects of water stress on the uptake of N, P, K and Fe in both root and shoot. Both levels of FeSO
4 and Fe-Glu were found equally effective in improving the uptake of N, P and K in shoot and root under limited water supply except to that of root P and Fe content in root and shoot that was significantly higher in water-stressed ajwain plants—supplied with both levels of Fe-Glu as compared with FeSO
4. Root and shoot Ca and Mg contents also reduced significantly due to water stress. However, an improvement in the uptake of Ca and Mg in root and shoot was recorded in water-stressed ajwain plants—supplied with both levels of Fe-Glu only, and the effect was the same at both the levels of Fe-Glu.
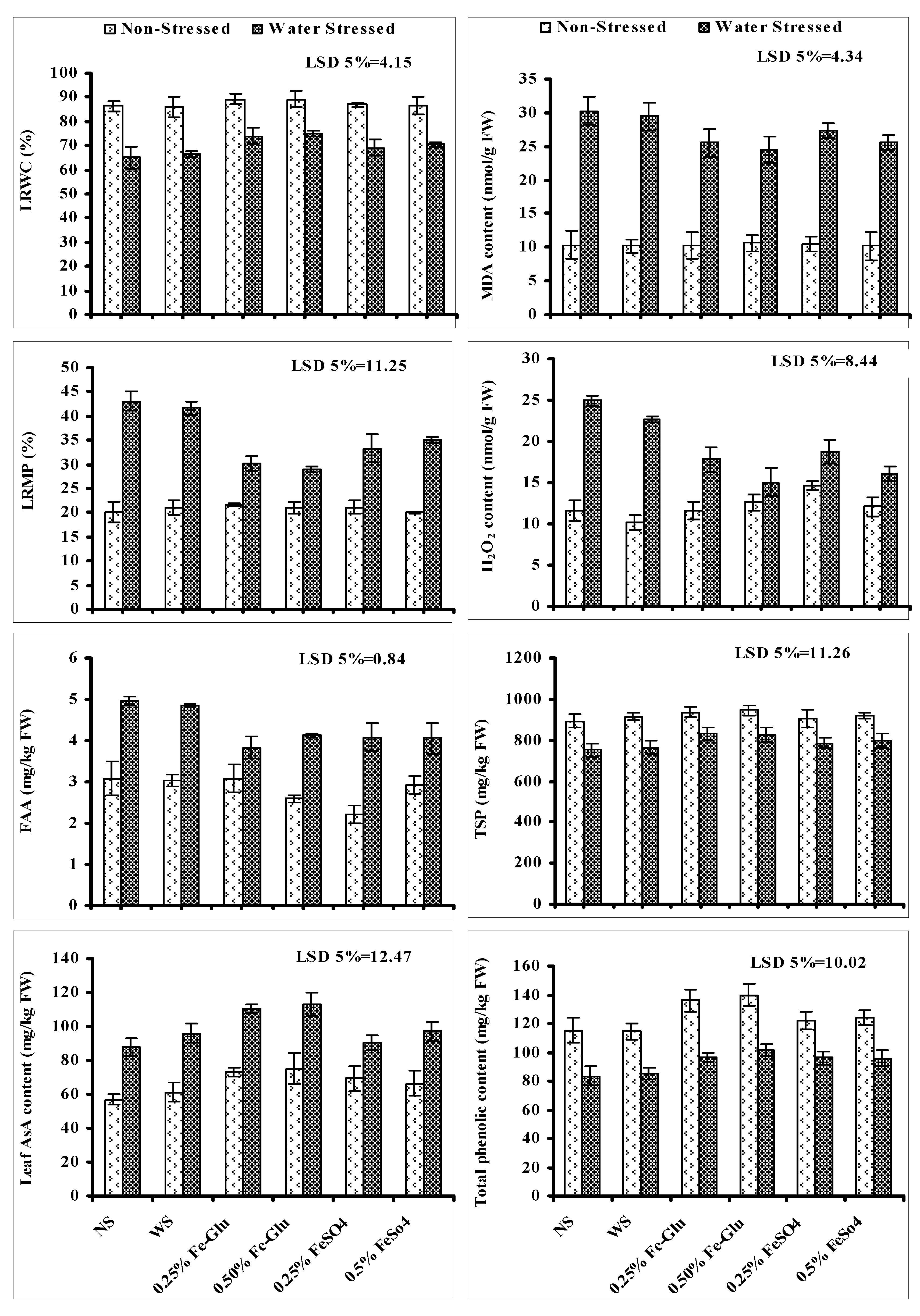
A significant reduction was recorded in LRWC, TSP, as well as the content of TPC of ajwain plants due to imposition of water stress. Foliar application of Fe-Glu was found effective in reducing the negative impacts of water deficit conditions on these attributes. Both levels of Fe-Glu were found more helpful in increasing the LRWC, TSP and TPC content in comparison to FeSO
4 of water-stressed ajwain plants (
Figure 3).
Significant increase in LRMP, leaf H
2O
2 and MDA accumulation was recorded in ajwain plants when grown under water stress. Foliar applied at both levels of FeSO
4 and Fe-glu significantly reduced the leaf LRMP, H
2O
2 and MDA accumulation in water-stressed ajwain plants. Both levels of Fe-Glu were found more effective in reducing these parameters in comparison to both levels of FeSO
4 under limited water supply. In the case of H
2O
2, the higher levels of both Fe-Glu and FeSO
4 were found more effective, but in the case of LRMP, higher level of Fe-Glu was better in this regard, while the opposite was true for FeSO
4 (
Figure 3).
Imposition of stress significantly increased the leaf AsA accumulation in ajwain plants. Treating plants with different levels of Fe-Glu was only found effective in improving the accumulation of leaf AsA in water-stressed ajwain plants. Both levels of Fe-Glu were found equally effective in further improving the leaf AsA content. However, under non-stress conditions, foliar spray of both FeSO
4 and Fe-Glu increased the accumulation of AsA and more increment was found in ajwain plants —supplied with both levels of Fe-Glu in comparison with FeSO
4 (
Figure 3).
Accumulation of leaf FFA increased significantly in water-stressed ajwain plants. Foliar-applied Fe-Glu and FeSO4 significantly reduced the accumulation of FAA in water-stressed ajwain plants. Both levels of FeSO4 and Fe-Glu were found equally effective in reducing the accumulation of FAA in water-stressed ajwain plants.
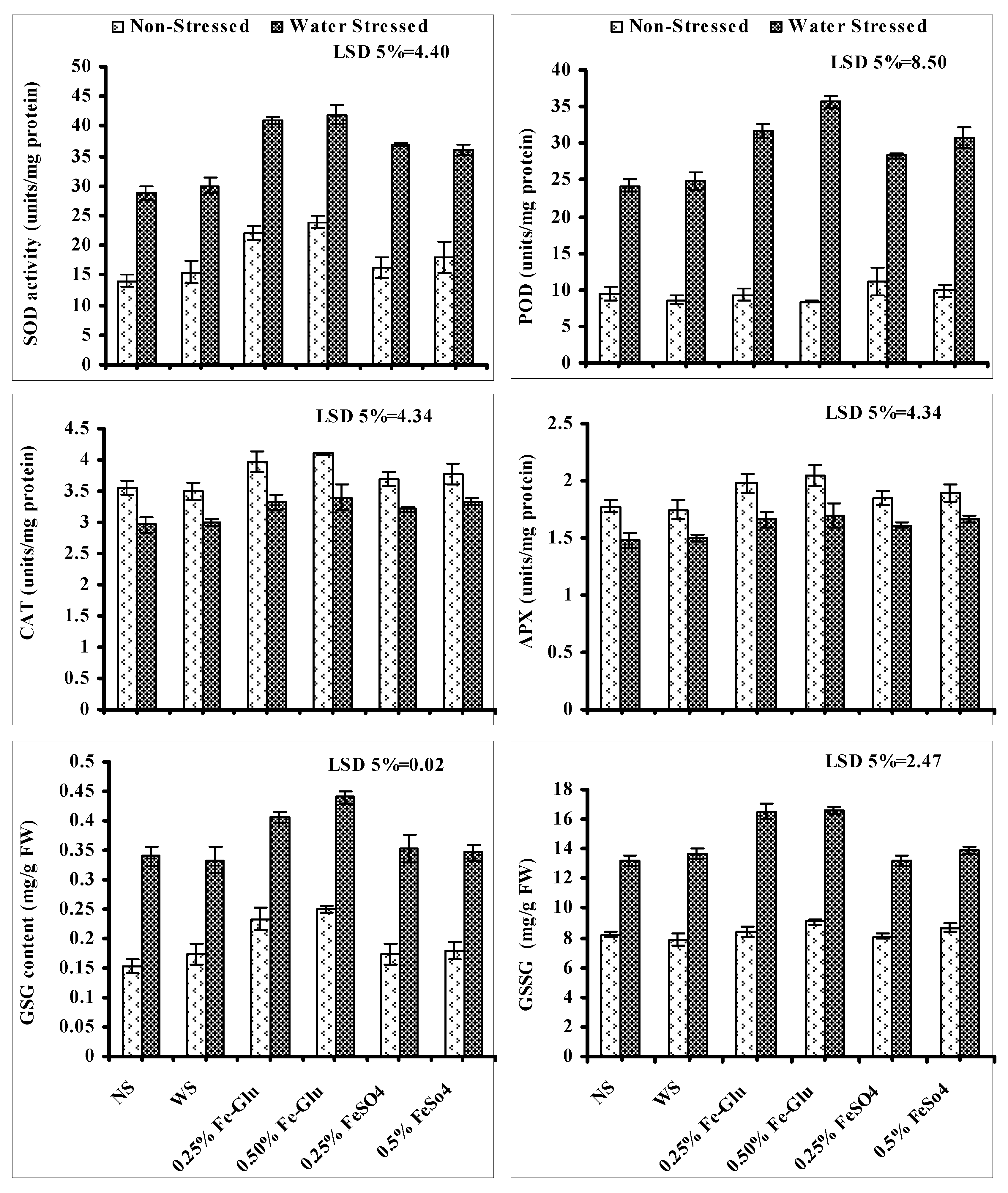
Activities of POD and SOD and the contents of GSG and GSSG increased significantly in water-stressed ajwain plants. Foliar application of both levels of FeSO
4 and Fe-Glu further improved the POD and SOD activities, and GSG and GSSG accumulation. More increase in the activities of SOD and POD, and contents of GSG and GSSG were found in plants treated with both levels of Fe-Glu in comparison with FeSO
4. The higher levels were found more effective except in the case of GSSG where both levels of FeSO
4 and Fe-Glu were found equally effective in increasing the content of GSSG (
Figure 4).
Leaf CAT and APX activities of water-stressed ajwain significantly decreased. Exogenously-applied different levels of Fe-Glu and FeSO
4 were found effective in decreasing the adverse impacts of water shortage on the activities of CAT and APX. Both levels of Fe-Glu were found more effective in reducing the negative impacts of drought on APX and CAT activities in comparison with both levels of FeSO
4 (
Figure 4).
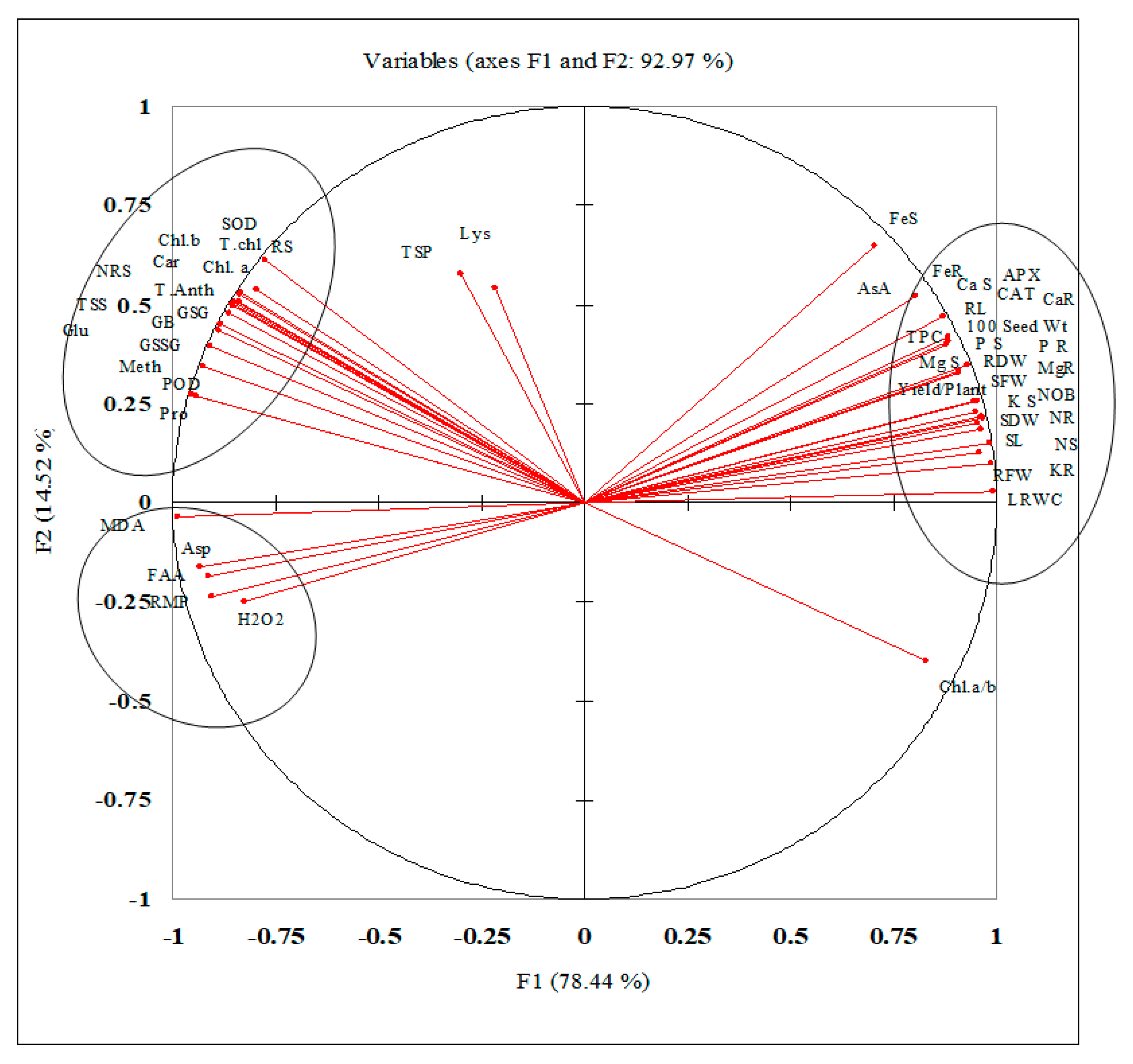
The correlation studies and PCA analysis for different studied attributes are being presented in
Table A1 (in
Appendix A) and
Figure 5. The data presented in the table shows that biomass production and yield has strong positive correlation with different antioxidative compounds such as TSP (0.962*** and 0.968***), AsA (0.932*** and 0.931***), CAT (0.932*** and 0.931***), APX (0.932*** and 0.931***) and nutrient uptake, while a negative correlation was with LRMP (−0.945*** and −0.923***) and MDA (−0.959*** and −0.963***) content. The PCA analysis shows that the studied attributes have been categorized into two major components, F1 and F2, that have major contribution in defining the variance among different studied attributes. Both components (F1 and F2) have a total contribution of 92.97%. The component F1 showed 78.44% contribution while the component F2 has 14.52% contribution in defining the variance among the attributes. The component F1 categorized the studied attributes in three major groups encircled on both sides of F1 axis. The parameters given encircled are closely positively correlated to each other (
Figure 5).
4. Discussion
For the induction of plant performance and drought tolerance, various strategies are being used for enhancing seed yield. Exogenous application of different chemicals (organic/inorganic) is found to be an effective technique being employed, including their exogenous applications through foliar spray [
17]. Studies depict that application of these chemicals as foliar sprays is being absorbed and take part in different cellular metabolic activities through their translocation to different plant parts, in order to induce stress tolerance [
61] and up-regulate their own metabolism [
7,
61,
62]. However, in the case of using chemical fertilizers as foliar sprays, the mobility and availability after their application is a problem [
20]. So, the use of these chelated compounds with improved uptake and mobilization lead to better utilization of applied compounds [
24,
63]. In chelation, both parts of the compound play key roles in a better way as compared with their individual application because the oxidation chances of metals increase [
21,
24,
28].
In the present study, an increase in glutamate and Fe contents was recorded both in root and shoot, when applied individually in chelated form. Furthermore, the mobility of Fe
2+ was higher in case of Fe-Glu application as a foliar spray, as compared with FeSO
4. This is because Fe-Glu supplied ajwain plants with more content of Fe which was found in root and shoot. This also confirms its lower oxidation when sprayed as Fe-Glu. Additionally, the increment in Fe was higher in the case of Fe-chelated glutamate as compared with FeSO
4, showing a better way to overcome plant Fe deficiency. Moreover, this increased accumulation of Fe
2+ and glutamate in root and shoot further describes their long-term translocation after its foliar application. In earlier studies, it has been reported that micronutrient-chelated amino acids after their absorption are taken as an amino acid, translocated to sink tissues in phloem tissue [
64,
65]. It has also been found that chelated amino acids in particular can increase the uptake and translocation of chelated metals [
24,
66], whereas metals in chemical form have normally low mobility and translocation within the plant [
27].
In the present study, an improvement in yield and biomass product of ajwain plants is recorded with the application of both Fe
2+ sources and is being found more in plants fertigated with Fe-Glu in comparison to FeSO
4. This improvement in water stress tolerance in terms of yield and growth might be due to the better uptake of Fe and the availability of glutamate. It is well known that the best yield is depending on the higher biomass production as found in the present study, where the increased yield improvement was found in ajwain plants fertigated with Fe-Glu rather than FeSO
4. It might be the fact that glutamate might also have played an additional role in improving the water stress tolerance of ajwain plants [
35]. It has been found earlier that foliar-supplied glutamate was found helpful in enhancing plant performance and alleviating the harmful effects of stress in faba bean and
Brassica napus plants [
33,
35], respectively. In rice, Jie et al. [
67] reported that amino acid- chelated Zn and Fe better improved the yield as compared with respective chemical fertilizers. In a field experiment conducted on onion, it was found that onion plants supplied with Zn-chelated amino acids performed better in terms of biomass production and yield as compared with ZnSO
4 [
68].
Normally, a better growth and seed yield is associated with a balanced plant water status, a significant and foremost effect that a plant faces under multiple abiotic stresses [
69,
70]. Similarly, in the present study, under applied water stress, a reduction in LRWC was found in ajwain plants. An improvement in LRWC due to foliar-applied FeSO
4 and Fe-Glu was recorded but comparatively was found better in ajwain plants supplied with Fe-Glu. This more improvement in LRWC due to Fe-Glu foliar application might be the role of glutamate in cellular water relations [
71] or its role in osmotic adjustment itself, or through its involvement in biosynthesis and accumulation of other amino acids that are important in improving the plant water relations through osmotic adjustment [
31]. It is well known that glutamate is involved indirectly in the proline biosynthesis under stressful conditions [
31,
71]. In the present study, Fe-Glu significantly improved the plant water relations, i.e., with increased accumulation of amino acids including proline and GB that might improve the plant cellular water status along with nutrient uptake [
21,
61]. Furthermore, this improvement in LRWC of ajwain plants is positively related with the improved growth and seed yield.
The better growth and seed yield are also associated with the better photosynthetic activity that is directly linked with better water relations and efficient photosynthetic pigments [
72,
73]. An increase in leaf Chl.
a,
b and total Chl. was recorded in ajwain plants, representing the defensive response of plants under stressful environment [
74,
75]. Rahbarian et al. [
73] reported that increase or decrease in leaf chlorophyll and carotenoid contents under water deficit conditions is genotype- specific. In the present investigation, foliar-applied FeSO
4 and Fe-Glu improved the biosynthesis of light-capturing compounds including Chl.
a,
b, total Chl. as well as Car. More significant improvement in this regard was found in plants sprayed with Fe-Glu as compared with FeSO
4. The similar findings have been reported in earlier studies, where an increase in photosynthetic rate (44%) of peanut plant was recorded along with increased biosynthesis of chlorophyll pigments, upon fertigation with Fe [
76]. Similarly, foliar application of Fe also improved the biosynthesis of Chl.
a,
b and total Chl. in groundnut [
77]. Furthermore, it is known that glutamate especially have a role in chlorophyll biosynthesis [
78] because it was reported that plant glutamate improved the leaf Chl. biosynthesis through lush green appearance of leaves [
32]. Similar pattern might be found in the present findings where foliar-applied Fe-Glu improved the biosynthesis of Chl.
a,
b, total Chl. as well as of Car. that might have helped in better leaf photosynthetic activity by increasing the light- capturing efficiency [
72,
73,
76] along with a better stomatal regulation by the maintenance of plant cellular water content through osmotic adjustment [
73], leading to better biomass production and seed yield. However, the studies reveal that the improvement in leaf photosynthetic pigments is not necessarily an indication of better photosynthetic efficiency because it also depends on the type of species and available light intensity [
72]. Photosynthetic rate has been associated with a number of different factors in different crops, including stomatal and mesophyll resistance, RuDP carboxylase activity, and mesophyll cell size [
79], as well as chlorophyll content [
80,
81] and chloroplast numbers [
82]. The improvement in the biosynthesis of photosynthetic pigments in plants sprayed with Fe-Glu might be due to better Fe availability [
21,
76], less oxidation, as well as the additional availability of the glutamate that resulted in increased capacity to absorb sunlight.
In the present study, better growth and seed yield of water-stressed ajwain is also associated with better antioxidative defense mechanisms. Normally, lipid peroxidation is a common phenomenon in plants and its severity increases under stressful environments due to the over- production of reactive oxygen species [
83]. To counteract the adversities of over-produced ROS, plants have a well-organized ROS revenging mechanism to reduce the membrane lipid peroxidation and oxidative damages to the other macromolecules [
84,
85,
86,
87]. The extent of oxidative damages due to ROS is being estimated by measuring the production of MDA or ROS. In the present study, under water stress, membrane lipid peroxidation was increased as measured in term MDA accumulation but decreased accumulation of MDA was found in plants sprayed with FeSO
4 and Fe-Glu. The more decrease was recorded in Fe-Glu-sprayed ajwain plants. This decrease in lipid peroxidation in the present study is associated with increased activities of antioxidative enzymes including CAT, POD, SOD, APX as well as the increased accumulation of AsA, TPC, phenolics, GSG and GSSG as well- known non-enzymatic antioxidant in water-stressed ajwain plants sprayed with FeSO
4 and Fe-Glu with a reduced accumulation of MDA, showing the role of these compounds in improving the antioxidative defense mechanism with reduced lipid per oxidation leading to improved biomass and seed yield. The more increase in antioxidative defense mechanism was in water-stressed ajwain plants foliar-supplied with Fe-Glu as compared with FeSO
4, showing a better potential of Fe-Glu in improving the antioxidative defense mechanism. It might be the fact that iron acts as a prosthetic group of many peroxidases and catalases [
88]. The reduced lipid peroxidation of the plants sprayed with Fe-Glu comparative with FeSO
4 shows the additional properties of glutamate as a signaling molecule [
35].
Plant nutrient acquisition also plays an important role in enhancing plant performance and counteracting adverse effects of water stress via regulating cellular metabolic activities, leading to better biomass and seed yield. Due to the soil lower osmotic potential under limited water supply, the disturbance in nutrient uptake is a common phenomenon that not only disturbs the cellular osmotic potential and the process of assimilation but also the final crop yield [
89]. In the present study, water stress reduced nutrient acquisition of ajwain plants in terms of its uptake from soil and root to the upper part that is cleared from the reduced content of studied nutrients in roots and shoots. Foliar-supplied Fe-Glu and FeSO
4 significantly improved the nutrient uptake from soil both in shoots and roots and the more uptake was in water-stressed ajwain plants foliarly-supplied with Fe-Glu. This might be due to the improved cellular water relations due to the amino acid metabolism. This improvement in the nutrient uptake might be due to the better cellular osmotic adjustment that resulted in more water uptake from soil along with the micro- and macro-nutrients from the rhizosphere in water-stressed ajwain plants. Furthermore, the improvement in nutrient quantity is positively correlated with improved biomass and seed yield. The findings correlate well with the previous findings where foliarly-applied glutamate at 200 ppm increased the nutrient uptake including P, N and K in leaves, stem and roots of
Codiaeum variegatum L. plants that ultimately improved plant growth and yield [
90]. In another study conducted by Ramezani et al. [
91], it was reported that foliarly-applied Fe as FeSO
4 considerably improved the uptake of Zn, Fe and Se in water-stressed alfalfa plants. Moreover, in the present study, foliarly-applied Fe-Glu and FeSO
4 were also found helpful in enhancing plant growth and yield under water stress and normal irrigation conditions.

 ,
, 


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}





