Growth Characteristics and Anti-Wind Erosion Ability of Three Tropical Foredune Pioneer Species for Sand Dune Stabilization

Abstract
:1. Introduction
2. Materials and Methods
2.1. Seed Collection
2.2. Seedling Raising
2.3. Growth Characteristics Observation
2.4. Uprooting Test
2.5. Root Tensile Test
2.6. Wind Tunnel Test
2.7. Statistical Data Analysis
3. Results
3.1. Growth Characteristics
3.2. Plant Uprooting Resistance
- For I. pes-caprae seedlings, Ur = − 0.263H + 0.026RSA − 72.846 (R2 = 0.910*, r = 0.954, p = 0.027, VIF = 3.151).
- For S. littoreus seedlings, Ur = 0.066H + 0.035RSA − 0.12 (R2 = 0.934*, r = 0.966, p = 0.017, VIF = 2.690).
- For V. rotundifolia seedlings, Ur = 0.115H + 0.212RSA − 7.757 (R2 = 0.990***, r = 0.995, p = 0.001, VIF = 3.167).
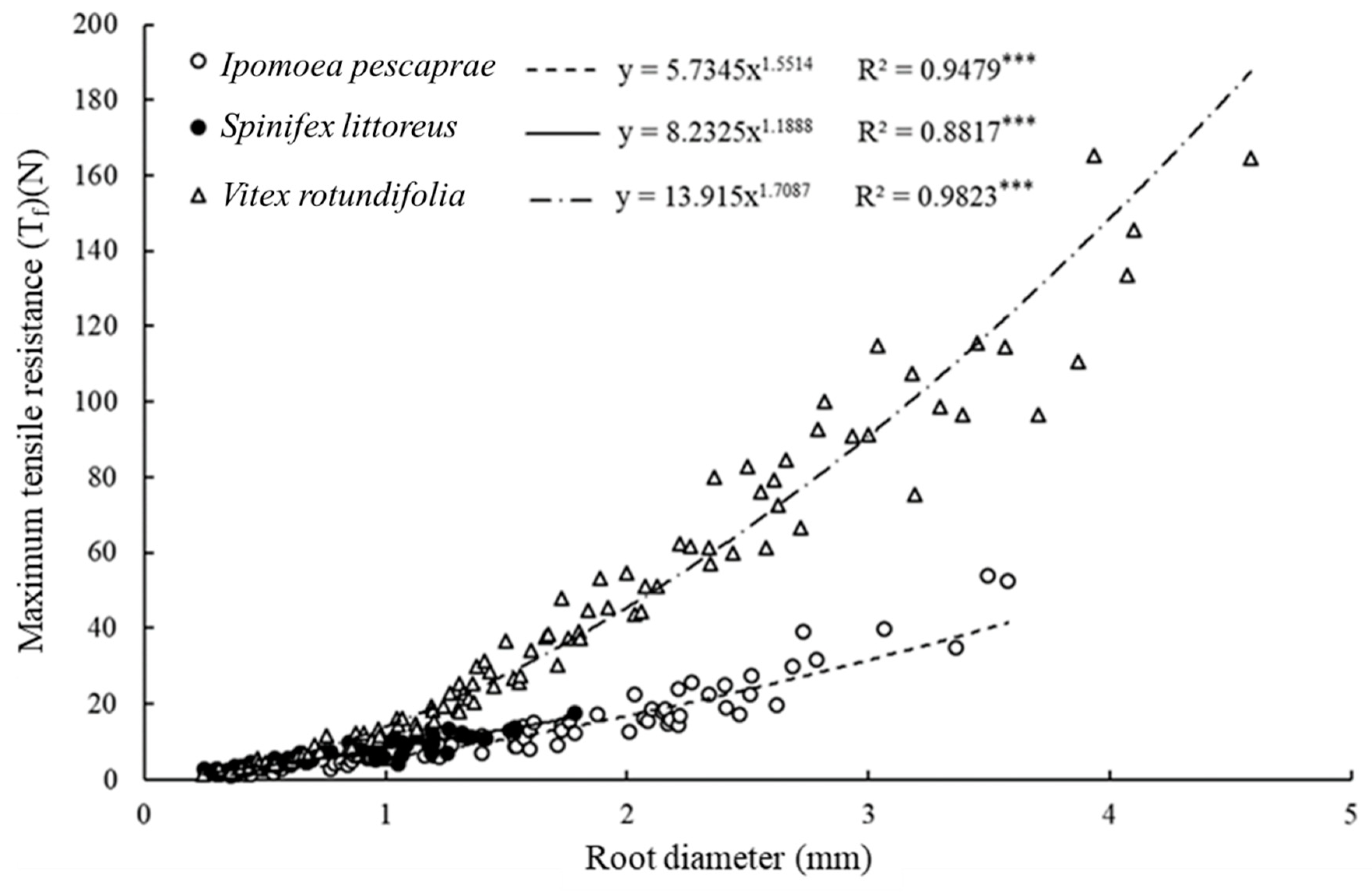
3.3. Root Tensile Strength
3.4. Anti-wind Erosion Ability
4. Discussion
4.1. Growth Characteristics
4.2. Plant Uprooting Resistance
4.3. Root Tensile Strength
4.4. Anti-wind Erosion Ability
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Chang, J.C. Natural hazards in Taiwan. GeoJournal 1996, 38, 251–257. [Google Scholar] [CrossRef]
- Tong, X.L.; Lin, T.Y. Dune restoration experiments during a typhoon season on Taiwan’s Si-Cao coast. J. Mar. Sci. Technol. 2016, 24, 1032–1040. [Google Scholar]
- Huang, W.P.; Yim, J.Z. Sand dune restoration experiments at Bei-Men coast, Taiwan. Ecol. Eng. 2014, 73, 409–420. [Google Scholar] [CrossRef] [Green Version]
- Antunes, C.; Pereira, A.J.; Fernandes, P.; Ramos, M.; Ascensao, L.; Correia1, O.; Maguas, C. Understanding plant drought resistance in a Mediterranean coastal sand dune ecosystem: Differences between native and exotic invasive species. J. Plant Ecol. 2018, 11, 26–38. [Google Scholar] [CrossRef] [Green Version]
- Miri, A.; Dragovich, D.; Dong, Z. Vegetation morphologic and aerodynamic characteristics reduce aeolian erosion. Sci. Rep. 2017, 7, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.; Xiao, H.B.; Li, Z.W.; Liu, C.; Wang, D.Y.; Wang, L.X.; Tang, C.J. Threshold effects of vegetation coverage on soil erosion control in small watersheds of the red soil hilly region in China. Ecol. Eng. 2019, 132, 109–114. [Google Scholar] [CrossRef]
- Capobianco, M.; Stive, M.J.F. Soft intervention technology as a tool for integrated coastal zone management. J. Coast. Conserv. 2000, 6, 33–40. [Google Scholar] [CrossRef]
- Chang, C.E. Ipomoea pes-caprae (L.) R. Br. subsp. Brasiliensis (L.) Oostst. In Flora of Taiwan, 2nd ed.; Editorial Committee of Flora of Taiwan: Taipei, Taiwan, 1998; Volume 4, pp. 367–368. [Google Scholar]
- Hsu, C.C. Spinifex littoreus (Burm. F.) Merr. In Flora of Taiwan, 2nd ed.; Editorial Committee of Flora of Taiwan: Taipei, Taiwan, 2000; Volume 5, pp. 518–519. [Google Scholar]
- Hsiao, J.Y. Vitex rotundigolia L. In Flora of Taiwan, 2nd ed.; Editorial Committee of Flora of Taiwan: Taipei, Taiwan, 1998; Volume 4, p. 428. [Google Scholar]
- Chen, T.H.; Yu, H.M.; Horng, F.W. The movement of shifting sand and the growth of sand stabilizing plants at Shitsugan coast, Taoyuan. Q. J. Chin. For. 2004, 37, 367–377. [Google Scholar]
- Sigren, J.M.; Figlus, J.; Armitage, A.R. Coastal sand dunes and dune vegetation: Restoration, erosion, and storm protection. Shore Beach 2014, 82, 5–12. [Google Scholar]
- Yen, C.P. Tree root patterns and erosion control. In Proceedings of the International Workshop on Soil Erosion and Its Counter-Measures; Jantawat, S., Ed.; Soil Conservation Society of Thailand: Bangkok, Thailand, 1987; pp. 92–111. [Google Scholar]
- Mickovski, S.B.; van Beek, L.P.H.; Salin, F. Uprooting resistance of vetiver grass (Vetiveria zizanioides). Plant Soil 2005, 278, 33–41. [Google Scholar] [CrossRef]
- Saifuddin, M.; Osman, N.; Rahman, M.M.; Boyce, A.N. Soil reinforcement capability of two legume species from plant morphological traits and mechanical properties. Curr. Sci. 2015, 108, 1340–1347. [Google Scholar]
- Lee, J.T.; Chu, M.Y.; Lin, Y.S.; Kung, K.N.; Lin, W.C.; Lee, M.J. Root traits and biomechanical properties of three tropical pioneer tree species for forest restoration in landslide areas. Forests 2020, 11, 179. [Google Scholar] [CrossRef] [Green Version]
- Kamakura, M.; Furukawa, A. Compensatory function for water transport by adventitious roots of Ipomoea pes-caprae. J. Plant Res. 2009, 122, 327–333. [Google Scholar] [CrossRef] [PubMed]
- Divyasree, M.; Raju, A.J.S. An ecological study of reproduction in Spinifex littoreus (Burm. F.) Merr. (Poaceae), a dominant species of sand dune ecosystem along the Visakhapatnam coast, Andhra Pradesh. Palynology 2019, 55, 35–54. [Google Scholar]
- He, D.X. Community features of Vitex rotundifolia var. simplicifolia and its adaptation to sandy land in Houtian area. Chin. J. Ecol. 1992, 11, 36–40. [Google Scholar]
- Li, Y.P.; Wang, Y.Q.; Wang, Y.J.; Ma, C. Effects of Vitex negundo properties on soil resitance caused by pull-out forces at different positions around the stem. Catena 2017, 158, 148–160. [Google Scholar] [CrossRef]
- Wasson, R.J.; Nanninga, P.M. Estimating wind transport of sand on vegetated surfaces. Earth Surf. Process. Landf. 1986, 11, 505–514. [Google Scholar] [CrossRef]
- Lancaster, N.; Baas, A. Influence of vegetation cover on sand transport by wind: Field studies at Owens Lake, California. Earth Surf. Process. Landf. 1998, 23, 69–82. [Google Scholar] [CrossRef]
- Udo, K.; Takewaka, S. Experimental study of blown sand in a vegetated area. J. Coast. Res. 2007, 23, 1175–1182. [Google Scholar] [CrossRef]
- Meng, Z.J.; Dang, X.H.; Gao, Y.; Ren, X.M.; Ding, Y.L.; Wang, M. Interactive effects of wind speed, vegetation coverage and soil moisture in controlling wind erosion in a temperate desert steppe, Inner Mongolia of China. J. Arid Land 2018, 10, 534–547. [Google Scholar] [CrossRef] [Green Version]
- Bouma, T.J.; Nielsen, K.L.; Koutstaal, K. Sample preparation and scanning protocol for computerized analysis of root length and diameter. Plant Soil 2000, 218, 185–196. [Google Scholar] [CrossRef]
- Pang, W.; Crow, W.T.; Luc, J.E.; McSorley, R.; Giblin-Davis, R.M.; Kruse, J.K. Comparison of water displacement and WinRHIZO software for plant root parameter assessment. Plant Dis. 2011, 95, 1308–1310. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Burylo, M.; Dutoit, T.; Rey, F. Species traits as practical tools for ecological restoration of Marly eroded lands. Restor. Ecol. 2014, 22, 633–640. [Google Scholar] [CrossRef]
- Gould, I.J.; Quinton, J.N.; Weigelt, A.; De Deyn, G.B.; Bardgett, R.D. Plant diversity and root traits benefit physical properties key to soil function in grasslands. Ecol. Lett. 2016, 19, 1140–1149. [Google Scholar] [CrossRef] [PubMed]
- Bischetti, G.B.; Chiaradia, E.A.; Simonato, T.; Speziali, B.; Vitali, B.; Vullo, P.; Zocco, A. Root strength and root area ratio of forest species in Lombardy (Northern Italy). Plant Soil 2005, 278, 11–22. [Google Scholar] [CrossRef]
- Genet, M.; Kokutse, N.; Stokes, A.; Fourcaud, T.; Caim, X.; Ji, J.; Mickovski, S.B. Root reinforcement in plantations of Cryptomeria japonica D. Don: Effect of tree age and stand structure on slope stability. For. Ecol. Manag. 2008, 256, 1517–1526. [Google Scholar] [CrossRef]
- De Baets, S.; Poeson, J.; Reubens, B.; Wemans, K.; Baerdemaeker, J.D.; Muys, B. Root tensile strength and root distribution of typical Mediterranean plant species and their contribution to soil shear strength. Plant Soil 2008, 305, 207–226. [Google Scholar] [CrossRef]
- Yang, Y.; Chen, L.; Li, N.; Zhang, Q. Effect of root moisture content and diameter on root tensile properties. PLoS ONE 2016, 11, e0151791. [Google Scholar] [CrossRef]
- Gray, D.H.; Sotir, R.B. Role of vegetation in the stability of slopes. In Biotechnical and Soil Bioengineering Slope Stabilization; Gray, D.H., Sotir, R.B., Eds.; John Wiley & Sons: New York, NY, USA, 1996; pp. 54–105. [Google Scholar]
- Lee, J.T.; Yen, L.Z.; Lee, M.J. Wind affects the growth, root anchorage and tensile strength of Australian pine (Casuarina equisetifolia) seedlings. J. For. Res. 2019, 24, 219–229. [Google Scholar] [CrossRef]
- Lee, F.C.; Yu, J.L.; Chiang, W.S.; Wen, J.T. Field measurements of aeolian sand transport at Yunlin coastal area. In Proceedings of the 25th Ocean Engineering Conference; National Taiwan Ocean University: Keelung, Taiwan, 2003; pp. 677–684. [Google Scholar]
- Lin, S.H.; Chiang, Y.C. Test on the relationships between sand moisture and its erodibility by wind. J. Chin. Soil Water Conserv. 1979, 10, 173–193. [Google Scholar]
- Huang, L.M.; Wu, C.H. A study on the estimate wind-blown sand in Taichung harbor. J. Chin. Soil Water Conserv. 2010, 42, 393–407. [Google Scholar]
- Davis, A.S.; Jacobs, D.F. Quantifying root system quality of nursery seedlings and relationship to outplanting performance. New For. 2005, 30, 295–311. [Google Scholar] [CrossRef]
- Morrissey, R.C.; Jacobs, D.F.; Davis, A.S.; Rathfon, R.A. Survival and competitiveness of Quercus rubra regeneration associated with planting stocktype and harvest opening intensity. New For. 2010, 40, 273–287. [Google Scholar] [CrossRef]
- Tsakaldimi, M.; Ganatsas, P.; Jacobs, D.F. Prediction of planted seedling survival of five Mediterranean species based on initial seedling morphology. New For. 2013, 44, 327–339. [Google Scholar] [CrossRef]
- Truong, P.; Loch, R. Vetiver system for erosion and sediment control. In Proceedings of the 13th International Soil Conservation Organization Conference, Brisbane, Australia, 1–4 July 2004; pp. 1–6. [Google Scholar]
- Ali, F. Use of vegetation for slope protection: Root biomechanical properties of some tropical seedlings. Int. J. Phys. Sci. 2010, 5, 496–506. [Google Scholar]
- Reubens, B.; Poesen, J.; Danjon, F.; Geudens, G.; Muys, B. The role of fine and coarse roots in shallow slope stability and soil erosion control with a focus on root system architecture: A review. Trees-Struct. Funct. 2007, 21, 385–402. [Google Scholar] [CrossRef]
- Fan, C.C.; Chen, Y.W. The effect of root architecture on the shearing resistance of root-permeated soils. Ecol. Eng. 2010, 36, 813–826. [Google Scholar] [CrossRef]
- Forde, B.G. Nitrogen signalling pathways shaping root system architecture: An update. Curr. Opin. Plant Biol. 2014, 21, 30–36. [Google Scholar] [CrossRef]
- Kiba, T.; Krapp, A. Plant nitrogen acquisition under low availability: Regulation of uptake and root architecture. Plant Cell Physiol. 2016, 57, 707–714. [Google Scholar] [CrossRef] [Green Version]
- Wasson, A.; Richards, R.; Chatrath, R.; Misra, S.; Prasad, S.S.; Rebetzke, G.; Kirkegaard, J.; Christopher, J.; Watt, M. Traits and selection strategies to improve root systems and water uptake in water-limited wheat crops. J. Exp. Bot. 2012, 63, 3485–3498. [Google Scholar] [CrossRef] [Green Version]
- Liu, K.; He, A.; Ye, C.; Liu, S.; Lu, J.; Gao, M.; Fan, Y.; Lu, B.; Tian, X.; Zhang, Y. Root morphological traits and spatial distribution under different nitrogen treatments and their relationship with grain yield in super hybrid rice. Sci. Rep. 2018, 8, 131. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Comas, L.H.; Becker, S.R.; Cruz, V.M.V.; Byrne, P.F.; Dierig, D.A. Root traits contributing to plant productivity under drought. Front. Plant Sci. 2013, 4, 442. [Google Scholar] [CrossRef] [Green Version]
- Kramer-Walter, K.R.; Bellingham, P.J.; Millar, T.R.; Smissen, R.D.; Richardson, S.J.; Laughlin, D.C. Root traits are multidimensional: Specific root length is independent from root tissue density and the plant economic spectrum. J. Ecol. 2016, 104, 1299–1310. [Google Scholar] [CrossRef]
- Li, D.; Nan, H.; Liang, J.; Cheng, X.; Zhao, C.Z.; Yin, H.J.; Yin, C.Y.; Liu, Q. Responses of nutrient capture and fine root morphology of subalpine coniferous tree Picea asperata to nutrient heterogeneity and competition. PLoS ONE 2017, 12, e0187496. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wendling, M.; Büchi, L.; Amossé, C.; Sinaj, S.; Walter, A.; Charles, R. Influence of root and leaf traits on the uptake of nutrients in cover crops. Plant Soil 2016, 409, 419–434. [Google Scholar] [CrossRef] [Green Version]
- Burylo, M.; Rey, F.; Roumet, C.; Buisson, E.; Dutoit, T. Linking plant morphological traits to uprooting resistance in eroded marly lands (Southern Alps, France). Plant Soil 2009, 324, 31–42. [Google Scholar] [CrossRef]
- Burylo, M.; Rey, F.; Mathys, N.; Dutoit, T. Plant root traits affecting the resistance of soils to concentrated flow erosion. Earth Surf. Process. Landf. 2012, 37, 1463–1470. [Google Scholar] [CrossRef]
- Katuwal, S.; Vermang, J.; Cornelis, W.M.; Gabriels, D.; Moldrup, P.; De Jonge, L.W. Effect of root density on erosion and erodibility of a loamy soil under simulated rain. Soil Sci. 2013, 178, 29–36. [Google Scholar] [CrossRef]
- Liu, Y.; Jia, Z.; Gu, L.; Gao, J. Vertical and lateral uprooting resistance of Salix matsudana Koidz in a riparian area. For. Chron. 2013, 89, 162–168. [Google Scholar] [CrossRef]
- Lee, J.T.; Tsai, S.M.; Lee, M.J. Uprooting resistance of two tropical tree species for sand dune stabilization. Afr. J. Agric. Res. 2017, 12, 3214–3220. [Google Scholar]
- Roering, J.J.; Schmidt, K.M.; Stock, J.D.; William, E.D.; David, R.M. Shallow landsliding, root reinforcement, and the spatial distribution of trees in the Oregon Coast Range. Can. Geotech. J. 2003, 40, 237–253. [Google Scholar] [CrossRef] [Green Version]
- Tardío, G.; Mickovski, S.B. Method for synchronisation of soil and root behaviour for assessment of stability of vegetated slopes. Ecol. Eng. 2015, 82, 222–230. [Google Scholar] [CrossRef] [Green Version]
- Tsige, D.; Senadheera, S.; Talema, A. Stability analysis of plant-root-reinforced shallow slopes along mountainous road corridors based on numerical modeling. Geosciences 2020, 10, 19. [Google Scholar] [CrossRef] [Green Version]
- Stokes, A.; Atger, C.; Bengough, A.G.; Fourcaud, T.; Sidle, R.C. Desirable plant root traits for protecting natural and engineered slopes against landslides. Plant Soil 2009, 324, 1–30. [Google Scholar] [CrossRef]
- Genet, M.; Stokes, A.; Salin, F.; Mickovski, S.B. The influence of cellulose content on tensile strength in tree roots. Plant Soil 2005, 278, 1–9. [Google Scholar] [CrossRef]
- Zhang, C.B.; Chen, L.H.; Jiang, J. Why fine tree roots are stronger than thicker roots: The role of cellulose and lignin in relation to slope stability. Geomorphology 2014, 206, 196–202. [Google Scholar] [CrossRef]
- Hong, C.; Liu, C.; Zou, X.; Li, H.; Kang, L.; Liu, B.; Li, J. Wind erosion rate for vegetated soil cover: A prediction model based on surface shear strength. Catena 2020, 187, 104398. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Growth Characteristics | I. pes-caprae | S. littoreus | V. rotundifolia | One-Way ANOVA (F) |
|---|---|---|---|---|
| H (cm) | 198.1 ± 38.3 a | 103.4 ± 34.1 c | 145.6 ± 31.5 b | 11.187 *** |
| SBD (mm) | 9.1 ± 0.6 a | 7.4 ± 1.6 a | 6.1 ± 2.5 b | 4.366 * |
| RT | 1206 ± 451.6 a | 734.7 ± 229.2 a | 1163.8 ± 816.8 a | 1.326 |
| TRL (cm) | 790 ± 135.3 a | 441.5 ± 150.3 b | 557.5 ± 292.5 b | 4.482 * |
| RB (g) | 9.6 ± 3.8 a | 0.5 ± 0.3 c | 3.9 ± 1.8 b | 11.993 *** |
| SB (g) | 46.5 ± 9.9 a | 9.6 ± 4.8 c | 27.4 ± 9.2 b | 6.306 ** |
| RD (kg m −3) | 0.18 ± 0.07 a | 0.02 ± 0.01 b | 0.07 ± 0.05 b | 12.256 *** |
| RLD (km m −3) | 0.15 ± 0.02 a | 0.08 ± 0.03 b | 0.1 ± 0.05 b | 4.515 * |
| RSA (cm2) | 696.5 ± 194.16 a | 187.43 ± 87.78 b | 304.71 ± 230.72 b | 12.969 *** |
| RTD (g cm −3) | 0.19 ± 0.06 a | 0.07 ± 0.03 b | 0.26 ± 0.11 a | 9.750 ** |
| RV (cm3) | 57.67 ± 34.25 a | 6.85 ± 4.51 b | 11.79 ± 9.27 b | 11.047 *** |
| SRL (m g−1) | 0.89 ± 0.23 c | 11.41 ± 4.78 a | 3.98 ± 0.91 b | 13.777 *** |
| Root Parameters | I. pes-caprae | S. littoreus | V. rotundifolia | One-Way ANOVA (F) |
|---|---|---|---|---|
| Root diameter (mm) | 1.61 ± 0.79 a | 0.76 ± 0.39 b | 1.73 ± 1.02 a | 34.270 *** |
| Tensile resistance force (N) | 13.68 ± 11.3 b | 6.24 ± 3.86 c | 42.69 ± 39.38 a | 49.477 *** |
| Tensile strength (MPa) | 6.65 ± 2.39 b | 16.68 ± 8.88 a | 16.48 ± 4.37 a | 76.298 *** |
| Root Diameter (mm) | Tensile Strength (MPa) | ||
|---|---|---|---|
| I. pes-caprae | S. littoreus | V. rotundifolia | |
| 0–1 | 9.38 ± 0.49 b | 19.76 ± 1.17 a | 20.53 ± 0.98 a |
| 1–2 | 6.18 ± 0.27 c | 8.76 ± 0.48 b | 16.37 ± 0.36 a |
| 2–5 | 4.97 ± 0.18 b | − | 13.37 ± 0.4 a |
| Species | Leeward Soil Surface Wind Speed (m s−1) | ||||
|---|---|---|---|---|---|
| Vegetation Cover | |||||
| 0% | 20% | 40% | 60% | 80% | |
| I. pescaprae | 6.28 ± 0.32 a 6.28 ± 0.32 a 6.28 ± 0.32 a | 4.58 ± 0.07 a 4.15 ± 0.07 b 4.63 ± 0.11 a | 4.02 ± 0.05 b 3.29 ± 0.04 c 4.26 ± 0.04 a | 3.24 ± 0.09 b 2.52 ± 0.05 c 3.63 ± 0.05 a | 2.77 ± 0.04 b 1.38 ± 0.06 c 3.16 ± 0.06 a |
| S. littoreus | |||||
| V. rotundifolia | |||||
| Species | Leeward Soil Surface Wind Speed (m s−1) | ||||
|---|---|---|---|---|---|
| Vegetation Cover | |||||
| 0% | 20% | 40% | 60% | 80% | |
| I. pescaprae | 10.78 ± 0.47 a 10.78 ± 0.47 a 10.78 ± 0.47 a | 6.55 ± 0.13 a 5.79 ± 0.11 b 6.43 ± 0.16 a | 5.5 ± 0.12 a 4.52 ± 0.06 c 5.4 ± 0.08 a | 4.89 ± 0.12 a 3.86 ± 0.09 b 4.82 ± 0.06 a | 4.04 ± 0.08 a 2.05 ± 0.09 b 4.16 ± 0.05 a |
| S. littoreus | |||||
| V. rotundifolia | |||||
| Species | Wind Erosion Rate (g m−2 s−1) | ||||
|---|---|---|---|---|---|
| Vegetation Cover (%) | |||||
| 0% | 20% | 40% | 60% | 80% | |
| I. pes-caprae | 8.21 ± 0.29 a 8.21 ± 0.29 a 8.21 ± 0.29 a | 5.91 ± 0.11 a 3.29 ± 0.05 b 6.02 ± 0.09 a | 3.94 ± 0.04 b 1.39 ± 0.04 c 4.71 ± 0.08 a | 1.09 ± 0.04 b 0.31 ± 0.02 c 1.71 ± 0.05 a | 0.41 ± 0.06 b 0.13 ± 0.01 c 0.58 ± 0.05 a |
| S. littoreus | |||||
| V. rotundifolia | |||||
| Species | Wind Erosion Rate (g m−2 s−1) | ||||
|---|---|---|---|---|---|
| Vegetation Cover (%) | |||||
| 0% | 20% | 40% | 6% | 80% | |
| I. pes-caprae | 12.56 ± 0.34 a 12.56 ± 0.34 a 12.56 ± 0.34 a | 8.25 ± 0.08 b 4.75 ± 0.09 c 9.14 ± 0.12 a | 5.96 ± 0.09 b 2.48 ± 0.08 c 6.64 ± 0.19 a | 1.58 ± 0.05 b 0.59 ± 0.05 c 2.09 ± 0.07 a | 0.69 ± 0.06 b 0.22 ± 0.03 c 0.99 ± 0.09 a |
| S. littoreus | |||||
| V. rotundifolia | |||||
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lee, J.-T.; Yen, L.-Z.; Chu, M.-Y.; Lin, Y.-S.; Chang, C.-C.; Lin, R.-S.; Chao, K.-H.; Lee, M.-J. Growth Characteristics and Anti-Wind Erosion Ability of Three Tropical Foredune Pioneer Species for Sand Dune Stabilization. Sustainability 2020, 12, 3353. https://0-doi-org.brum.beds.ac.uk/10.3390/su12083353
Lee J-T, Yen L-Z, Chu M-Y, Lin Y-S, Chang C-C, Lin R-S, Chao K-H, Lee M-J. Growth Characteristics and Anti-Wind Erosion Ability of Three Tropical Foredune Pioneer Species for Sand Dune Stabilization. Sustainability. 2020; 12(8):3353. https://0-doi-org.brum.beds.ac.uk/10.3390/su12083353
Chicago/Turabian StyleLee, Jung-Tai, Lin-Zhi Yen, Ming-Yang Chu, Yu-Syuan Lin, Chih-Chia Chang, Ru-Sen Lin, Kung-Hsing Chao, and Ming-Jen Lee. 2020. "Growth Characteristics and Anti-Wind Erosion Ability of Three Tropical Foredune Pioneer Species for Sand Dune Stabilization" Sustainability 12, no. 8: 3353. https://0-doi-org.brum.beds.ac.uk/10.3390/su12083353