Time-Dependent Effects of Bentazon Application on the Key Antioxidant Enzymes of Soybean and Common Ragweed

 ,
,  , ,
, , 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Materials
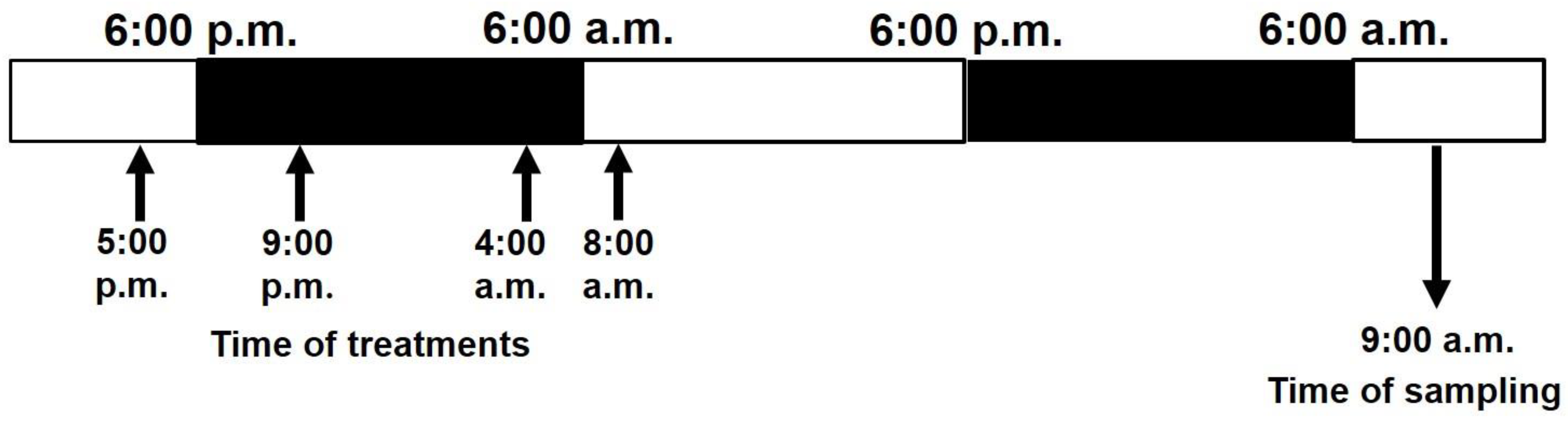
2.2. Treatments
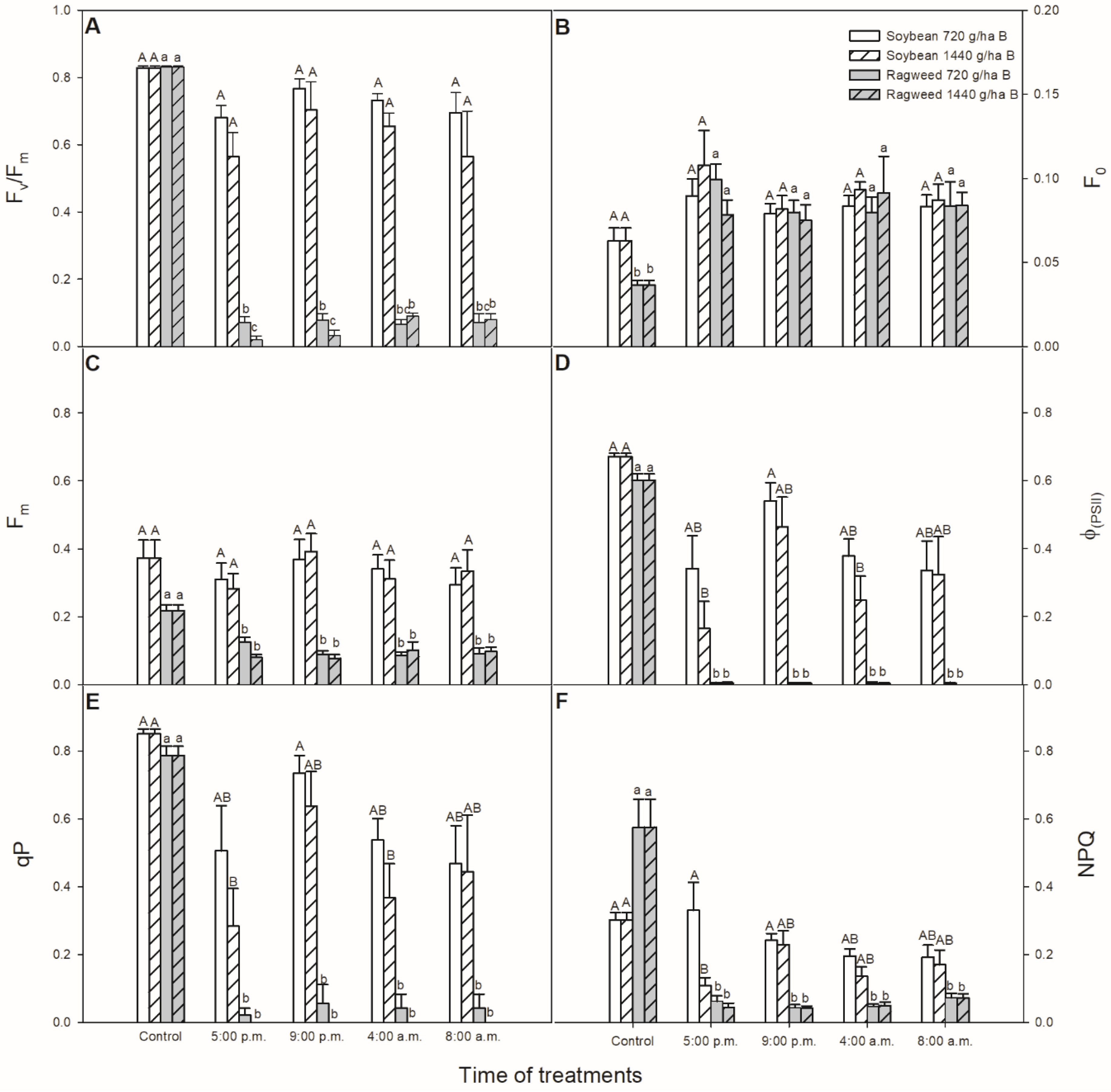
2.3. Measurement of Photosynthetic Activity
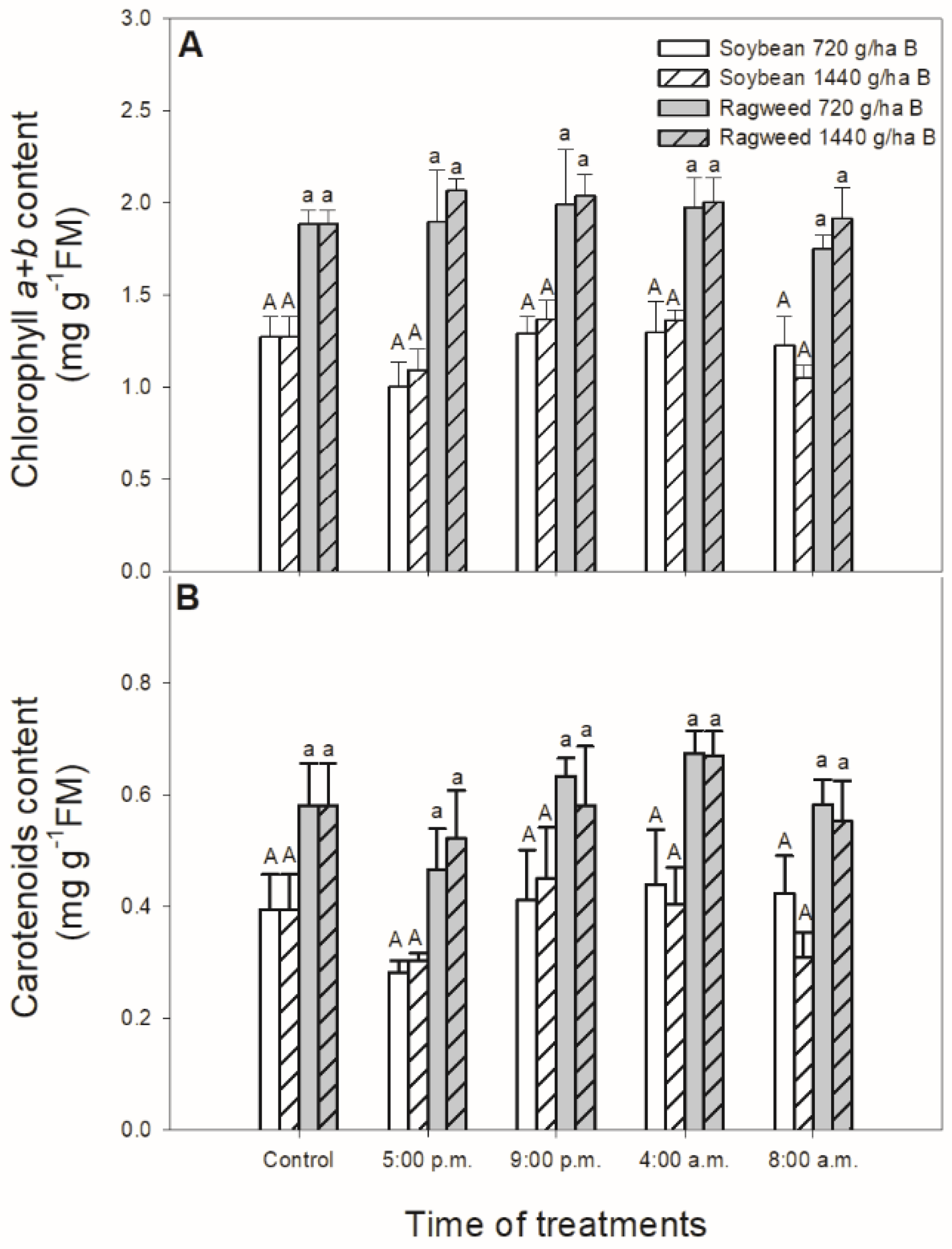
2.4. Determination of Photosynthetic Pigment Content
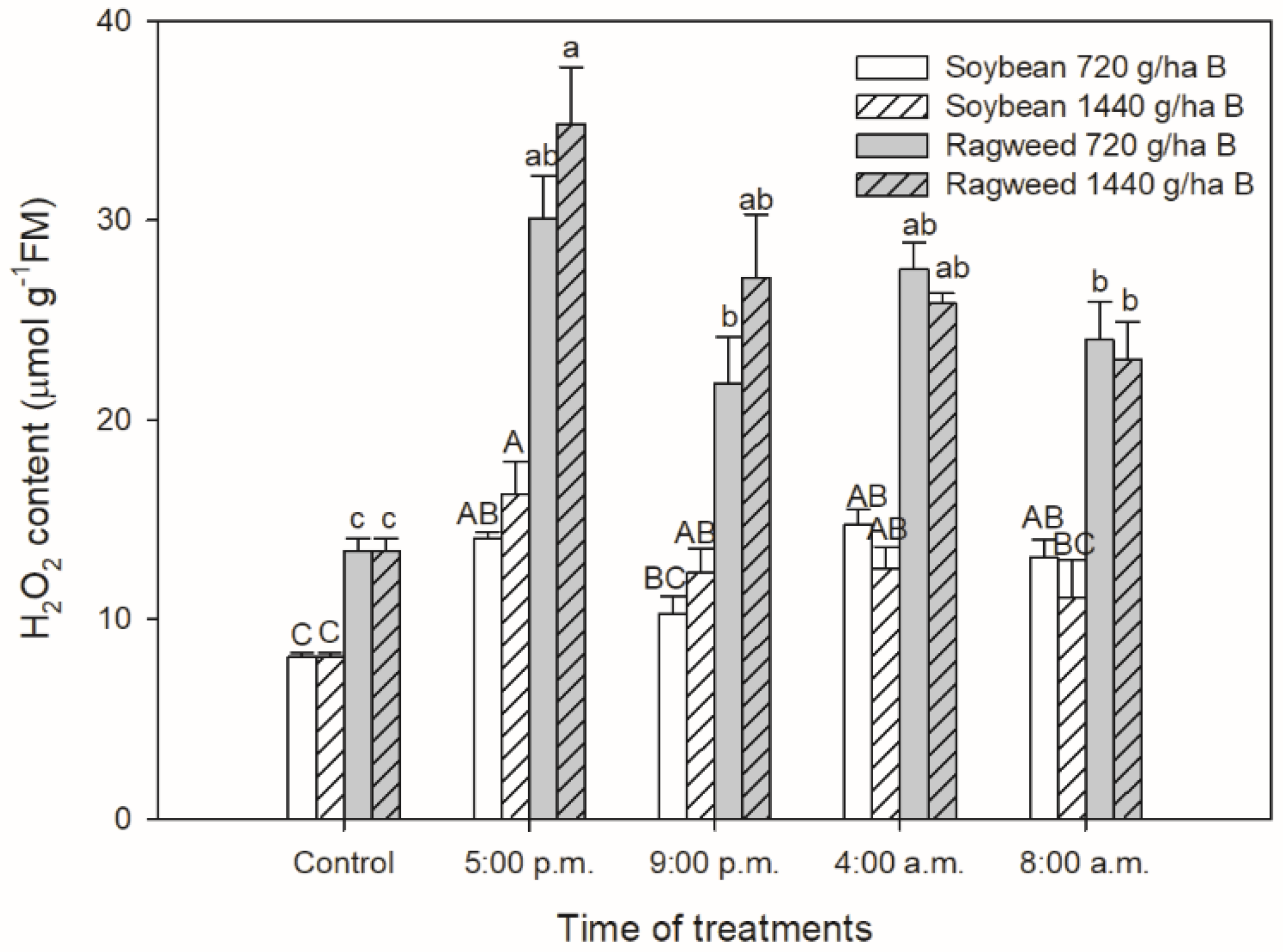
2.5. Determination of H2O2 Content
2.6. Determination of Activities of Key Antioxidant Enzymes
2.7. Measurements of Ascorbate (AsA) and Glutathione (GSH) Levels
2.8. Determination of Glutathione S-Transferase (GST) Activity
2.9. Determination of Malondialdehyde (MDA) Content
2.10. Determination of Electrolyte Leakage (EL) and Leaf Fresh Mass (FM)
2.11. Statistical Analysis
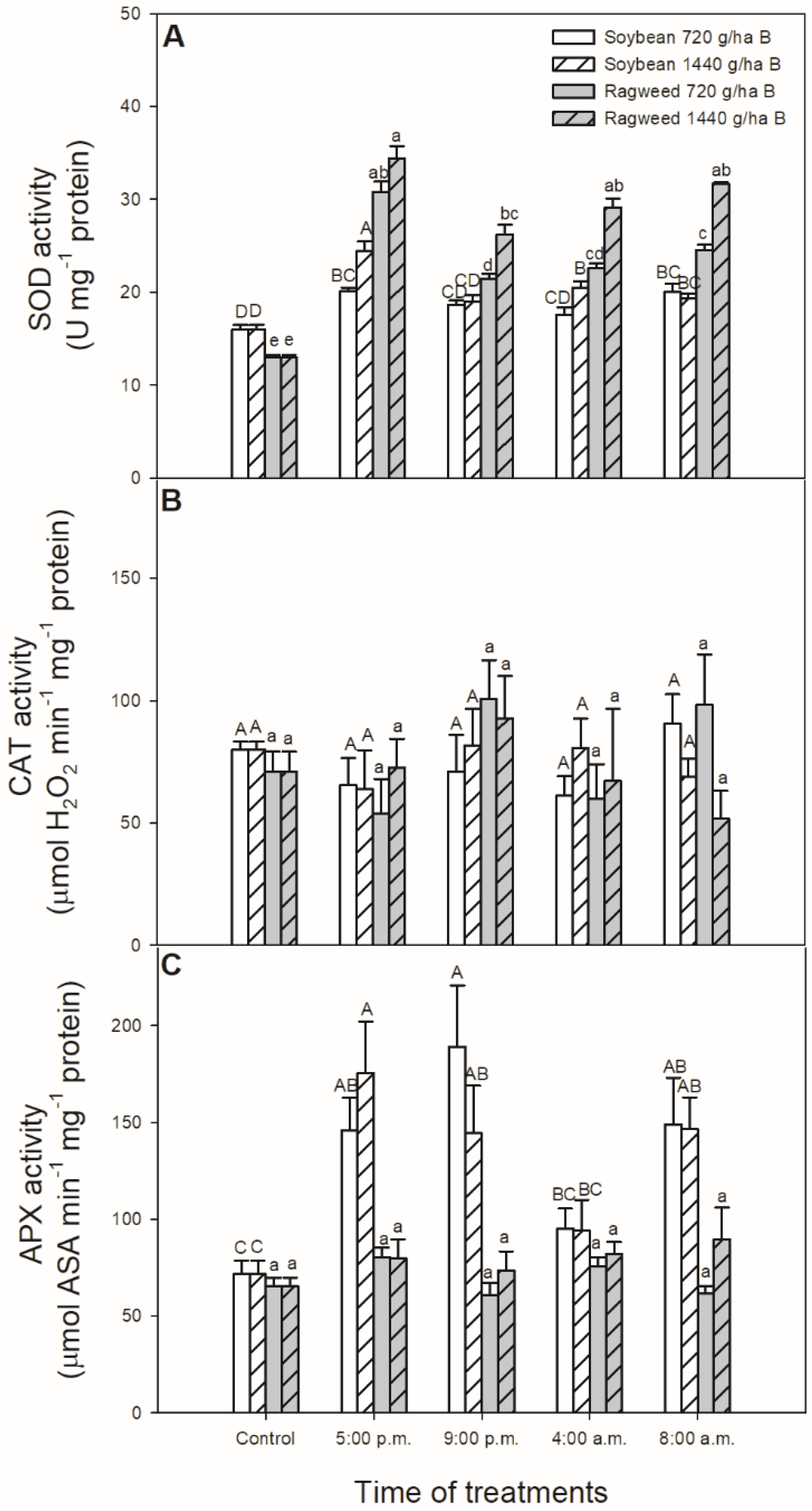
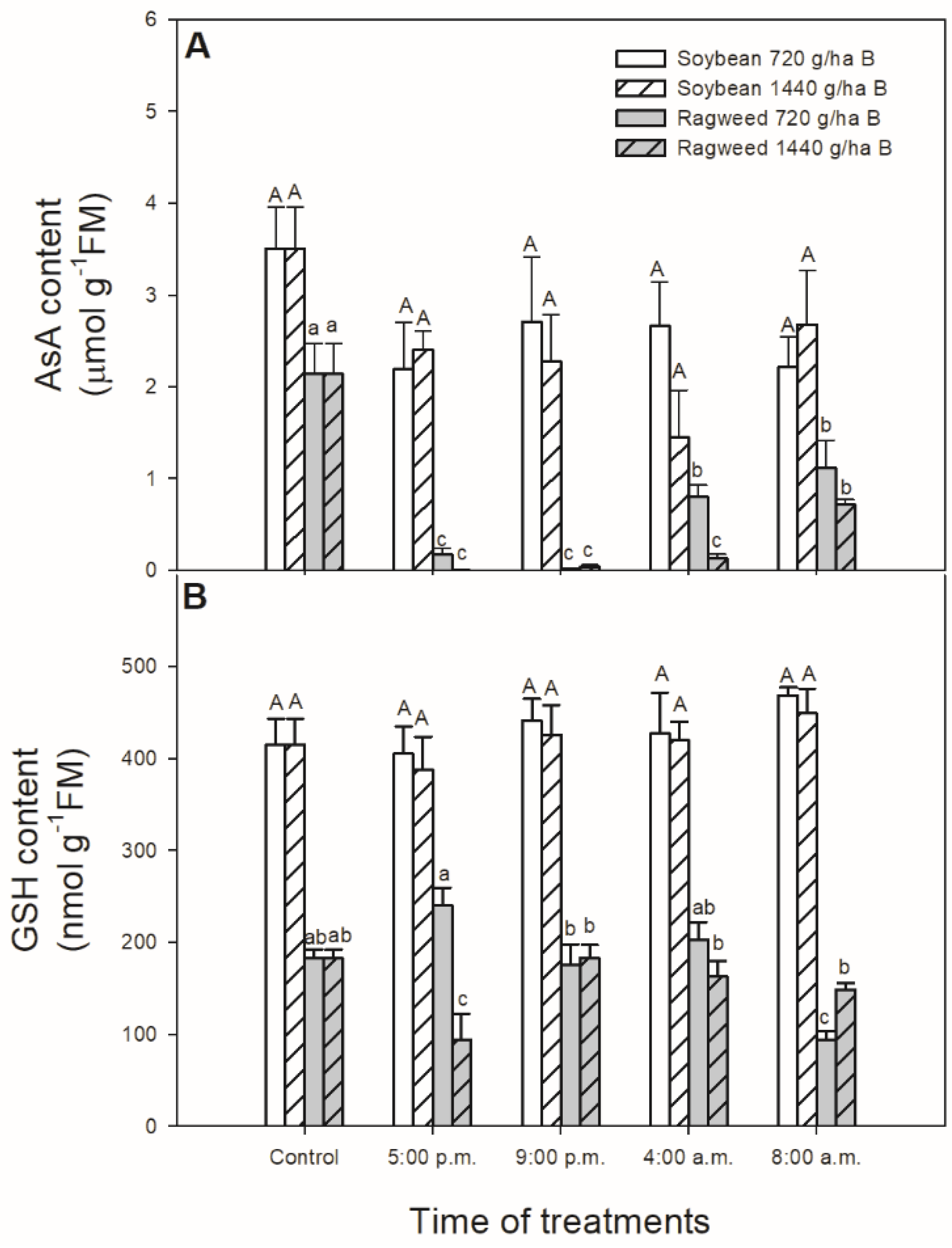
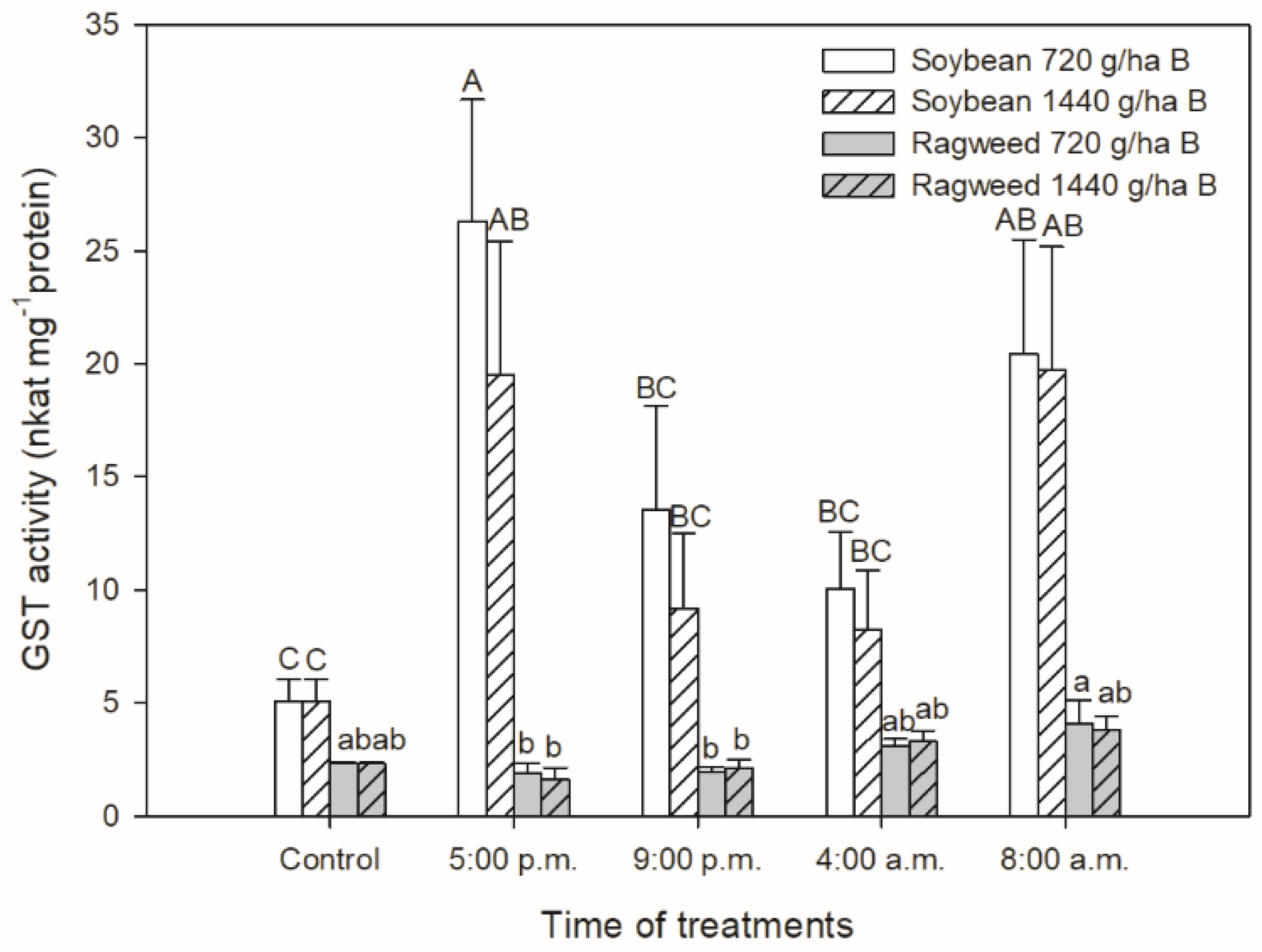
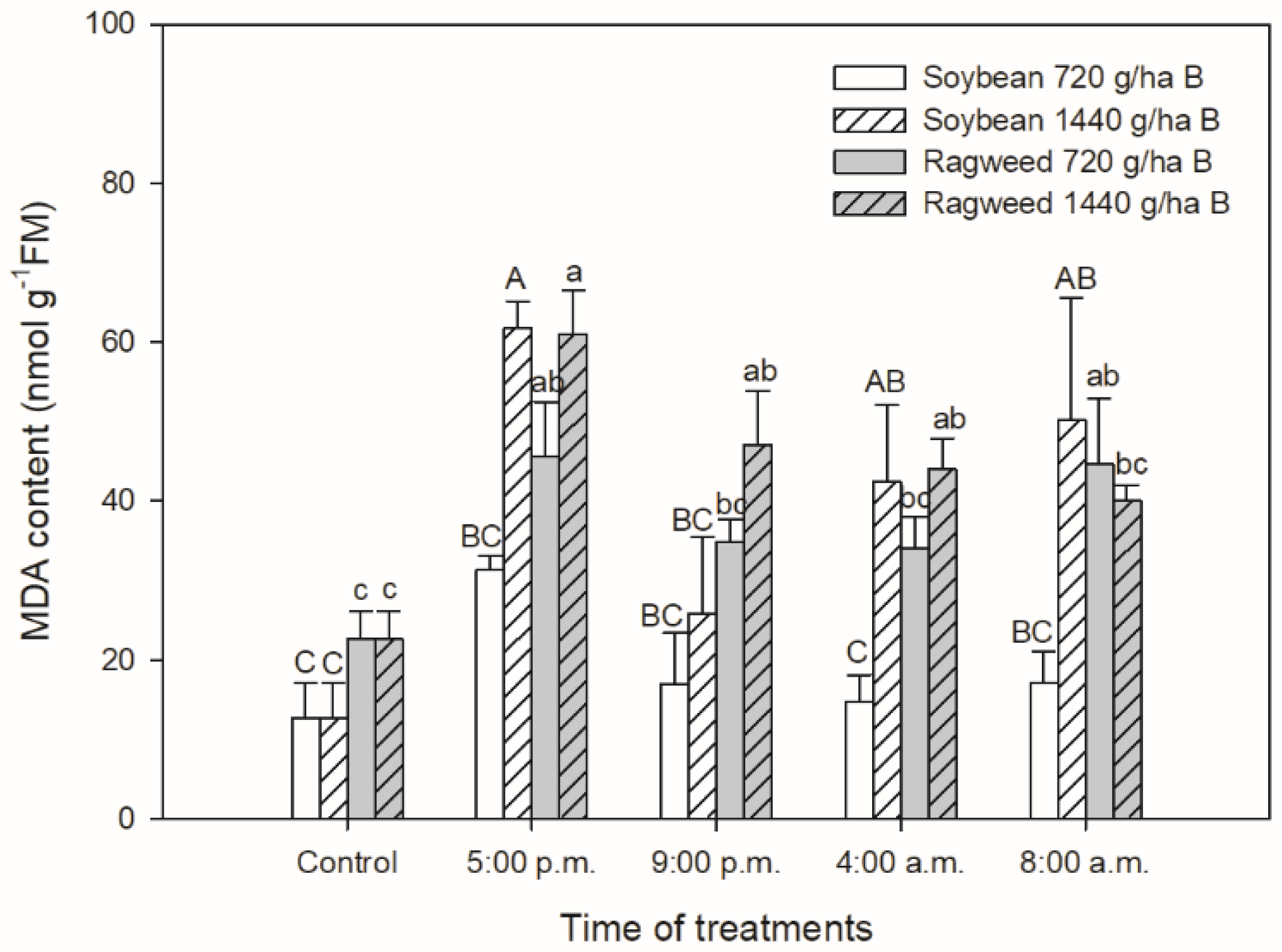
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Hartman, G.L.; West, E.D.; Herman, T.K. Crops that feed the World 2. Soybean—Worldwide production, use, and constraints caused by pathogens and pests. Food Secur. 2011, 3, 5–17. [Google Scholar] [CrossRef]
- Bish, M.D.; Bradley, K.W. Survey of Missouri pesticide applicator practices, knowledge, and perceptions. Weed Technol. 2017, 31, 165–177. [Google Scholar] [CrossRef] [Green Version]
- Soltani, N.; Dille, J.A.; Burke, I.C.; Everman, W.J.; VanGessel, M.J.; Davis, V.M.; Sikkema, P.H. Perspectives on potential soybean yield losses from weeds in North America. Weed Technol. 2017, 31, 148–154. [Google Scholar] [CrossRef] [Green Version]
- Nandula, V.K. Herbicide resistance traits in maize and soybean: Current Status and future outlook. Plants 2019, 8, 337. [Google Scholar] [CrossRef] [Green Version]
- Hart, S.E.; Roskamp, G.K. Soybean (Glycine max) response to thifensulfuron and bentazon combinations. Weed Technol. 1998, 12, 179–184. [Google Scholar] [CrossRef]
- Lycan, D.W.; Hart, S.E. Physiological response of soybean (Glycine max) and two weed species to thifensulfuron and bentazon combinations. Weed Sci. 1999, 47, 143–148. [Google Scholar] [CrossRef]
- Stewart, C.L.; Nurse, R.E.; Hamill, A.S.; Sikkema, P.H. Environment and soil conditions influence pre-and postemergence herbicide efficacy in soybean. Weed Technol. 2010, 24, 234–243. [Google Scholar] [CrossRef]
- Hager, A.; Renner, K. Common ragweed (Ambrosia artemisiifolia) control in soybean (Glycine max) with bentazon as influenced by imazethapyr or thifensulfuron tank-mixes. Weed Technol. 1994, 8, 766–771. [Google Scholar] [CrossRef]
- Ganie, Z.A.; Jhala, A.J. Glyphosate-resistant common ragweed (Ambrosia artemisiifolia) in Nebraska: Confirmation and response to postemergence corn and soybean herbicides. Weed Technol. 2017, 31, 225–237. [Google Scholar] [CrossRef]
- Weaver, S.E. Impact of lamb’s-quarters, common ragweed and green foxtail on yield of corn and soybean in Ontario. Can. J. Plant Sci. 2001, 81, 821–828. [Google Scholar] [CrossRef]
- Pospíšil, P. Production of reactive oxygen species by photosystem II. Biochim. Biophys. Acta 2009, 1787, 1151–1160. [Google Scholar] [CrossRef] [Green Version]
- Galhano, V.; Peixoto, F.; Gomes-Laranjo, J. Bentazon triggers the promotion of oxidative damage in the Portuguese ricefield cyanobacterium Anabaena cylindrica: Response of the antioxidant system. Environ. Toxicol. 2010, 25, 517–526. [Google Scholar] [CrossRef]
- Roach, T.; Krieger-Liszkay, A. Regulation of photosynthetic electron transport and photoinhibition. Curr. Protein Pept. Sci. 2014, 15, 351–362. [Google Scholar] [CrossRef] [Green Version]
- Das, P.K.; Bagchi, S.N. Bentazone and bromoxynil induce H+ and H2O2 accumulation, and inhibit photosynthetic O2 evolution in Synechococcous elongatus PCC7942. Pest. Biochem. Physiol. 2010, 97, 256–261. [Google Scholar] [CrossRef]
- Jervekani, M.T.; Karimmojeni, H.; Razmjo, J.; Tseng, T.M. Common sage (Salvia officinalis L.) tolerance to herbicides. Ind. Crops Prod. 2018, 121, 46–53. [Google Scholar] [CrossRef]
- Radwan, D.E.M.; Mohamed, A.K.; Fayez, K.A.; Abdelrahman, A.M. Oxidative stress caused by Basagran® herbicide is altered by salicylic acid treatments in peanut plants. Heliyon 2019, 5, e01791. [Google Scholar] [CrossRef] [Green Version]
- Gomes, M.P.; Smedbol, E.; Chalifour, A.; Hénault-Ethier, L.; Labrecque, M.; Lepage, L.; Lucotte, M.; Juneau, P. Alteration of plant physiology by glyphosate and its by-product aminomethylphosphonic acid: An overview. J. Exp. Bot. 2014, 65, 4691–4703. [Google Scholar] [CrossRef] [Green Version]
- Foyer, C.H.; Noctor, G. Redox regulation in photosynthetic organisms: Signaling, acclimation, and practical implications. Antioxid. Redox Signal. 2009, 11, 861–905. [Google Scholar] [CrossRef]
- Van Aken, O.; Van Breusegem, F. Licensed to kill: Mitochondria, chloroplasts, and cell death. Trends Plant Sci. 2015, 20, 754–766. [Google Scholar] [CrossRef]
- Czarnocka, W.; Karpiński, S. Friend or foe? Reactive oxygen species production, scavenging and signaling in plant response to environmental stresses. Free Radic. Biol. Med. 2018, 122, 4–20. [Google Scholar] [CrossRef]
- Noctor, G.; Reichheld, J.P.; Foyer, C.H. ROS-related redox regulation and signaling in plants. Semin. Cell Dev. Biol. 2018, 80, 3–12. [Google Scholar] [CrossRef] [Green Version]
- Knörzer, O.C.; Burner, J.; Boger, P. Alterations in the antioxidative system of suspension-cultured soybean cells (Glycine max) induced by oxidative stress. Phys. Plant. 1996, 97, 388–396. [Google Scholar] [CrossRef]
- Malenčić, D.; Miladinović, J.; Popović, M. Effects of linuron and dimethenamid on antioxidant systems in weeds associated with soybean. Open Life Sci. 2008, 3, 155–160. [Google Scholar] [CrossRef]
- Gomes, M.P.; Bicalho, E.M.; Smedbol, E.; Cruz, F.V.D.S.; Lucotte, M.; Garcia, Q.S. Glyphosate can decrease germination of glyphosate-resistant soybeans. J. Agric. Food Chem. 2017, 65, 2279–2286. [Google Scholar] [CrossRef]
- Hamim, H.; Violita, V.; Triadiati, T.; Miftahudin, M. Oxidative stress and photosynthesis reduction of cultivated (Glycine max L.) and wild soybean (G. tomentella L.) exposed to drought and paraquat. Asian J. Plant Sci. 2017, 16, 65–77. [Google Scholar] [CrossRef] [Green Version]
- Poór, P.; Ördög, A.; Czékus, Z.; Borbély, P.; Takács, Z.; Kovács, J.; Tari, I. Regulation of the key antioxidant enzymes by developmental processes and environmental stresses in the dark. Biol. Plant. 2018, 62, 201–210. [Google Scholar] [CrossRef]
- Kerdnaimongkol, K.; Bhatia, A.; Joly, R.J.; Woodson, W.R. Oxidative stress and diurnal variation in chilling sensitivity of tomato seedlings. J. Am. Soc. Hortic. Sci. 1997, 122, 485–490. [Google Scholar] [CrossRef] [Green Version]
- Dutilleul, C.; Garmier, M.; Noctor, G.; Mathieu, C.; Chétrit, P.; Foyer, C.H.; De Paepe, R. Leaf mitochondria modulate whole cell redox homeostasis, set antioxidant capacity, and determine stress resistance through altered signaling and diurnal regulation. Plant Cell 2003, 15, 1212–1226. [Google Scholar] [CrossRef] [Green Version]
- Pignocchi, C.; Fletcher, J.M.; Wilkinson, J.E.; Barnes, J.D.; Foyer, C.H. The function of ascorbate oxidase in tobacco. Plant Physiol. 2003, 132, 1631–1641. [Google Scholar] [CrossRef] [Green Version]
- Mhamdi, A.; Noctor, G.; Baker, A. Plant catalases: Peroxisomal redox guardians. Arch. Biochem. Biophys. 2012, 525, 181–194. [Google Scholar] [CrossRef]
- Poór, P.; Takács, Z.; Bela, K.; Czékus, Z.; Szalai, G.; Tari, I. Prolonged dark period modulates the oxidative burst and enzymatic antioxidant systems in the leaves of salicylic acid-treated tomato. J. Plant Physiol. 2017, 213, 216–226. [Google Scholar] [CrossRef]
- Nianiou-Obeidat, I.; Madesis, P.; Kissoudis, C.; Voulgari, G.; Chronopoulou, E.; Tsaftaris, A.; Labrou, N.E. Plant glutathione transferase-mediated stress tolerance: Functions and biotechnological applications. Plant Cell Rep. 2017, 36, 791–805. [Google Scholar] [CrossRef]
- Zechmann, B. Diurnal changes of subcellular glutathione content in Arabidopsis thaliana. Biol. Plant. 2017, 61, 791–796. [Google Scholar] [CrossRef]
- Gallé, Á.; Czékus, Z.; Bela, K.; Horváth, E.; Csiszár, J.; Poór, P. Diurnal changes in tomato glutathione transferase activity and expression. Acta Biol. Hung. 2018, 69, 505–509. [Google Scholar] [CrossRef]
- Gallé, Á.; Czékus, Z.; Bela, K.; Horváth, E.; Ördög, A.; Csiszár, J.; Poór, P. Plant glutathione transferases and light. Front. Plant Sci. 2019, 9, 1944. [Google Scholar] [CrossRef] [Green Version]
- Guóth, A.; Benyó, D.; Csiszár, J.; Gallé, Á.; Horváth, F.; Cseuz, L.; Erdei, L.; Tari, I. Relationship between osmotic stress-induced abscisic acid accumulation, biomass production and plant growth in drought-tolerant and-sensitive wheat cultivars. Acta Physiol. Plant 2010, 32, 719–727. [Google Scholar]
- Czékus, Z.; Poór, P.; Tari, I.; Ördög, A. Effects of light and daytime on the regulation of chitosan-induced stomatal responses and defence in tomato plants. Plants 2020, 9, 59. [Google Scholar] [CrossRef] [Green Version]
- Genty, B.; Briantais, J.M.; Baker, N.R. The relationship between the quantum yield of photosynthetic electron transport and quenching of chlorophyll fluorescence. Biochim. Biophys. Acta 1989, 990, 87–92. [Google Scholar] [CrossRef]
- Kramer, D.M.; Johnson, G.; Kiirats, O.; Edwards, G.E. New fluorescence parameters for the determination of QA redox state and excitation energy fluxes. Photosyn. Res. 2004, 79, 209. [Google Scholar] [CrossRef]
- Lichtenthaler, H.K.; Wellburn, A.R. Determinations of total carotenoids and chlorophylls a and b of leaf extracts in different solvents. Biochem. Soc. Trans. 1983, 11, 591–592. [Google Scholar] [CrossRef] [Green Version]
- Velikova, V.; Yordanov, I.; Edreva, A. Oxidative stress and some antioxidant systems in acid rain-treated bean plants: Protective role of exogenous polyamines. Plant Sci. 2000, 151, 59–66. [Google Scholar] [CrossRef]
- Tari, I.; Csiszár, J.; Horváth, E.; Poór, P.; Takács, Z.; Szepesi, Á. The alleviation of the adverse effects of salt stress in the tomato plant by salicylic acid shows a time-and organ-specific antioxidant response. Acta Biol. Cracov. Bot. 2015, 57, 21–30. [Google Scholar] [CrossRef]
- Bradford, M.M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- Szepesi, Á.; Csiszár, J.; Gallé, Á.; Gémes, K.; Poór, P.; Tari, I. Effects of long-term salicylic acid pre-treatment on tomato (Lycopersicon esculentum Mill. L.) salt stress tolerance: Changes in glutathione S-transferase activities and anthocyanin contents. Acta Agron. Hung. 2008, 56, 129–138. [Google Scholar] [CrossRef]
- Ederli, L.; Pasqualini, S.; Batini, P.; Antonielli, M. Photoinhibition and oxidative stress: Effects on xanthophyll cycle, scavenger enzymes and abscisic acid content in tobacco plants. J. Plant Physiol. 1997, 151, 422–428. [Google Scholar] [CrossRef]
- Sun, J.; Li, L.; Liu, M.; Wang, M.; Ding, M.; Deng, S.; Chen, S. Hydrogen peroxide and nitric oxide mediate K+/Na+ homeostasis and antioxidant defense in NaCl-stressed callus cells of two contrasting poplars. Plant Cell Tissue Organ Cult. 2010, 103, 205–215. [Google Scholar] [CrossRef]
- Bourguet, D.; Guillemaud, T. The hidden and external costs of pesticide use. In Sustainable Agriculture Reviews, 1st ed.; Lichtfouse, E., Ed.; Springer: Cham, Switzerland, 2016; Volume 19, pp. 35–120. [Google Scholar]
- Sharma, A.; Kumar, V.; Shahzad, B.; Tanveer, M.; Sidhu, G.P.S.; Handa, N.; Kohli, S.K.; Yadav, P.; Bali, A.S.; Parihar, R.D.; et al. Worldwide pesticide usage and its impacts on ecosystem. SN Appl. Sci. 2019, 1, 1446. [Google Scholar] [CrossRef] [Green Version]
- Kristoffersen, P.; Rask, A.M.; Grundy, A.C.; Franzen, I.; Kempenaar, C.; Raisio, J.; Schroeder, H.; Spijker, J.; Verschwele, A.; Zarina, L. A review of pesticide policies and regulations for urban amenity areas in seven European countries. Weed Res. 2008, 48, 201–214. [Google Scholar] [CrossRef]
- Gill, S.S.; Tuteja, N. Reactive oxygen species and antioxidant machinery in abiotic stress tolerance in crop plants. Plant Physiol. Biochem. 2010, 48, 909–930. [Google Scholar] [CrossRef]
- Kocsy, G.; Tari, I.; Vanková, R.; Zechmann, B.; Gulyás, Z.; Poór, P.; Galiba, G. Redox control of plant growth and development. Plant Sci. 2013, 211, 77–91. [Google Scholar] [CrossRef]
- Lati, R.N.; Mou, B.; Rachuy, J.S.; Fennimore, S.A. Evaluation of cycloate followed by evening two-leaf–stage phenmedipham application in fresh market spinach. Weed Technol. 2016, 30, 464–471. [Google Scholar] [CrossRef]
- Frenkel, E.; Matzrafi, M.; Rubin, B.; Peleg, Z. Effects of environmental conditions on the fitness penalty in herbicide resistant Brachypodium hybridum. Front. Plant Sci. 2017, 8, 94. [Google Scholar] [CrossRef] [Green Version]
- Lei, Y.B.; Zheng, Y.L.; Dai, K.J.; Duan, B.L.; Cai, Z.Q. Different responses of photosystem I and photosystem II in three tropical oilseed crops exposed to chilling stress and subsequent recovery. Trees 2014, 28, 923–933. [Google Scholar] [CrossRef]
- Demmig-Adams, B.; Cohu, C.M.; Muller, O.; Adams, W.W. Modulation of photosynthetic energy conversion efficiency in nature: From seconds to seasons. Photosynth. Res. 2012, 113, 75–88. [Google Scholar] [CrossRef]
- Kummerová, M.; Váňová, L.; Krulová, J.; Zezulka, Š. The use of physiological characteristics for comparison of organic compounds phytotoxicity. Chemosphere 2008, 71, 2050–2059. [Google Scholar] [CrossRef]
- Sterling, T.M.; Balke, N.E.; Silverman, D.S. Uptake and accumulation of the herbicide bentazon by cultured plant cells. Plant Phys. 1990, 92, 1121–1127. [Google Scholar] [CrossRef] [Green Version]
- Kato, S.; Yokota, Y.; Suzuki, R.; Fujisawa, Y.; Sayama, T.; Kaga, A.; Anai, T.; Komatsu, K.; Oki, N.; Kikuchi, A.; et al. Identification of a cytochrome P450 hydroxylase, CYP81E22, as a causative gene for the high sensitivity of soybean to herbicide bentazon. Theor. Appl. Genet. 2020, 1–11. [Google Scholar] [CrossRef]
- Murchie, E.H.; Lawson, T. Chlorophyll fluorescence analysis: A guide to good practice and understanding some new applications. J. Exp. Bot. 2013, 64, 3983–3998. [Google Scholar] [CrossRef] [Green Version]
- Liu, Y.F.; Qi, M.F.; Li, T.L. Photosynthesis, photoinhibition, and antioxidant system in tomato leaves stressed by low night temperature and their subsequent recovery. Plant Sci. 2012, 196, 8–17. [Google Scholar] [CrossRef]
- Poór, P.; Borbély, P.G.; Bódi, N.; Bagyánszki, M.; Tari, I. Effects of salicylic acid on photosynthetic activity and chloroplast morphology under light and prolonged darkness. Photosynthetica 2019, 57, 367–376. [Google Scholar] [CrossRef] [Green Version]
- Zhang, G.; Liu, Y.; Ni, Y.; Meng, Z.; Lu, T.; Li, T. Exogenous calcium alleviates low night temperature stress on the photosynthetic apparatus of tomato leaves. PLoS ONE 2014, 9, e97010. [Google Scholar] [CrossRef]
- Demmig-Adams, B.; Adams, W.W., III. The role of xanthophyll cycle carotenoids in the protection of photosynthesis. Trends Plant Sci. 1996, 1, 21–26. [Google Scholar] [CrossRef]
- Cheng, D.D.; Liu, M.J.; Sun, X.B.; Zhao, M.; Chow, W.S.; Sun, G.Y.; Zhang, Z.S.; Hu, Y.B. Light suppresses bacterial population through the accumulation of hydrogen peroxide in tobacco leaves infected with Pseudomonas syringae pv. tabaci. Front. Plant Sci. 2016, 7, 512. [Google Scholar] [CrossRef]
- Karapetyan, S.; Dong, X. Redox and the circadian clock in plant immunity: A balancing act. Free Radic. Biol. Med. 2018, 119, 56–61. [Google Scholar] [CrossRef]
- Alscher, R.G.; Erturk, N.; Heath, L.S. Role of superoxide dismutases (SODs) in controlling oxidative stress in plants. J. Exp. Bot. 2002, 53, 1331–1341. [Google Scholar] [CrossRef]
- Cole, D.J. Detoxification and activation of agrochemicals in plants. Pestic. Sci. 1994, 42, 209–222. [Google Scholar] [CrossRef]
- Cummins, I.; Cole, D.J.; Edwards, R. A role for glutathione transferases functioning as glutathione peroxidases in resistance to multiple herbicides in black-grass. Plant J. 1999, 18, 285–292. [Google Scholar] [CrossRef]
- Batish, D.R.; Singh, H.P.; Setia, N.; Kaur, S.; Kohli, R.K. 2-Benzoxazolinone (BOA) induced oxidative stress, lipid peroxidation and changes in some antioxidant enzyme activities in mung bean (Phaseolus aureus). Plant Physiol. Biochem. 2006, 44, 819–827. [Google Scholar] [CrossRef] [PubMed]
- Bajpai, J.; Srivastava, A. In vitro response of black gram genotypes to herbicide stress and elevation of antioxidative defence system. Acta Physiol. Plant 2015, 37, 182. [Google Scholar] [CrossRef]
- Jiang, Z.; Ma, B.; Erinle, K.O.; Cao, B.; Liu, X.; Ye, S.; Zhang, Y. Enzymatic antioxidant defense in resistant plant: Pennisetum americanum (L.) K. Schum during long-term atrazine exposure. Pest. Biochem. Physiol. 2016, 133, 59–66. [Google Scholar] [CrossRef] [PubMed]









© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Czékus, Z.; Farkas, M.; Bakacsy, L.; Ördög, A.; Gallé, Á.; Poór, P. Time-Dependent Effects of Bentazon Application on the Key Antioxidant Enzymes of Soybean and Common Ragweed. Sustainability 2020, 12, 3872. https://0-doi-org.brum.beds.ac.uk/10.3390/su12093872
Czékus Z, Farkas M, Bakacsy L, Ördög A, Gallé Á, Poór P. Time-Dependent Effects of Bentazon Application on the Key Antioxidant Enzymes of Soybean and Common Ragweed. Sustainability. 2020; 12(9):3872. https://0-doi-org.brum.beds.ac.uk/10.3390/su12093872
Chicago/Turabian StyleCzékus, Zalán, Máté Farkas, László Bakacsy, Attila Ördög, Ágnes Gallé, and Péter Poór. 2020. "Time-Dependent Effects of Bentazon Application on the Key Antioxidant Enzymes of Soybean and Common Ragweed" Sustainability 12, no. 9: 3872. https://0-doi-org.brum.beds.ac.uk/10.3390/su12093872