
Soil Nutrient Supply in Cultivated Bush Bean–Potato Intercropping Grown in Subarctic Soil Managed with Agroforestry

Abstract
:1. Introduction
2. Materials and Methods
2.1. Location
2.2. Site Preparations
2.3. Ion Exchange Resin Probes
2.4. Statistical Analysis
3. Results
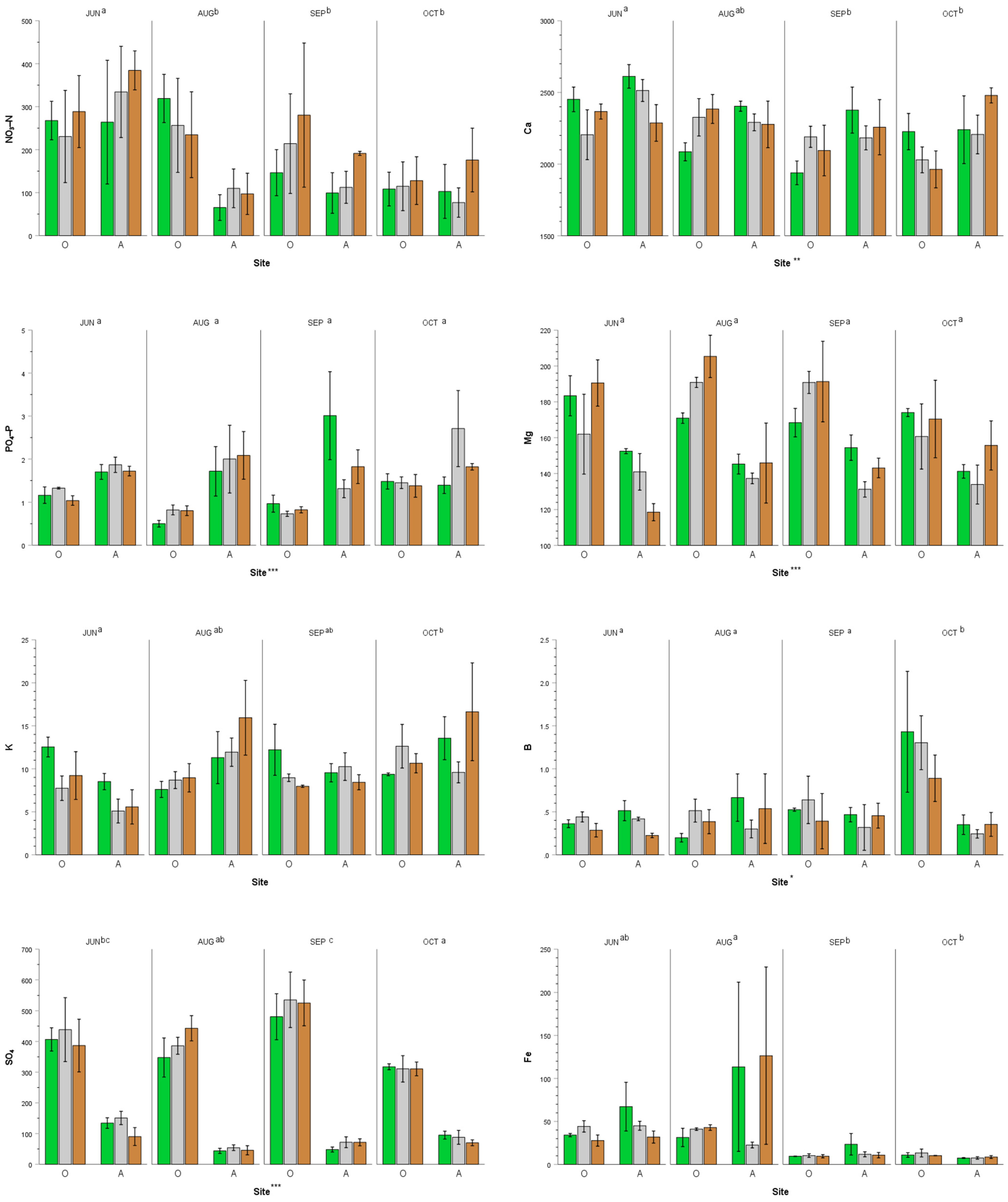
3.1. Spatiotemporal Soil Nutrient Supply Rate in Agroforestry and Open Sites
3.2. The Relationship between Soil Nutrient Supply Rate and Leaf Tissue Chemistry
3.3. The Relationship between Soil Nutrient Supply Rate and Yield
4. Discussion
4.1. Agroforestry Sites Had Greater Supply Rates of Phosphorus, but It Is Still Inadequate for Maximum Crop Yield
4.2. Spatiotemporal Dynamics of Macronutrient Supply Rates in Subarctic Cultivated Soil
4.3. Spatiotemporal Dynamics of Micronutrient Supply Rates in Subarctic Cultivated Soil
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Altdorff, D.; Borchard, N.; Young, E.H.; Galagedara, L.; Sorvali, J.; Quideau, S.; Unc, A. Agriculture in boreal and Arctic regions requires an integrated global approach for research and policy. Agron. Sustain. Dev. 2021, 41, 23. [Google Scholar] [CrossRef]
- Hannah, L.; Roehrdanz, P.R.; Krishna Bahadur, K.C.; Fraser, E.D.G.; Donatti, C.I.; Saenz, L.; Wright, T.M.; Hijmans, R.J.; Mulligan, M.; Berg, A.; et al. The environmental consequences of climate-driven agricultural frontiers. PLoS ONE 2020, 15, e0228305. [Google Scholar] [CrossRef]
- Barbeau, C.D.; Wilton, M.J.; Oelbermann, M.; Karagatzides, J.D.; Tsuji, L.J.S. Local food production in a subarctic Indigenous community: The use of willow (Salix spp.) windbreaks to increase the yield of intercropped potatoes (Solanum tuberosum) and bush beans (Phaseolus vulgaris). Int. J. Agric. Sust. 2017, 16, 29–39. [Google Scholar] [CrossRef]
- Chen, A.; Natcher, D. Greening Canada’s Arctic food system: Local food procurement strategies for combating food insecurity. Can. Food Stud. 2019, 6, 140–154. [Google Scholar] [CrossRef] [Green Version]
- Kedir, A.J.; Zhang, M.; Unc, A. Understanding soil fertility status in Newfoundland from standard farm soil tests. Can. J. Soil Sci. 2021, 1–15. [Google Scholar] [CrossRef]
- Smith, J.; Pearce, B.D.; Wolfe, M.S. Reconciling productivity with protection of the environment: Is temperate agroforestry the answer? Renew. Agric. Food Syst. 2013, 28, 80–92. [Google Scholar] [CrossRef]
- Bhardwaj, D.R.; Navale, M.R.; Sharma, S. Agroforestry practices in temperate regions of the world. In Agroforestry; Dagar, J., Tewari, V., Eds.; Springer: Singapore, 2017; pp. 163–187. [Google Scholar] [CrossRef]
- Hynynen, J.; Niemistö, P.; Viherä-Aarnio, A.; Brunner, A.; Hein, S.; Velling, P. Silviculture of birch (Betula pendula Roth and Betula pubescens Ehrh.) in northern Europe. Forestry 2010, 83, 103–119. [Google Scholar] [CrossRef]
- Dubois, H.; Verkasalo, E.; Claessens, H. Potential of birch (Betula pendula Roth and B. pubescens Ehrh.) for forestry and forest-based industry sector within the changing climatic and socio-economic context of Western Europe. Forests 2020, 11, 336. [Google Scholar] [CrossRef] [Green Version]
- Pohjanmies, T.; Jašková, A.; Hotanen, J.P.; Manninen, O.; Salemaa, M.; Tolvanen, A.; Merilä, P. Abundance and diversity of edible wild plants in managed boreal forests. For. Ecol. Manag. 2021, 491, 119151. [Google Scholar] [CrossRef]
- Spiegelaar, N.F.; Tsuji, L.J.S.; Oelbermann, M. The potential use of agroforestry community gardens as a sustainable import-substitution strategy for enhancing food security in subarctic Ontario, Canada. Sustainability 2013, 5, 4057–4075. [Google Scholar] [CrossRef] [Green Version]
- Skinner, K.; Hanning, R.M.; Desjardins, E.; Tsuji, L.J. Giving voice to food insecurity in a remote indigenous community in subarctic Ontario, Canada: Traditional ways, ways to cope, ways forward. BMC Public Health 2013, 13, 427. [Google Scholar] [CrossRef] [Green Version]
- Barbeau, C.D.; Oelbermann, M.; Karagatzides, J.D.; Tsuji, L.J.S. Sustainable agriculture and climate change: Producing potatoes (Solanum tuberosum L.) and bush beans (Phaseolus vulgaris L.) for improved food security and resilience in a Canadian subarctic First Nations community. Sustainability 2015, 7, 5664–5681. [Google Scholar] [CrossRef] [Green Version]
- Birch, P.R.; Bryan, G.; Fenton, B.; Gilroy, E.M.; Hein, I.; Jones, J.T.; Prashar, A.; Taylor, M.A.; Torrance, L.; Toth, I.K. Crops that feed the world 8: Potato: Are the trends of increased global production sustainable? Food Secur. 2012, 4, 477–508. Available online: https://0-link-springer-com.brum.beds.ac.uk/article/10.1007/s12571-012-0220-1 (accessed on 21 May 2021). [CrossRef]
- Spiegelaar, N.F.; Tsuji, L.J. Impact of Euro-Canadian agrarian practices: In search of sustainable import-substitution strategies to enhance food security in subarctic Ontario, Canada. Rural Remote Health 2013, 13, 2211. [Google Scholar] [CrossRef] [PubMed]
- Rodriguez, C.; Carlsson, G.; Englund, J.E.; Flöhr, A.; Pelzer, E.; Jeuffroy, M.H.; Makowski, D.; Jensen, E.S. Grain legume-cereal intercropping enhances the use of soil-derived and biologically fixed nitrogen in temperate agroecosystems. A meta-analysis. Eur. J. Agron. 2020, 118, 126077. [Google Scholar] [CrossRef]
- Gitari, H.I.; Gachene, C.K.; Karanja, N.N.; Kamau, S.; Nyawade, S.; Sharma, K.; Schulte-Geldermann, E. Optimizing yield and economic returns of rain-fed potato (Solanum tuberosum L.) through water conservation under potato-legume intercropping systems. Agric. Water Manag. 2018, 208, 59–66. [Google Scholar] [CrossRef]
- Sharaiha, R.K.; Hadidi, N.A. Micro-environmental effects on potato and bean yields grown under intercropping system. Agron. Ser. 2008, 51, 209–219. Available online: http://www.uaiasi.ro/revagrois/PDF/2008_1_209.pdf (accessed on 21 May 2021).
- Techen, A.K.; Helming, K.; Brüggemann, N.; Veldkamp, E.; Reinhold-Hurek, B.; Lorenz, M.; Vogel, H.J. Soil research challenges in response to emerging agricultural soil management practices. Adv. Agron. 2020, 161, 179–240. [Google Scholar] [CrossRef]
- Wilton, M.J.; Karagatzides, J.D.; Tsuji, L.J. Nutrient concentrations of bush bean (Phaseolus vulgaris L.) and potato (Solanum tuberosum L.) cultivated in subarctic soils managed with intercropping and willow (Salix spp.) agroforestry. Sustainability 2017, 9, 2294. [Google Scholar] [CrossRef] [Green Version]
- Environment Canada. Available online: https://climate.weather.gc.ca/climate_normals/ (accessed on 26 April 2021).
- Kenney, W.A. A method for estimating windbreak porosity using digitized photographic silhouettes. Agric. For. Meteorol. 1987, 39, 91–94. [Google Scholar] [CrossRef]
- Western Ag Technologies. Available online: https://www.westernag.ca/ (accessed on 30 April 2021).
- Mohamed, M.A.; Stamatakis, A.; Keramidas, V. Anionic resin–extractable phosphorus as an index of phosphorus availability in calcareous soils of Crete amended or not amended with pig manure compost. Commun. Soil Sci. Plant Anal. 2013, 44, 50–61. [Google Scholar] [CrossRef]
- SPSS 26; SPSS Inc.: Chicago, IL, USA, 2017.
- Van Meeteren, M.J.M.; Tietema, A.; Westerveld, J.W. Regulation of microbial carbon, nitrogen, and phosphorus transformations by temperature and moisture during decomposition of Calluna vulgaris litter. Biol. Fert. Soils 2007, 44, 103–112. [Google Scholar] [CrossRef] [Green Version]
- Rydin, H.; Jeglum, J.K.; Hooijer, A. Nutrients, light and temperature. In The Biology of Peatlands, 2nd ed.; Rydin, H., Jeglum, J.K., Eds.; Oxford University Press: New York, NY, USA, 2013; pp. 176–198. [Google Scholar] [CrossRef]
- Sun, D.; Bi, Q.; Li, K.; Dai, P.; Yu, Y.; Zhou, W.; Lv, T.; Liu, X.; Zhu, J.; Zhang, Q.; et al. Significance of temperature and water availability for soil phosphorus transformation and microbial community composition as affected by fertilizer sources. Biol. Fertil. Soils 2018, 54, 229–241. [Google Scholar] [CrossRef]
- Foereid, B.; Bro, R.; Mogensen, V.O.; Porter, J.R. Effects of windbreak strips of willow coppice—Modelling and field experiment on barley in Denmark. Agric. Ecosyst. Environ. 2002, 93, 25–32. [Google Scholar] [CrossRef]
- Brandle, J.R.; Hodges, L.; Zhou, X.H. Windbreaks in North American agricultural systems. Agrofor. Syst. 2004, 61, 65–78. [Google Scholar]
- Mungai, N.W.; Motavalli, P.P.; Kremer, R.J.; Nelson, K.A. Spatial variation of soil enzyme activities and microbial functional diversity in temperate alley cropping systems. Biol. Fertil. Soils 2005, 42, 129–136. [Google Scholar] [CrossRef]
- Tamang, B.; Andreu, M.G.; Rockwood, D.L. Microclimate patterns on the leeside of single-row tree windbreaks during different weather conditions in Florida farms: Implications for improved crop production. Agrofor. Syst. 2010, 79, 111–122. [Google Scholar] [CrossRef]
- Rinnan, R.; Michelsen, A.; Bååth, E.; Jonasson, S. Mineralization and carbon turnover in subarctic heath soil as affected by warming and additional litter. Soil Biol. Biochem. 2007, 39, 3014–3023. [Google Scholar] [CrossRef]
- Ruess, L.; Michelsen, A.; Schmidt, I.K.; Jonasson, S. Simulated climate change affecting microorganisms, nematode density and biodiversity in subarctic soils. Plant Soil 1999, 212, 63–73. [Google Scholar] [CrossRef]
- Manimel Wadu, M.C.; Ma, F.; Chang, S.X. Phosphorus availabilities differ between cropland and forestland in shelterbelt systems. Forests 2019, 10, 1001. [Google Scholar] [CrossRef] [Green Version]
- Rao, K.P.C.; Verchot, L.V.; Laarman, J. Adaptation to climate change through sustainable management and development of agroforestry systems. J. SAT Agric. Res. 2007, 4, 1–30. [Google Scholar]
- Shukla, A.; Kumar, A.; Jha, A.; Chaturvedi, O.P.; Prasad, R.; Gupta, A. Effects of shade on arbuscular mycorrhizal colonization and growth of crops and tree seedlings in Central India. Agrofor. Syst. 2009, 76, 95–109. [Google Scholar] [CrossRef]
- Seiter, S.; Ingham, E.R.; William, R.D. Dynamics of soil fungal and bacterial biomass in a temperate climate alley cropping system. Appl. Soil Ecol. 1999, 12, 139–147. [Google Scholar] [CrossRef]
- Lacombe, S.; Bradley, R.L.; Hamel, C.; Beaulieu, C. Do tree-based intercropping systems increase the diversity and stability of soil microbial communities? Agric. Ecosyst. Environ. 2009, 131, 25–31. [Google Scholar] [CrossRef]
- Banerjee, S.; Baah-Acheamfour, M.; Carlyle, C.N.; Bissett, A.; Richardson, A.E.; Siddique, T.; Bork, E.W.; Chang, S.X. Determinants of bacterial communities in Canadian agroforestry systems. Environ. Microbiol. 2016, 18, 1805–1816. [Google Scholar] [CrossRef]
- Nelson, J.D.J.; Schoenau, J.J.; Malhi, S.S.; Gill, K.S. Burning and cultivation effects on greenhouse gas emissions and nutrients in wetland soils from Saskatchewan, Canada. Nutr. Cycl. Agroecosyst. 2007, 78, 291–303. [Google Scholar] [CrossRef]
- Richardson, A.E.; Lynch, J.P.; Ryan, P.R.; Delhaize, E.; Smith, F.A.; Smith, S.E.; Harvey, P.R.; Ryan, M.H.; Veneklaaas, E.J.; Lambers, H.; et al. Plant and microbial strategies to improve the phosphorus efficiency of agriculture. Plant Soil 2011, 349, 121–156. [Google Scholar] [CrossRef]
- Ehrmann, J.; Ritz, K. Plant: Soil interactions in temperate multi-cropping production systems. Plant Soil 2014, 376, 1–29. [Google Scholar] [CrossRef] [Green Version]
- Adams, P.W.; Lamoureux, S. The Use of Native Northern Plants for the Re-Vegetation of Arctic Mine Tailings and Mine Waste; Environment and Natural Resources; Government of Northwest Territories: Yellowknife, NT, Canada, 2005; 67p.
- Kedir, A.J.; Nyiraneza, J.; Galagedara, L.; Cheema, M.; Hawboldt, K.A.; McKenzie, D.B.; Unc, A. Phosphorus adsorption characteristics in forested and managed podzolic soils. Soil Sci. Soc. Am. J. 2021, 85, 249–262. [Google Scholar] [CrossRef]
- Magdziak, Z.; Mleczek, M.; Kaczmarek, Z.; Golinski, P. Influence of Ca/Mg ratio and Cd2+ and Pb2+ elements on low molecular weight organic acid secretion by Salix viminalis L. roots into the rhizosphere. Trees 2013, 27, 663–673. [Google Scholar] [CrossRef] [Green Version]
- Kuzovkina, Y.A.; Volk, T.A. The characterization of willow (Salix L.) varieties for use in ecological engineering applications: Co-ordination of structure, function and autecology. Ecol. Eng. 2009, 35, 1178–1189. [Google Scholar] [CrossRef]
- Gordon, A.; Newman, S. Temperate Agroforestry Systems; CAB International: Cambridge, MA, USA, 1997; pp. 263–269. [Google Scholar]
- Rytter, R.M. Fine-root production and turnover in a willow plantation estimated by different calculation methods. Scand. J. For. Res. 1999, 14, 526–537. [Google Scholar] [CrossRef]
- Phillips, C.J.; Marden, M.; Suzanne, L.M. Observations of root growth of young poplar and willow planting types. N. Z. J. For. Sci. 2014, 44, 15. [Google Scholar] [CrossRef] [Green Version]
- Wilton, M.J.; Karagatzides, J.D.; Tsuji, L.J.S. Fertilizing bush beans with locally made compost in a remote subarctic community. Agrosyst. Geosci. Environ. 2020, 3, e20109. [Google Scholar] [CrossRef]
- Ghosh, U.; Chatterjee, A.; Bremer, E. Determining the moisture and plant effect on nutrient release, and plant nutrient uptake using ion exchange resin membrane. Commun. Soil Sci. Plant Anal. 2018, 49, 782–790. [Google Scholar] [CrossRef]
- Sierra, J. Temperature and soil moisture dependence of N mineralization in intact soil cores. Soil Biol. Biochem. 1997, 29, 1557–1563. [Google Scholar] [CrossRef]
- Evers, A.K.; Bambrick, A.; Lacombe, S.; Dougherty, M.C.; Peichl, M.; Gordon, A.M.; Thevathasan, N.V.; Whalen, J.; Bradley, R.L. Potential greenhouse gas mitigation through temperate tree-based intercropping systems. Open Agric. J. 2010, 4, 49–57. [Google Scholar] [CrossRef] [Green Version]
- Aronsson, P.; Perttu, K. Willow vegetation filters for wastewater treatment and soil remediation combined with biomass production. For. Chron. 2001, 77, 293–299. [Google Scholar] [CrossRef] [Green Version]
- Ryszkowski, L.; Kędziora, A. Modification of water flows and nitrogen fluxes by shelterbelts. Ecol. Eng. 2007, 29, 388–400. [Google Scholar] [CrossRef]
- Bergeron, M.; Lacombe, S.; Bradley, R.L.; Whalen, J.; Cogliastro, A.; Jutras, M.F.; Arp, P. Reduced soil nutrient leaching following the establishment of tree-based intercropping systems in eastern Canada. Agrofor. Syst. 2011, 83, 321–330. [Google Scholar] [CrossRef]
- Verwijst, T. Willows: An underestimated resource for environment and society. For. Chron. 2001, 77, 281–285. [Google Scholar] [CrossRef] [Green Version]
- Smart, L.B.; Cameron, K.D. Genetic improvement of willow (Salix spp.) as a dedicated bioenergy crop. In Genetic Improvement of Bioenergy Crops; Vermerris, W., Ed.; Springer: New York, NY, USA, 2008; pp. 347–376. [Google Scholar] [CrossRef]
- Myers-Smith, I.H.; Forbes, B.C.; Wilmking, M.; Hallinger, M.; Lantz, T.; Blok, D.; Hik, D.S. Shrub expansion in tundra ecosystems: Dynamics, impacts and research priorities. Environ. Res. Lett. 2011, 6, 045509. [Google Scholar] [CrossRef] [Green Version]
- Sader, J.A.; Hattori, K.; Hamilton, S.; Brauneder, K. Metal binding to dissolved organic matter and adsorption to ferrihydrite in shallow peat groundwaters: Application to diamond exploration in the James Bay Lowlands, Canada. Appl. Geochem. 2011, 26, 1649–1664. [Google Scholar] [CrossRef]
- Sundström, E.; Magnusson, T.; Hånell, B. Nutrient conditions in drained peatlands along a north-south climatic gradient in Sweden. For. Ecol. Manag. 2000, 126, 149–161. [Google Scholar] [CrossRef]
- Jalali, M.; Arian, T.M.; Ranjbar, F. Selectivity coefficients of K, Na, Ca, and Mg in binary exchange systems in some calcareous soils. Environ. Monit. Assess. 2020, 192, 80. [Google Scholar] [CrossRef]
- Øien, D.I.; Moen, A. Nutrient limitation in boreal plant communities and species influenced by scything. Appl. Veg. Sci. 2001, 4, 197–206. [Google Scholar] [CrossRef]
- Tsialtas, I.T.; Shabala, S.; Baxevanos, D.; Matsi, T. Cation selectivity in cotton (Gossypium hirsutum L.) grown on calcareous soil as affected by potassium fertilization, cultivar and growth stage. Plant Soil 2017, 415, 331–346. [Google Scholar] [CrossRef]
- Assunção, N.S.; Ribeiro, N.P.; da Silva, R.M.; Soratto, R.P.; Fernandes, A.M. Tuber yield and allocation of nutrients and carbohydrates in potato plants as affected by limestone type and magnesium supply. J. Plant Nutr. 2020, 43, 51–63. [Google Scholar] [CrossRef]
- Kaya, M.; Küçükyumuk, Z.; Erdal, I. Effects of elemental sulfur and sulfur-containing waste on nutrient concentrations and growth of bean and corn plants grown on a calcareous soil. Afr. J. Biotech. 2009, 8, 4481–4489. [Google Scholar]
- Wilhelm Scherer, H. Sulfur in soils. J. Plant Nutr. Soil Sci. 2009, 172, 326–335. [Google Scholar] [CrossRef]
- Orlova, J.; Branfireun, B.A. Surface water and groundwater contributions to streamflow in the James Bay Lowland, Canada. Arct. Antarct. Alp. Res. 2014, 46, 236–250. [Google Scholar] [CrossRef] [Green Version]
- Kayser, A.; Schröder, T.J.; Grünwald, A.; Schulin, R. Solubilization and plant uptake of zinc and cadmium from soils treated with elemental sulfur. Int. J. Phytoremediat. 2001, 3, 381–400. [Google Scholar] [CrossRef]
- Rahman, M.M.; Soaug, A.A.; Darwish, F.H.A.; Golam, F.; Sofian-Azirun, M. Growth and nutrient uptake of maize plants as affected by elemental sulfur and nitrogen fertilizer in sandy calcareous soil. Afr. J. Biotech. 2011, 10, 12882–12889. [Google Scholar] [CrossRef] [Green Version]
- Rezapour, S. Effect of sulfur and composted manure on SO4-S, P and micronutrient availability in a calcareous saline–sodic soil. Chem. Ecol. 2014, 30, 147–155. [Google Scholar] [CrossRef]
- Ruiz, J.M.; López-Cantarero, I.; Rivero, R.M.; Romero, L. Sulphur phytoaccumulation in plant species characteristic of gypsiferous soils. Int. J. Phytoremediat. 2003, 5, 203–210. [Google Scholar] [CrossRef] [PubMed]
- Plaster, E.J. Soil Science and Management, 6th ed.; Delmar: Clifton Park, NY, USA, 2014; pp. 300–301. [Google Scholar]




{kind=link}
{kind=link}
{kind=link}
| Site | p | Crop | p | Month | p | Site × Crop | p | Site × Month | p | Crop × Month | p | Site × Crop × Month | p | |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| NO3 | 2.19 | 0.146 | 0.93 | 0.402 | 5.25 | 0.003 | 0.26 | 0.774 | 2.656 | 0.059 | 0.295 | 0.937 | 0.19 | 0.978 |
| PO4 | 31.23 | 1.1 × 10−6 | 0.11 | 0.898 | 1.01 | 0.395 | 0.02 | 0.980 | 1.41 | 0.251 | 1.74 | 0.133 | 1.13 | 0.359 |
| K | 0.85 | 0.361 | 0.74 | 0.485 | 3.57 | 0.021 | 0.87 | 0.426 | 3.82 | 0.016 | 1.24 | 0.301 | 0.82 | 0.557 |
| Ca | 9.59 | 0.003 | 0.32 | 0.730 | 4.58 | 0.007 | 0.60 | 0.556 | 0.60 | 0.615 | 0.70 | 0.648 | 2.08 | 0.073 |
| Mg | 61.22 | 4.1 × 10−10 | 1.18 | 0.316 | 0.88 | 0.456 | 1.854 | 0.168 | 0.91 | 0.445 | 0.69 | 0.659 | 1.37 | 0.247 |
| SO4 | 301.76 | 2.4 × 10−22 | 0.38 | 0.683 | 4.89 | 0.005 | 0.36 | 0.699 | 6.29 | 0.001 | 0.47 | 0.829 | 0.099 | 0.996 |
| B | 4.90 | 0.032 | 0.59 | 0.561 | 3.32 | 0.027 | 1.06 | 0.354 | 6.02 | 0.001 | 0.19 | 0.979 | 0.46 | 0.837 |
| Al | 3.44 | 0.070 | 1.61 | 0.211 | 4.34 | 0.009 | 0.45 | 0.639 | 0.57 | 0.637 | 1.39 | 0.240 | 0.69 | 0.659 |
| Cu | 1.22 | 0.274 | 0.28 | 0.756 | 5.32 | 0.003 | 0.53 | 0.593 | 2.08 | 0.116 | 0.25 | 0.956 | 0.59 | 0.737 |
| Fe | 1.68 | 0.201 | 0.38 | 0.685 | 4.28 | 0.009 | 0.81 | 0.450 | 0.89 | 0.454 | 0.53 | 0.780 | 0.43 | 0.855 |
| Mn | 2.81 | 0.100 | 0.94 | 0.398 | 1.79 | 0.162 | 1.29 | 0.285 | 0.78 | 0.511 | 0.66 | 0.678 | 0.88 | 0.517 |
| Zn | 28.08 | 2.9 × 10−6 | 1.74 | 0.187 | 43.50 | 9.9 × 10−14 | 0.26 | 0.773 | 2.75 | 0.053 | 0.78 | 0.594 | 0.98 | 0.449 |
| Bush Bean | June PRS + Leaves | August PRS + Leaves | Yield | |||
| Soil | r | P | r | p | r | p |
| NO3–N | 0.57 | 0.06 | 0.28 | 0.38 | 0.05 | 0.88 |
| PO4–P | 0.60 | 0.04 | 0.25 | 0.42 | 0.56 | 0.06 |
| K | 0.45 | 0.14 | 0.14 | 0.66 | −0.16 | 0.61 |
| Ca | −0.27 | 0.40 | 0.41 | 0.19 | 0.72 | 0.01 |
| Mg | −0.20 | 0.53 | −0.37 | 0.23 | −0.28 | 0.38 |
| B | −0.05 | 0.44 | 0.16 | 0.31 | −0.20 | 0.54 |
| Fe | −0.22 | 0.50 | −0.01 | 0.99 | −0.22 | 0.50 |
| Mn | −0.21 | 0.52 | −0.24 | 0.46 | −0.15 | 0.64 |
| Zn | 0.52 | 0.08 | 0.76 | 0.004 | 0.70 | 0.01 |
| Cu | −0.15 | 0.65 | −0.34 | 0.28 | −0.30 | 0.35 |
| Al | -- | -- | -- | -- | 0.16 | 0.63 |
| SO4 | -- | -- | -- | -- | −0.54 | 0.07 |
| Potato | June PRS + Leaves | August PRS + Leaves | Yield | |||
| Soil | r | p | r | P | r | p |
| NO3–N | 0.50 | 0.10 | 0.43 | 0.16 | −0.05 | 0.88 |
| PO4–P | 0.71 | 0.01 | 0.69 | 0.01 | 0.72 | 0.01 |
| K | 0.06 | 0.84 | 0.13 | 0.69 | 0.26 | 0.42 |
| Ca | −0.03 | 0.92 | −0.58 | 0.05 | 0.36 | 0.25 |
| Mg | 0.46 | 0.14 | 0.34 | 0.28 | −0.61 | 0.04 |
| B | −0.10 | 0.75 | 0.06 | 0.87 | −0.54 | 0.07 |
| Fe | −0.08 | 0.80 | −0.24 | 0.44 | 0.46 | 0.13 |
| Mn | −0.30 | 0.34 | −0.19 | 0.55 | 0.42 | 0.18 |
| Zn | −0.12 | 0.72 | 0.25 | 0.44 | 0.51 | 0.09 |
| Cu | −0.06 | 0.85 | 0.26 | 0.42 | 0.60 | 0.04 |
| Al | -- | -- | -- | -- | −0.81 | 0.002 |
| SO4 | -- | -- | -- | -- | −0.79 | 0.002 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Karagatzides, J.D.; Wilton, M.J.; Tsuji, L.J.S. Soil Nutrient Supply in Cultivated Bush Bean–Potato Intercropping Grown in Subarctic Soil Managed with Agroforestry. Sustainability 2021, 13, 8185. https://0-doi-org.brum.beds.ac.uk/10.3390/su13158185
Karagatzides JD, Wilton MJ, Tsuji LJS. Soil Nutrient Supply in Cultivated Bush Bean–Potato Intercropping Grown in Subarctic Soil Managed with Agroforestry. Sustainability. 2021; 13(15):8185. https://0-doi-org.brum.beds.ac.uk/10.3390/su13158185
Chicago/Turabian StyleKaragatzides, Jim D., Meaghan J. Wilton, and Leonard J. S. Tsuji. 2021. "Soil Nutrient Supply in Cultivated Bush Bean–Potato Intercropping Grown in Subarctic Soil Managed with Agroforestry" Sustainability 13, no. 15: 8185. https://0-doi-org.brum.beds.ac.uk/10.3390/su13158185