
Predicting the Areas of Suitable Distribution for Zelkova serrata in China under Climate Change

1
College of Horticultural Science and Technology, Suzhou Polytechnic Institute of Agriculture, Suzhou 215008, China
2
College of Horticulture and Plant Protection, Yangzhou University, Yangzhou 225009, China
*
Author to whom correspondence should be addressed.
Sustainability 2021, 13(3), 1493; https://0-doi-org.brum.beds.ac.uk/10.3390/su13031493
Submission received: 17 December 2020
/
Revised: 21 January 2021
/
Accepted: 28 January 2021
/
Published: 1 February 2021
(This article belongs to the Special Issue Hydrology and Climate-Change Impact Assessment and Adaptation)
Abstract
:Predicting the geographic distribution of a species together with its response to climate change is of great significance for biodiversity conservation and ecosystem sustainable development. Zelkova serrata is an excellent shelterbelt tree species that is used for soil and water conservation due to the fact of its well-developed root system, strong soil fixation, and wind resistance. However, the wild germplasm resources of Z. serrata have been increasingly depleted due to the fact of its weak ability to regenerate naturally and the unprecedented damage humans have caused to the natural habitats. The present work using Maxent aimed to model the current potential distribution of this species as well as in the future, assess how various environmental factors affect species distribution, and identify the shifts in the distribution of this species in various climate change scenarios. Our findings show habitat in provinces in the southern Qinling and Huai river basins have high environmental suitability. Temperature seasonality, annual precipitation, annual mean temperature, and warmest quarter precipitation were the most important factors affecting its distribution. Under a climate change scenario, the appropriate habitat range showed northeastward expansion geographically. The results in the present work can lay the foundation for the cultivation and conservation of Z. serrata.
1. Introduction
Climate has been suggested to be the major factor that affects the distribution of many species at a large scale [1,2]. Global climate changes have led to distribution shifts of numerous species, which may become a direct leading cause of the extinction of many species in the near future; alternatively, it may act synergistically with other extinction drivers [2,3,4,5]. These climate effects are generally referred to as alterations in climate that affect the distribution [5,6], structure, and composition of species [7,8] in the forests. Also, climate affects the fruiting and flowering phenology [2], life history characters [9,10,11], as well as the habitat requirements of many species [2,6]. As a result, predicting the effects of future climate change on the spatial distribution of habitat across the landscape for individual species this will help managers address any possible issues that can potentially change species distribution and range [11].
Notably, species distribution modeling (SDM) plays a vital role in predicting species ecological requirements together with the possible spatial distribution of species distribution in the fields of regional ecology and biogeography, when limited distribution information is available [12]. A variety of SDM models have been adopted for predicting distribution area, ecological response, and ecological requirements [12]. Among these modeling approaches, bioclimatic envelope models such as CLIMEX, Maximum entropy modeling (Maxent), Domain, and GARP, are extensively used for predicting the potential distributions of species based on their ecological and climatic profiles [13], with the assumption that climate is the primary determinant of the distribution of plant species [14]. Maxent has been extensively applied and found to perform well in many situations including with small sample sizes [3,11,15,16,17,18].
Zelkova serrata (Ulmaceae), a common deciduous tree, is mainly distributed in the south of the Qinling Mountains and Huai River basins and grows in the river valley and sparse forest habitat beside streams at elevations of 500–1900 m [19]. Because of its well-developed root system, strong ability to promote soil conservation and to prevent soil erosion, it is an excellent tree species for use in wind breaks, habitat restoration, and in soil and water conservation. At the same time, Z. serrata is a good ornamental tree because of its tall trunk and rich seasonal changes in leaf color [20,21,22,23,24,25]. Previous studies have shown that an increase in winter temperature decreased seedling emergence in the follow spring [24]. Furthermore, due to the fact of its weak natural regeneration ability, tourism, unsustainable harvesting, and the increased frequency of extreme weather, the wild resources are nearly exhausted, so that Z. serrata has been listed as the national second-class key protected plant [20]. Previous studies have mainly focused on horticulture and landscape uses [22], seedling technology [23], seedling growth [24], and the mechanism involved in leaf color change [25], but knowledge related to its habitat, distribution, and ecological requirements that affect the sustainability of harvests remain largely unclear. Therefore, it is highly significant to explore the relationship between Z. serrata and environment factors as well as to determine those factors that influence the distribution of this species.
In this work, a wide spectrum of field survey data and geo-referenced collections were analyzed by means of Maxent modeling in predicting the distribution of Z. serrata. Specifically, this work aimed to (1) identify the most significant environmental variables affecting the range and distribution of Z. serrata, (2) examine the spatial extent of appropriate habitat of the species by using present and future climate modeling scenarios based on natural and variation trends, and (3) quantify these patterns of spatial variation within the extent of appropriate habitat by using future climate scenarios to facilitate the formulation of suitable conservation measures while providing a theoretical reference for afforestation, management, and use of this species.
2. Materials and Methods
2.1. Collection of Occurrence Locations
The complete locations (latitude and longitude) with distribution of Z. serrata across China were extracted based on the Chinese Virtual Herbarium (http://v5.cvh.org.cn/) and the Global Biodiversity Information Facility (http://www.gbif.org/) databases. In addition, certain distribution data were acquired through field investigation and literature review. The dataset was assessed by the criteria as suggested by Boitani et al. [26]. Moreover, this study only used those locations offered from online search engines online or the local flora, so as to guarantee that such location data represented naturally or permanently present occurrence area. Meanwhile, data with inaccurate locations and/or lacking precise geo-coordinates were eliminated out from this study. Duplicates were eliminated, and the remaining points were then screened so that one point was plotted in each 1.0 × 1.0 km2 grid cell. Altogether, 172 geo-referenced records were adopted for modelling.
2.2. Environmental Factors
A total of 30 environmental factors were screened as possible predictors of Z. serrata habitat distribution with reference to other research on SDMs as well as analyzing the biological relevance. The following data were included. First, 19 bioclimatic parameters were used, which had a spatial resolution of 30 s (approximately 1 km) and were collected based in the World Climate Database (www.worldclim.org) [27]. In addition, elevation data were collected using the Geospatial Data Cloud (http://www.gscloud.cn) based on a digital elevation model with a resolution of 30 × 30 m2. Moreover, soil organic carbon (SOC), soil pH (SpH), and growing degree day (GDD) data were collected based on the Center for Sustainability and the Global Environment (http://www.sage.wisc.edu/atlas/index.php) [28]. In addition, vapor pressure (VAP), wet-day frequency (WET), and ground-frost frequency (FRS) were extracted based on the Intergovernmental Panel on Climate Change (IPCC) database (http://www.ipcc-data.org/obs/cru_ts2_1.html); data on global UV-B radiation (UVB1-4) were obtained based on gIUV database (http://www.ufz.de/gluv/) [29].
The Beijing Climate Center – Climate System Model (BCC-CSM) 1.1 modeling data were used as the simulated future climate at Representative Concentration Pathway (RCP) 2.6, 4.5, and 8.5 for 2070, which were released in the the IPCC Assessment Report 5 (AR5). Typically, BCC-CSM 1.1 is suggested to be used in climate change studies in China [30]. The RCP 2.6 scenario can reflect the possible radiative forcing in 2100 relative to the positive + 2.6 W/m2 pre-industrial value; by contrast, RCP 8.5 is more negative, which stands for the extensive greenhouse gas emissions, with a radiative forcing value of 8.5 W/m2 by 2100.
The spatial resolution of 1 km was used for resampling each environmental parameter. In addition, each parameter was clipped within the research region. Subsequently, ArcGIS 10.0 and a WGS84 projection system were used to process each layer with the identical spatial extent and cell size. Thereafter, all variables were analyzed through the principal component analysis (PCA) and Pearson correlation analysis. We only chose one parameter for great cross-correlation (r2 > 0.90) according to the biological significance to distribution of Z. serrata. Principal component analyses were used to selected which variables were retained (for more detail see Yi et al. [11]). Ultimately, only 21 predictors were used (Table 1).
2.3. Fitting of Model
This study adopted MaxEnt (MaxEnt 3.3.3) in modelling. Generally, Maxent can produce models according to the maximum entropy principle (details are available in Phillips et al. [31]), which produces models through identifying the closest distribution to the uniform distribution (namely, the maximum entropy) for every environmental parameter in the research region. In this study, 75% data were adopted for training, while the remaining 25% were used to test the capacity of our model to predict the distribution of species. Altogether 1000 iterations were conducted, so that our model had enough convergence time. In addition, this study adopted 1 × 10−6 to be the threshold for convergence [32]. We tested various regularization multiplier values and then chose the default (i.e., 1) option because it performed best; that is, provided the best representation of the known current distribution. In addition, we used the default “autofeatures,” and the “logistic” output format.
For model calibration and robustness validation, threshold-independent ROC curve analysis was carried out. Then, the AUC values were analyzed to verify the model precision. Additionally, every parameter was assessed for its relative importance using a Jackknife test. For Z. serrata, every pixel observed on the potential distribution map was classified between 0 and 1. Several habitat suitability classification methods have been used in the past, most of which were based on experience [33,34,35,36]. Li et al. [37] pointed that grid points with probabilities higher than 0.6 should be considered as the most suitable habitat or core area. The main objective of the present study was to find the location of the regions with the most suitable habitats, and provide guidelines related to the conservation of Z. serrata. Based on our experience, we classified such girds as four possible habitat types: “high” (>0.6), “moderate” (0.4–0.6), “low” (0.2–0.4), and “not potential” (<0.2) habitat.
When the current habitat appropriate for Z. serrata was modeled, Maxent conducted the modeling for the four kinds of the abovementioned future climate situations based on the available climate data. Therefore, the appropriate future habitats might be predicted for those species. In addition, SDM projections were calculated by Maxent software. The future appropriate habitat areas of Z. serrata were crosschecked relative to those present distribution areas, so as to identify regions with little change, including suitable or unsuitable areas. Additionally, for every region type, its area was analyzed.
2.4. Shift in the Distribution Center of Suitable Habitat
The SDM toolbox, the GIS toolkit operating based on Python, was employed to calculate and compare centroids between present and future appropriate regions for this species [38]. This study used an SDM toolbox (including present as well as future SDMs) for calculating changes in the distributions of the above two binary SDMs [38], while analyzing those primary shifts in Z. serrata distribution. The species distributions were reduced to one individual centroid point. Then, for those time-dependent estimated variations, their magnitudes and directions were created. The movement of centroid of various SDMs were tracked to examine the distributional shifts.
3. Results
3.1. Species Distribution Model as well as Model Accuracy
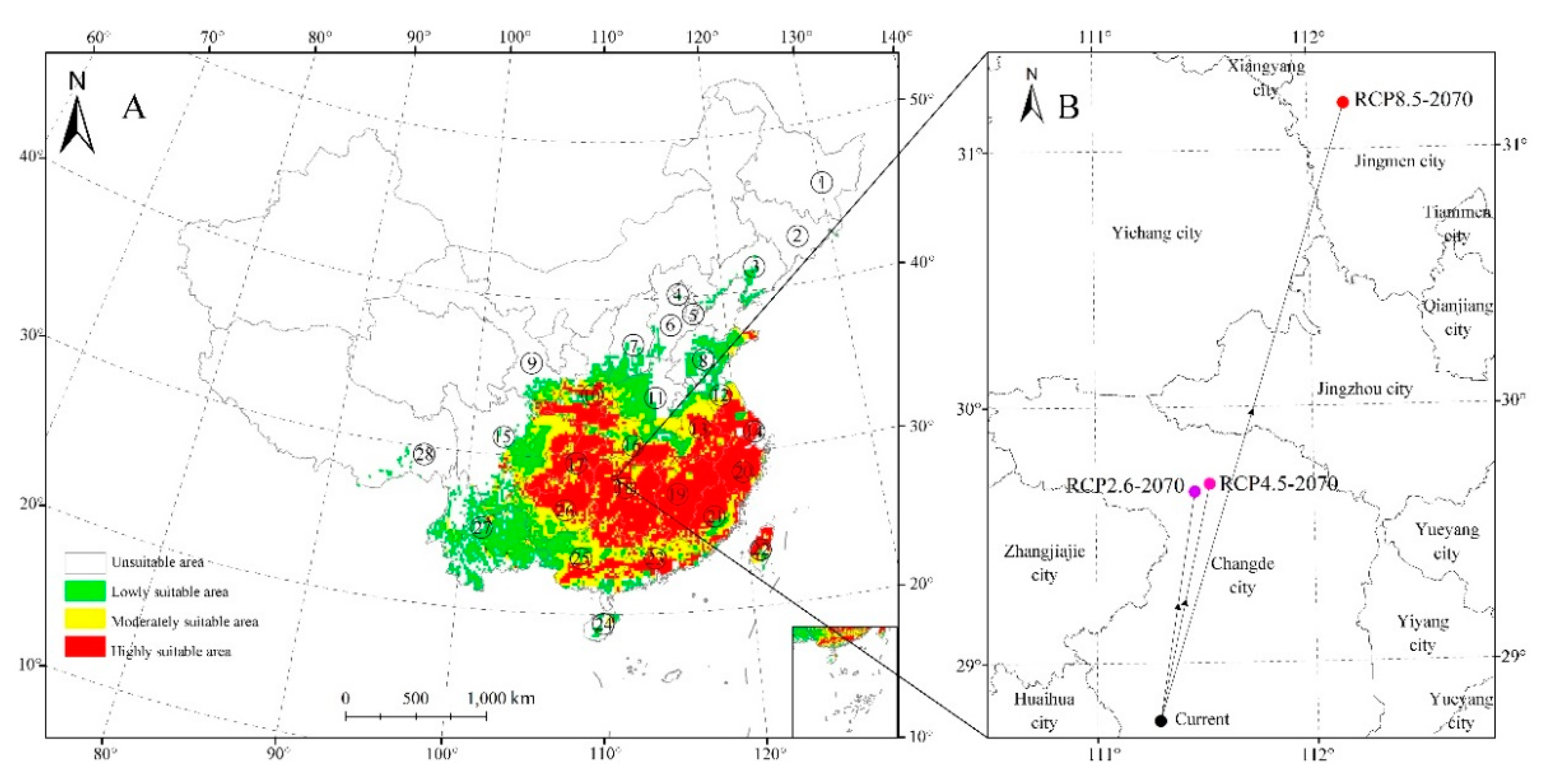
For our model, the mean value of the AUC for the training subset of samples was 0.924 (Figure 1). We predicted that the currently highly appropriate habitats for Z. serrata were in southern Jiangsu, southern Anhui, Shanghai, Zhejiang, Jiangxi, southern Shaanxi, northern Guizhou, Hunan, southern Hubei, Chongqing, eastern Fujian, eastern central Taiwan, central Guangdong, and central Guangxi (Figure 2A). The area of highly suitable areas accounts for 11.9% of total distribution areas.
3.2. Important Environmental Factors Affecting the Distribution of Habitat
Of those 21 parameters used for model establishment, annual precipitation (Bio 12, 44.9%), annual mean temperature (Bio 1, 17%), warmest quarter precipitation (Bio 18, 8.1%), together with temperature seasonality (Bio 4, 6%) showed high permutation importance (Table 1). For the above four parameters, their accumulated contribution rate reached up to 76%, indicating that such factors exerted a vital role in possible Z. serrata distribution. The preferred climate of Z. serrata was generated based on the above four variables in Maxent (Figure 3). Overall, a positive nonlinear response was observed for Bio 12, while a negative nonlinear response was observed for Bio 4. The optimum annual mean temperature and precipitation of warmest quarter were ca 18 °C and 520 mm, respectively.
3.3. Variations of Future Spatial Appropriate Habitat Extent
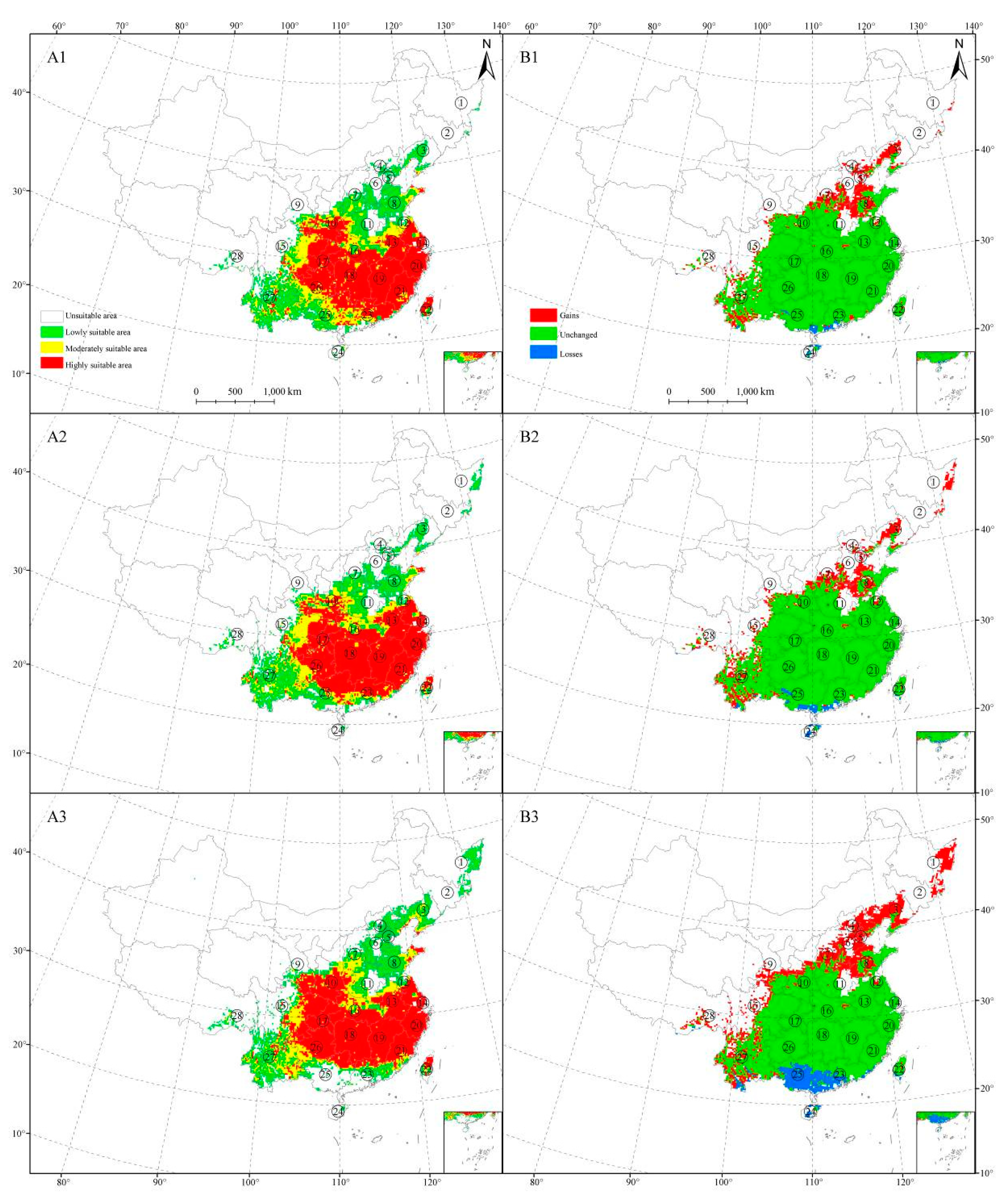
In general, a limited expansion of range was seen across the appropriate habitat area under climate change scenarios. According to the scenario that had low emission levels of greenhouse gas (RCP 2.6), Maxent predicted that, the appropriate habitat extent increased in western Yunnan, central and western Shandong, central Shanxi, southern Hebei, central Liaoning, Tianjin, and Beijing, at the same time, the area of suitable habitat losses will mainly be scattered in central Yunnan and southern Guangdong (Figure 4A1,B1). Under the RCP 4.5 scenario, Maxent predicted a small expansion of its habitat than the climate scenario of RCP 2.6 and the change area occurred the same provinces at the climate scenario of RCP 2.6 (Figure 4A2,B2). In the RCP 8.5 scenario, the distribution area continued to expand. There is an increase in the distribution area in the east of Heilongjiang, while the decrease in the distribution area mainly concentrated in Yunnan (Xishuangbanna), Guangdong (Zhanjiang, Maoming, Yangjiang, Jiangmen, Yunfu, Dongguan, and Zhongshan) and most parts of Guangxi and central Hainan (Figure 4A3,B3).
3.4. Shift of Appropriate Habitat Distribution Center
Currently, the Z. serrata distribution centroid was predicted to be in Heishui County of Sichuan Province (111°17′ E, 28°15′ N; Figure 3B). In addition, the appropriate habitat centroid was estimated to show a shift to 29°41′ N and 117°27′ E, 29°43′ N, and 112°70′ E, and 31°13′ E and 112°7′ N under RCP2.6, 4.5 and 8.5, respectively. In brief, the distribution center of suitable habitat will shift to north (northeast).
4. Discussion
Zelkova serrata is an excellent protective forest and soil and water conservation tree species. However, its habitat distribution is being anthropogenically disturbed and influenced by climate change. This work comprehensively analyzed the kinds of habitat that are appropriate for Z. serrata and provides a vital link to help land managers to develop policies and plans to support the sustainably use and management of this important species. In this study, Maxent predicts a wide distribution of Z. serrata across China. In particular, these areas are usually in provinces of south Qinling and the Huai River. The models constructed in previous research conformed to ours in this work [19]. Nonetheless, this study predicted a broader possible extent of the range of this species in the future. Yunnan, Guangxi, and eastern Sichuan have large-scale areas that were predicted to serve as appropriate habitat, which offers an enlarged region for Z. serrata cultivation.
Of the 21 elements employed in the model, annual precipitation, annual mean temperature, warmest quarter precipitation, and temperature seasonality were the dominant parameters that drove the potential distribution of Z. serrata, which suggested that these parameters were important for species distribution. The amount and patterns of annual precipitation are the vital parameters for plant survival, regeneration and additional functions of the ecosystem [39]. Precipitation directly affects the growth and morphology of Z. serrata as well as affects the accumulation of plant biomass [40]. Reductions in the amount of rainfall resulted in reductions in plant height and biomass accumulation in Z. serrata and in decreased seed production. Previous studies have shown that the plastic response of seedling growth [40], seed germination [41], biomass allocation [42], physiological characteristic [43], and phenology [42] varies with annual precipitation.
The tolerance to a wide temperature range accounts for a vital factor for explaining the species latitudinal distribution [44]. Zelkova serrata occurs extensively in the subtropical and warm temperate areas in China [19]. It tolerates high temperatures with an optimum annual mean temperature of 18 °C for this species. Temperature changes have a certain effect on the distribution of Z. serrata by impacting growth, germination, photosynthesis, water absorption, transpiration, reproduction and respiration. It has previously been suggested that, for Z. serrata, its seedling emergence were under the direct influence of winter temperature [40]. As a result, the above hydrothermal parameters might affect Z. serrata distribution.
Besides the environmental factors listed above, the suitable habitats of this plant can be influenced by some other factors, including competition between species, natural enemy predation, low seed quality, anthropogenic impact, human activities, geographical barrier, and changes in habitats [45,46,47]. Those factors were not taken into consideration in the distribution model due to the lack of robust data. Research is still required to qualify the effects of human activity on the adaptation of this species to future climates.
Currently, unprecedented attention has been placed on the effects of climatic change worldwide [4]. Extreme climate events show an increasing trend in terms of the severity and occurrence [1,5]. The increased temperature can boost a number of physiological processes. For instance, photosynthesis can achieve the upper limit within plants as the temperature becomes elevated, regardless of the varying responses of diverse plant species. Therefore, this may result in extirpation of species regional or local scales, or perhaps the loss of entire ecosystems which may be replaced by other ecosystems [39,45]. In this study, according to our applied models, the appropriate spatial climate extent for Z. serrata showed expansion in geography, in particular in the northeastward direction with climatic warming. According to the above discussion, temperature and precipitation were the vital parameters that affected the distribution of Z. serrata. The species will be unable to tolerate a continuous increase in temperature. Additionally, changes of precipitation and temperature might lead to phenological shifts of Z. serrata, while this probably had an indirect effect on dependent floral and faunal species. On the other hand, such alterations might adversely affect numerous terrestrial mammals, birds, and insects directly or indirectly if they depend on seeds of Z. serrata or its flowers and fruits [48,49,50]. Therefore, the area of suitable areas in Guangdong, Yunnan, Guangxi, and Hainan will be significantly reduced. The northeastward movement of suitable areas can alleviate the harm caused by high temperatures. This also explains why the areas in southern Hebei, central Liaoning, Tianjin, and Beijing has increased significantly.
Results obtained by the model we established can be tailored to satisfy the Z. serrata conservation guidelines by identifying the habitats that are climatically appropriate and critically susceptible to climate change; artificial regeneration should be taken into consideration in the process of reforestation. For instance, our models predicted that the suitable Z. serrata habitats shifted northeastward beyond the present natural distribution area. Nonetheless, the natural migration rate is not high; as a result, assisted Z. serrata migration may be used as a possible conservation strategy when the anticipated significant future climate changes generate novel climatically suitable habitat [45]. In this regard, the creation of Z. serrata plantations is proposed in the new climatically suitable habitats. However, it is important to note that the proposal for more northern plantations should be supported by the identification of forest areas to be targeted, which are realistic and practical and not competing with other land uses. In addition, the natural regeneration for this species should be preserved in places that are identified to be of high risk in future climate scenarios. Additionally, the gain or non-change in climate space observed across different ecoregions of Z. serrata are identified as possible climate change refugia. At present, the Z. serrata trees grow in different types of climate, which facilitate species adaptation to new climates in the natural habitats [51,52,53]. As a result, applying phenotypic plasticity and selecting genotypes that are able to adapt to scenarios in future climates can improve the persistence of Z. serrata.
For promoting the sustainable development and management of Z. serrata as well as the relevant forest ecosystems, the strategies below are recommended. (1) Ex situ and in situ wild Z. serrata conservation strategies together with relevant information networks must be formulated [45]. (2) Efforts should be made to minimize anthropogenic disturbance. (3) Attention should be paid to establish a database that can be used to systemically and persistently monitor the Z. serrata population dynamics [40]. (4) For those high-risk areas experiencing climate change, forest managers must attempt to introduce different known species suitable for certain climate scenarios, but not further formulate the novel use of Z. serrata plantations [45]. (5) Forest management agencies should be provided with increased funding to support comprehensive surveys for analyzing and understanding the current situation associated with wild Z. serrata [40,41,42]. Therefore, land managers should make efforts to formulate management strategies and practical measures that can be used to conserve valuable wild Z. serrata resources [29].
5. Conclusions
Using Maxent modeling, this study identified extensive habitat for Z. serrata in the presence of climate scenarios at present and in the future. Annual precipitation (Bio 12, 44.9%), annual mean temperature (Bio 1, 17%), warmest quarter precipitation (Bio 18, 8.1%), together with temperature seasonality (Bio 4, 6%) are the vital factors that shape Z. serrata distribution. Under climate change scenarios, appropriate habitat extent shows a northeastward expansion geographically. In this work, the maps generated help us to quantitatively assess regional climate change-related risks that might potentially affect the Z. serrata cultivation. As a result, this work can provide precious data associated with forest management planning in response to the global climate change. Moreover, our results can be used for optimizing field surveys and detecting those unknown populations in the future.
Author Contributions
Conceptualization, C.C. and J.T.; methodology, C.C.; formal analysis, C.C. and J.T.; data curation, J.T.; writing—original draft preparation, C.C.; writing—review and editing, C.C. and J.T.; supervision, J.T. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by the Forestry Science and Technology Promotion and Demonstration Fund of Central Finance (Grant No. (2018)TG08) and the Construction of Jiangsu Modern Agricultural Industry Technology System (Grant No. JATS (2019)448).
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
No new data were created or analyzed in this study. Data sharing is not applicable to this article.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Poortinga, W.; Whitmarsh, L.; Steg, L.; Böhm, G.; Fisher, S. Climate change perceptions and their individual-level determinants: A cross-European analysis. Global Environ. Chang. 2019, 55, 25–35. [Google Scholar] [CrossRef]
- Hossain, M.S.; Arshad, M.; Qian, L.; Kächele, H.; Khan, I.; Islam, M.D.I.; Mahboob, M.G. Climate change impacts on farmland value in Bangladesh. Ecol. Indic. 2020, 112, 106181. [Google Scholar] [CrossRef]
- Pearson, R.G.; Stanton, J.C.; Shoemaker, K.T.; Aiello-Lammens, M.E.; Ersts, P.J.; Horning, N.; Fordham, D.A.; Raxworthy, C.J.; Ryu, H.Y.; McNees, J.; et al. Life history and spatial traits predict extinction risk due to climate change. Nat. Clim. Chang. 2014, 4, 217–221. [Google Scholar] [CrossRef] [Green Version]
- Román-Palacios, C.; Wiens, J.J. Recent responses to climate change reveal the drivers of species extinction and survival. Proc. Natl. Acad. Sci. USA 2020, 117, 4211–4217. [Google Scholar] [CrossRef] [PubMed]
- Timm, K.M.; Maibach, E.W.; Boykoff, M.; Myers, T.A.; Broeckelman-Post, M.A. The prevalence and rationale for presenting an opposing viewpoint in climate change reporting: Findings from a US national survey of TV weathercasters. Weather Clim. Soc. 2020, 12, 103–115. [Google Scholar] [CrossRef]
- Yan, H.; Feng, L.; Zhao, Y.; Feng, L.; Wu, D.; Zhu, C. Prediction of the spatial distribution of Alternanthera philoxeroides in China based on ArcGIS and Maxent. Glob. Ecol. Conserv. 2020, 21, e00856. [Google Scholar] [CrossRef]
- Negrini, M.; Fidelis, E.G.; Picanço, M.C.; Ramos, R.S. Mapping of the Steneotarsonemus spinki invasion risk in suitable areas for rice (Oryza sativa) cultivation using Maxent. Exp. Appl. Acarol. 2020, 80, 445–461. [Google Scholar] [CrossRef]
- Gonzalez, P.; Kroll, B.; Vargas, C.R. Tropical rainforest biodiversity and aboveground carbon changes and uncertainties in the Selva Central, Peru. Forest Ecol. Manag. 2014, 312, 78–91. [Google Scholar] [CrossRef]
- Peterson, A.T.; Papes, M.; Eaton, M. Transferability and model evaluation in ecological niche modeling: A comparison of GARP and Maxent. Ecography 2007, 30, 550–560. [Google Scholar] [CrossRef]
- Zhang, W.X.; Kou, Y.X.; Zhang, L.; Zeng, W.D.; Zhang, Z.Y. Suitable distribution of endangered species Pseudotaxus chienii (Cheng) Cheng (Taxaceae) in five periods using niche modeling. Chin. J. Ecol. 2020, 39, 600–613. [Google Scholar]
- Yi, Y.J.; Cheng, X.; Yang, Z.F.; Zhang, S.H. Maxent modeling for predicting the potential distribution of endangered medicinal plant (H. riparia Lour) in Yunnan, China. Ecol. Eng. 2016, 92, 260–269. [Google Scholar] [CrossRef]
- Guisan, A.; Zimmermann, N.E. Predictive habitat distribution models in ecology. Ecol. Model. 2000, 135, 147–186. [Google Scholar] [CrossRef]
- Butt, N.; Seabrook, L.; Maron, M.; Law, B.S.; Dawson, T.P.; Syktus, J.I.; McAlpne, A. Cascading effects of climate extremes on vertebrate fauna through changes to low-latitude tree flowering and fruiting phenology. Global Change Biol. 2015, 21, 3267–3277. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, Y.; Tang, Z.Y.; Yan, Y.J.; Wang, K.; Cai, L.; He, J.S.; Gu, S.; Yao, Y.J. Incorporating species distribution model into the red list assessment and conservation of macrofungi: A case study with Ophiocordyceps sinensis. Biodiver. Sci. 2020, 28, 99–106. [Google Scholar]
- Elith, J.; Graham, C.H.; Anderson, R.P.; Dudík, M.; Ferrier, S.; Guisan, A.; Hijmans, R.J.; Huettmann, F.; Leathwick, J.R.; Lehmann, A.; et al. Novel methods improve prediction of species’ distributions from occurrence data. Ecography. 2006, 29, 129–151. [Google Scholar] [CrossRef] [Green Version]
- Elith, J.; Kearney, M.; Phillips, S. The art of modelling range-shifting species. Methods Ecol. Evol. 2010, 1, 330–342. [Google Scholar] [CrossRef]
- Hernandez, P.A.; Franke, I.; Herzog, S.K.; Pacheco, V.; Paniagua, L.; Quintana, H.L.; Soto, A.; Swenson, J.J.; Tovar, C.; Valqui, T.H.; et al. Predicting species distributions in poorly-studied landscapes. Biodivers. Conserv. 2008, 17, 1353–1366. [Google Scholar] [CrossRef]
- Støa, B.; Halvorsen, R.; Stokland, J.N.; Gusarov, V.I. How much is enough? Influence of number of presence observations on the performance of species distribution models. Sommerfeltia 2019, 39, 1–28. [Google Scholar]
- Fu, L.; Xin, Y.; Alan, W. Ulmaceae. In Flora of China; Wu, C.Y., Raven, P.H., Hong, D.Y., Eds.; Science Press: Beijing, China; Missouri Botanical Garden Press: St. Louis, MI, USA, 2003; Volume 5, pp. 1–19. [Google Scholar]
- Wang, L.D.; Zhang, R.Q. The research progress of Zelkova serrata. Guangxi Forest. Sci. 2005, 4, 188–191. [Google Scholar]
- Zhang, C.T.; Hu, X.J.; Luo, X.M.; Zhong, F.X.; Jin, X.L. Cutting propagation techniques of Zelkova serrata. Nonwood For. Res. 2011, 29, 108–110. [Google Scholar]
- Ru, L.M.; Zhang, Y.W.; Jiang, W.B. Cultural connotations of Zelkova serrata and its application in landscape architecture. Guangdong Landsc. Archit. 2007, 6, 50–52. [Google Scholar]
- Kang, R. The Research on Zelkova serrata Seed Germination and Seedling Growth Characteristics of Laws. Master’s Thesis, Central South University of Forestry & Technology, Changsha, China, 2010. [Google Scholar]
- Li, S.Q. Breeding Methods of Zelkova serrata. J. Jiangsu For. Sci. Tech. 2000, 3, 39–41. [Google Scholar]
- Zhang, M.; Huang, L.B.; Zhou, B.; Qian, M.; Dou, Q.Q. Physiological and biochemical changes in Zelkova serrata leaves during leaf color transformation in autumn. Sci. Silvae Sin. 2015, 51, 44–51. [Google Scholar]
- Boitani, L.; Maiorano, L.; Baisero, D.; Falcucci, A.; Visconti, P.; Rondinini, C. What spatial data do we need to develop global mammal conservation strategies? Philos. Trans. R. Soc. Lond. B Biol. Sci. 2011, 366, 2623–2632. [Google Scholar] [CrossRef] [Green Version]
- Hijmans, R.J.; Cameron, S.E.; Parra, J.L.; Jones, P.G.; Jarvis, A. Very high resolution interpolated climate surfaces for global land areas. Int. J. Climatol. 2015, 25, 1965–1978. [Google Scholar] [CrossRef]
- New, M.; Hulme, M.; Jones, P. Representing twentieth-century space-time climate variability. Part I: Development of a 1961–1990 mean monthly terrestrial climatology. J. Clim. 1999, 12, 829–856. [Google Scholar] [CrossRef]
- Beckmann, M.; Václavík, T.; Manceur, A.M.; Šprtová, L.; Wehrden, H.; Welk, E.; Cord, A.F. glUV: A global UV-B radiation data set for macroecological studies. Methods Ecol. Evol. 2014, 5, 372–383. [Google Scholar] [CrossRef]
- Wu, T.W.; Song, L.C.; Li, W.P.; Wang, Z.Z.; Zhang, H.; Xin, X.G.; Zhang, Y.W.; Zhang, L.; Li, J.L.; Wu, F.H.; et al. An overview of BCC climate system model development and application for climate change studies. J. Meteorol. Res. 2019, 28, 34–56. [Google Scholar] [CrossRef]
- Phillips, S.J.; Anderson, R.P.; Schapire, R.E. Maximum entropy modeling of species geographic distributions. Ecol. Model. 2006, 190, 231–259. [Google Scholar] [CrossRef] [Green Version]
- Hernandez, P.A.; Graham, C.H.; Master, L.L.; Albert, D.L. The effect of sample size and species characteristics on performance of different species distribution modeling methods. Ecography 2006, 29, 773–785. [Google Scholar] [CrossRef]
- Zhang, K.; Zhang, Y.; Tao, J. Predicting the potential distribution of Paeonia veitchii (Paeoniaceae) in China by incorporating climate change into a Maxent model. Forests 2019, 10, 190. [Google Scholar] [CrossRef] [Green Version]
- Liu, C.; White, M.; Newell, G. Selecting thresholds for the prediction of species occurrence with presence-only data. J. Biogeogr. 2013, 40, 778–789. [Google Scholar] [CrossRef]
- Newbold, T.; Gilbert, F.; Zalat, S.; El-Gabbas, A.; Reader, T. Climate-based models of spatial patterns of species richness in Egypt’s butterfly and mammal fauna. J. Biogeogr. 2009, 36, 2085–2095. [Google Scholar] [CrossRef]
- Dormann, C.F.; Elith, J.; Bacher, S.; Buchmann, C.; Carl, G.; Carré, G.; Marquéz, J.R.G.; Gruber, B.; Lafourcade, B.; Leitão, P.J.; et al. Collinearity: A review of methods to deal with it and a simulation study evaluating their performance. Ecography 2013, 36, 27–46. [Google Scholar] [CrossRef]
- Li, G.; Xu, G.; Guo, K.; Du, S. Mapping the global potential geographical distribution of black locust (Robinia pseudoacacia L.) using herbarium data and a maximum entropy model. Forests 2014, 5, 2773–2792. [Google Scholar] [CrossRef] [Green Version]
- Zhang, K.L.; Yao, L.J.; Meng, J.S.; Tao, J. Maxent modeling for predicting the potential geographical distribution of two peony species under climate change. Sci. Total Environ. 2018, 634, 1326–1334. [Google Scholar] [CrossRef]
- Poorter, H.; Nagel, O. The role of biomass allocation in the growth response of plants to different levels of light, CO2, nutrients and water: A quantitative review. Aust. J. Plant. Physiol. 2000, 27, 595–607. [Google Scholar]
- Wang, X.L.; Wang, T.; Han, H.Z. Advances in biological characteristics and propagation of Zelkova serrata. Anhui Agr. Sci. Bull. 2017, 23, 122–123. [Google Scholar]
- Zhang, C.H. The Research of Breaking Dormancy and Germination Mechanism of Zelkova serrata Seeds. Master’s Thesis, Shandong Agricultural University, Taian, China, 2018. [Google Scholar]
- Fan, F.F.; Yuan, W.G.; Li, T.T.; Wu, C.P.; Zhang, J.; Jiao, J.J. Effect of water logging and drainage on growth and physiological properties of Zelkova serrata. J. Zhejiang For. Sci. Technol. 2018, 38, 62–68. [Google Scholar]
- Zeng, Y. The Physiological Response of Three Zelkova serrata Varieties to Water Stress. Master’s Thesis, Central South University of Forestry & Technology, Changsha, Hunan, China, 2018. [Google Scholar]
- Li, Y.F.; Zhu, X.J.; Chen, A.S.; Chen, D.P.; Chen, P. The research on introduction and high efficiency cultivation technology of Zelkova serrata. Modern Agr. Sci. Tech. 2014, 20, 142–160. [Google Scholar]
- Deb, J.C.; Phinn, S.; Butt, N.; McAlpin, C.A. Climatic-induced shifts in the distribution of teak (Tectona grandis) in tropical Asia: Implications for forest management and planning. Environ. Manag. 2017, 60, 422–435. [Google Scholar] [CrossRef] [PubMed]
- Zhang, K.; Liu, H.; Pan, H.; Shi, W.; Zhao, Y.; Li, S.; Liu, J.; Tao, J. Shifts in potential geographical distribution of Pterocarya stenoptera under climate change scenarios in China. Ecol. Evol. 2020, 10, 4828–4837. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, K.; Zhang, Y.; Zhou, C.; Meng, J.S.; Sun, J.; Zhou, T.H.; Tao, J. Impact of climate factors on future distributions of Paeonia ostii across China estimated by MaxEnt. Ecol. Inform. 2019, 50, 62–67. [Google Scholar] [CrossRef]
- Thomas, C.D.; Cameron, A.; Green, R.E.; Bakkenes, M.; Beaumont, L.J.; Collingham, Y.C.; Erasmus, B.F.; de Siqueira, M.F.; Grainger, A.; Hannah, L.; et al. Extinction risk from climate change. Nature 2004, 427, 145–148. [Google Scholar] [CrossRef]
- Zhang, K.; Zhang, Y.; Jia, D.; Tao, J. Species distribution modeling of Sassafras tzumu and implications for forest management. Sustainability 2020, 12, 4132. [Google Scholar] [CrossRef]
- Hällfors, M.; Aikio, S.; Fronzek, S.; Hellmann, J.; Ryttäri, T.; Heikkinen, R. Assessing the need and potential of assisted migration using species distribution models. Biol. Conserv. 2016, 196, 60–68. [Google Scholar] [CrossRef] [Green Version]
- Liu, Q.; Wang, Y.K.; Peng, P.H.; Lu, Y.F.; Chen, Y.F.; Wang, S. Characteristics of distribution and migration of species in Sichuan under the climate change. Mount. Res. 2016, 34, 716–723. [Google Scholar]
- Yang, L.; Yang, L.; Li, J.X.; Zhang, C.; Huo, Z.M.; Luan, X.F. Potential distribution and conservation priority areas of five species in Northeast China. Acta Ecol. Sin. 2019, 39, 1082–1094. [Google Scholar]
- Wiens, J.A.; Stralberg, D.; Jongsomjit, D.; Howell, C.A.; Snyder, M.A. Niches, models, and climate change: Assessing the assumptions and uncertainties. Proc. Natl. Acad. Sci. USA 2009, 106, 19729–19736. [Google Scholar] [CrossRef] [Green Version]
Figure 1.
The ROC curve predicting the distribution of Zelkova serrata based on the Maxent algorithm.
Figure 1.
The ROC curve predicting the distribution of Zelkova serrata based on the Maxent algorithm.
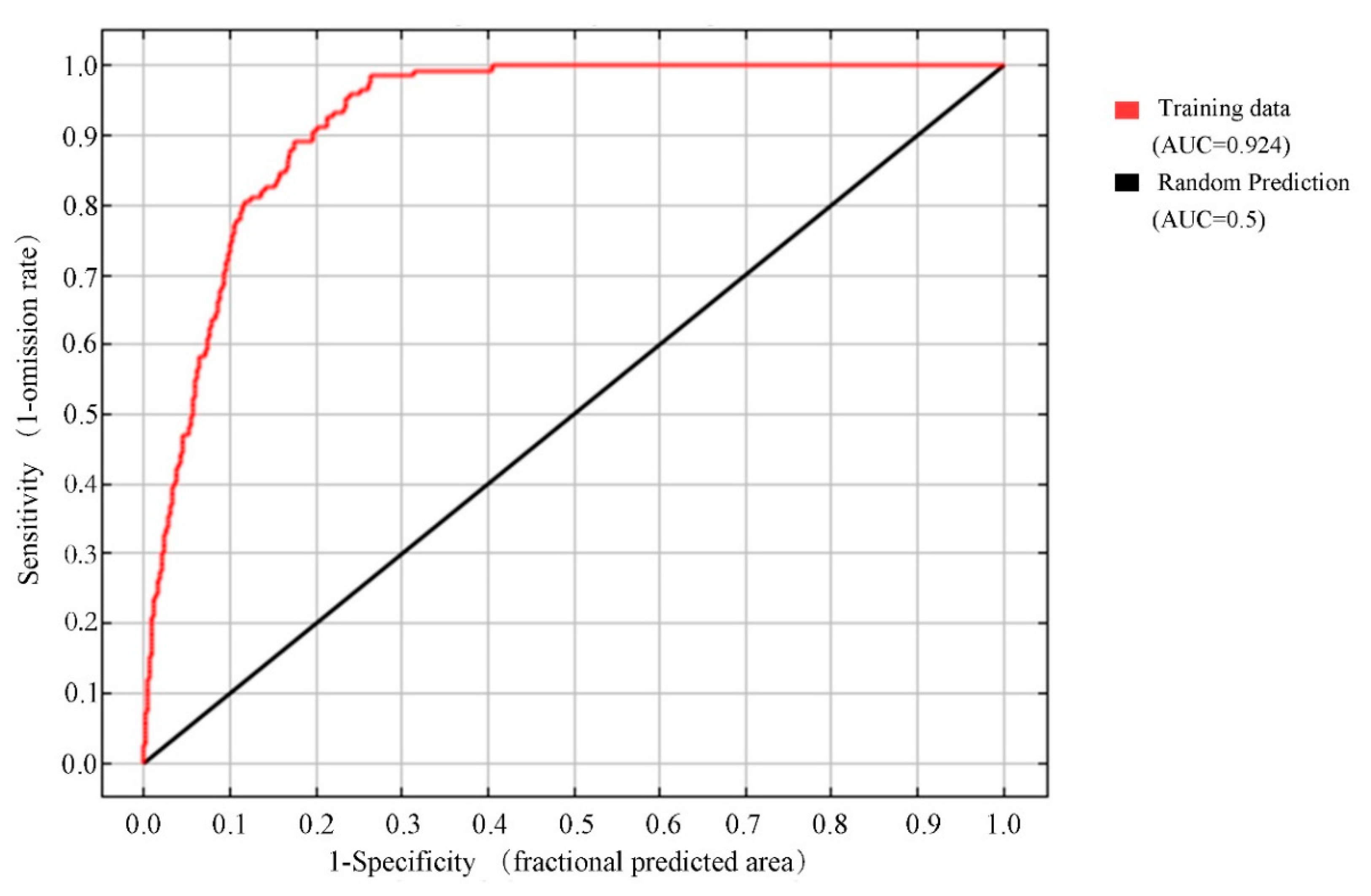
Figure 2.
Potential suitable distribution of Zelkova serrata (A) and the predicted shifts in core distributional (B) using the Maxent algorithm.
Figure 2.
Potential suitable distribution of Zelkova serrata (A) and the predicted shifts in core distributional (B) using the Maxent algorithm.
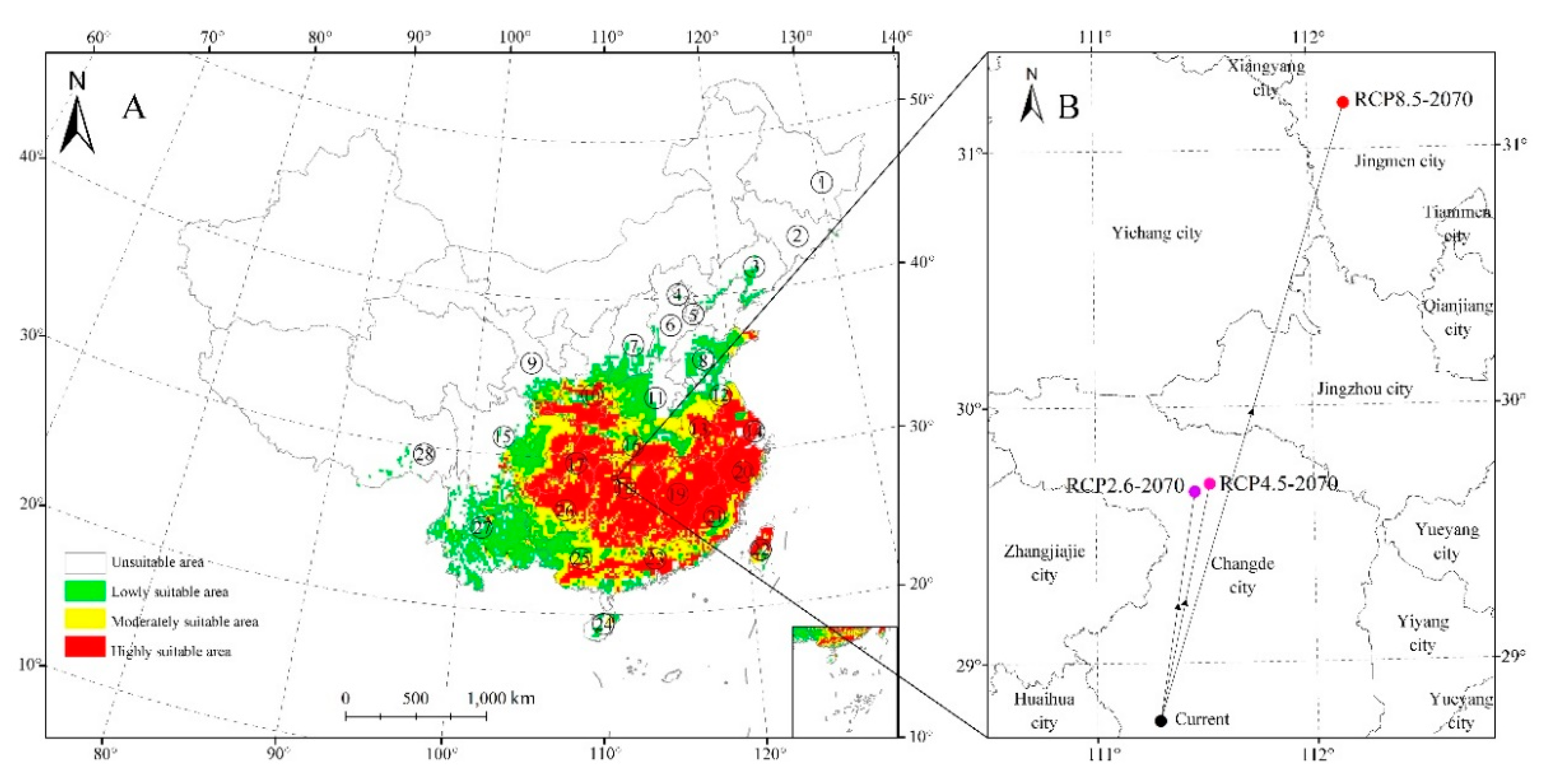
Figure 3.
Response curves for (A) annual precipitation (Bio 12), (B) annual mean temperature (Bio 1), (C) warmest quarter precipitation (Bio 18), and (D) temperature seasonality (Bio 4) in the species distribution model for Zelkova serrata based on the Maxent algorithm.
Figure 3.
Response curves for (A) annual precipitation (Bio 12), (B) annual mean temperature (Bio 1), (C) warmest quarter precipitation (Bio 18), and (D) temperature seasonality (Bio 4) in the species distribution model for Zelkova serrata based on the Maxent algorithm.
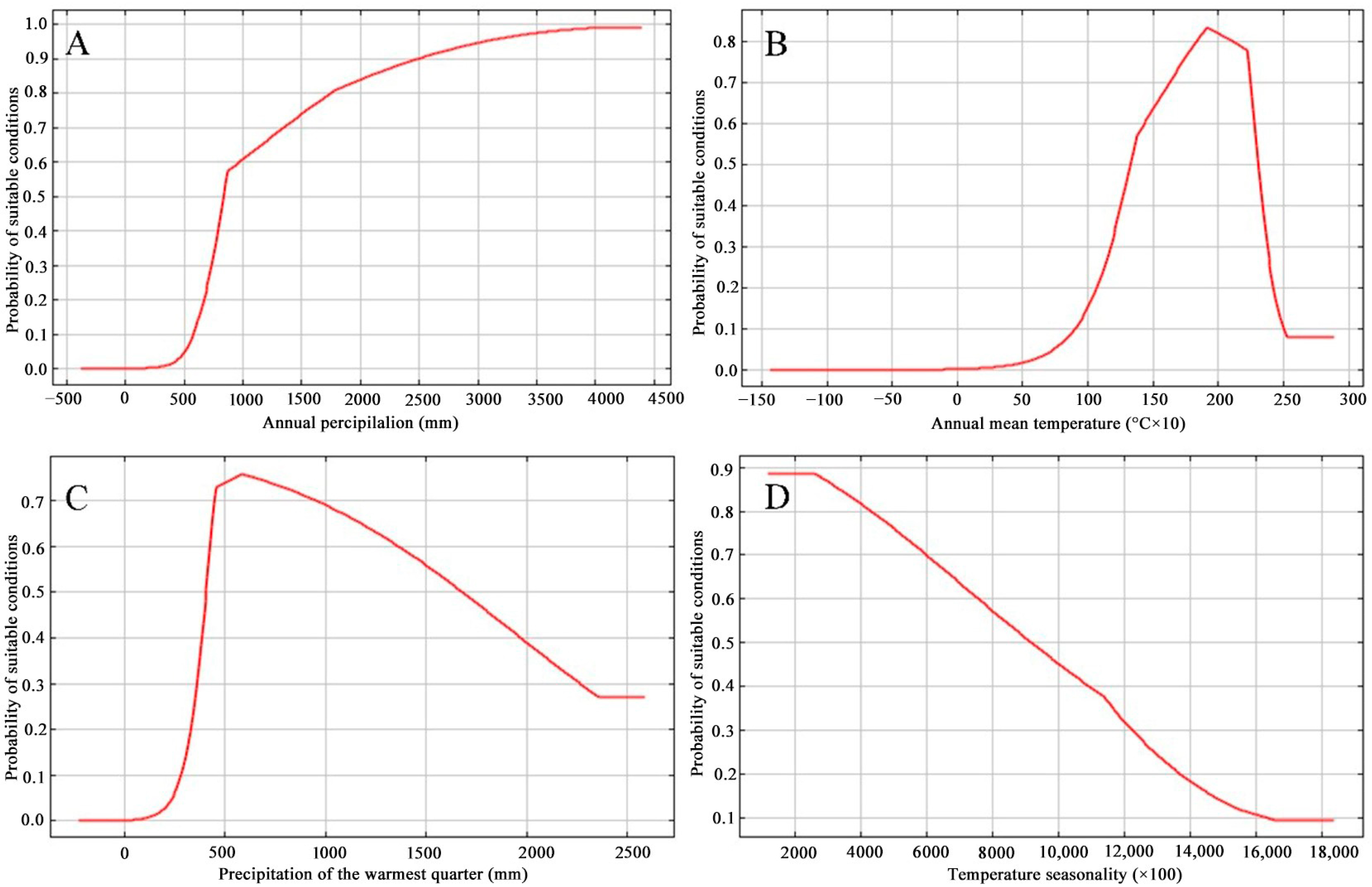
Figure 4.
Potential suitable distribution of Zelkova serrata predicted by Maxent. (A1–A3) are the maps of the suitable areas in 2070 in different scenarios of RCP 2.6, RCP 4.5, and RCP 8.5, separately; (B1–B3) are changes in the distribution areas in 2070 in the scenarios of RCP 2.6, RCP 4.5, and RCP 8.5, separately.
Figure 4.
Potential suitable distribution of Zelkova serrata predicted by Maxent. (A1–A3) are the maps of the suitable areas in 2070 in different scenarios of RCP 2.6, RCP 4.5, and RCP 8.5, separately; (B1–B3) are changes in the distribution areas in 2070 in the scenarios of RCP 2.6, RCP 4.5, and RCP 8.5, separately.

{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
Environment variables used for predicting the potential distribution of Zelkova serrata.
| Code | Environment Variables | Unit | Permutation Importance |
|---|---|---|---|
| Bio 12 | Annual precipitation | mm | 44.9 |
| Bio 1 | Annual mean temperature | °C × 10 | 17 |
| Bio 18 | Precipitation of the warmest quarter | mm | 8.1 |
| Bio 4 | Temperature seasonality | ×100 | 6 |
| Vap | Vapor pressure | kPa | 4.8 |
| Wet | Wet-day frequency | 4.1 | |
| Soilph | Soil pH | ×10 | 3.3 |
| Alt | Altitude | m | 3.3 |
| Gdd | Growing degree days | day | 1.9 |
| Bio 8 | Mean temperature of the wettest quarter | °C × 10 | 1.7 |
| Bio 14 | Precipitation of the driest quarter | mm | 1.4 |
| Bio 10 | Mean temperature of the warmest quarter | °C × 10 | 1.1 |
| Frs | Ground-frost frequency | 0.9 | |
| UVB 4 | Mean UV-B of lowest month | J m−2·day−1 | 0.4 |
| UVB 3 | Mean UV-B of highest month | J m−2·day−1 | 0.3 |
| Bio 15 | Precipitation seasonality | % | 0.2 |
| Bio 3 | Isothermality | ×100 | 0.2 |
| UVB 2 | UV-b seasonality | J m−2·day−1 | 0.2 |
| Bio 2 | Mean diurnal range | °C × 10 | 0.1 |
| UVB 1 | Annual mean UV-b | J m−2·day−1 | 0.1 |
| Soilorc | Soil organic carbon | g/kg | 0.0 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Cao, C.; Tao, J. Predicting the Areas of Suitable Distribution for Zelkova serrata in China under Climate Change. Sustainability 2021, 13, 1493. https://0-doi-org.brum.beds.ac.uk/10.3390/su13031493
AMA Style
Cao C, Tao J. Predicting the Areas of Suitable Distribution for Zelkova serrata in China under Climate Change. Sustainability. 2021; 13(3):1493. https://0-doi-org.brum.beds.ac.uk/10.3390/su13031493
Chicago/Turabian StyleCao, Chunyan, and Jun Tao. 2021. "Predicting the Areas of Suitable Distribution for Zelkova serrata in China under Climate Change" Sustainability 13, no. 3: 1493. https://0-doi-org.brum.beds.ac.uk/10.3390/su13031493
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.