
Effects of Diet Selection by Goats Grazing on the Vegetation of a Protected Pine Forest in Doñana Natural Park (SW Spain)

Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Area
2.2. Goat Grazing
2.3. Species Abundance
2.4. Data Analysis
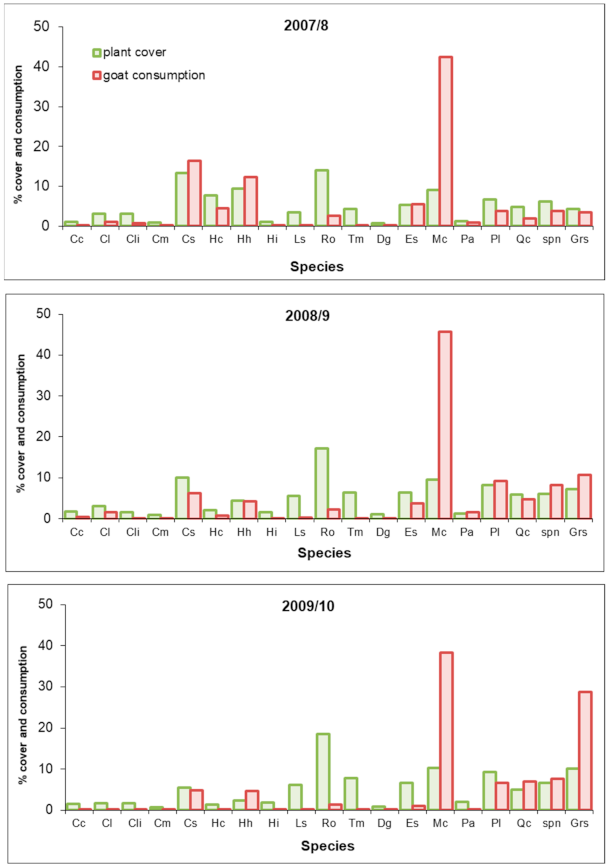
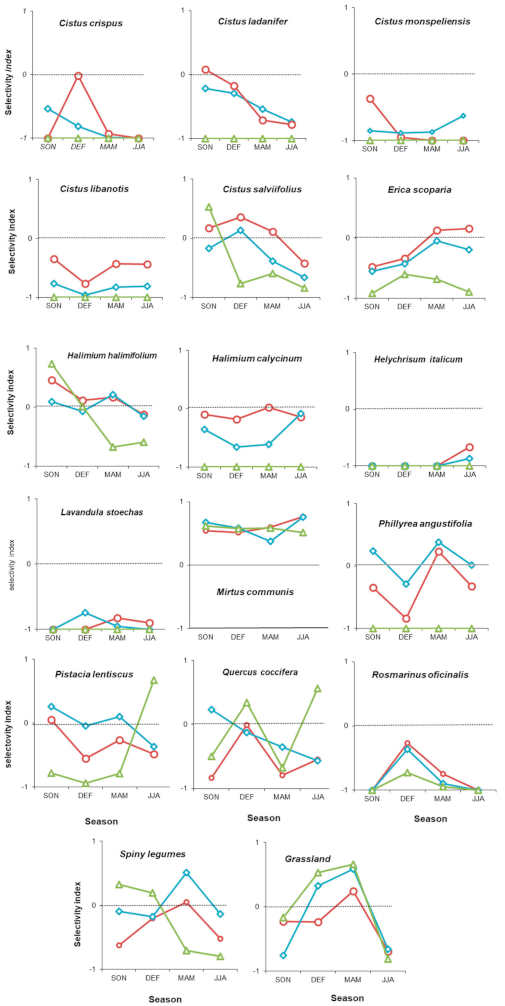
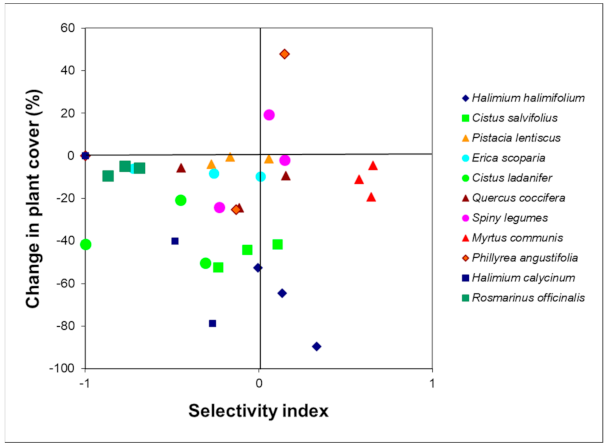
3. Results
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Food and Agriculture Organization of the United Nations (FAOSTAT). Statistics Database, Crop Statistics. Available online: http://www.fao.org/faostat/en/#data/QC (accessed on 4 March 2021).
- Southgate, E.W.R. People and the Land Through Time. Linking Ecology and History; Yale University Press: New Haven, CT, USA, 2019. [Google Scholar]
- Naveh, Z.; Lieberman, A.S. Landscape Ecology: Theory and Application; Springer Science and Business Media: New York, NY, USA, 2013. [Google Scholar]
- Harper, J.L. Population Biology of Plants; Academic Press: London, UK, 1977. [Google Scholar]
- Baraza, E.; Hódar, J.A.; Zamora, R. Consequences of plant–chemical diversity for domestic goat food preference in Mediterranean forests. Acta Oecol. 2009, 35, 117–127. [Google Scholar] [CrossRef]
- Morales-Jerrett, E.; Mancilla-Leytón, J.M.; Delgado-Pertíñez, M.; Mena, Y. The contribution of traditional meat goat farming systems to human wellbeing and its importance for the sustainability of this livestock subsector. Sustainability 2020, 12, 1181. [Google Scholar] [CrossRef] [Green Version]
- Pareja, J.; Baraza, E.; Ibáñez, M.; Domenech, O.; Bartolomé, J. The role of feral goats in maintaining firebreaks by using attractants. Sustainability 2020, 12, 7144. [Google Scholar] [CrossRef]
- Mancilla-Leytón, J.M.; Hernando, C.; Cambrollé, J.; Muñoz-Vallés, S.; Pino-Mejías, R.; MartínVicente, A. Can shrub flammability be affected by goat grazing? Flammability parameters of Mediterranean shrub species under grazing. Sustainability 2021, 13, 1555. [Google Scholar] [CrossRef]
- Mancilla-Leytón, J.M.; Parejo-Farnés, C.; Martín Vicente, A. Selection of browse species and energy balance of goats grazing on forest understory vegetation in Doñana Natural Park (SW Spain). Livest. Sci. 2012, 148, 237–242. [Google Scholar] [CrossRef]
- Kilgour, R.J. In pursuit of “normal”: A review of the behaviour of cattle at pasture. Appl. Anim. Behav. Sci. 2012, 138, 1–11. [Google Scholar] [CrossRef]
- Jáuregui, B.; Celaya, R.; García, U.; Osoro, K. Vegetation dynamics in burnt heather-gorse shrublands under different grazing management with sheep and goats. Agrofor. Syst. 2007, 70, 103–111. [Google Scholar] [CrossRef]
- Meuret, M.; Bartiaux-Thill, N.; Bourbouze, A. Evaluation de la consommation d’un troupeau de chèvres laitières sur parcours forestier—Méthode d’observation directe des coups de dents—Méthode du marqueur oxyde de chrome. Ann. Zootech. 1985, 34, 159–180. [Google Scholar] [CrossRef]
- Mancilla-Leytón, J.M.; Pino Mejías, R.; Martín Vicente, A. Do goats preserve the forest? Evaluating the effects of grazing goats on combustible Mediterranean scrub. Appl. Veg. Sci. 2013, 16, 63–73. [Google Scholar] [CrossRef]
- Daget, P.; Poissonet, J. A method of plant analysis of pastures. Ann. Agron. 1971, 22, 5–41. [Google Scholar]
- Valdés, B.; Talavera, S.; Fernández-Galiano, E. Flora Vascular de Andalucía Occidental; Ketres Editora: Barcelona, Spain, 1987. [Google Scholar]
- Haberman, S.J. The analysis of residuals in cross-classified tables. Biometrics 1973, 29, 205–220. [Google Scholar] [CrossRef]
- Ivlev, V.S. Experimental Ecology of the Feeding of Fishes; Yale University Press: New Haven, CT, USA, 1975. [Google Scholar]
- Papachristou, T.G.; Dziba, L.E.; Provenza, F.D. Foraging ecology of goats and sheep on wooded rangelands. Small Rumin. Res. 2005, 59, 141–156. [Google Scholar] [CrossRef]
- Goetsch, A.L.; Gipson, T.A.; Askar, A.R.; Puchala, R. Invited review: Feeding behavior of goats. J. Anim. Sci. 2010, 88, 361–373. [Google Scholar] [CrossRef] [PubMed]
- Mancilla-Leytón, J.M.; Joffre, R.; Martín Vicente, A. Effect of grazing and season on the chemical composition of Mediterranean shrub species in Doñana Natural Park, Spain. J. Arid Environ. 2014, 108, 10–18. [Google Scholar] [CrossRef]
- Rogosic, J.; Moe, S.R.; Skobic, D.; Knezovic, Z.; Rozic, I.; Zivkovic, M.; Pavlicevic, J. Effect of supplementation with barley and activated charcoal on intake of biochemically diverse Mediterranean shrubs. Small Rumin. Res. 2009, 81, 79–84. [Google Scholar] [CrossRef]
- Vasta, V.; Ventura, V.; Luciano, G.; Andronico, V.; Pagano, R.I.; Scerra, M.; Priolo, A. The volatile compounds in lamb fat are affected by the time of grazing. Meat Sci. 2012, 90, 451–456. [Google Scholar] [CrossRef] [PubMed]
- Benítez, G.; González-Tejero, M.R.; Molero-Mesa, J. Knowledge of ethnoveterinary medicine in the Province of Granada, Andalusia, Spain. J. Ethnopharm. 2012, 139, 429–439. [Google Scholar] [CrossRef] [PubMed]
- Mithöfer, A.; Boland, W. Plant defense against herbivores: Chemical aspects. Ann. Rev. Plant Biol. 2012, 63, 431–450. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mancilla-Leyton, J.M.; Sánchez-Lineros, V.; Martín Vicente, A. Influence of grazing on the decomposition of Pinus pinea L. needles in a silvopastoral system in Doñana, Spain. Plant Soil 2013, 373, 173–181. [Google Scholar] [CrossRef]
- Emanuelsson, U. The Rural Landscapes of Europe: How Man Has Shaped European Nature; Swedish Research Council Formas: Värnamo, Sweden, 2009. [Google Scholar]




{kind=link}
{kind=link}
{kind=link}
| Species | Initial Cover | Total Consumption | Change in Abundance from 2007 to 2010 (%) | |
|---|---|---|---|---|
| % | % | Ungrazed | Grazed | |
| Cistus crispus (Cc) | 1.1 | 0.23 | 1.10 | −29.2 |
| Cistus ladanifer (Cl) | 3.0 | 1.19 | 45.6 | −72.2 |
| Cistus libanotis (Cli) | 3.2 | 0.37 | −5.90 | −69.2 |
| Cistus monspeliensis (Cm) | 0.9 | 0.07 | 0.60 | −62.9 |
| Cistus salviifolius (Cs) | 13.3 | 10.14 | −65.5 | −80.6 |
| Halimium calycinum (Hc) | 7.7 | 2.14 | −60.8 | −87.4 |
| Halimium halimifolium (Hh) | 9.4 | 7.61 | −26.9 | −91.1 |
| Helichrysum italicum (Hi) | 1.1 | 0.04 | 161.1 | 19.2 |
| Lavandula stoechas (Ls) | 3.5 | 0.19 | 24.0 | 17.1 |
| Rosmarinus officinalis (Ro) | 14.0 | 2.23 | 66.8 | −18.0 |
| Thymus mastichina (Tm) | 4.3 | 0.00 | 14.9 | 26.6 |
| Daphne gnidium (Dg) | 0.7 | 0.00 | 27.8 | −29.3 |
| Erica scoparia (Es) | 5.4 | 4.04 | 25.7 | −21.8 |
| Myrtus communis (Mc) | 9.1 | 43.30 | 30.8 | −30.8 |
| Phillyrea angustifolia (Pa) | 1.2 | 1.10 | 39.7 | 10.4 |
| Pistacia lentiscus (Pl) | 6.6 | 6.63 | 11.7 | −5.90 |
| Quercus coccifera (Qc) | 4.8 | 3.88 | 17.4 | −34.9 |
| Spiny legumes (spn) | 6.1 | 6.33 | 20.0 | −8.10 |
| Grassland (Grs) | 4.3 | 10.5 | 13.3 | 82.0 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mancilla-Leytón, J.M.; Fernández-Alés, R.; Martín Vicente, Á. Effects of Diet Selection by Goats Grazing on the Vegetation of a Protected Pine Forest in Doñana Natural Park (SW Spain). Sustainability 2021, 13, 3950. https://0-doi-org.brum.beds.ac.uk/10.3390/su13073950
Mancilla-Leytón JM, Fernández-Alés R, Martín Vicente Á. Effects of Diet Selection by Goats Grazing on the Vegetation of a Protected Pine Forest in Doñana Natural Park (SW Spain). Sustainability. 2021; 13(7):3950. https://0-doi-org.brum.beds.ac.uk/10.3390/su13073950
Chicago/Turabian StyleMancilla-Leytón, Juan Manuel, Rocío Fernández-Alés, and Ángel Martín Vicente. 2021. "Effects of Diet Selection by Goats Grazing on the Vegetation of a Protected Pine Forest in Doñana Natural Park (SW Spain)" Sustainability 13, no. 7: 3950. https://0-doi-org.brum.beds.ac.uk/10.3390/su13073950