
Biological Concepts as a Source of Inspiration for Efficiency, Consistency, and Sufficiency



1
Cluster of Excellence livMatS @ FIT—Freiburg Center for Interactive Materials and Bioinspired Technologies, University of Freiburg, 79110 Freiburg, Germany
2
Plant Biomechanics Group @ Botanic Garden Freiburg, University of Freiburg, 79104 Freiburg, Germany
3
Oeko-Institut, Institute for Applied Ecology, 79100 Freiburg, Germany
*
Author to whom correspondence should be addressed.
†
These authors contributed equally to this work.
Sustainability 2022, 14(14), 8892; https://0-doi-org.brum.beds.ac.uk/10.3390/su14148892
Submission received: 18 January 2022
/
Revised: 12 July 2022
/
Accepted: 15 July 2022
/
Published: 20 July 2022
(This article belongs to the Special Issue Nature-Inspired Sustainable Development)
Abstract
:Sustainable development is a global challenge addressed by the 2030 Agenda with internationally adopted goals. The consideration of the three major sustainability strategies of efficiency, consistency, and sufficiency can guide us toward more sustainable policy approaches, product manufacturing, service offers, and consumption lifestyles. We select the growth form “liana”, which has evolved several times independently, to identify traits of lianas and general biological concepts derived therefrom. Even though sustainability is an anthropocentric approach and does not exist in biology, we can attribute biological concepts to sustainability strategies. The biological concepts of lightweight construction, modularity, function-related tissue formation, and trade-off can be attributed to efficiency; the concepts of zero waste, best fit, and damage repair to consistency; and the change of growth form and the concept of less is more and good enough to sufficiency. We discuss the analogies between parasitic architecture and the “structural parasitism” of lianas on host trees and between polymers with switchable autonomous properties and ontogenetic changes in the lianescent growth form. Efficiency can be analyzed quantitatively and consistency qualitatively, whereas sufficiency, as an aspect of human consumption patterns, cannot be mathematically measured. Biological concepts can thus serve as a source of inspiration for improving sustainability in the technosphere.
1. Introduction
The understanding of sustainability has repeatedly changed over the last 300 years [1]. Its inflationary use in politics, science, and society in the last few decades has rendered it increasingly arbitrary, and it often blurs the view of its core meaning. The term “sustainability” was originally introduced more than 300 years ago by chief miner Hans-Carl von Carlowitz with regard to economic motives during a serious timber shortage [2]. Sustainability as a mission statement for the technosphere was taken up in the debate about the scarcity of natural resources in the 1970s [3] and experienced a renaissance through its further development in terms of content. An important milestone in the framing of sustainability as a statement with normative relevance was achieved in the World Commission on Environment and Development. Its report, the so-called Brundtland Report, describes sustainable development as a necessary transformation process of the economy and society toward a sustainable development, with particular emphasis on innovations in the technosphere that respect the link between humans and nature [4]. Finally, the adoption of the 2030 Agenda by the member states of the United Nations in the year 2015 led to an internationally agreed set of 17 Sustainability Development Goals (= SDGs) and 169 targets [5]. These goals can be considered as the interdisciplinary normative basis of sustainability research, covering ecological, economic, and social aspects.
Beyond the goals and targets of the 2030 Agenda, three major strategies have emerged from the aforementioned mission statements and milestones for targeted action toward a sustainable development. Efficiency, consistency, and sufficiency offer a range of possibilities and guidance that help us achieve more sustainable policy approaches, product and service manufacturing, and consumption lifestyles. Efficiency raises the question: “How can we reduce the use of resources and associated environmental impacts at the lowest possible input?”. The strategy can be applied not only to the manufacturing process and the usage of products and services but also to policy strategies (e.g., funding schemes for CO2 reduction). Following the approach “doing more with less”, efficiency strategies primarily aim to optimize manufacturing and/or the use of products and services in order to reduce resource use and environmental impact throughout their entire life cycle. Consistency involves the question: “How do we use resources while reducing the associated environmental impacts?” and is aimed at various technological or product-related solutions that are embedded in nature [6]. Hence, the ultimate objective of consistency is not less resource utilization but rather the integration of anthropogenic material and production systems and the related material and energy flows into natural cycles [7]. Consistency as a sustainability strategy has been further cemented by the development of product life cycle analysis [8] and product sustainability analysis [9,10] and by the concepts of industrial ecology [11] and of cradle-to-cradle [12] and circular economy [13]. Sufficiency deals with the question: “How much do we need and what is superfluous?”, with the primary goal of reducing resource use and environmental impacts through changes in political framework conditions and consumption patterns [14]. The discourse on sufficiency is often associated with the discourse on the needs for a “good life” [15], renunciation, and the correct level of consumption [16]. According to Paech [17], sufficiency can be perceived more as liberation rather than renunciation because by decluttering consumption patterns, one can turn to the remaining consumption activities intensively and with pleasure. Moreover, sufficiency embraces aspects of efficiency and consistency.
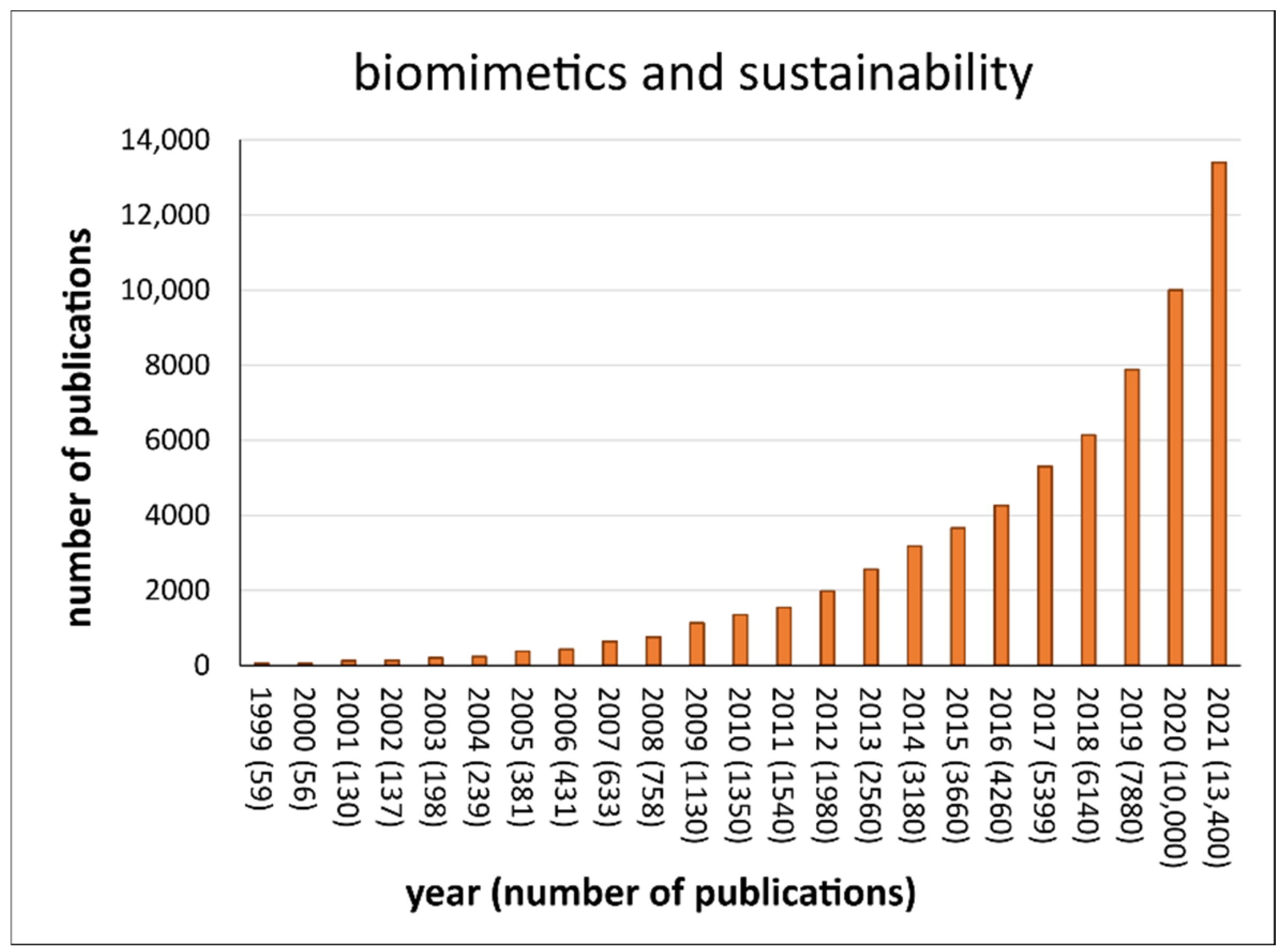
Interestingly, the proclamation of the Anthropocene [18] occurred simultaneously with considerations that biomimetic products ought to contribute to a more sustainable future. Since 2000, there has been an exponential increase in publications dealing with “sustainability and Anthropocene” and ”biomimetics and Anthropocene” [19]. Figure 1 shows publications that study the relationship of “biomimetics and sustainability” also increasing exponentially from 2000 to 2021. However, the question arises as to whether this scientific interest is also a safeguard for targeted action in the direction of sustainability. For instance, policymakers need to translate scientific findings into legislative frameworks as well as entrepreneurs into products and services that create the basis for more sustainable consumption and lifestyle patterns among the population.
Generally, learning from nature is linked with the hope that we can thereby promote the discovery and development of sustainable solutions in the technosphere. The idea flow from living nature to technology seems to promise technical solutions with extraordinary qualities, namely ones that are low risk, evolutionary optimized, and ecologically adapted. Von Gleich et al. [20] have summarized this normative aspect under the term “biomimetic promise”. Without question, sustainable development is a human-made mission statement that puts humankind and intergenerational equity at the center and provides targeted guidance for achieving the goal of sustainability. Living nature, however, is the result of biological evolution, which is characterized by the evolutionary factors of mutation, recombination, and selection, and it is indisputably neither anthropocentric nor teleological. In general, learning from nature can take place on three levels that differ with respect to the biological model, the knowledge transferred, and the technical development (Table 1).
Since biological models and mechanisms and processes of evolution are not per se sustainable, biomimetic products, biomimetically optimized components, and bioinspired strategies cannot automatically be sustainable either [27]. However, engineers can ensure a contribution to more sustainability of their biomimetic developments if a special ethos and respectful interaction with nature complement their practice [28]. Even though sustainability is an anthropocentric approach and does not exist in biological models, we can attribute several biological concepts to the sustainability strategies of efficiency, consistency, and sufficiency.
Against this background, in this paper, we will focus on the example of the plant growth form “liana”, which has evolved independently several times during the course of biological evolution, and present biological concepts that can sharpen and develop our understanding of efficiency, consistency, and sufficiency. The aim of this work is not to write a comprehensive review on the subject of sustainability strategies and/or the growth form of plants but to find answers to the following scientific question: “Can the sustainability strategies of efficiency, consistency, and sufficiency be sharpened and further concretized through the attribution of biological concepts?“
2. Biological Concepts of the Growth Form of Lianas
In the following, we will introduce our selected plant model, the growth form “liana”. We decided against randomly searching for as many unlinked examples as possible from the animal and plant kingdom. Instead, we wish to show that the characteristics of lianas are in a form–function relationship, which, despite its complexity, only allows a limited range of biological concepts and attributions to sustainability strategies (Table 2).
In addition to the systematic classification of family, genus, and species, growth forms classify categories of plants that have the same growth habit but that are not necessarily related [29,30,31]. The growth forms of plants are, for example, trees, shrubs, lianas, semi-self-supporters, succulents, epiphytes, and herbs. In the following, we focus on woody plants that have the most interesting features for our study. In contrast to the typically smaller annual to perennial herbaceous plants, woody plants have a typically longer lifetime and larger plant body. Climbing plants differ drastically from self-supporting plants or semi-self-supporting plants in their mechanical performance, their underlying stem structure, and especially the changes of these features during ontogeny (= development of an individual organism).
The growth form liana is evolutionarily highly successful, which is reflected by the fact that climbing plants exist in at least 133 families of spermatophytes and that more than half of the families of vascular plants have climbing species that all originated from originally self-supporting ancestors (best fit concept) [32,33,34,35].
Lianas and their interplay with host trees represent prime examples for the complex mutual influence and interdependence of various plant species that are nevertheless viable independently of each other. Furthermore, lianas play an important role in highly complex and interconnected ecosystems, such as tropical rainforests. Because of the high temperature and humidity in the rainforest, microorganisms rapidly and completely decompose dead plant parts that have fallen to the ground (zero waste concept).
In contrast to self-supporting shrubs and trees, which merely increase in plant body size (i.e., basal stem diameter and stem height), lianas change their growth form during ontogeny (change of growth form concept). Young liana saplings and young terminal searcher stems of lianas are mechanically self-supporting until they find support. Figure 2a shows a young self-supporting sapling and Figure 2b an old lianescent ribbon-shaped stem of Bauhinia guianensis. In their establishment phase, lianas have to overcome three main challenges: they need (1) to find new support (typically by autonomous circumnutation movements), (2) to bridge the distance to a new (distant) support (by stiff searcher stems), and (3) to attach safely to the new support (by various attachment structures). For these partly conflicting requirements, the most apical stem parts of a liana, the searcher stems, have developed trade-offs in terms of being sufficiently stiff and lightweight to prevent them drooping under their own weight. However, they must also be flexible enough to enable circumnutation movements that cover a large space volume and that are essential for finding new supports (trade-off concept) [36,37,38,39].
With their thin stems, young liana saplings are just stiff enough not to bend. However, even a thin woolen thread attached to the stem tip as an additional weight leads to a drooping of the stem [30]. Young lianas form just enough material to provide them with a safety factor of slightly over 1.0 (good enough concept), i.e., they can carry their own weight and extremely small additional loads, such as light wind gusts or raindrops. In contrast, other herbaceous plant stems have safety factors of between 1.7 and 2.9, whereas large old record-size trees are mechanically “overbuilt” with safety factors of approximately 4.0 [40,41]. In many liana species, the searcher stem can intertwine and form braided systems (modularity concept). Figure 2c shows the intertwining stems of Aristolochia macrophylla. In these braids, several stems of the same specimen are intertwined around each other and provide mutual support. Braiding typically occurs only in limited segments and not over the entire length of the nearly parallel searcher shoots. The longest (typically oldest) searcher stem of the braid can thus achieve a considerable lateral reach and reliably maintain its posture and the apical flexibility necessary for circumnutation [42].
Older lianas, however, climb plants with mostly woody stems that are rooted in the ground and that climb up trees or other supporting structures (Figure 2b). Liana stems remain much smaller in diameter and volume compared with self-supporting trees of a comparable age. Figure 2d shows the difference in cross-sectional diameter between the coiled liana stem of Condylocarpon guianense and the trunk of its host tree. Compared with trees, lianas can grow over crowns of several supporting trees because they invest in enormous stem length (500 m and more have been recorded), which is much more than the maximal stem height of trees at approximately 130 m (Figure 2e) [35,38,43]. Furthermore, huge diameter vessels in the flexible wood allow for sufficient water transport and for mechanical stability and elasticity in the long thin stems (vessel diameter up to 800 µm) (multifunctionality concept) [30,37,38,44]. Thus, the non-self-supporting growth form enables climbing lianas in dense forests to reach the canopy and thus an advantageous position for pollination, fruit, or seed dispersal and for receiving direct sun light for photosynthesis but with little investment in their own stem material (concept of external support). The resource savings from the exploitation of external climbing support allows the liana to use its metabolic energy for other tasks. One of the most important of these tasks is the production of various attachment structures, such as hooks and tendrils with or without adhesive pads, adhesion roots with root hairs, and twining stems (Figure 2f) that allow climbing lianas to secure their contact with the supporting trees and that represent an inevitable consequence of the non-self-supporting growth form (change of function concept) [38,45,46,47].
The growth of the woody plant body of climbing plants is generally accompanied by a marked change in rigidity with increasing age. Flexural rigidity and torsional rigidity are determined by the size and the mechanical properties of the stems, branches, or twigs. In this context, the bending modulus and the torsional modulus are measures for resistance to bending or torsion deformation, ultimately resulting in a trade-off between being stiff enough to withstand drooping by gravity and flexible enough to be streamlined under wind loads and, in lianas, to follow the movement of the host trees (trade-off concept). With respect to material properties, the bending elastic moduli of old basal stem parts of self-supporting plants are typically up to 10 times higher than those of young flexible apical twigs. In semi-self-supporting plants, the bending elastic modulus remains mostly constant during ontogeny. In non-self-supporting climbing lianas, a significant decrease in the bending elastic modulus and torsional modulus by a factor of up to 15 occurs from the relatively stiff young apical searcher stems or saplings (Figure 2a) to the old basal stem part, which is highly flexible in bending and torsion (Figure 2b) [30,31,34,35,36,38,39]. What is the basis of the described changes of material properties that occur during the ontogeny of lianas? Plants can form various tissues with different material properties (concept of function-related tissue formation). Stiff young searcher stems are either stabilized by dense stiff wood (e.g., Condylocarpon guianense, Bauhinia guianensis) or by a closed peripheral ring of sclerenchyma fibers (e.g., Aristolochia marcophylla, see magenta arrows in Figure 2g, left, and Clematis vitalba). In contrast, old stems are flexible in bending and torsion caused by less dense flexible wood (concept of lightweight construction), huge wood rays, a thick flexible cortex, ribbon-like or indented stems (e.g., Bauhinia guianensis, Passiflora glandulifera), and/or a highly segmented (e.g., Aristolochia marcophylla, see yellow arrows in Figure 2g, right) or a star-shaped (e.g., Maripa scandens, Condylocarpon guianense) wood cylinder.
Figure 2g shows transverse sections of a one-year-old stem and of a two-year-old stem of Aristolochia marcophylla. The transverse section of the one-year-old stem reveals a peripheral closed ring of sclerenchyma (Figure 2g, left, see magenta arrows) that is responsible for the high flexural and torsional rigidity of the self-supporting young plant stems and searcher stems that cannot yet rely on support from a host tree. During secondary growth, following the secure attachment of the stem to a host tree (in the case of Aristolochia marcophylla by twining around host stems and branches), the newly formed flexible wood inside the closed ring increases in volume to such an extent that, in the two-year-old stem, the strengthening ring is ruptured (concept of less is more). The fissures of the ring are rapidly sealed by adjacent thin-walled sealing cells (concept of damage repair). In the subsequent healing phase, further thin-walled sealing cells are formed that can safely seal the fissures but that cannot restore the original mechanical property (good enough concept). Some of the thin-walled parenchyma cells develop into thick-walled strengthening cells (sclerenchyma), leading to a partial recovery of the original mechanical property (Figure 2g, see yellow arrows). Compared with the stems with a closed ring of lignified sclerenchyma cells, the stems with a segmented ring exhibit a more flexible behavior in bending and torsion. The described sealing principle provided the inspiration for a self-sealing foam coating for pneumatic systems in technology [48].
The high flexibility in bending and torsion of their older stems enables lianas to survive the breaking off of large branches to which they are attached and even the falling down of entire support trees. After such an event, the lianas slip down the main stem (Figure 2d) or may even be found coiled like a rope on the forest floor (Figure 2e) and are then able to colonize new support trees with the aid of stiff reiterative axes built on the old stems (Ref [30]; TS: field observations). On the other hand, climbing lianas rely on the existence of self-supporting host trees, which makes them dependent on the existence of stable forest ecosystems. This can be observed in disturbed areas, e.g., after natural tree breakage or logging, where lianas, because of their vigorous growth, may densely cover the area and can hinder and/or delay the emergence and growth of young trees that would provide new support (TS: field observations).
3. Analogies in the Technosphere
In this study, we compiled numerous evolutionary characteristics from which we derived various biological concepts (Table 2). These characteristics are of high selective advantage for the growth form “liana“. The particularly remarkable characteristics in this respect include the small investment in liana’s own stem material as part of a “structural parasitic” growth habit and the change of material properties, i.e., the transformation from stiff self-supporting searcher stem or sapling to flexible climbing plant stem. Our scientific motivation was to investigate whether these biological concepts can sharpen and develop our understanding of efficiency, consistency, and sufficiency. In particular, we will use these two extraordinary characteristics of liana as examples in order to investigate to what extent they can act as inspiration for the technosphere. Indeed, some rare examples can be found here that demonstrate the existence of comparable strategies in the technosphere: parasitic architecture and polymer systems with switchable autonomous properties.
3.1. Parasitic Architecture
Based on the definitions of biology, most lianas are not parasitic plants that harm their host by means of extracting water and nutrients from the host plant. However, with regard to the climbing support provided by the host, we can speak of a “structural parasitism” that can be used as a source of inspiration in the field of architecture.
Analogies with the interplay of lianas and their host trees can thus be found in the approach of parasitic architecture, which refers to a wide array of small-scale architectonic “additions” to existing larger buildings. Independent of being legal or illegal, temporary or permanent, all of these additions aim to expand volumes of their architectonic “hosts” and sometimes even further develop their functions [49,50]. An example of a cutting-edge application of parasitic architecture is the concept of tiny houses (i.e., small stand-alone homes that mimic a modern home on a smaller scale) as complete living units occupying the roofs of existing buildings but making use of the staircases and installation ducts of the latter [49]. With “solar rooftops” (Figure 3), a solar-plus-energy residential unit also aims to create additional high-quality living space on the top of existing buildings. Previously unused roof space is replaced by a pre-fabricated, one- or two-story lightweight wooden construction, with the roof cladding almost completely covered with photovoltaic modules. On an annual average, these photovoltaic modules produce more electricity than the rooftop residential unit’s own requirements. The energy surplus can be used by the residential units of the existing buildings below or fed into the public grid [51]. Essentially, with the latter characteristics, parasitic architecture harbors great potential to contribute to net-zero CO2 emission scenarios in urban settings [52].
3.2. Polymer Systems with Switchable Autonomous Properties
The transformation from stiff searcher stems to flexible old stems represents another characteristic feature of lianas that could be mimicked and further developed in the technosphere. The corresponding engineering analog would be polymer systems with switchable autonomous properties, as chosen in our second example. An interesting approach in this respect are hydrophilic porous polymer coatings (PPC) that are based on ethyl cellulose and that are capable of switching upon being wetted with water (Figure 4). If applied in exterior paint coatings, ethyl cellulose systems can switch in response to precipitation. In the dry state, PPCs are opaque and reflect sunlight; once exposed to rainwater, they turn transparent, and sunlight is transmitted. Such coatings can passively heat or cool buildings, depending on the season, in regions with dry summers and wet winters [53]. It is passive nature makes this functionality highly appealing in terms of sustainability, since cooling is achieved without electricity, refrigerants, or mechanical pumps. In this sense, ethyl cellulose-based PPCs can be considered as an autonomous (pre-) programed material system in which complex structural and mechanical changes occur interactively, triggered by an external environmental stimulus.
4. Concretization of Sustainability Strategies by Means of Biological Concepts
As a basis for the discussion of our answers to the scientific question “Can the sustainability strategies of efficiency, consistency, and sufficiency be sharpened and further concretized through the attribution of biological concepts?“, we now provide some preliminary remarks that are important in this interdisciplinary context in order to avoid any misunderstandings.
4.1. Preliminary Remarks
Nature and culture represent a dualistic word pair, with culture denoting everything anthropogenic as opposed to nature, which is not human made [54]. The natural sciences (e.g., physics, chemistry, biology) and technology (e.g., engineering, design) also represent a dualistic word pair. Unfortunately, nature and technology are often juxtaposed, but they obviously do not belong to the same category [27]. In the study presented here, we make a clear distinction between biological concepts and sustainability strategies. Biological concepts have developed during biological evolution, which is characterized by the evolutionary factors of mutation, recombination, and selection. Sustainability strategies, however, are guidelines for human action designed to achieve predetermined sustainability goals. In contrast to biological concepts, sustainability strategies are anthropocentric and teleological. Since the term function is used in both biology and technology, we will provide two brief explanations. In the life sciences, no experiments can be conducted to determine the function of a biological structure that has evolved during the course of evolution; rather, functional attributions are made, since organisms are not deliberately constructed machines whose components serve intended purposes or goals [55]. Therefore, in the biological context, function is understood in the sense of traits that have evolved to contribute to fitness [27]. In a technical context, however, function is understood as the performance characteristics of a product or service system that can be quantified in the form of a functional unit [8]. The function of a technical product system needs to be evaluated based on its benefits [56]. Finally, we will discuss the difference between goals and targets, two terms that play a major role in the 2030 Agenda, which includes 17 Sustainability Development Goals (SDGs) and 169 targets. Goals are general statements of what is to be achieved but without giving precise figures or setting deadlines for completion. By contrast, targets define specific and measurable results to be achieved within a given time period.
4.2. Efficiency
Originally, the definition of efficiency referred to economic performance and costs. Within the context of sustainability research, the definition focuses more on physical resources, such as raw materials and energy carriers. According to Behrendt et al. [57], efficiency means the creation of economic performance with the lowest possible use of materials and energy by improving the input–output ratio. Since the economic performance of a product or service system is defined by its function (as defined above), the following Equation (1) can be derived that is applicable to both technical products and biological structures.
Thus, efficiency can be compared quantitatively. Basically, the more functions that a technical product/service or biological structure has and the fewer resources that are needed (or associated impacts are generated) to produce or build it, the greater the efficiency. The calculation is rather simple when only one resource or one impact is considered. It becomes more complicated, however, when the total environmental impact needs to be assessed, since the individual impacts have to be weighed against each other, and aggregations are necessary [58]. According to the biological concepts, the input–output ratio [57] can be improved by reducing the input for a function in the sense of using the minimum quantity of resources and/or by increasing the output in terms of considering multiple, (partly) conflicting, and improved functions while minimizing resources.
Within this context, we should note that efficiency and effectiveness are often used synonymously or confused with each other. However, they are clearly defined: efficiency means doing things right, whereas effectiveness means doing the right things.
Furthermore, after the implementation of efficiency strategies, rebound effects can often be observed in practice. Rebound effects are secondary effects that occur as a result of an efficiency measure but that counteract the original objectives of the primary measure (i.e., energy or resource savings). Hence, rebound effects are triggered by the measure and reduce its associated and usually intended effect or, in the worst case, even overcompensate for it [59]. Energy-efficient LED bulbs are a striking example in this respect; since they consume less energy than conventional light bulbs, many users tend to switch on more bulbs or leave them switched on for longer periods of time.
Interestingly, we can observe an analogous effect in lianas. As mentioned above, resource saving by exploitation of the external climbing support allows the use of metabolic energy for other tasks of the lianas, such as the development of attachment structures for a secure adherence to the host tree or the increased formation of leaves in the canopy for photosynthesis. The load that climbing lianas apply to their host trees and the potential increase in crown size and density caused by the additional liana leaves increase the danger of mechanical failure (mostly by wind loads) of branches or the entire support tree. In these cases, the resource savings do not lead to energy efficiency but even result in increased consumption; this is called the “rebound effect” in the technosphere.
4.3. Consistency
Consistency stands for the compatibility of technological production processes with nature. Huber [6] claims that consistent material flows are those “which are operated largely fail-safe in their own closed technical cycle, or which correspond to the metabolic processes of the surrounding nature in such a way that they can be integrated into them relatively easily, even in large volumes.” (Ref. [6], p. 81; translated into English by the authors). Thus, when focusing on resource use and waste, consistency can be achieved by the establishment of closed-loop material cycles as postulated by the concept of a circular economy [13]. In contrast to a linear economy, in which each product eventually becomes waste (cradle-to-grave), the circular economy creates closed-loop material cycles with a constant reuse of the resources employed (cradle-to-cradle). The circular economy thus describes a socio-economic system that aims at using equipment, materials, and products as long as and as efficiently as possible in order to minimize waste. Waste can be avoided either by the use of fully biodegradable materials or by keeping the products in the cycle with strategies such as reuse, repair, refurbishment, remanufacture, repurpose, recycling, and recovery [60]. Nevertheless, we need to stress that waste minimization is not the exclusive goal of the consistency strategy; instead, resource utilization should also be improved.
Against this background, a quantitative formulation of consistency has to refer to the aspect of circularity (Equation (2)). In this sense, the ideal consistency in terms of optimized resource use and a minimum of waste generation is achieved if perfect circularity is created.
In the technosphere, however, circularity is currently more of a vision than an established approach; other than a few material flows (such as paper/cardboard, glass or aluminum), recycling rates are still comparatively low, especially with regard to material recycling [61]. Haas et al. [62] also report a low level of circularity when material flows are used as a proxy. They calculated that only 7% of all materials entering the global economy are in closed loops.
In the context of the biological concepts found in lianas, we learned that the growth form liana has evolved independently in at least 133 plant families, and despite all differences, each of them is well embedded in its respective ecosystem. Especially in the tropical rain forests, microorganisms completely and rapidly decompose dropped liana parts, following the zero-waste concept. However, under anaerobic decomposition, nature itself produces natural waste in the form of fossilized organic material below ground level. Fossil fuels, such as coal, natural gas, and crude oil, consist mostly of carbon, which reacts with oxygen to produce the greenhouse gas carbon dioxide (CO2) when burnt by humans for heating or transportation [63]. Nevertheless, waste reduction through damage prevention and damage management is widespread in plants. Plants can prevent damage through the formation of gradient transitions by means of geometric characteristics and biomechanical properties, or they can respond and acclimate structurally and mechanically to withstand higher loads without damage. Damage management of plants includes the sealing and healing of wounds (concept of damage repair) and the formation of abscission zones, where damage is controlled spatially and temporarily, such as during leaf fall [64]. The concept of function change is common in the plant kingdom. Several types of structures have evolved that allow the support and attachment of liana and that originate from various plant organs or tissues. Tendrils, for example, are specialized stems, leaves, or petioles; thorns are modified branches or stems; and prickles are extensions of the epidermis.
4.4. Sufficiency
Although the strategies of efficiency and consistency are centered on technological progress, sufficiency requires personal lifestyle changes that respect ecological limits and the ability to restrain superfluous consumption [61]. Fischer and Grießhammer (Ref [14], p. 10; translated into English by the authors) define sufficiency as “changes in consumption patterns that help remain within the Earth’s ecological carrying capacity, whereby the benefit aspects of consumption change”. Paech [17] defines sufficiency as a process of decluttering by means of discarding superfluous items. In practice, the sufficiency approach involves borrowing instead of possessing things, traveling by train instead of flying, repairing damaged products instead of throwing them away or buying new consumer goods, and eating local foods instead of exotic ones [17].
However, we cannot analyze the degree of sufficiency solely by the quantity of renounced items. Sufficiency has to be conceived of in a much more multifaceted manner than the individual renunciation of superfluous consumption. According to Heyen et al. [65], the practice of sufficiency cannot be left to the individual alone. Instead, the demanded changes in consumption patterns must be operationalized, i.e., encouraged and promoted by a targeted sufficiency policy and with a suitable framework. For example, one can appeal to people to fly less; a governmental fixed CO2 price of EUR 180 per ton of CO2 would, however, act as a more effective control, leading to less air traffic.
When we try to take into account the multifaceted framing of sufficiency, and particularly its political and social implications, we soon see that this strategy cannot be expressed by an all-embracing mathematical–scientific formula.
Biological concepts attributed to sufficiency are associated with the change of growth form concept from a self-supporting searcher stem or sapling to a non-self-supporting climbing liana. In this context, the less is more concept in terms of the growth-induced disintegration of the closed peripheral ring of sclerenchyma cells into individual parts and the good enough concept of filling the gaps of the strengthening ring with thin-walled parenchyma cells demonstrate that these biological concepts are not the expression of the further strengthening of the existing structures but, on the contrary, the abandonment of previous structures. Moreover, the good enough concept of the young searcher stems and saplings is especially interesting, since young plants invest as few resources as possible in this early establishment phase, which is limited in time (time to host contact) and space (distance to host).
4.5. Perception Shift of the Sustainability Strategies
In the framework of this interdisciplinary study between biology and sustainability science, we noted that the boundaries between the three sustainability strategies of efficiency, consistency, and sufficiency are blurred to some extent. For example, the consistency approach often includes efficiency aspects, a phenomenon that has not been adequately taken into account in the classic sustainability discussion so far.
Our attempts to translate the strategies of efficiency, consistency, and sufficiency into formulae in order to generate measurable figures yielded mixed success. Efficiency can be calculated quantitatively, and the resulting numbers can be compared with each other (Equation (1)). Consistency can be captured qualitatively by the level of circularity (Equation (2)). No universal formula can, however, be derived for sufficiency. The performance of a quantitative analysis, as can be undertaken in the cases of efficiency and (to a more limited extent) consistency, has the potential to transfer a general goal into a particular target that can be formulated and evaluated. Since sufficiency is a multifaceted strategy, covering, in particular, consumption patterns and corresponding political frameworks, a quantitative analysis based on a mathematical–scientific formula is not possible.
We were able to identify several biological concepts with the potential to sharpen and further concretize our understanding and management of sustainability strategies. All biological concepts are self-explanatory by their very naming, are further explained by using the liana characteristics described, and can thus be attributed to concepts in the field of sustainability. We attributed the biological concepts of lightweight construction, modularity, function-related material formation, and trade-off to efficiency; the concepts of zero waste, best fit, and damage repair to consistency; and the concepts of change of growth form, less is more, and good enough to sufficiency.
5. Conclusions and Outlook
We provided some answers to our scientific question as to whether we can sharpen and further develop the sustainability strategies of efficiency, consistency, and sufficiency through the attribution of biological concepts. Even though sustainability is an anthropocentric approach and does not exist in biology, we can indeed attribute biological concepts to the three sustainability strategies. However, biological concepts and sustainability strategies are not opposite sides of the same coin or two separate realms of reality. Within our study, we were able to identify biological concepts with the potential of deepening our understanding and sharpening the definition of the three sustainability strategies. In the case of efficiency and consistency, for example, we derived formulae that can be applied in principle in both biology and the technosphere. In the case of sufficiency, however, we were unable to develop a mathematical–scientific formula, since this strategy is based primarily on political and social science principles.
Analogous to the biological concepts, smooth transitions exist between the three strategies. Furthermore, the sustainability strategies are not equivalent to each other in terms of their definition or with respect to their quantitative and qualitative analyses. Efficiency and consistency refer to product manufacturing, whereas sufficiency refers to changes in political frameworks and consumption patterns. Nevertheless, human consumption habits are of great relevance for the realization of all sustainability strategies.
We also wish to present here an outlook on the development of sustainability strategies. Efficiency is certainly the best known sustainability strategy in the public perception. In this regard, the public has to rely on future advances in technology for further improvements in efficiency. The Achilles’ heel of efficiency is the rebound effect, of which some people are not even aware. Consistency aims at changing the “quality” of energy and material cycles toward compatibility with nature. This entails a technical and economic structural change with further consequences for legal, political, and cultural changes. The high transformational potential of the consistency strategy is counteracted by the long-lasting processes involved [6,57]. Sufficiency is not a matter of renunciations and prohibitions by politicians but of rethinking and of changing our habits. A compelling argument is that sufficiency measures do not require new, expensive, and time-consuming technologies. Indeed, everyone can immediately commence a sufficient lifestyle [17]. We were able to show that biological concepts can serve as a source of inspiration for enhancing the sustainability performance of specific products in the technosphere; parasitic architecture and switchable polymers are interesting but seldom used approaches in this context. In particular, parasitic architecture can unlock great potential in terms of net-zero initiatives. Thus, our biological concepts can be used to find further ways of dealing with the technosphere. Further studies of biological concepts and their potential applications with regard to the various materials, products, and sectors of the technosphere should certainly help promote sustainability.
Author Contributions
Conceptualization, all authors; methodology, all authors; writing—original draft preparation, O.S. and M.M.; writing—review and editing, all authors; visualization, O.S. and T.S.; supervision, R.G. and T.S.; project administration, R.G.; funding acquisition, R.G. All authors have read and agreed to the published version of the manuscript.
Funding
Funded by the Deutsche Forschungsgemeinschaft (DFG, German Research Foundation) under Germany’s Excellence Strategy—EXC-2193/1—390951807. The article processing charge was partly funded by the University of Freiburg (Germany) in the funding program Open Access Publishing.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Acknowledgments
We thank R. Theresa Jones for improving the English.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Grober, U. Sustainability: A Cultural History; Green Books: Cambridge, UK, 2012; ISBN 9780857840455. [Google Scholar]
- Von Carlowitz, H.C. Sylvicultura Oeconomica, Oder Haußwirthliche Nachricht und Naturmäßige Anweisung zur Wilden Baum-Zucht; Johann Friedrich Braun: Leipzig, Germany, 1713. [Google Scholar]
- Meadows, D.H.; Meadows, D.L.; Randers, J.; Behrens, W.W., III. The Limits to Growth: A Report for the Club of Rome’s Project on the Predicament of Mankind; Universe Books: New York, NY, USA, 1972; ISBN 0876631650. [Google Scholar]
- World Commission on Environment and Development. Our Common Future; Oxford University Press: Oxford, UK, 1987; ISBN1 019282080X. ISBN2 9780192820808. [Google Scholar]
- United Nations. Transforming Our World: The 2030 Agenda for Sustainable Development: A/RES/70/1. 2015. Available online: https://www.un.org/en/development/desa/population/migration/generalassembly/docs/globalcompact/A_RES_70_1_E.pdf (accessed on 7 February 2021).
- Huber, J. Industrielle Ökologie. Konsistenz, Effizienz und Suffizienz in zyklusanalytischer Betrachtung. In Global Change; Simonis, U.E., Ed.; Nomos: Baden-Baden, Germany, 2000; Available online: http://www.rla-texte.de/texte/16%20Lehre%20Regional%C3%B6konomie/Huber%20industrial%20umweltkonsistenz.pdf (accessed on 4 October 2021).
- Linz, M. Warum Suffizienz unentbehrlich ist. In Von Nichts Zu Viel-Suffizienz Gehört zur Zukunftsfähigkeit; Linz, M., Ed.; Wuppertal Papers: Wuppertal, Germany, 2002; Volume 125, pp. 7–14. Available online: https://epub.wupperinst.org/frontdoor/deliver/index/docId/1512/file/WP125.pdf (accessed on 4 October 2021).
- International Standard Organization. ISO 14040; Environmental Management–Life Cycle Assessment–Principles and Framework. Beuth: Berlin, Germany, 2006.
- Öko-Institut. Produktlinienanalyse–Bedürfnisse, Produkte Und Ihre Folgen; Kölner Volksblatt Verlag: Köln, Germany, 1987. [Google Scholar]
- Grießhammer, R.; Buchert, M.; Gensch, C.-O.; Hochfeld, C.; Manhart, A.; Reisch, L.; Rüdenauer, I. PROSA–Product Sustainability Assessment, Guideline; Öko-Institut: Freiburg, Germany, 2007. [Google Scholar]
- Frosch, R.A.; Gallopoulos, N.E. Strategies for Manufacturing. Sci. Am. 1989, 261, 144–152. Available online: http://0-www-jstor-org.brum.beds.ac.uk/stable/24987406 (accessed on 4 October 2021).
- Braungart, M.; McDonough, W. Cradle to Cradle: Remaking the Way We Make Things; North Point Press: New York, NY, USA, 2002. [Google Scholar]
- MacArthur, F.E. (Ed.) Towards the Circular Economy Vol. 1: An Economic and Business Rationale for An Accelerated Transition; Seacourt: Oxford, UK, 2013; Available online: https://ellenmacarthurfoundation.org/towards-the-circular-economy-vol-1-an-economic-and-business-rationale-for-an (accessed on 4 October 2021).
- Fischer, C.; Grießhammer, R. Mehr als nur weniger. Suffizienz: Begriff, Begründung und Potenziale. Öko-Institut Working Paper; Öko-Institut: Freiburg, Germany, 2013; p. 2. Available online: https://www.oeko.de/oekodoc/1836/2013-505-de.pdf (accessed on 4 October 2021).
- Schneidewind, U.; Zahrnt, A. Damit Gutes Leben Einfacher Wird. Perspektiven Einer Suffizienzpolitik; Oekom Verlag: München, Germany, 2013. [Google Scholar]
- Scherhorn, G. Die Logik der Suffizienz. In Von Nichts zu Viel—Suffizienz Gehört zur Zukunftsfähigkeit; Linz, M., Ed.; Wuppertal Papers: Wuppertal, Germany, 2002; Volume 125, pp. 15–26. Available online: https://epub.wupperinst.org/frontdoor/deliver/index/docId/1512/file/WP125.pdf (accessed on 4 October 2021).
- Paech, N. Lob der Reduktion. Polit. Okol. 2013, 135, 16–22. Available online: https://klimaherbst.de/2016/wp-content/uploads/2018/11/A_016_22_paech_18_11.pdf (accessed on 4 October 2021).
- Crutzen, P.J.; Stoermer, E.F. The “Anthropocene”. Glob. Change Newsl. 2000, 41, 17–18. [Google Scholar]
- Möller, M.; Höfele, P.; Kiesel, A.; Speck, O. Reactions of sciences to the Anthropocene. Highlighting inter-and transdisciplinary practices in biomimetics and sustainability research. Elem. Sci. Anthr. 2021, 9. [Google Scholar] [CrossRef]
- Von Gleich, A.; Pade, C.; Petschow, U.; Pissarskoi, E. Potentials and Trends in Biomimetics; Springer Science & Business Media: Berlin/Heidelberg, Germany, 2010; ISBN 978-3-642-44406-7. [Google Scholar]
- Neinhuis, C.; Barthlott, W. Characterization and distribution of water-repellent, self-cleaning plant surfaces. Ann. Bot 1997, 79, 667–677. [Google Scholar] [CrossRef] [Green Version]
- Mattheck, C.; Tesari, I. Design in nature. In Development & Application of Computer Techniques to Environmental Studies VII; Brebbia, C.A., Zannetti, P., Ibarra-Berasteg, G., Eds.; WIT Press: Southampton, UK, 2000; Volume 41, pp. 217–226. [Google Scholar]
- Van Wassenbergh, S.; van Manen, K.; Marcroft, T.A.; Alfaro, M.E.; Stamhuis, E.J. Boxfish swimming paradox resolved: Forces by the flow of water around the body promote manoeuvrability. J. Roy. Soc. Interface 2015, 12, 20141146. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vincent, J.F. Survival of the cheapest. Mater. Today 2002, 5, 28–41. [Google Scholar] [CrossRef]
- Knippers, J.; Menges, A. ICD/ITKE Research Pavilion 2011. In Architecture Research Building: ICD/ITKE 2010-2020; Menges, A., Knippers, J., Eds.; Birkhäuser: Berlin, Germany; Boston, MA, USA, 2020; pp. 56–65. ISBN 9783035620399. [Google Scholar]
- ICD/ITKE Research Pavilion 2011. Available online: https://www.icd.uni-stuttgart.de/projects/icditke-research-pavilion-2011/ (accessed on 21 September 2021).
- Speck, O.; Speck, D.; Horn, R.; Gantner, J.; Sedlbauer, K.P. Biomimetic bio-inspired biomorph sustainable? An attempt to classify and clarify biology-derived technical developments. Bioinsp. Biomim. 2017, 12, 011004. [Google Scholar] [CrossRef]
- MacKinnon, R.B.; Oomen, J.; Pedersen Zari, M. Promises and presuppositions of biomimicry. Biomimetics 2020, 5, 33. [Google Scholar] [CrossRef]
- Merriam-Webster-Definition of Growth Form. Available online: https://www.merriam-webster.com/dictionary/growth%20form (accessed on 4 October 2021).
- Speck, T.; Rowe, N.P. A quantitative approach for analytically defining size, form and habit in living and fossil plants. In The Evolution of Plant Architecture; Kurmann, M.H., Hemsley, A.R., Eds.; Royal Botanic Gardens: Kew, UK, 1999; pp. 447–479. [Google Scholar]
- Rowe, N.; Speck, T. Plant growth forms: An ecological and evolutionary perspective. New Phytol. 2005, 166, 61–72. [Google Scholar] [CrossRef]
- Putz, F.E. The natural history of lianas on Barro Colorado Island, Panama. Ecology 1984, 65, 1713–1724. [Google Scholar] [CrossRef]
- Gentry, A.H. The distribution and evolution of climbing plants. In The biology of Vines; Putz, F.E., Mooney, H.A., Eds.; Cambridge University Press: Cambridge, UK, 1991; pp. 3–49. [Google Scholar]
- Putz, F.E.; Mooney, H.A. (Eds.) The Biology of Vines; Cambridge University Press: Cambridge, UK, 1991. [Google Scholar]
- Schnitzer, S.; Bongers, F.; Burnham, R.; Putz, F. (Eds.) The Ecology of Lianas; Wiley-Blackwell: Chichester, UK, 2015. [Google Scholar]
- Rowe, N.P.; Isnard, S.; Speck, T. Diversity of mechanical architectures in climbing plants: An evolutionary perspective. J. Plant Growth Regul. 2004, 23, 108–128. [Google Scholar] [CrossRef]
- Rowe, N.P.; Speck, T. Hydraulics and mechanics of plants: Novelty, innovation and evolution. In The Evolution of Plant Physiology; Hemsley, A.R., Poole, I., Eds.; Academic Press: London, UK, 2004; pp. 301–329. [Google Scholar]
- Rowe, N.P.; Speck, T. Stem biomechanics, strength of attachment, and developmental plasticity of vines and lianas. In The Ecology of Lianas; Schnitzer, S., Bongers, F., Burnham, R., Putz, F., Eds.; Wiley-Blackwell: Chichester, UK, 2015; pp. 323–341. [Google Scholar] [CrossRef]
- Rowe, N.P.; Isnard, S.; Gallenmüller, F.; Speck, T. Diversity of mechanical architectures in climbing plants: An ecological perspective. In Ecology and Biomechanics: A Mechanical Approach to the Ecology of Animals and Plants; Herrel, A., Speck, T., Rowe, N.P., Eds.; Dekker Publishers/Taylor & Francis Group: Boca Raton, FL, USA, 2006; pp. 35–59. [Google Scholar]
- Niklas, K.J. Plant Biomechanics: An Engineering Approach to Plant Form and Function; University of Chicago Press: Chicago, IL, USA, 1992. [Google Scholar]
- Langer, M.; Kelbel, M.C.; Speck, T.; Müller, C.; Speck, O. Twist-to-bend ratios and safety factors of petioles having various geometries, sizes and shapes. Front. Plant Sci. 2021, 12, 765605. [Google Scholar] [CrossRef]
- Gallentine, J.; Wooten, M.B.; Thielen, M.; Walker, I.D.; Speck, T.; Niklas, K.J. Searching and intertwining: Climbing plants and GrowBots. Front. Robot. AI 2020, 7, 118. [Google Scholar] [CrossRef]
- Speck, T.; Schmitt, M.; Stahmer, E.-M.; Uhl, G.; Lange, J.; Parnesar, A. Tabellen. In Biologie im Überblick, Lexikon der Biologie; Schmitt, M., Ed.; Herder: Freiburg, Germany, 1992; Volume 10, pp. 187–328. [Google Scholar]
- Caballé, G. Liana structure, function and selection: A comparative study of xylem cylinders of tropical rainforest species in Africa and America. Bot. J. Linn. Soc. 1993, 113, 41–60. [Google Scholar] [CrossRef]
- Steinbrecher, T.; Danninger, E.; Harder, D.; Speck, T.; Kraft, O.; Schwaiger, R. Quantifying the attachment strength of climbing plants: A new approach. Acta Biomater. 2010, 6, 1497–1504. [Google Scholar] [CrossRef]
- Melzer, B.; Steinbrecher, T.; Seidel, R.; Kraft, O.; Schwaiger, R.; Speck, T. The attachment strategy of English Ivy: A complex mechanism acting on several hierarchical levels. J. R. Soc. Interface 2010, 7, 1383–1389. [Google Scholar] [CrossRef] [Green Version]
- Bohn, H.F.; Günther, F.; Fink, S.; Speck, T. A passionate free climber: Structural development and functional morphology of the adhesive tendrils in Passiflora discophora. Int. J. Plant Sci. 2015, 176, 294–305. [Google Scholar] [CrossRef]
- Rampf, M.; Speck, O.; Speck, T.; Luchsinger, R.H. Investigation of a fast mechanical self-repair mechanism for inflatable structures. Int. J. Eng. Sci. 2013, 63, 61–70. [Google Scholar] [CrossRef]
- Bardzinska-Bonenberg, T. Parasitic Architecture: Theory and Practice of the Postmodern Era. In Advances in Human Factors, Sustainable Urban Planning and Infrastructure, Proceedings of the AHFE 2017 International Conference on Human Factors, Sustainable Urban Planning and Infrastructure, Los Angeles, CA, USA, 17–21 July 2017; Charytonowicz, J., Ed.; Springer: Cham, Switzerland, 2018; Volume 600, pp. 3–12. [Google Scholar] [CrossRef]
- Baroš, T.; Katunský, D. Parasitic architecture. Sel. Sci. Pap.—J. Civ. Eng. 2020, 15, 19–28. [Google Scholar] [CrossRef]
- UdK Rooftop Building. 2021. Available online: http://www.solar-rooftop.de/en/index.html (accessed on 24 August 2021).
- Hachem-Vermette, C.; Dara, C.; Kane, R. Towards net zero energy modular housing: A case study. In Proceedings of the Modular and Offsite Construction (MOC) Summit Proceedings, Hollywood, FL, USA, 22–25 March 2018; Volume 8. [Google Scholar] [CrossRef]
- Mandal, J.; Jia, M.; Overvig, A.; Fu, Y.; Che, E.; Yu, N.; Yang, Y. Porous polymers with switchable optical transmittance for optical and thermal regulation. Jule 2019, 3, 3088–3099. [Google Scholar] [CrossRef]
- Haila, Y. Beyond the nature-culture dualism. Biol. Phil. 2000, 15, 155–175. [Google Scholar] [CrossRef]
- Krohs, U. Der Funktionsbegriff in der Biologie. In Wissenschaftstheorie. Texte zur Einführung; Bartels, A., Stöckler, M., Eds.; Mentis: Paderborn, Germany, 2009; pp. 287–306. [Google Scholar]
- Möller, M.; Antony, F.; Grießhammer, R.; Gröger, J.; Prieß, R. PROSA—Product Sustainability Assessment. In PROSAplus Guideline; Öko-Institut: Freiburg, Germany, 2020; Available online: https://www.prosa.org/fileadmin/user_upload/pdf/PROSA_Guideline_final.pdf (accessed on 8 October 2021).
- Behrendt, S.; Göll, E.; Korte, F. Effizienz, Konsistenz, Suffizienz, Strategieanalytische Betrachtung für eine Green Economy, Inputpapier im Rahmen des Projekts Evolution2Green—Transformationspfade zu einer Green Economy 2016. Available online: https://evolution2green.de/sites/evolution2green.de/files/documents/evolution2green_inputpapier_effizient_konsisten_suffizienz.pdf (accessed on 4 October 2021).
- Grießhammer, R.; Bunke, C.; Gensch, C. EcoGrade–die integrierte ökologische Bewertung. Entscheid. Für Unternehm. Umw. 2002, 10, 47–51. [Google Scholar]
- De Haan, P.; Peters, A.; Semmling, E.; Marth, H. Rebound-Effekte: Ihre Bedeutung für die Umweltpolitik; UBA-Texte: Berlin, Germany, 2015; Volume 31, Available online: https://www.bmu.de/fileadmin/Daten_BMU/Pools/Forschungsdatenbank/fkz_3711_14_104_rebound_effekte_bf.pdf (accessed on 21 November 2021).
- Potting, J.; Hekkert, M.P.; Worrell, E.; Hanemaaijer, A. Circular Economy: Measuring Innovation in the Product Chain, No. 2544; PBL Netherlands Environmental Assessment Agency: The Hague, The Netherlands, 2017; Available online: https://www.pbl.nl/en/publications/circular-economy-measuring-innovation-in-product-chains (accessed on 4 October 2021).
- Will, M. Towards a Sustainable Circular Economy–Remarks on plastics and wood-waste sector. Cent. Eur. Rev. Econ. Manag. 2019, 3, 149–183. [Google Scholar] [CrossRef]
- Haas, W.; Krausmann, F.; Wiedenhofer, D.; Heinz, M. How circular is the global economy?: An assessment of material flows, waste production, and recycling in the European Union and the world in 2005. J. Ind. Ecol. 2015, 19, 765–777. [Google Scholar] [CrossRef]
- Horn, R.; Dahy, H.; Gantner, J.; Speck, O.; Leistner, P. Bio-inspired sustainability assessment for building product development—Concept and case study. Sustainability 2018, 10, 130. [Google Scholar] [CrossRef] [Green Version]
- Speck, O.; Langer, M.; Mylo, M.D. Plant-inspired damage control–An inspiration for sustainable solutions in the Anthropocene. Anthr. Rev. 2022, 9, 220–236. [Google Scholar] [CrossRef]
- Heyen, D.A.; Fischer, C.; Barth, R.; Brunn, C.; Grießhammer, R.; Keimeyer, F.; Wolff, F. Mehr als nur Weniger. Suffizienz: Notwendigkeit und Optionen politischer Gestaltung; Öko-Institut Working Paper; Öko-Institut: Freiburg, Germany, 2013; Available online: https://www.oeko.de/oekodoc/1837/2013-506-de.pdf (accessed on 4 October 2021).
Figure 1.
Exponential increase in articles with the words “biomimetics and sustainability” published from 1999 to 2021. Number of publications is given in brackets after the year. (Raw data retrieved from Google Scholar on 27 June 2022).
Figure 1.
Exponential increase in articles with the words “biomimetics and sustainability” published from 1999 to 2021. Number of publications is given in brackets after the year. (Raw data retrieved from Google Scholar on 27 June 2022).

Figure 2.
Selected climbing plants. (a) Young self-supporting plant and (b) old ribbon-shaped stem of Bauhinia guianensis; (c) Intertwining stems of Aristolochia macrophylla; (d) Stem of Condylocarpon guianense that has slipped downward after losing its hold on the supporting tree; (e) Climbing plants (Actinida arguta) growing up a house wall and towering over the house; (f) Tendril of the Boston ivy (Parthenocissus tricuspidata) with adhesive pads as an example of an attachment structure; (g) Microscopic images of transverse sections of stems of Aristolochia macrophylla. Left side shows a one-year-old stem with a closed ring of strengthening tissue in the periphery (magenta arrows) causing a high flexural and torsional rigidity of the self-supporting young plant stems. Right side shows a two-year-old stem with newly formed flexible wood with huge vessels (blue arrows) and a segmented but healed ring of strengthening tissue in the periphery that cause a more flexible behavior in bending and torsion (yellow arrows); (h) Coiled stem of Bauhinia guianensis on the forest floor after tree breakage. (a,b,d,h) Tropical rain forest of French Guiana, France; (c,e–g) Outdoor area of the Botanic Garden Freiburg, Germany.
Figure 2.
Selected climbing plants. (a) Young self-supporting plant and (b) old ribbon-shaped stem of Bauhinia guianensis; (c) Intertwining stems of Aristolochia macrophylla; (d) Stem of Condylocarpon guianense that has slipped downward after losing its hold on the supporting tree; (e) Climbing plants (Actinida arguta) growing up a house wall and towering over the house; (f) Tendril of the Boston ivy (Parthenocissus tricuspidata) with adhesive pads as an example of an attachment structure; (g) Microscopic images of transverse sections of stems of Aristolochia macrophylla. Left side shows a one-year-old stem with a closed ring of strengthening tissue in the periphery (magenta arrows) causing a high flexural and torsional rigidity of the self-supporting young plant stems. Right side shows a two-year-old stem with newly formed flexible wood with huge vessels (blue arrows) and a segmented but healed ring of strengthening tissue in the periphery that cause a more flexible behavior in bending and torsion (yellow arrows); (h) Coiled stem of Bauhinia guianensis on the forest floor after tree breakage. (a,b,d,h) Tropical rain forest of French Guiana, France; (c,e–g) Outdoor area of the Botanic Garden Freiburg, Germany.

Figure 3.
Plus-energy rooftop unit positioned on a pre-existing building. Reprinted with permission from Ref. [51]. 2021, UdK Berlin.
Figure 3.
Plus-energy rooftop unit positioned on a pre-existing building. Reprinted with permission from Ref. [51]. 2021, UdK Berlin.

Figure 4.
Spectral hemispherical transmittance of a PPC device in the wet state (sunlight transmitted) and dry state (sunlight reflected) (with permission of Ref. [53]).
Figure 4.
Spectral hemispherical transmittance of a PPC device in the wet state (sunlight transmitted) and dry state (sunlight reflected) (with permission of Ref. [53]).


{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
Three levels of learning from living nature for technical development.
| Biological Model | Knowledge Transfer | Technical Development |
|---|---|---|
| Form–function–structure relationship | Functional principle | Biomimetic product |
| Example: Self-cleaning surfaces [21] | Micro-rough and hydrophobic surfaces plus surface tension of water droplets | Self-cleaning products with the trademark Lotus-Effect® |
| Mechanisms of evolution | Algorithm for biomimetic optimization | Biomimetically optimized components |
| Example: Growth laws of trees and bones [22] | Computer Aided Optimization (CAO) and Soft Skill Option (SKO) | Mercedes-Benz bionic car [23] |
| Processes of evolution | Biological concepts | Bioinspired strategies |
| Example: Survival of the cheapest [24] | Modularity concept | Modular design ICD/ITKE Research Pavilion 2011 built from wood modules [25,26] |
Table 2.
Compilation of biological concepts found in the growth form of lianas (green) and attribution of the concepts to sustainability strategies of efficiency, consistency, and sufficiency (blue).
Table 2.
Compilation of biological concepts found in the growth form of lianas (green) and attribution of the concepts to sustainability strategies of efficiency, consistency, and sufficiency (blue).
| Characteristics of Lianas | Biological Concepts | Strategy Features | Sustainability Strategy |
|---|---|---|---|
| growth form has evolved several times independently in different taxa | best fit | optimal embedding in ecosystems | consistency |
| rapid and complete decomposition | zero waste | improvement of circular economy through recycling | consistency |
| from a self-supporting searcher stem to a climbing plant stem | change of growth form | change of lifestyle with reduction in material use | sufficiency |
| ratio of flexural and torsional rigidity | trade-off | creation of different (partly conflicting) functions while minimizing resources | efficiency |
| parenchyma cells instead of sclerenchyma/wood cells fill the fissures | good enough | creation of a sufficient function | sufficiency |
| intertwining stems | modularity | improvement of a function while minimizing resources | efficiency |
| wood provides water transport and mechanical stability | multifunctionality | creation of multiple functions with a minimum quantity of resources | efficiency |
| little investment in own stem material | external support | creation of a function with a minimum quantity of resources | efficiency |
| attachment structures newly evolved from an organ with a formerly different function | change of function | new function for embedding a product in its environment | consistency |
| various tissue types | function-related tissue formation | creation of a function while minimizing resources | efficiency |
| formation of less dense wood | lightweight construction | creation of a function with a minimum quantity of resources | efficiency |
| disintegration of a continuous strengthening ring into individual parts | less is more | renunciation of superfluous functions | sufficiency |
| self-sealing and self-healing | damage repair | improvement of circular economy through repair | consistency |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Speck, O.; Möller, M.; Grießhammer, R.; Speck, T. Biological Concepts as a Source of Inspiration for Efficiency, Consistency, and Sufficiency. Sustainability 2022, 14, 8892. https://0-doi-org.brum.beds.ac.uk/10.3390/su14148892
AMA Style
Speck O, Möller M, Grießhammer R, Speck T. Biological Concepts as a Source of Inspiration for Efficiency, Consistency, and Sufficiency. Sustainability. 2022; 14(14):8892. https://0-doi-org.brum.beds.ac.uk/10.3390/su14148892
Chicago/Turabian StyleSpeck, Olga, Martin Möller, Rainer Grießhammer, and Thomas Speck. 2022. "Biological Concepts as a Source of Inspiration for Efficiency, Consistency, and Sufficiency" Sustainability 14, no. 14: 8892. https://0-doi-org.brum.beds.ac.uk/10.3390/su14148892
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.