
Pluralistic Valuation of Codling Moth Regulation by Brown Long-Eared Bats in English Apple Orchards
1
Sustainability Research Institute, School of Earth and Environment, The University of Leeds, Leeds LS2 9JT, UK
2
Community Development and Applied Economics, The University of Vermont, Burlington, VT 05401, USA
*
Author to whom correspondence should be addressed.
Sustainability 2022, 14(19), 11966; https://0-doi-org.brum.beds.ac.uk/10.3390/su141911966
Submission received: 27 June 2022
/
Revised: 7 September 2022
/
Accepted: 15 September 2022
/
Published: 22 September 2022
(This article belongs to the Collection Sustainability in Agricultural Systems and Ecosystem Services)
Abstract
:The benefits humans utilise from the ecological interactions of the Brown Long-eared bat (Plecotus auritus) have gone relatively unnoticed in the UK. This study aimed to expose one such interaction through a pluralistic valuation of Codling moth (Cydia pomonella) regulation by P. auritus in the context of English apple orchards. A stage-structured model linking C. pomonella population dynamics through consumption by P. auritus was created to understand crop loss reduction estimates due to this predation. The model found that the presence of a single P. auritus individual resulted in a significant reduction of 81.06–83.68% crop lost per hectare. The results of the avoided costs methods yielded a range of economic estimates with between GBP 307.59 and 604.95/ha private, ecological and social costs being avoided. Traditional biodiverse apple orchards that support P. auritus habitats are at risk from land development and agricultural intensification which would have serious consequences on biodiversity. The findings of this study could form the basis of further research that informs apple orchard managers, local communities dependent on apple production and policymakers of the importance of ecosystem services in sustainable agriculture.
1. Introduction
In 2019, the estimated combined value of dessert and culinary apples grown in the UK was valued at GBP 182.8 million, and GBP 24.4 million for cider apples and pears [1] Despite much of the nation’s fruit consumption being supplemented with internationally imported apples providing cheap produce throughout the year [2], the production of UK apples represents a significant market. That said, the importance of apple orchards is not limited to purely financial significance. Traditional apple orchards, which are low-intensity managed systems, provide a variety of habitats which in turn support a high level of biodiversity. The ongoing Traditional Orchards project by the People’s Trust for Endangered Species has to date found that this important habitat has declined by 90% in England since 1950, with key pressures being cited as agricultural intensification, land development and neglect [3,4]. These findings in part led to traditional apple orchards being designated a Priority Habitat under the Natural Environment and Rural Communities (NERC) Act in 2006. Bats (Chiroptera spp.) are just some of the varied species found in traditional orchards and are also a priority species under NERC Act.
Globally, humans utilise many ecosystem interactions of bats to benefit production including pollination, insect regulation, nutrient distribution through guano and seed dispersal services [5]. Collectively, these benefits can be referred to as ecosystem services. Ecosystem services are defined by the UK National Ecosystem Assessment (UKNEA) as being benefits to human life provided by ecosystems. This definition focuses upon the final benefit obtained by humans and fails to recognise the intrinsic value of the complex ecological systems behind the benefits. Fisher et al. [6] provides an alternative definition: “Ecosystem services are the aspects of ecosystems utilised (actively or passively) to produce human well-being.” It is this definition that was used throughout the study. Although the concept of ecosystem services is intrinsically anthropocentric, the Fisher et al. [6] definition takes a more ecologically focused perspective, acknowledging that ecosystems occur regardless of whether humans derive benefits from them. Conflicting definitions of ecosystem services bring about difficulties for developing standardised valuation methods. A discussion of the disparities of valuation methods has been delineated in the literature review with a focus on comparing monistic and pluralistic approaches to valuation methods [7,8].
This paper considers to what extent Brown long-eared bats (P. auritus) contribute to UK apple production through regulation of the pest, Codling moth (Cydia pomenella). The overarching aim of the study was to pluralistically assess the extent of C. pomonella regulation by P. auritus in the context of English apple orchards. More specifically, the aim is to show how pes-control may be considered through non-traditional means, i.e., the ecosystem service provided by P. Auritus to elucidate alternatives to single-financial metric orchard management. To reach this aim a mathematical stage-structured C. pomonella population model was created which featured consumption by P. auritus and then further valued using an avoided costs methodology.
We hope that the findings of this study will aid apple orchard managers in implementing integrated pest management strategies, reducing the quantity of pesticides required, and in turn potentially ease the environmental, social, and financial pressures of pest control. As this study only addresses one specific part of P. auritus ecology, we believe the findings have the potential to provide a baseline for further research into the complex interactions of the P. auritus with English apple orchards and other agricultural systems.
2. Background
2.1. Ecosystem Services and Economics
Ecosystem services are the product of complex ecological and environmental cycles that human existence relies upon [9] despite our ‘urbanisation’ and technological advancements. The term in its current mainstream form was coined by Paul and Anne Ehrlich in 1981 in ‘Extinction: The Causes and Consequences of the Disappearance of Species’, although variants of this term existed prior [10,11,12]. The broad definition of ecosystem services can be broken down into four categories: provisioning (physical goods; food, fuel, fibre), regulating (regulation of climate, disease, water, pollination), supporting (nutrient cycling and soil formation) or cultural services (aesthetics, cultural heritage, spiritual) [6]. There is no single agreed upon definition of ecosystem services; the field is extensive and only a brief discussion of the fundamental ideas has been presented here.
Increasingly, environmental economists advocate for valuation and ‘internalising’ of ecosystem services in what Spash [13] has referred to as a ‘new environmental pragmatism’. Under orthodox economic theory, the costs or benefits where neither the producer nor the consumer bear the full costs of or benefits to production are referred to as ‘externalities’ [14]. For example, with regard to the production of apples, ecosystem services may come in the form of pest regulation by birds, bats and predatory arthropods [15,16,17], pollination of the apple flowers [18], or the provision of nutrients in the soil which are used by the trees [19]. All of these are beneficial to apple production, but the grower does not pay for these ‘positive externalities.’ On the contrary, the eutrophication of water bodies by farm fertiliser run-off is also not incorporated in the market price and so is a ‘negative externality.’ Neo-classical economics posits that the internalisation of externalities in the market price will allow the market to efficiently allocate resources, ensuring production output reaches a more sustainable equilibrium point. Some environmental economists question such a monistic approach but nevertheless maintain that communicating the significance of nature in the language of economics (e.g., price, supply, and demand) will encourage policy-makers to conserve ecosystems [20].
Contrastingly, ecological economists criticise the mainstream monetary valuation of ecosystem services as the commodification of nature. Even from a new environmental pragmatism approach it is argued this ultimately leads to the same conclusion due to the structures in which environmental policy currently exists in [21]. As Spash and Aslaksen [8] argue, to incorporate ecological systems into the market results in ecosystems and biodiversity only being deemed necessary in their support of the economic system. Once those ecosystems or species are deemed “uneconomic” then there stands little reason under the market system to sustain them. Furthermore, the incorporation of finite ecosystems into an economic system that strives for continued economic growth, is to impose the same growth imperative on a system which will eventually reach its limit, leading to complete ecosystem crash.
De Groot [22] argues that quantification of ecosystems in monetary terms should always be an addition to intrinsic values, not a replacement of. Importantly, ‘tipping points’ are a feature of most ecosystems, which if crossed the ecosystem will rapidly change to an alternative steady state [6]. Monetary valuation alone does not reflect this. Similarly, the neoclassical concept of marginal analysis is not applicable to ecosystems as say, the loss of just a single species from an ecosystem could have innumerable unknown consequences on the wider system through loss of species-specific interactions [23]. Whilst the provision of financial value as a single metric is problematic, it may still have its merits within a pluralistic approach under some contexts. Gómez-Baggethun and Martín-López [24], amongst many others [8,22] advocate for a ‘value pluralism’ approach. A rejection of reductionism, pluralistic valuations feature multiple metrics, often in contradiction with one another, to evaluate the ecological, social and economic values. It is this pluralistic approach that was employed in this study.
Numerous methods have been deployed in attempts to measure the value of ecosystem services with many being aimed towards financial quantification. Typically for services involved in the production of goods (i.e., regulation services), an avoided-costs methodology may be utilised [9] where the value of the service is equal to the costs (financial, social and/or ecological) that would be incurred in the absence of the service. Problems occur however when determining which avoided impacts should be included in the cost and how to measure said impacts. Taking the example of apple production, should the avoided cost cease at the avoided private costs of crop loss? Or should the avoided cost extend to ecological and social costs to the production of apples (‘negative’ externalities) such as pollution of communal water supplies through the use of fertilisers and pesticides, impacts of said chemicals to fauna using the orchards or on workers’ health [25]? Often avoided cost models barely scratch the surface of the true value of a service by failing to incorporate the wider environmental or social consequences. Whilst some avoided cost calculations are relatively simple to implement (i.e., avoided crop loss) others such as avoided impacts to workers health or pollution of public/communal waterways are more difficult. Where the social and environmental costs of production are attempted to be quantified there can be large disparities in the estimated values depending on the methods used, the species in question or the internal biases the researcher brings to the study [9].
2.2. Apples and Arthropods
Apple trees are considered relatively short-lived hardwood trees and so begin to exhibit ageing features, such as rot holes, peeling bark, and hollow trunks, which support a large number of species including invertebrates. Traditional apple orchards support a vast number of arthropod species, with some interactions being detrimental to the production of fruit and others benefiting both arthropod and trees. Cross et al. [17] estimated that unsprayed apple orchards support over 2000 arthropod species, some of which engage in a mutualistic relationship with the trees. Apple trees are typically self-incompatible and are consequently reliant upon pollinating insects to ensure the fruit is set and for the production of larger apples [18,26]. Carrack and Williams [18] estimated that the value of pollinators to UK apple trees was worth approximately GBP 81.7 million in 1996, or approximately GBP 170.2 million in 2020 inflation-adjusted pounds [27]. In contrast, approximately one quarter of the 2000 arthropods identified in unsprayed apple orchards pose some threat of damage to apple trees [17]. C. pomonella is just one of the arthropod species with the potential to cause vast damage to apples, with Woolly Aphids (Eriosomatinae spp.) and Dysaphis plantaginea (Rosy Apple Aphid) also being cited by orchard managers as causing significant damage [28]. C. pomonella, the codling moth, emerges from overwintering pupae into the final adult form. Eggs are laid singly on the leaves of the apple tree or on the skin of the apples [29]. Gestation generally occurs within 7–10 days [30], releasing larvae that then searches for apples to infest, a process which usually takes just 24 h. Once inside the apple the larvae will consume the fruit until it is ready to pupate. Pupation occurs outside of the apple, typically under loose tree bark [31] hidden out of sight of predators until the cycle begins again.
Human control of C. pomonella often comes in one of three methods: pesticide application, mating disruption, and/or deployment of a granulosis virus. Use of microbial control with granulosis virus is usually seen in organic orchards. Mating disruption is the release of a synthetic pheromone to cause olfactory confusion and prevent the moth’s ability to find a mate [32,33]. The effectiveness of insecticides in their role is well documented, however, the wider impacts of insecticide use should be evaluated. Firstly, pesticides exert a selective pressure on pest populations, favouring those individuals with resistance, something which is accelerated through overapplication [34]. Pesticide resistance in C. pomonella has been recorded in varying degrees [35,36]. In C. pomonella’s final stages resistance yields current pesticides futile and incurs greater costs to orchard managers. Other recorded possible adverse effects of pesticide use include loss of biodiversity in aquatic ecosystems [37,38], non-target effects on beneficial arthropods [39] and an array of human health issues [40]. Neonicotinoids accounted for approximately 22% of pesticides used on UK orchards in 2018 [28] despite growing concerns surrounding their adverse effects on bee populations [41,42]. Even where attempts are made to increase the abundance of ‘beneficial’ arthropods, for example through the sowing of wildflower strips, pest regulation and pollination services by those arthropods are restricted through the use of pesticides [43].
2.3. Bat Ecology
There are 17 species of bat (Chiroptera spp.) known to breed in the UK, all of which are nocturnal and obligate insectivores. Most species of UK bats primarily feed, communicate, and navigate using echolocation [44], the emitting of high frequency sounds and subsequent return of sound waves to inform the bat of its surroundings and prey. Traditionally, apple orchards feature rows of evenly spaced trees surrounded by windbreak hedges providing essential linear landscape features used by echolocating bats for navigation and hunting opportunities [45]. Linear landscape features, the abundance of prey, and the relative temporal stability of the traditional orchard landscape provide suitable habitats for bats.
Of the 17 species of bats known to breed in the UK, only 4 of these have been explicitly recorded in existing studies as consuming C. pomonella: Eptesicus serotinus [46], Pipistrellus pygmaeus [47], Plecotus auritus [48], Rhinolophus hipposideros [49]. P. auritus was chosen as the focus of this study due to the relative abundance of available population dynamics data including one study specific to the South-East UK [50]. P. auritus is widespread throughout the British Isles with some exceptions on the smaller Western Isles. Low wing aspect ratio, large ears and relatively large eyes allow P. auritus to hunt, primarily moths, within open woodland and parkland [44] and so are well adapted to the scattered trees and pasture landscape of traditional orchards.
2.4. Bat Pest Regulation Services
As all UK bat species are insectivorous, the focus of this study will be its potential insect regulation services. In their role as an insectivorous predator across many habitats in the UK, some species of bats are considered as indicators of biodiversity and wider landscape change [51]. Despite their recognised importance as key predators, the DEFRA/AHDB ‘Apple best practice guide’, a guide for apple producers, fails to highlight bats (or birds) as natural enemies to apple insect pests. Only 26% of Spanish apple growers interviewed by Martínez-Sastre et al. [52] acknowledged bats as a source of pest regulation. Contrary to this general unawareness, across the reviewed literature there is a general consensus that bat feeding ecology has a significant deprecating impact on several crop pest populations and resulting reduction in damage to crops [53,54,55,56,57,58]. Internationally, studies exist for bat pest regulation ecosystem services in a wide array of crops, the greatest proportion focussed on North American landscapes and species, followed by European studies. None of the European studies of bat pest regulation ecosystem services, however, were based in, or upon species that also occur in, the UK.
It is important to understand in depth the pest regulation services provided by natural enemies, how they can be enhanced, and how these services are impacted by human interventions. It is possible that P. auritus could considerably contribute to C. pomonella regulation and therefore to the production of apples in the UK and yet there is no literature on this potential contribution nor how current artificial pest regulation strategies could be impacting this service. Being mindful of the criticisms of orthodox ecosystem service valuation, the ecological economic concepts obtained from this literature review were applied in the creation of a mathematical model as outlined in Section 3.
3. Method
London and the South-east of England has the greatest proportion of apple orchards in England accounting for 39% of UK orchards whilst disproportionately counting for 67% of pesticide applications [28]. For this reason, the study was based upon a hypothetical 1 hectare orchard in this area. In regard to adult P. auritus, non-pregnant female and male individuals are required to consume on average approximately 1.8 g (dry mass) per night. Due to the increased energetic requirements of pregnancy, pregnant and lactating individuals consume approximately 2 g (dry mass) per night [59]. To incorporate these differences in consumption and to further control for variation such as intra-population dynamics, the study observed a single bat’s impact. Observations regarding life stage timings specific to C. pomonella and P. auritus, the first moth flight typically occurs within the same time period as P. auritus females will be pregnant whereas the second moth flight (if it occurs) would coincide with the end of the lactation period. For this reason, the ‘reproductive’ female P. auritus will be henceforth referred to as a pregnant female throughout the study although the daily energetic requirement of pregnant and lactating P. auritus are believed to be equal [60].
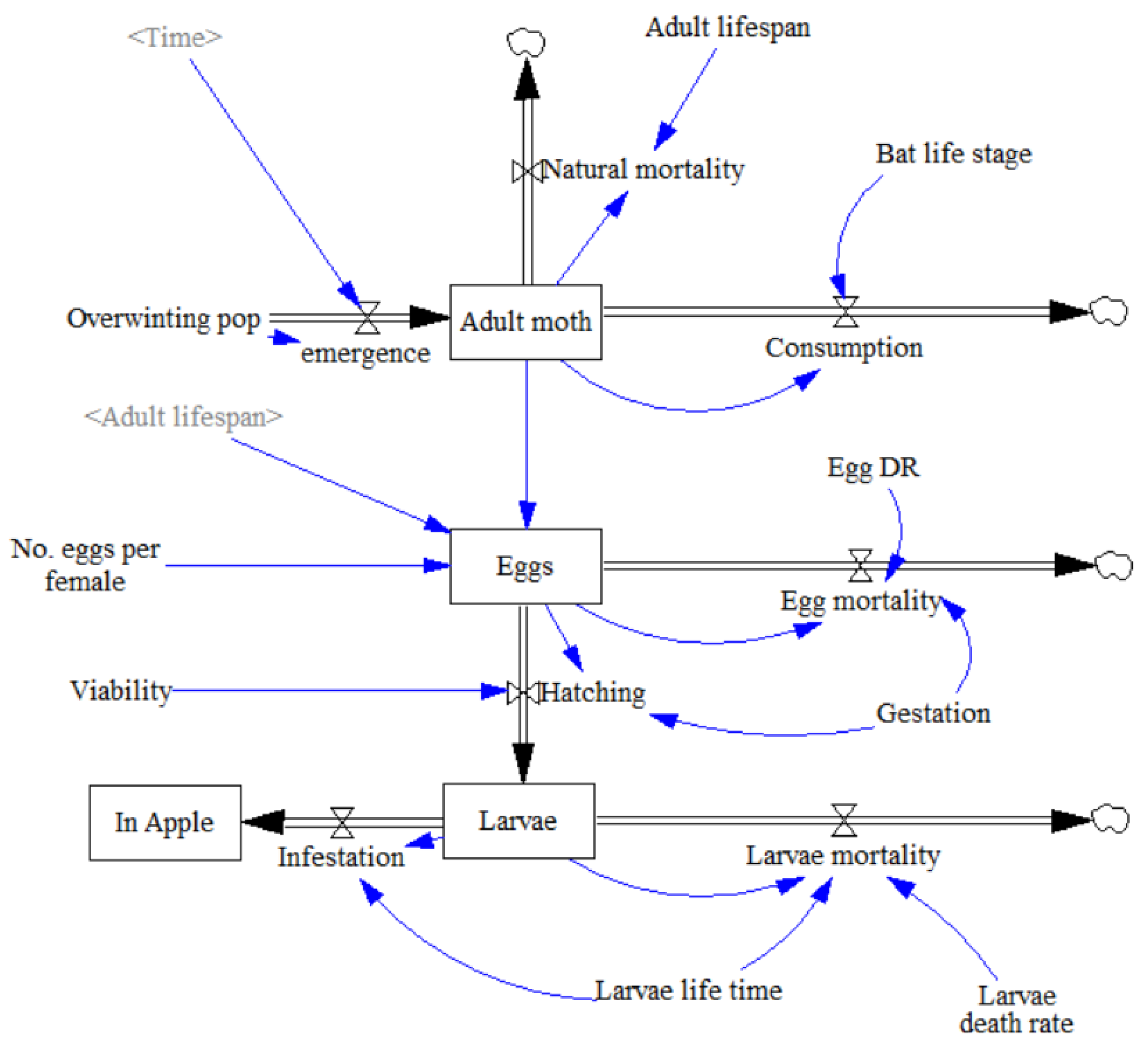
3.1. Establishing the Model
An intra-seasonal, stage-structured mathematical model was constructed in Vensim PLE 8.2.0 based upon the system dynamics of a Cydia pomonella population and the predator-prey interactions of a single Plecotus auritus in a UK apple orchard (Figure 1). This mathematical model drew upon the methods of Cleveland et al. [53] and Federico et al. [54]. C. pomonella life stages were first established. The model spanned 122 simulated days representing a typical period between May and the end of August. In particularly warm years C. pomonella can undergo 2 generation cycles in a single season. To simulate an average year and accommodate for comparison, the study was restricted to just one generation of adult moths and resulting larvae damage. The single generation is represented in the model as the larvae enter the apples, but this does not loop back round to the adult moth population. Within the real system the larvae would then leave the apples to pupate before emerging as adult moths. The assumption was made that the system was closed with no migration in or out of the model population, which for C. pomonella is likely often true [61]. Likewise, P. auritus also show very high roost fidelity [44,62,63].
Following the establishment of the C. pomonella life stages, the transitory equations were formulated. Each number below is a reference to the numbers on Figure 1.
- 1
- Spring emergence of adult moths from their overwintering pupating stage, with a peak between May and June, was first established using a differential equation:where P = Overwintering population; te = Time at end of emergence period (day); t = Current time (day).
- 2
- Natural mortality rate of adults was calculated through current number of adults divided by average life expectancy assuming that all adult moths which are not consumed by the bat live out their full life expectancy and fecundity.where N = Current C. pomonella population; Ta = Adult C. pomonella lifespan.
- 3
- Consumption by the bat was also dependent upon the density of adult moths as follows:where C = P. auritus nightly consumption (g); η = Proportion of C. pomonella in P. auritus diet; γ = Mean C. pomonella body weight (mg); N = Current C. pomonella population; κ = Initial overwintering C. pomonella population.As well as being dependent on bat reproductive stage, nightly pest consumption rate is also dependent on the weight of prey and proportion of prey in the bat’s diet. Studies attempting to calculate the average proportion of Lepidoptera in P. auritus diet have attained widely different values. Although a West of Ireland study by Shiel et al. [64] is the geographically closest estimation, a more recent faecal analysis study in Switzerland by Andriollo et al. [65] utilised DNA metabarcoding, identifying a broader taxonomic diversity, Andriollo et al. [65] found that C. pomonella occurred in 7% of P. auritus guano from across 9 different roost sites. It was assumed C. pomonella constituted up to 7% of P. auritus diet. The current moth population expressed as a proportion of the peak population ensures that the bat was not able to consume more moths than were currently present. Where the current population of C. pomonella reached peak population size, codling moth was assumed to account for the full 7% in the P. auritus diet as it was deemed abundant.
- 4
- The egg-laying function was integrated into the egg life stage parameter. Number of eggs at any given time was calculated as “Eggs laid–Egg mortality–Hatching”. Assuming a sex ratio of 1:1 [66], half of the adult moth population were assumed to be female and lay a set number of eggs in their full lifetime, established with a rate of number of eggs divided by adult life expectancy. This can be visualised in the equation:where N = Current C. pomonella population; Ef = Number of eggs per female; Ta = Adult C. pomonella life expectancy.
- 5
- Egg mortality was calculated by number of eggs at given time multiplied by egg death rate divided by gestation time. Dividing egg death rate by gestation time ensured that the net proportion of eggs dying was equal to the egg death rate.where Et = Number of eggs at time t; De = Egg death rate; Tg = Gestation time.
- 6
- Hatching was calculated as a rate of viable eggs over gestation time:where Et = Number of eggs at time t; V = Proportion of viable eggs; Tg = Gestation time.
- 7
- Larvae mortality was calculated in a similar manner to egg mortality with number of larvae (i.e., Eggs hatched) multiplied by Larvae death rate over larvae life time.where Lt = Number of larvae at time t; Dl = Larvae death rate; Tl = Larvae life expectancy.
- 8
- The equation for infestation was number of larvae at time t divided by larvae life expectancy. It was assumed that one larva infests one apple and multiple larvae cannot infest the same apple [67].where Lt = Number of larvae at time t; Tl = Larvae life expectancy.
3.2. Data Collection
Parameter values for C. pomonella were gathered from a wide selection of sources (Table 1). Papers were selected from areas with a comparable climate to South-East England or where this was not possible, the most similar. Graf et al. [68] provided in-depth captive estimates of C. pomonella life and seasonal history, providing the basis for the model timings. Viability and stage-specific mortality rates came from three papers: Kuyulu & Genc [69], Glen & Milsom [70], and Glen [71]. Whilst Kuyulu and Genc [69] obtained the findings from laboratory experiments, Glen [71] and Glen and Milstrom [70] directly observed C. pomonella in the field in South-West England making this study the closest geographically and likely to be highly applicable to populations in the South-East. Egg and larvae mortality rates were inclusive of predation from arthropods or birds, respectively, predation which is unlikely to exert competition pressure on the bats feeding on adult moths. The baseline model parameter values were generally the reference value given by the respective paper or where this was not available, the median value of the range (Table 1, see also Appendix A). A range of potential values was also typically given, and these ranges were explored in the sensitivity analysis.
Spatial population estimates for C. pomonella adults are not readily available. A study by Adams et al. [72] attempted to estimate actual pest densities from the number of male moths caught per pheromone trap, a popular method of monitoring moth pests. ADHB suggests one trap per orchard to monitor C. pomonella numbers and a threshold of 5 male moths caught per trap before taking reactive measures. According to the Adams et al. [72] estimates, 5 male moths approximately translate to a population of 24 C. pomonella individuals (male and females) per hectare. A UK study monitoring codling moth found the peak number of male C. pomonella caught per trap in the South-east of England, averaged between 1972 and 1974, was 6.3 male moths [73]. Using the Adams et al., estimate this equates to approximately 30 moths per hectare at the peak of the population. Reversing the emergence equation of the model and accounting for natural adult mortality, in the absence of the bat, a peak of 30 moths at day 7 equates to 60 overwintering moths.
3.3. Sensitivity Analysis
For a greater in-depth analysis of the model and individual parameters, the model was passed through Jupyter Notebook using the PYSD library. To test the limits of the model and determine relative impacts individual parameters have on the final output, each parameter was first manipulated individually within its biologically feasible range in ten increments. Final number of infested apples (i.e., crop damage) was used as the primary output for the sensitivity analysis. The non-reproductive bat was included for the sensitivity analysis as the presence of the predator changes the expression of some of the parameters and as the model was focussed on the predator-prey interactions, it was deemed important to observe the impact of individual parameters within the confines of this interaction.
Two extreme scenarios were developed. With the baseline model representing an ‘average’ season condition, the two extreme scenarios were: a year of high C. pomonella reproduction and a low reproductive year. Daily temperatures account for a large extent of the variation seen within C. pomonella populations [74,75]. Warmer temperatures typically coincide with higher reproduction and vice versa. In the high reproduction year C. pomonella timings are at their shortest (such as adult and larvae lifespan) allowing for rapid life cycles, low natural mortality rates and greatest number of eggs per female. Whilst number of eggs produced has no significant correlation with temperature, higher temperatures correlate with a higher and earlier laying peak [68]. The low reproduction scenario represents the opposite with long life stage timings, higher natural mortality rates and a more prolonged egg laying period [68]. Given the relationships between atmospheric temperature and C. pomonella breeding conditions, these extreme scenarios were deemed particularly important to predict the impact under various seasonal conditions.
3.4. Avoided Costs
Avoided costs methods of valuing regulating ecosystem services are the costs that otherwise would have been incurred due to the pest in the absence of the regulator (P. auritus). This is inclusive of the avoided costs of crop loss to the pest and the costs of insecticide that would have been required to counter the pest population. The avoided costs of chemical control can be broken down into private, environmental and social costs. In this study the private costs come from crop loss (apples) infested by C. pomonella larvae and the market price for pesticides to counter the pest population. Annual average cost of apples in the UK for the last full economic year was available through DEFRA [1] and was given in sterling pounds per kilogram. Cider apples account for the greatest area of orchard crops grown in the UK (38%) followed by dessert apples (non-Cox) [28]. In the South-East of England, apple cider is traditionally made from dessert apples [76] and so the estimate value of the crop in this study focussed on desert apples (non-Cox). The two most common non-Cox dessert apples grown in the UK in 2018 were Gala and Braeburn [28]. As most fruit are sold by the kilogram due to the great variation in individual fruit weights, it is difficult to obtain the average weight of a single apple. Using supermarket estimates the mean price for dessert apples in 2020 in the UK was GBP 1.02/kg which, when using an average weight of 0.175 kg [77], roughly equates to GBP 0.18 per apple.
4. Results
The model was considered logistically sound in that it did not return negative population values, followed the timings and shape of C. pomonella populations observed in the literature [68,74] and bat consumption followed the relative abundance of adult moths. As population estimates for codling moth on a local scale are not readily available, the number of adult moths in the population could not be verified against recent local field estimates. All population output values were rounded to the nearest integer to accurately represent the ecosystem as partial moths are not consumed and partial apples are not infested in the field.
4.1. Baseline Model Results
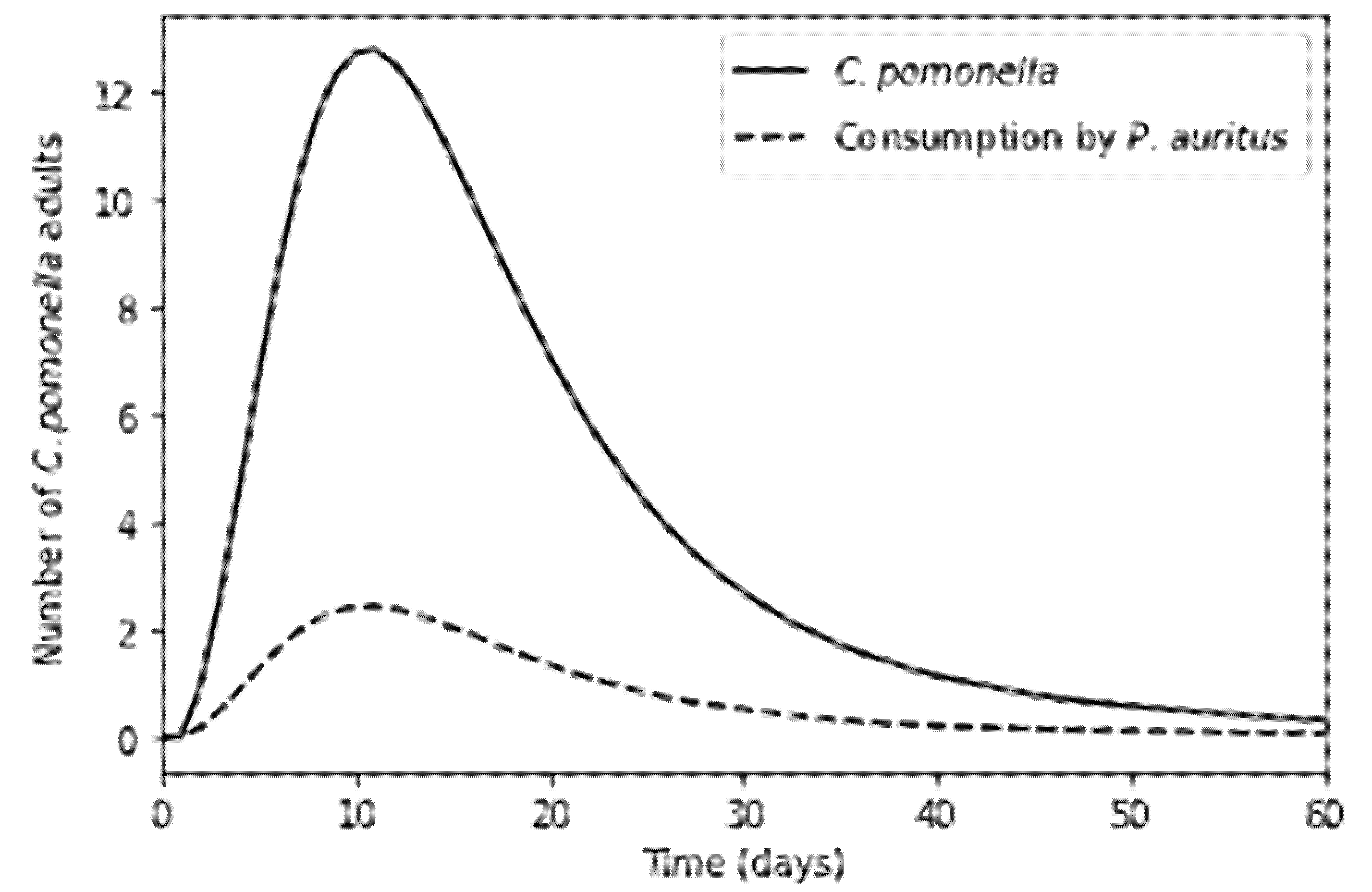
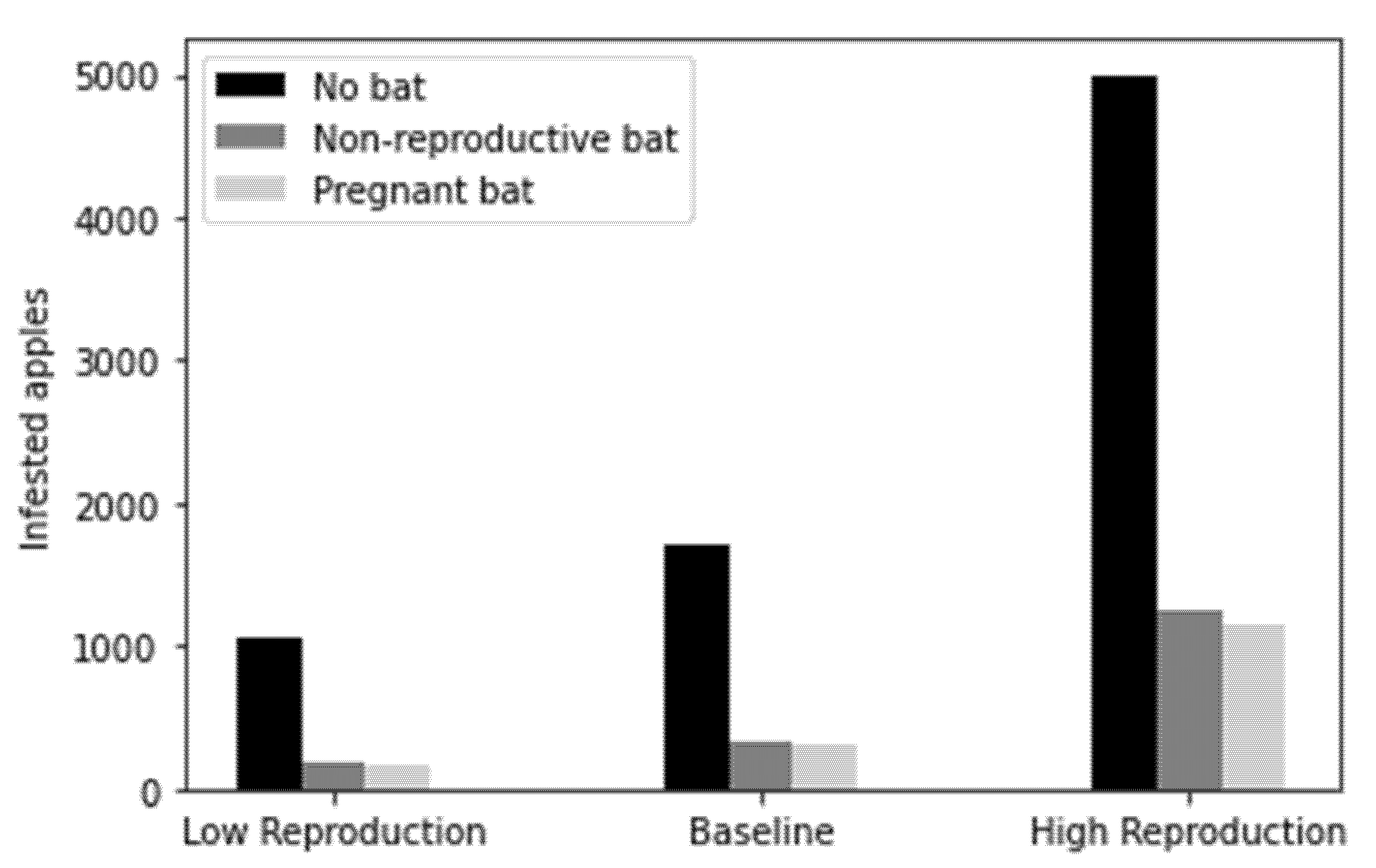
Bat consumption followed the adult moth population trajectory as intended (Figure 2), with the greatest consumption rates occurring at peak moth population and then dropping again as the population narrowed to zero. Although there were 60 initial overwintering moths as pupae, the emergence rate, natural mortality rate and consumption by the bat, mean that the number of adult moths only reaches 12 individuals at its peak. The adult population of C. pomonella entirely died off by day 60, after which point the bat’s diet would presumably shift to alternative prey entirely. For the baseline model, with a starting population of 60 overwintering C. pomonella, 47.95 or 79.92% of that adult moth population were consumed by a single non-reproductive P. auritus over the 60-day period in the model. This resulted in a decrease in infested apples by 79.95%, whilst a pregnant bat caused a decrease of 81.59% through the consumption of 81.50% of adult moths. The damage to crops from an overwintering population of 60 moths in the absence of the bat was 1705 apples infested yielding them unsellable, whereas in the presence of a non-reproductive bat 342 apples were infested and 314 apples infested in the presence of a pregnant bat (Figure 3).
4.2. Sensitivity Analysis
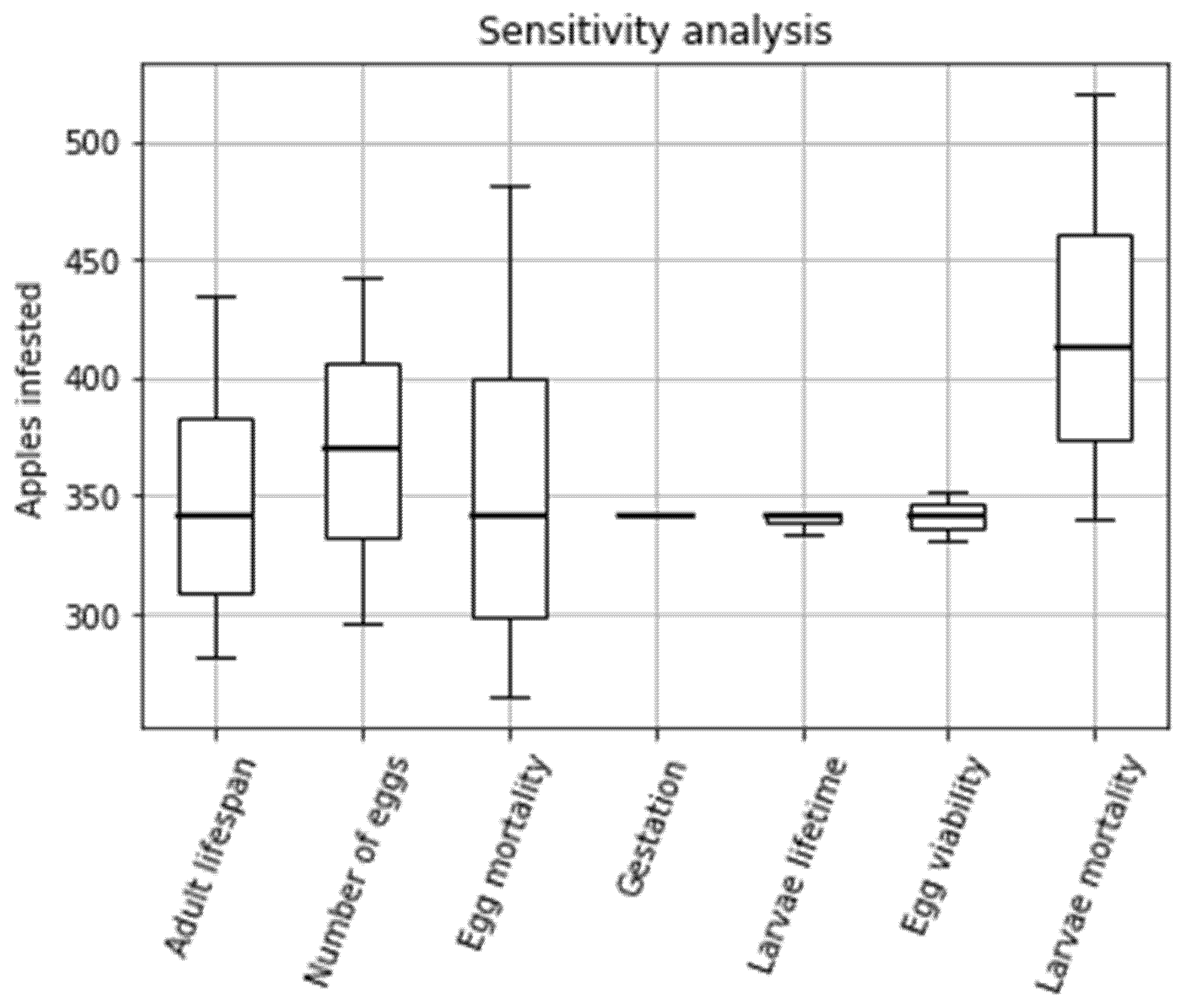
Through incremental analysis three parameters were identified as accounting for the greatest variation in the model (Figure 4). ‘Egg mortality’ accounted for the greatest variation with a range of 265–482 infested apples, followed by ‘Larvae death rate’ (range of 180 infested apples) and ‘Number of eggs per female’ (range of 147 apples). ‘Egg mortality ’, as expected, causes such a large difference in infested apples as at low mortality rates, a greater number of eggs are surviving to larvae and infesting apples. The same applies for ‘Larvae mortality ’. With regard to ‘Number of eggs per female’ an increase in eggs led to an increase in larvae which infest a greater number of apples. One would expect ‘egg viability’ to follow a similar pattern but instead yielded a range of just 21 apples. It could be assumed that this is due to the smaller value range for ‘Egg viability’ as recorded in the literature. ‘Adult lifespan’ also produced a relatively large range of results (153 apples) which can be explained by the fact that with longer activity times, there are more opportunities there are to be consumed by the bat and as the period of egg laying is proportional to temperature as is lifespan [68], the longer lifespan (under colder conditions) does not necessarily equate to greater productivity as the warmer temperature will increase the speed of egg-laying.
The low reproductive year resulted in a loss of 185 apples, whilst the baseline scenario and high reproductive scenario saw a loss of 342 and 1245 apples, respectively (Figure 3). For a pregnant P. auritus the difference in crop loss for the three scenarios is slightly larger, with the most and least preferential years resulting in a loss of 1149 and 169 apples, respectively (Figure 3). As the reproductive success of C. pomonella increases so does the amount of yield lost to a single adult through an increased number of successful larvae, therefore increasing the avoided cost value per moth consumed.
4.3. Avoided Costs
Using the reference values from the stochastic model and the estimated average price of dessert apples at GBP 0.18/apple, the avoided crop loss is valued between GBP 191.88 and 489.24 for a non-reproductive P. auritus and GBP 221.58 and 324.90 for a pregnant P. auritus (Table 2).
Regarding the avoided private costs of insecticide use (in this case chlorantraniliprole), the avoided private cost to the orchard manager was calculated using the previous estimate of an average of 34 g of chlorantraniliprole active substance used per dose per hectare [28]. Chlorantraniliprole containing pesticides typically contain 200 g/L of active substance [78], meaning approximately 170 mL of pesticide was used per hectare in 2018. The average cost of popular chlorantraniliprole containing pesticide, Dupont Coragen, was approximately GBP 58.74 for a 170 mL dose (based upon prices from four UK retailers). Where applied, an average of 1.95 applications was required [28] using the average Coragen cost, this would cost GBP 114.54.
The University of Hertfordshire’s Pesticide Properties Database refers to chlorantraniliprole as environmentally persistent, high toxicity to aquatic invertebrates, moderate toxicity to aquatic plants and birds, and low oral mammalian toxicity but with possible implications on human reproduction/development. Per Leach & Mumford [79] and Kovach et al. [80] ‘Environmental Impact’ was calculated at GBP 1.17 per 170 mL application. With an average of 1.95 applications per hectare, this equates to a hidden cost of GBP 2.28.
5. Discussion
5.1. Analysis
This study has shone light on an otherwise elusive species, P. auritus, and its hidden pest control contribution to apple orchards in South-East England. This study’s findings emphasise the significant impact bats have on insect pest populations as identified by Maine and Boyles [57] and Federico et al. [54]. Although other studies also found moderate impacts [53,55,56,58], the results were closest to the findings of those two papers. Direct comparisons between studies is difficult due to variations in methods and study context (i.e., geographical location, study species, crop type). Outside of these two studies the literature estimates drop in significance with one study estimating a crop loss reduction of just 9–23% [81]. To deeper analyse how these discrepancies can occur, the two most methodologically similar studies will be compared below.
The methods of the current study drew from two similar North American studies of cotton pest regulation services by bats in Texas, USA [53,54] but yields rather different estimates. Both studies shared many of the same researchers but followed different methods of valuation. Cleveland et al. [53] employed an avoided-costs method based in the literature, estimating the pest consumption of a single female bat and then scaling this up whereas Federico et al. [54] opted for a population mathematical model, recording the pest consumption as a direct response of bat population dynamics and again using an avoided-costs method to calculate the financial value. For comparison the results of these two studies were converted into 2021 GBP to account for inflation and to a per hectare estimate. Although comparisons have been drawn between the current study and Cleveland et al. [53] percentage reduction in crop loss, a monetary value of just GBP 186.97/hectare was calculated in the Cleveland study. In further contrast, Federico et al. [54] estimated an avoided-cost of GBP 81.28/hectare. Unlike its predecessor, the Federico et al. [54] study omitted the ‘hidden’ social and ecological costs of pesticide use, however this does not account for the full difference in estimates. This omission not only exposes the subjective nature of monetary valuation but represents a neoclassical view concerned only with the direct costs to the individual.
The scaling up of the behaviour of an individual to entire populations also depends on some potentially unrealistic assumptions. Unlike the Cleveland et al. [53] study (amongst others, e.g., Taylor et al. [81]), an attempt was not made to extrapolate the estimated impact of a single bat to provide an estimate for the wider population (in the case of Cleveland et al. [53], 1.5 million individuals). Scaling to such extents comes with a degree of uncertainty in national population estimates, spatial variation and population-dependent effects. On a UK basis, estimates of bat populations are often contradictory of each other, possibly owing to the secretive life habitats of nocturnal species, making accurate estimates for entire populations difficult. Additional uncertainties arise from potential population-density effects such as competition for resources [82] which could lower the pest consumption of an individual. It is however known that bats are typically sociable animals and so it is likely that more than one individual would be foraging throughout the same orchard or even multiple given P. auritus being known to forage within 0.5–1 km of their roosts [44]. Entwistle et al. [63] found through mark and recapture techniques, an average P. auritus colony size of 30–50 individuals. The composite impact of this many bats in a single area could yield substantial positive results for crop protection however more research in a local context is required first before such a calculation can be reasonably attempted.
This study was not without its own limitations. The main limitation to the methods was restrictions to primary data collection due to the pandemic lockdown of 2021 and resulting dependence on the availability of secondary data which at times was very limited or unavailable. The use of secondary data also comes with some questions of validity as methods are not standardised between studies and in many cases, studies from other geographical areas are not comparable with a UK specific context [8]. Similarly, some of the data used in the model may not reflect recent population dynamics in the face of global environmental change. We chose to use older data within the literature rather than model the complexities of global change on populations. Particular difficulty came in the sourcing of financial data, with cost of active insecticide substances varying so greatly between online suppliers. The recalculation of this study with locally specific primary data could further validate the findings. Although daily temperature accounts for a significant proportion of the variation, the landscape of the orchard will also account for some variation such as edge effects or understory vegetation cover [83] that is likely not accounted for in the model.
As discussed, debate surrounds the presentation of ‘value’. The results of the model were attempted to be presented pluralistically, echoing concepts derived in ecological economics. Opposing values representing the same ecosystem service often act in contradiction with one another and for this reason are more representative than a single metric [24]. The same is true of the output values of this study. Percentage crop loss reduction was calculated between simulations (no bat, with bat) with the same parameters and so a single percentage could yield a wide array of gross apples lost and subsequently, economic avoided cost values. Hence, different expressions of value favour different applications and a just evaluation can only be delivered through pluralism.
Financial valuation of ecosystem services does have its merits in the current economic system in that it makes the scale of the service more applicable to the orchard manager as a business owner. However, assigning a financial value to a species deducts from the intrinsic value of nature and implies that for such a value, the service could be replaced with a technological alternative. This line of thought is inherently damaging to the conservation of all species where the natural end-point is to only conserve those with instrumental value to humans. It could be argued that a fine balance between conserving species for the intrinsic or instrumental value can be achieved, yet the contradictions between these two opposed values deems this balance not possible and under the current competitive economic system will lead to the instrumental value dominating the discourse. Although the same could be said to some degree for the core concept of ecosystem services, in that it is an anthropocentric perspective on nature; pluralism and a biophysical dominated valuation represents a far less utilitarian position.
5.2. Integrated Pest Management
Insect and bird exclusion netting has been suggested as an alternative to insecticide use by Sauphanor et al. [84]. Whilst netting does divert away from pesticide use, the Sauphanor study fails to account for the possible negative impacts on apple yield through the exclusion of pollinators as apples are generally self-incompatible [26]. Netting would also impact the ability for some bat species to glean prey from vegetation, as P. auritus has been observed doing. Likewise, netting would reduce opportunities for avian predators to take pest larvae (as identified in the literature) and for nesting in the trees.
Alternatively, through orchard field studies, Glen and Milsom [70] reported on the estimated reductions in C. pomonella larvae through various means, the results of which formed the larvae mortality parameter for the model. It was found that Blue (Cyanistes caeruleus) and Great Tits (Parus major) reduced larvae numbers on apple trees by 47%. Contrastingly, Solomon et al. [31] and Solomon and Glen [85] found considerably larger reductions of between 94.7 and 95%. Reduction in C. pomonella by these species of birds occurs through regulation of the larval and pupae stages and so are highly unlikely to come into competition with bat species but rather act as a composite regulator [56], enhancing the overall crop protection service.
P. auritus and C. pomonella are both widespread throughout the UK, only being absent from some of the islands surrounding Scotland, meaning it is possible that this predator-prey interaction occurs across the British Isles. Although the wide array of environmental conditions across the UK indicate that the value and scale of this service likely differ between regions and possibly even on a sub-regional level. Assuming the findings of this model accurately represent the impact of bats in apple orchard pest regulation, then it is curious that the services that orchard managers gain from bat populations goes so under noticed by both individuals [52] and key organisations/governing bodies (i.e., ADHB and DEFRA). Focusing specifically on pesticides, Olimpi and Philpott [86] found that on farms where pesticide use was less frequent, greater prey biomass correlated with a greater amount of bat activity. These findings hint at a worrying cycle of increased pesticide use reducing prey availability for bats, leading to a decrease in local bat presence, which when combined with the findings of the study model, would see an increase in C. pomonella numbers and a subsequent increased need for pesticide use.
Pest regulation services by P. auritus present an uncommon win-win situation between human economic goals and nature conservation [87]; the provision of habitat by orchard managers in turn supports bat populations which then benefits the farmer through avoided crop loss. Whilst many ecosystem services benefiting humans on a society level have required Payment for Ecosystem Service schemes (PES), usually funded through state subsidies to reward stewardship [88] this ecosystem service directly rewards apple orchard managers on an individual level for their stewardship. There is no need to create a market for this service, thus likely avoiding the potential for commodification. Additionally, orchard managers benefit from natural pest regulation as they do not have to be reliant on trap feedback loops in which moths are feeding while traps are filling, creating a lag.
5.3. Wider Implications
The range of economic values calculated from this model correspond to the regulation of just one crop pest species and so is not representative of the worth of a P. auritus individual. Along with the regulation of numerous other insect species, P. auritus may have value in nutrient cycling, cultural significance or intrinsic value [5]. It is also true that P. auritus consumes natural arthropod predators of C. pomonella, though moths are the largest proportion of the P. auritus diet while other natural predators are a lower proportion of their diet [64,89]. Surface valuation of just one service provided by a species belies the complex cycles and ecosystems behind the individual. Despite attempting to account for the hidden/social costs of avoided pesticide use it is possible that yet more, immeasurable impacts could be prevented through the pest regulation service. Given that chlorantraniliprole is recorded as being highly toxic to water invertebrates and persistent in the ecosystem it is possible that this could have further negative impacts on the aquatic ecosystem. Devastation of primary consumer trophic level species such as Daphnia spp. could lead to population decline in species which feed upon these invertebrates through unavailability of food, eventually reducing populations up the ecosystem to top predators such as fish. Fish population decline reduces opportunities for local fishing or possibly even have negative impacts on tourism particularly if this leads to a decline in characteristic species (for example kingfishers or otters). Similarly significant loss of apple crop yield could have negative impacts in the community through job reductions and possibly even loss of community cultural identity particularly in areas where apple production is characteristic of the community as is the case in many rural areas of Southern England. Apple production has a long history in the UK and is arguably part of its cultural heritage [90]. Whilst humans may indirectly financially gain from the presence of P. auritus, there is likely much more to lose than this monetary value if P. auritus were lost from the UK.
The high C. pomonella reproduction sensitivity scenario presented in Section 4.2 provides a glimpse into a possible future as the conditions for reproduction are linked with ambient temperature. In line with global warming and temperatures rising further, it could be assumed from the literature that conditions in South-East England will become more favourable for C. pomonella reproduction, thus increasing the strain on apple orchards through increases in annual populations. In this future situation, the pest regulation services of a single P. auritus is likely to have a greater impact on the larger moth population and therefore are likely to be more valued by future generations. Coupled with potential increases in pesticide resistance in C. pomonella emphasises the importance of conserving these bats for future generations.
6. Conclusions
The model developed in this study found that P. auritus provide significant pest control services of C. pomonella in English apple orchards. Through 1000 iterations of the model, a reference reduction in crop loss of between 81.06–83.68% per hectare was observed, which, using an avoided costs method corresponded to saving the orchard manager an annual GBP 308.70–606.06 per hectare (2021 GDP). The financial valuation included not only the private costs but also the social and ecological costs of pesticide applications which were avoided in all study scenarios. P. auritus pest regulation has serious potential for integrated pest management. Nevertheless, it is likely that continued use of pesticides may inhibit the capabilities of the bat, along with other natural enemies of C. pomonella. To utilise the full potential of P. auritus in orchards, historic roosts must be protected, new suitable roosting habitat be provided and the reduction or altogether stopping of pesticide use.
The decline or loss of P. auritus from orchards has potential to indirectly impact far beyond this monetary value with potential further impacts into other sectors (for example, fishing sector) or the wider community and therefore it is in the direct interest of farmers and land managers to provide habitats for the conservation of bat populations. Due to the far-reaching nature and complexity of ecological systems, the study emphasises the need for a pluralistic approach to ecosystem service valuation in line with ecological economic theory. No single metric can fully represent the antagonistic forces of an ecosystem. The results of this study were represented through various alternative and at times contradictory metrics to better aid decision making, policy and the conservation of natural systems.
In the context of English apple orchards, field data from apple orchards in the South-East region would be required to test the validity of the model findings with a particular research focus on estimating actual population densities of C. pomonella and P. auritus as this accounts for much variation. It is possible that the findings of this model could form just one section of an integrated multi-faceted ecosystem service evaluation for Plecotus auritus in the UK. To obtain a more diverse and multi-value outlook for this pest regulation service, further study could also look at qualitative impacts of the service through interviews and focus groups. The potential impacts of loss of this ecosystem service on the cultural identity of apple producing communities could be evaluated in this manner. In a broader sense, the model created for this study has particular potential to be applied to the impact of the other three UK bat species known to consume C. pomonella. It also has potential to be expanded to other bat and insect crop pest dynamics for a range of other seasonal crops. As two thirds of bat species globally are insectivorous [5], the potential for such research is compelling.
Author Contributions
The manuscript and the model were created and written by the lead author F.M. The secondary author J.A. provided guidance on writing and modelling and contributed to the discussion of ecosystem services and ecological economics. All authors have read and agreed to the published version of the manuscript.
Funding
This manuscript and the study within were created independent of funding sources.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Not applicable.
Conflicts of Interest
We declare no conflict of interest.
Appendix A. Sensitivity Analysis Results

{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table A1.
Full table of results for the step-by-step analysis. Each parameter range was divided into ten even increments. The output is the number of apples lost to C. pomonella larvae.
Table A1.
Full table of results for the step-by-step analysis. Each parameter range was divided into ten even increments. The output is the number of apples lost to C. pomonella larvae.
| Parameter | Parameter Value | Cumulative Crop Loss (Apples Infested) |
|---|---|---|
| Adult lifespan (days) | 16.100 17.389 18.678 19.967 21.256 22.544 23.833 25.122 26.411 27.700 | 434.61 409.87 387.79 367.98 350.09 333.86 319.06 305.53 293.09 281.63 |
| Number of eggs per female | 161.8 170.7 179.6 188.5 197.4 206.3 215.2 224.1 233.0 241.9 | 296.20 312.49 328.78 345.07 361.37 377.66 393.95 410.24 426.54 442.83 |
| Egg death rate (proportion of eggs dying per day) | 0.12 0.20 0.28 0.37 0.45 0.53 0.61 0.70 0.78 0.86 | 481.74 441.59 407.60 378.45 353.19 331.08 311.58 294.24 278.73 264.77 |
| Gestation (days) | 7.00 7.33 7.67 8.00 8.33 8.67 9.00 9.33 9.67 10.00 | 342.00 341.94 341.89 341.85 341.80 341.76 341.71 341.66 341.60 341.55 |
| Larvae lifespan (days) | 13.00 16.79 20.58 24.37 28.16 31.95 35.74 39.53 43.32 47.11 | 343.12 342.90 342.62 342.20 341.59 340.72 339.54 338.02 336.17 333.98 |
| Egg viability (proportion of eggs hatching) | 0.7170 0.7306 0.7441 0.7577 0.7712 0.7848 0.7983 0.8119 0.8255 0.8390 | 330.84 333.37 335.84 338.25 340.62 342.93 345.20 347.41 349.59 351.72 |
| Larvae death rate | 0.241000 0.317111 0.393222 0.469333 0.545444 0.621556 0.697667 0.773778 0.849889 0.926000 | 521.14 492.71 467.05 443.80 422.66 403.37 385.70 369.48 354.54 340.73 |
References
- National Statistics: Latest Horticulture Statistics; Department for Environment, Food & Rural Affairs: London, UK, 2020.
- Trade Map—Trade Statistics for International Business Development; International Trade Centre: Geneva, Switzerland, 2021.
- Burrough, A.E.; Oines, C.M.; Oram, S.P.; Robertson, H.J. Traditional Orchard Project in England—The Creation of an Inventory to Support the UK Habitat Action Plan. In Natural England Commissioned Reports; Natural England: Worcester, UK, 2010. [Google Scholar]
- Gimber, M. Traditional Orchard Decline; People’s Trust for Endangered Species: London, UK, 2021. [Google Scholar]
- Kunz, T.H.; De Torrez, E.B.; Bauer, D.; Lobova, T.; Fleming, T.H. Ecosystem services provided by bats. Ann. N. Y. Acad. Sci. 2011, 1223, 1–38. [Google Scholar] [CrossRef] [PubMed]
- Fisher, B.; Turner, K.; Zylstra, M.; Brouwer, R.; de Groot, R.; Farber, S.; Ferraro, P.; Green, R.; Hadley, D.; Harlow, J.; et al. Ecosystem services and economic theory: Integration for policy-relevant research. Ecol. Appl. 2008, 18, 2050–2067. [Google Scholar] [CrossRef] [PubMed]
- Norgaard, R.B. The case for methodological pluralism. Ecol. Econ. 1989, 1, 37–57. [Google Scholar] [CrossRef]
- Spash, C.L.; Aslaksen, I. Re-establishing an ecological discourse in the policy debate over how to value ecosystems and biodiversity. J. Environ. Manag. 2015, 159, 245–253. [Google Scholar] [CrossRef] [PubMed]
- Daily, G.C. (Ed.) Nature’s Services: Societal Dependence on Natural Ecosystems; Island Press: Washington, DC, USA, 1997. [Google Scholar]
- Study of Critical Environmental Problems (SCEP). Man’s Impact on the Global Environment; MIT Press: Cambridge, MA, USA, 1970. [Google Scholar]
- Holdren, J.P.; Ehrlich, P.R. Human population and the global environment. Am. Sci. 1974, 62, 282–292. [Google Scholar] [PubMed]
- Westman, W.E. How much are nature’s services worth? Science 1977, 197, 960–964. [Google Scholar] [CrossRef]
- Spash, C.L. The New Environmental Pragmatists, Pluralism and Sustainability. Environ. Values 2009, 18, 253–256. [Google Scholar] [CrossRef]
- Hanley, N.; Shogren, J.F.; White, B. Introduction to Environmental Economics; Oxford University Press: Oxford, UK, 2001. [Google Scholar]
- Mols, C.M.M.; Visser, M.E. Great tits can reduce caterpillar damage in apple orchards. J. Appl. Ecol. 2002, 39, 888–899. [Google Scholar] [CrossRef]
- Glen, D.M. The effects of predators on the eggs of codling moth Cydia pomonella, in a cider-apple orchard in south-west England. Ann. Appl. Biol. 1975, 80, 115–119. [Google Scholar] [CrossRef]
- Cross, J.; Fountain, M.; Markó, V.; Nagy, C. Arthropod ecosystem services in apple orchards and their economic benefits. Ecol. Entomol. 2015, 40, 82–96. [Google Scholar] [CrossRef]
- Carreck, N.; Williams, I. The economic value of bees in the UK. Bee World 1998, 79, 115–123. [Google Scholar] [CrossRef]
- Zhang, D.; Wang, C.; Li, X.; Yang, X.; Zhao, L.; Liu, L.; Zhu, C.; Li, R. Linking plant ecological stoichiometry with soil nutrient and bacterial communities in apple orchards. Appl. Soil Ecol. 2017, 126, 1–10. [Google Scholar] [CrossRef]
- Helm, D. Natural Capital: Valuing the Planet; Yale University Press: New Haven, CT, USA; London, UK, 2016. [Google Scholar]
- Gómez-Baggethun, E.; Ruiz-Pérez, M. Economic valuation and the commodification of ecosystem services. Prog. Phys. Geogr. Earth Environ. 2011, 35, 613–628. [Google Scholar] [CrossRef]
- de Groot, R. Investing in Natural Capital: The Ecological Economics Approach To Sustainability; Jansson, A., Hammer, M., Folke, C., Costanza, R., Eds.; Island Press: Washington, DC, USA, 1994; ISBN 978-1-55963-316-1. [Google Scholar]
- Gowdy, J.M. Terms and concepts in ecological economics. Wildl. Soc. Bull. 2000, 28, 26–33. [Google Scholar]
- Gomez-Baggethun, E.; Martin-Lopez, B. Ecological economics perspectives on ecosystem services valuation. In Handbook of ecological economics; Edward Elgar Publishing: Cheltenham, UK, 2015; pp. 260–282. [Google Scholar] [CrossRef]
- Naranjo, S.E.; Ellsworth, P.C.; Frisvold, G.B. Economic Value of Biological Control in Integrated Pest Management of Managed Plant Systems. Annu. Rev. Entomol. 2015, 60, 621–645. [Google Scholar] [CrossRef]
- Garratt, M.P.D.; Truslove, L.; Coston, D.; Evans, R.; Moss, E.; Dodson, C.; Jenner, N.; Biesmeijer, J.; Potts, S. Pollination deficits in UK apple orchards. J. Pollinat. Ecol. 2013, 12, 9–14. [Google Scholar] [CrossRef]
- EUR Exchange Rates. Bank of England. Available online: https://0-www-bankofengland-co-uk.brum.beds.ac.uk/boeapps/database/Rates.asp?TD=13&TM=Sep&TY=2021&into=EUR&rateview=D (accessed on 13 September 2021).
- Mace, A.; Ridley, L.; Parrish, G.; MacArthur, R.; Rainford, J.; Garthwaite, D. Pesticide Usage Survey Report 286: Orchards in the United Kingdom 2018; Land Use and Sustainability Team, Fera Science Ltd.: York, UK, 2018. [Google Scholar]
- Jackson, D.M. Codling Moth1 Egg Distribution on Unmanaged Apple Trees2. Ann. Entomol. Soc. Am. 1979, 72, 361–368. [Google Scholar] [CrossRef]
- AHDB and Department for Environment, Food and Rural Affairs. Apple Best Practice Guide: Codling Moth (Cydia pomonella L.). 2021. Available online: https://apples.ahdb.org.uk/codling-moth/ (accessed on 10 July 2021).
- Solomon, M.E.; Glen, D.M.; Kendall, D.A.; Milsom, N.F. Predation of Overwintering Larvae of Codling Moth (Cydia pomonella (L.)) by Birds. J. Appl. Ecol. 1976, 13, 341. [Google Scholar] [CrossRef]
- Witzgall, P.; Stelinski, L.; Gut, L.; Thomson, D. Codling Moth Management and Chemical Ecology. Annu. Rev. Entomol. 2008, 53, 503–522. [Google Scholar] [CrossRef]
- Judd, G.J.R.; Gardiner, M.G.T.; Thomson, D.R. Control of codling moth in organically-managed apple orchards by combining pheromone-mediated mating disruption, post-harvest fruit removal and tree banding. Entomol. Exp. Appl. 1997, 83, 137–146. [Google Scholar] [CrossRef]
- Dewar, A.; Foster, S. Overuse of Pyrethroids may be implicated in the Recent BYDV Epidemics in Cereals. Outlooks Pest Manag. 2017, 28, 7–12. [Google Scholar] [CrossRef]
- Voudouris, C.C.; Sauphanor, B.; Franck, P.; Reyes, M.; Mamuris, Z.; Tsitsipis, J.A.; Vontas, J.; Margaritopoulos, J.T. Insecticide resistance status of the codling moth Cydia pomonella (Lepidoptera: Tortricidae) from Greece. Pestic. Biochem. Physiol. 2011, 100, 229–238. [Google Scholar] [CrossRef]
- Franck, P.; Reyes, M.; Olivares, J.; Sauphanor, B. Genetic architecture in codling moth populations: Comparison between microsatellite and insecticide resistance markers. Mol. Ecol. 2007, 16, 3554–3564. [Google Scholar] [CrossRef] [PubMed]
- Relyea, R.A. The impact of insecticides and herbicides on the biodiversity and productivity of aquatic communities. Ecol. Appl. 2005, 15, 618–627. [Google Scholar] [CrossRef]
- Szöcs, E.; Brinke, M.; Karaoglan, B.; Schäfer, R.B. Large Scale Risks from Agricultural Pesticides in Small Streams. Environ. Sci. Technol. 2017, 51, 7378–7385. [Google Scholar] [CrossRef]
- Flexner, J.; Lighthart, B.; Croft, B. The effects of microbial pesticides on non-target, beneficial arthropods. Agric. Ecosyst. Environ. 1986, 16, 203–254. [Google Scholar] [CrossRef]
- Koureas, M.; Tsakalof, A.; Tsatsakis, A.; Hadjichristodoulou, C. Systemic Review of Biomonitoring Studies to Determine the Association between Exposure to Organophosphorus and Pyrethroid Insecticides and Human Health Outcomes. Toxicol. Lett. 2012, 210, 155–168. [Google Scholar] [CrossRef]
- Whitehorn, P.R.; O’Connor, S.; Wackers, F.L.; Goulson, D. Neonicotinoid Pesticide Reduces Bumble Bee Colony Growth and Queen Production. Science 2012, 336, 351–352. [Google Scholar] [CrossRef]
- Woodcock, B.A.; Bullock, J.M.; Shore, R.F.; Heard, M.S.; Pereira, M.G.; Redhead, J.W.; Ridding, L.; Dean, H.; Sleep, D.; Henrys, P.A.; et al. Country-specific effects of neonicotinoid pesticides on honey bees and wild bees. Science 2017, 356, 1393–1395. [Google Scholar] [CrossRef]
- McKerchar, M.; Potts, S.; Fountain, M.; Garratt, M.; Westbury, D. The potential for wildflower interventions to enhance natural enemies and pollinators in commercial apple orchards is limited by other management practices. Agric. Ecosyst. Environ. 2020, 301, 107034. [Google Scholar] [CrossRef]
- Altringham, J.D. British Bats; Harper Collins Publishers: New York, NY, USA, 2003. [Google Scholar]
- Verboom, B.; Huitema, H. The importance of linear landscape elements for the pipistrelle Pipistrellus pipistrellus and the serotine bat Eptesicus serotinus. Landsc. Ecol. 1997, 12, 117–125. [Google Scholar] [CrossRef]
- Mikula, P.; Čmoková, A. Lepidopterans in the summer diet of Eptesicus serotinus in Central Bohemia. Vespertilio 2012, 16, 197–201. [Google Scholar]
- Jay, M.; Roincé, C.B.; Ricard, J.M.; Garcin, A.; Mandrin, J.F.; Lavigne, C.; Bouvier, J.C.; Tupinier, Y.; Puechmaille, S. Biodiversité fonctionelle en verger de pommier: Les chauves-souris consomment-elles des ravageurs? Infos.-Ctifl. 2012, 286, 28–34. [Google Scholar]
- Andriollo, T.; Michaux, J.R.; Ruedi, M. Food for everyone: Differential feeding habits of cryptic bat species inferred from DNA metabarcoding. Mol. Ecol. 2021, 30, 4584–4600. [Google Scholar] [CrossRef]
- Baroja, U.; Garin, I.; Aihartza, J.; Arrizabalaga-Escudero, A.; Vallejo, N.; Aldasoro, M.; Goiti, U. Pest consumption in a vineyard system by the lesser horseshoe bat (Rhinolophus hipposideros). PLoS ONE 2019, 14, e0219265. [Google Scholar] [CrossRef] [PubMed]
- Boyd, I.L.; Stebbings, R.E. Population Changes of Brown Long-Eared Bats (Plecotus auritus) in Bat Boxes at Thetford Forest. J. Appl. Ecol. 1989, 26, 101–112. [Google Scholar] [CrossRef]
- UK Biodiversity Indicators 2020; Department for Environment, Food & Rural Affairs: London, UK, 2020.
- Martínez-Sastre, R.; García, D.; Miñarro, M.; Martín-López, B. Farmers’ perceptions and knowledge of natural enemies as providers of biological control in cider apple orchards. J. Environ. Manag. 2020, 266, 110589. [Google Scholar] [CrossRef]
- Cleveland, C.J.; Betke, M.; Federico, P.; Frank, J.D.; Hallam, T.G.; Horn, J.; López, J.D.; McCracken, G.F.; Medellín, R.A.; Moreno-Valdez, A.; et al. Economic value of the pest control service provided by Brazilian free-tailed bats in south-central Texas. Front. Ecol. Environ. 2006, 4, 238–243. [Google Scholar] [CrossRef]
- Federico, P.; Hallam, T.G.; McCracken, G.; Purucker, S.T.; Grant, W.E.; Correa-Sandoval, A.N.; Westbrook, J.K.; Medellín, R.A.; Cleveland, C.J.; Sansone, C.G.; et al. Brazilian free-tailed bats as insect pest regulators in transgenic and conventional cotton crops. Ecol. Appl. 2008, 18, 826–837. [Google Scholar] [CrossRef]
- Boyles, J.G.; Cryan, P.M.; McCracken, G.F.; Kunz, T.H. Economic Importance of Bats in Agriculture. Science 2011, 332, 41–42. [Google Scholar] [CrossRef]
- Maas, B.; Clough, Y.; Tscharntke, T. Bats and birds increase crop yield in tropical agroforestry landscapes. Ecol. Lett. 2013, 16, 1480–1487. [Google Scholar] [CrossRef]
- Maine, J.J.; Boyles, J.G. Bats initiate vital agroecological interactions in corn. Proc. Natl. Acad. Sci. USA 2015, 112, 12438–12443. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Puig-Montserrat, X.; Torre, I.; López-Baucells, A.; Guerrieri, E.; Monti, M.M.; Ràfols-García, R.; Ferrer, X.; Gisbert, D.; Flaquer, C. Pest control service provided by bats in Mediterranean rice paddies: Linking agroecosystems structure to ecological functions. Mamm. Biol. 2015, 80, 237–245. [Google Scholar] [CrossRef]
- McLean, J.A.; Speakman, J.R. Energy budgets of lactating and non-reproductive Brown Long-Eared Bats (Plecotus auritus) suggest females use compensation in lactation. Funct. Ecol. 2002, 13, 360–372. [Google Scholar] [CrossRef]
- Speakman, J.R.; Racey, P.A. The Energetics of Pregnancy and Lactation in the Brown Long-Eared Bat, Plecotus Auritus. In Recent Advances in the Study of Bats; Racey, P.A., Rayner, J.M.V., Eds.; Cambridge University Press: Fenton, MB, USA, 1987; pp. 367–393. [Google Scholar]
- Margaritopoulos, J.T.; Voudouris, C.C.; Olivares, J.; Sauphanor, B.; Mamuris, Z.; Tsitsipis, J.A.; Franck, P. Dispersal ability in codling moth: Mark-release-recapture experiments and kinship analysis. Agric. For. Entomol. 2012, 14, 399–407. [Google Scholar] [CrossRef]
- Harris, S.; Yalden, D.W. Mammals of the British Isles Handbook; The Mammal Society: London, UK, 2008. [Google Scholar]
- Entwistle, A.C.; Racey, P.A.; Speakman, J.R. Social and Population Structure of a Gleaning Bat, Plecotus Auritus. J. Zool. 2000, 252, 11–17. [Google Scholar] [CrossRef]
- Shiel, C.; McAney, C.; Fairley, J. Analysis of the Diet of Natterer’s Bat Myotis Nattereri and the Common Long-eared Bat Plecotus Auritus in the West of Ireland. J. Zool. 1991, 223, 299–305. [Google Scholar] [CrossRef]
- Andriollo, T.; Gillet, F.; Michaux, J.R.; Ruedi, M. The menu varies with metabarcoding practices: A case study with the bat Plecotus auritus. PLoS ONE 2019, 14, e0219135. [Google Scholar] [CrossRef]
- Glen, D.M.; Milsom, N.F.; Wiltshire, C.W. The Effect of Predation by Blue-Tits (Parus caeruleus) on the Sex-Ratio of Codling Moth (Cydia pomonella). J. Appl. Ecol. 1981, 18, 133. [Google Scholar] [CrossRef]
- Plantwise Knowledge Bank: Codling Moth, Cydia Pomonella; Centre for Agriculture and Bioscience (CAB) International: Oxfordshire, UK, 2021.
- Graf, B.; Höhn, H.; Höpli, H.; Kuske, S. Predicting the phenology of codling moth, Cydia pomonella, for sustainable pest management in Swiss apple orchards. Entomol. Exp. Appl. 2018, 166, 618–627. [Google Scholar] [CrossRef]
- Kuyulu, A.; Genc, H.B.; Codling Moth, L.R. Cydia pomonella (L.) (Lepidoptera: Tortricidae) on Its Natural Host “Green Immature Apple” Malus domestica (Borkh; Rosaceae); Türk Tarim Ve Doga Bilimleri Derg: Rosales, Philippines, 2019; Volume 6. [Google Scholar]
- Glen, D.M.; Milsom, N.F. Survival of mature larvae of codling moth (Cydia pomonella) on apple trees and ground. Ann. Appl. Biol. 1978, 90, 133–146. [Google Scholar] [CrossRef]
- Glen, D.M. Predation of Codling Moth Eggs, Cydia pomonella, the Predators Responsible and Their Alternative Prey. J. Appl. Ecol. 1977, 14, 445–456. [Google Scholar] [CrossRef]
- Adams, C.G.; Schenker, J.H.; McGhee, P.S.; Gut, L.J.; Brunner, J.F.; Miller, J.R. Maximizing Information Yield from Pheromone-Baited Monitoring Traps: Estimating Plume Reach, Trapping Radius, and Absolute Density of Cydia pomonella (Lepidoptera: Tortricidae) in Michigan Apple. J. Econ. Entomol. 2017, 110, 305–318. [Google Scholar] [CrossRef] [PubMed]
- Alford, D.V.; Carden, P.W.; Dennis, E.B.; Gould, H.J.; Vernon, J.D.R. Monitoring codling and tortrix moths in United Kingdom apple orchards using pheromone traps. Ann. Appl. Biol. 1979, 91, 165–178. [Google Scholar] [CrossRef]
- Glenn, P.A. Codling-moth Investigations of the State Entomologist’s Office. Ill. Nat. Hist. Surv. Bull. 1915, 14, 219–289. [Google Scholar] [CrossRef]
- Sæthre, M.-G.; Hofsvang, T. Effect of Temperature on Oviposition Behavior, Fecundity, and Fertility in Two Northern European Populations of the Codling Moth (Lepidoptera: Tortricidae). Environ. Entomol. 2002, 31, 804–815. [Google Scholar] [CrossRef]
- Apples & Orchards. The National Association of Cider Makers. Available online: https://cideruk.com/orchards-apples/ (accessed on 14 September 2022).
- Waitrose & Partners. Available online: https://www.waitrose.com/ecom/products/essential-loose-braeburn-apples/088640-45525-45526 (accessed on 14 June 2021).
- Pesticide Fact Sheet: Chlorantraniliprole; United States Environmental Protection Agency: Washington, DC, USA, 2008.
- Leach, A.; Mumford, J. Pesticide Environmental Accounting: A method for assessing the external costs of individual pesticide applications. Environ. Pollut. 2008, 151, 139–147. [Google Scholar] [CrossRef] [PubMed]
- Kovach, J.; Petzoldt, C.; Degnil, J.; Tette, J. A Method to Measure the Environmental Impacts of Pesticides. N. Y. Food Life Sci. Bull. 1992, 139, 1–8. [Google Scholar]
- Taylor, P.J.; Grass, I.; Alberts, A.J.; Joubert, E.; Tscharntke, T. Economic value of bat predation services—A review and new estimates from macadamia orchards. Ecosyst. Serv. 2018, 30, 372–381. [Google Scholar] [CrossRef]
- Corcoran, A.J.; Conner, W.E. Bats jamming bats: Food competition through sonar interference. Science 2014, 346, 745–747. [Google Scholar] [CrossRef]
- Ricci, B.; Franck, P.; Bouvier, J.-C.; Casado, D.; Lavigne, C. Effects of hedgerow characteristics on intra-orchard distribution of larval codling moth. Agric. Ecosyst. Environ. 2011, 140, 395–400. [Google Scholar] [CrossRef]
- Sauphanor, B.; Severac, G.; Maugin, S.; Toubon, J.F.; Capowiez, Y. Exclusion netting may alter reproduction of the codling moth (Cydia pomonella) and prevent associated fruit damage to apple orchards. Entomol. Exp. et Appl. 2012, 145, 134–142. [Google Scholar] [CrossRef]
- Solomon, M.E.; Glen, D.M. Prey Density and Rates of Predation by Tits Parus Spp. on Larvae of Codling Moth Cydia pomonella (L.) under Bark. J. Appl. Ecol. 1979, 16, 49–59. [Google Scholar] [CrossRef]
- Olimpi, E.M.; Philpott, S.M. Agroecological farming practices promote bats. Agric. Ecosyst. Environ. 2018, 265, 282–291. [Google Scholar] [CrossRef]
- Farber, S.C.; Costanza, R.; Wilson, M.A. Economic and ecological concepts for valuing ecosystem services. Ecol. Econ. 2002, 41, 375–392. [Google Scholar] [CrossRef]
- Gómez-Baggethun, E.; Muradian, R. In markets we trust? Setting the boundaries of Market-Based Instruments in ecosystem services governance. Ecol. Econ. 2015, 117, 217–224. [Google Scholar] [CrossRef]
- de Roincé, C.B.; Lavigne, C.; Ricard, J.-M.; Franck, P.; Bouvier, J.-C.; Garcin, A.; Symondson, W.O.C. Predation by generalist predators on the codling moth versus a closely-related emerging pest the oriental fruit moth: A molecular analysis. Agric. For. Entomol. 2012, 14, 260–269. [Google Scholar] [CrossRef]
- Crosby, J. The Social and Cultural Value of the Apple and the Orchard in Victorian England. Ph.D. Thesis, University of Essex, Colchester, UK, 2020. [Google Scholar]
Figure 1.
System dynamics model representing a single C. pomonella reproductive cycle.

Figure 2.
Number of C. pomonella adults and number consumed by a single P. auritus against model time (days).
Figure 2.
Number of C. pomonella adults and number consumed by a single P. auritus against model time (days).

Figure 3.
Number of infested apples by C. pomonella larvae (i.e., crop loss) in the presence of a pregnant or non-reproductive P. auritus individual for high and low sensitivity scenarios.
Figure 3.
Number of infested apples by C. pomonella larvae (i.e., crop loss) in the presence of a pregnant or non-reproductive P. auritus individual for high and low sensitivity scenarios.

Figure 4.
Box-plot visualisation of the results of step-by-step sensitivity analysis of manipulatable model parameters.
Figure 4.
Box-plot visualisation of the results of step-by-step sensitivity analysis of manipulatable model parameters.

Table 1.
Literature sources used for data parameters in model.
| C. pomonella Parameter | Baseline Value | Range | Author(s) | Methods of Obtaining | Country of Study |
|---|---|---|---|---|---|
| Adult lifespan | 21.9 days (female) (at mean 22 °C) | 16.1–27.7 days | Graf et al. [68] | Captive study of individuals in climate chambers at six constant temperatures. | Switzerland |
| Number of eggs per female moth | 186.7 | 161.8–241.9 | Graf et al. [68] | As above. | Switzerland |
| Egg viability | 0.778 | 0.717–0.839 | Kuyulu & Genc [69] | Rearing of C. pomonella on Malus domestica cv. ‘Gala’ in laboratory conditions | Turkey |
| Egg mortality (inc. arthropod predation) | 0.49 | 0.12–0.86 | Glen [71] | Field experiment. Known number of eggs glued to known locations over regular intervals in a cider apple orchard. | South-West England |
| Gestation | 8.5 days | 7–10 days | AHDB, DEFRA [30,51] | Unknown | UK |
| Larvae mortality | 0.92 | 0.241–0.926 Kuyulu & Genc [69] | Glen & Milsom [70] | Field experiment. Apple trees kept in different conditions with some excluding ground predatory beetles or birds. | South-West England |
| Adult body mass | Males = 0.0164 g, Females = 0.02785 g | Unknown | Kuyulu & Genc [69] | As above. | Turkey |
| Larvae lifespan | 27.15 days | 13–47.1 days | Graf et al. [68] | As above. | Switzerland |
Table 2.
Summary of valuation of the pest regulation service provided by a non-reproductive and a pregnant P. auritus under varying conditions for C. pomonella reproduction. The first value provided under ‘Crop loss’ is the price of avoided crop loss in the presence of a non-reproductive P. auritus. The second value is in the presence of a pregnant individual.
Table 2.
Summary of valuation of the pest regulation service provided by a non-reproductive and a pregnant P. auritus under varying conditions for C. pomonella reproduction. The first value provided under ‘Crop loss’ is the price of avoided crop loss in the presence of a non-reproductive P. auritus. The second value is in the presence of a pregnant individual.
| Conditions for C. pomonella Reproduction | Low Reproduction | Stochastic Mode Reference | High Reproduction |
|---|---|---|---|
| Percentage crop loss reduction | 82.45–83.94% | 81.06–83.68% | 75.07–77.00% |
| Gross avoided crop loss | 868–884 apples | 1066–2718 apples | 3751–3847 apples |
| Avoided costs | |||
| Crop loss (non-reprod.) Crop loss (pregnant) | GBP 156.24 GBP 159.12 | GBP 191.88–489.24 GBP 221.58–324.90 | GBP 657.18 GBP 692.46 |
| Pesticide application | GBP 114.54 | GBP 114.54 | GBP 114.54 |
| Hidden pesticide cost | GBP 2.28 | GBP 2.28 | GBP 2.28 |
| Total | GBP 273.06–275.94 | GBP 308.70–606.06 | GBP 774.00–809.28 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Murphy, F.; Ament, J. Pluralistic Valuation of Codling Moth Regulation by Brown Long-Eared Bats in English Apple Orchards. Sustainability 2022, 14, 11966. https://0-doi-org.brum.beds.ac.uk/10.3390/su141911966
AMA Style
Murphy F, Ament J. Pluralistic Valuation of Codling Moth Regulation by Brown Long-Eared Bats in English Apple Orchards. Sustainability. 2022; 14(19):11966. https://0-doi-org.brum.beds.ac.uk/10.3390/su141911966
Chicago/Turabian StyleMurphy, Francis, and Joe Ament. 2022. "Pluralistic Valuation of Codling Moth Regulation by Brown Long-Eared Bats in English Apple Orchards" Sustainability 14, no. 19: 11966. https://0-doi-org.brum.beds.ac.uk/10.3390/su141911966
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.