
Flowering, Nutritional Status, and Content of Chloroplast Pigments in Leaves of Gladiolus hybridus L. ‘Advances Red’ after Application of Trichoderma spp.


Abstract
:1. Introduction
2. Materials and Methods
2.1. Cultivation of Plants
2.2. Inoculum
2.3. Parameters
2.4. Content of Chloroplast Pigments
2.5. Content of Macro- and Microelements
2.6. Root Colonization
2.7. Data Analysis
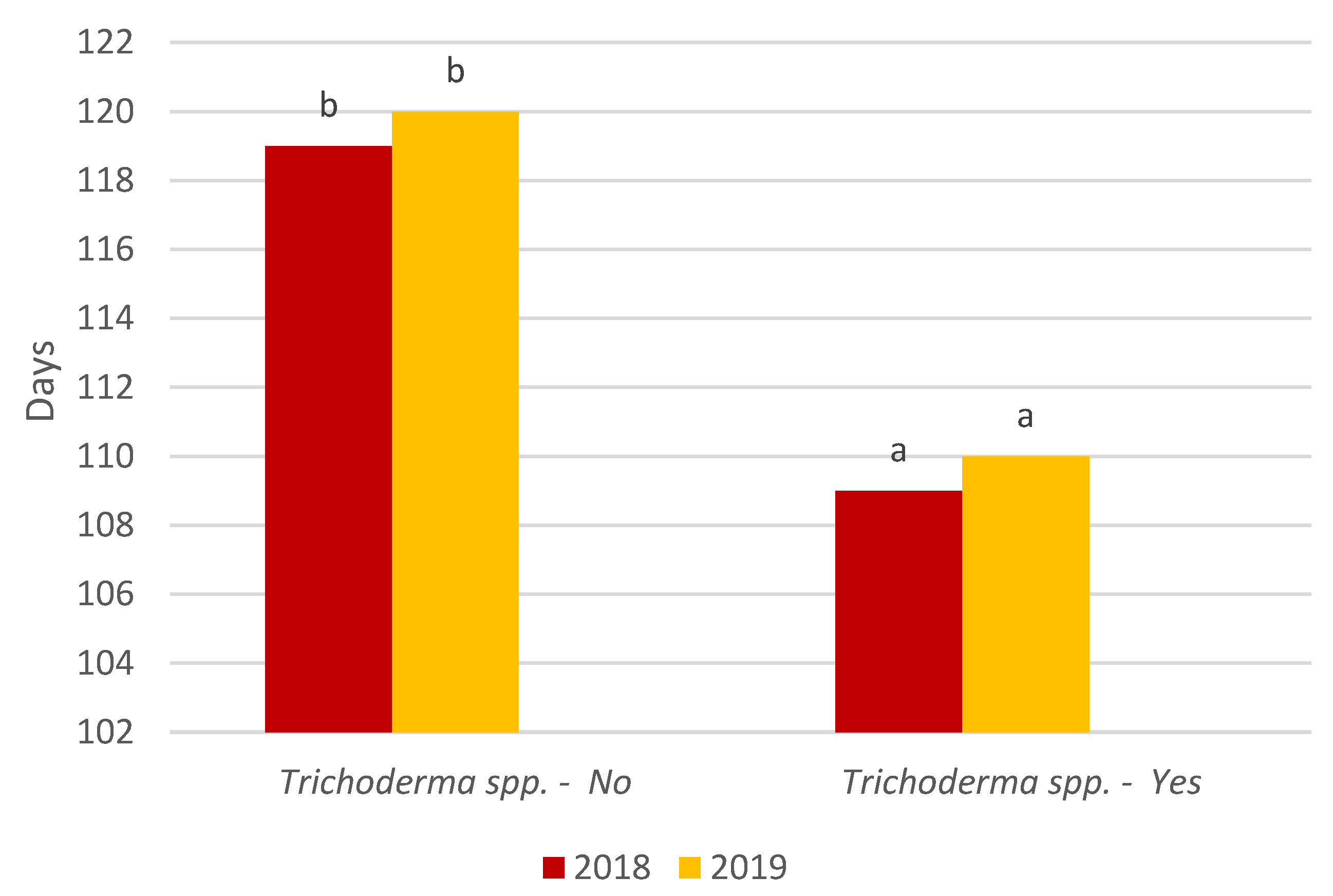
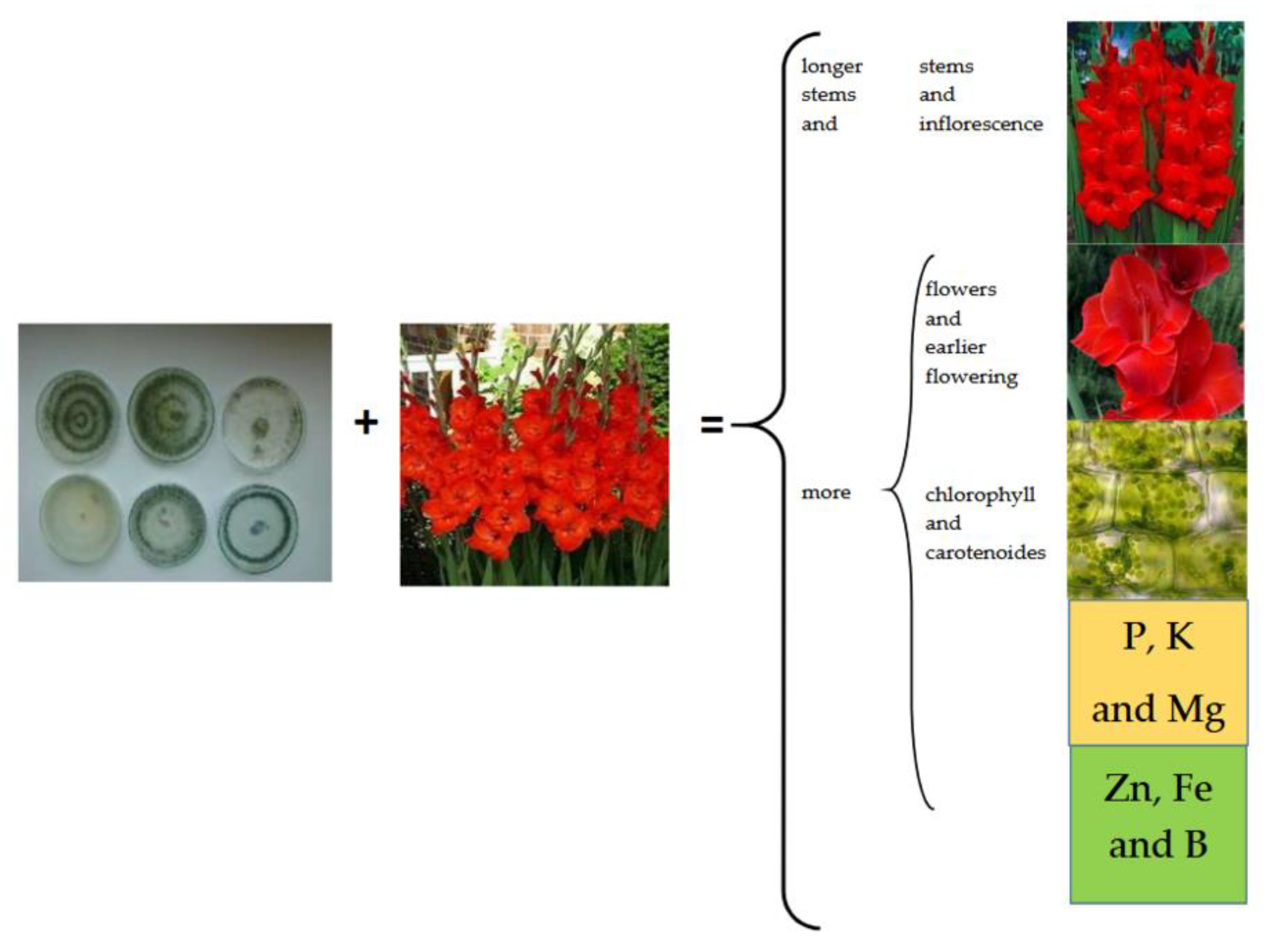
3. Results and Discussion
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Chandra, H.; Kumari, P.; Prasad, R.; Chandra Gupta, S.; Yadav, S. Antioxidant and antimicrobial activity displayed by a fungal endophyte Alternaria alternata isolated from Picrorhiza kurroa from Garhwal Himalayas, India. Biocat. Agric. Biotechnol. 2021, 33, 101955. [Google Scholar] [CrossRef]
- Hewedy, O.A.; Abdel Lateif, K.S.; Seleiman, M.F.; Shami, A.; Albarakaty, F.M.; MEl-Meihy, R. Phylogenetic diversity of Trichoderma strains and their antagonistic potential against soil-borne pathogens under stress conditions. Biology 2020, 9, 189. [Google Scholar] [CrossRef] [PubMed]
- Wojtkowiak-Gebarowska, E. Mechanizmy zwalczania fitopatogenow glebowych przez grzyby z rodzaju Trichoderma. Post. Mikrobiol. 2006, 45, 261–273. [Google Scholar]
- Benìtez, T.; Rincòn, A.M.; Limòn, M.C.; Codòn, A.C. Biocontrol mechanisms of Trichoderma strains. Int. Microbiol. 2004, 7, 249–260. [Google Scholar]
- Świerczyńska, I.; Korbas, M.; Horoszkiewicz-Janka, J.; Danielewicz, J. Antagonistyczne oddziaływanie Trichoderma viride na patogeny z rodzaju Fusarium w obecności biopreparatów (Antagonistic effect of Trichoderma viride on pathogenic fungi of the genus Fusarium in the presence of biopreparations). J. Res. Appl. Agric. Eng. 2011, 56, 157–160. [Google Scholar]
- di Lelio, I.; Coppola, M.; Comite, E.; Molisso, D.; Lorito, M.; Woo, S.L.; Pennacchio, F.; Rao, R.; Digilio, M.C. Temperature differentially influences the capacity of Trichoderma species to induce plant defense responses in Tomato against insect pests. Front. Plant Sci. 2021, 12, 678830. [Google Scholar] [CrossRef]
- Poveda, J. Trichoderma as biocontrol agent against pests: New uses for a mycoparasite. Biol. Control 2021, 159, 104634. [Google Scholar] [CrossRef]
- Kosicka, D.; Wolna-Maruwka, A.; Trzeciak, M. Aspekty stosowania Trichoderma sp. w ochronie roślin i rozkładzie materii organicznej (Aspects of the use of Trichoderma sp. in crop protection and distribution of organic matter). Kosmos 2014, 63, 635–642. [Google Scholar]
- Vey, A.; Hoagland, R.E.; But, T.M. Toxic metabolites of fungal biocontrol agents. In Fungi as Biocontrol Agents: Progress, Problems and Potential; Butt, T.M., Jackson, C., Magan, N., Eds.; CABI Publishing: Egham, UK, 2001; pp. 311–346. [Google Scholar]
- Howell, C.R. Understanding the mechanisms employed by Trichoderma virens to effect biological control of cotton diseases. Phytopathology 2006, 96, 178–180. [Google Scholar] [CrossRef] [Green Version]
- Reino, J.L.; Guerrero, R.F.; Hernández-Galán, R.; Collado, I.G. Secondary metabolites from species of the biocontrol agent Trichoderma. Phytochem. Rev. 2008, 7, 89–123. [Google Scholar] [CrossRef]
- Stewart, A.; Hill, R. Applications of Trichoderma in plant growth promotion. In Biotechnology and Biology of Trichoderma; Gupta, V.K., Schmoll, M., Herrera-Estrella, A., Upadhyay, R.S., Druzhinina, I., Tuohy, M.G., Eds.; Elsevier: Amsterdam, The Netherlands, 2014; pp. 415–428. [Google Scholar] [CrossRef]
- Zin, N.A.; Badaluddin, N.A. Biological functions of Trichoderma spp. for agriculture applications. Ann. Agric. Sci. 2020, 65, 168–178. [Google Scholar] [CrossRef]
- Hoyos-Carvajal, L.; Orduz, S.; Bissett, J. Growth stimulation in bean (Phaseolus vulgaris L.) by Trichoderma. Biol. Control 2009, 51, 409–416. [Google Scholar] [CrossRef]
- Nieto-Jacobo, M.F.; Steyaert, J.M.; Salazar-Badillo, F.B.; Nguyen, D.V.; Rostás, M.; Braithwaite, M.; de Souza, J.T.; Jimenez-Bremont, J.F.; Ohkura, M.; Stewart AMendoza-Mendoza, A. Environmental growth conditions of Trichoderma spp. affects indole acetic acid derivatives, volatile organic compounds, and plant growth promotion. Front. Plant Sci. 2017, 8, 102. [Google Scholar] [CrossRef] [Green Version]
- Contreras-Cornejo, H.A.; Macías-Rodríguez, L.; Cortés-Penagos, C.; López-Bucio, J. Trichoderma virens, a plant beneficial fungus, enhances biomass production and promotes lateral root growth through an auxin-dependent mechanism in Arabidopsis. Plant Physiol. 2009, 149, 1579–1592. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yedidia, I.; Srivastava, A.K.; Kupalnik, Y.; Chet, I. Effect of Trichoderma harzianum on microelement concentrations and increased growth of Cucumber plants. Plant Soil. 2001, 235, 235–242. [Google Scholar] [CrossRef]
- Harman, G.E.; Howell, C.R.; Viterbo, A.; Chet, I.; Lorito, M. Trichoderma species–opportunistic, avirulent plant symbionts. Nat. Rev. Microbiol. 2004, 2, 43–56. [Google Scholar] [CrossRef]
- Lorito, M.; Woo, S.; Harman, G.; Monte, E. Translational research on Trichoderma: From ‘omics to the field. Annu. Rev. Phytopathol. 2010, 48, 395–417. [Google Scholar] [CrossRef] [Green Version]
- Simkin, A.J.; Gaffe, J.; Alcaraz, J.P.; Carde, J.P.; Bramley, P.M.; Fraser, P.D.; Kuntz, M. Fibrillin influence on plastid ultrastructure and pigment content in tomato fruit. Phytochemistry 2007, 68, 1545–1556. [Google Scholar] [CrossRef]
- Langi, P.; Kiokias, S.; Varzakas, T.; Proestos, C. Carotenoids: From plants to food and feed industries. Methods Mol. Biol. 2018, 1852, 57–71. [Google Scholar]
- Meléndez-Martínez, A.J.; Mandić, A.I.; Bantis, F.; Böhm, V.; Borge, G.I.A.; Brnĉiĉ, M.; Bysted, A.; Cano, M.P.; Dias, M.G.; Elgersma, A.; et al. A comprehensive review on carotenoids in foods and feeds: Status quo, applications, patents, and research needs. Crit. Rev. Food Sci. Nutr. 2022, 62, 1999–2049. [Google Scholar] [CrossRef]
- Havale, V.B.; Tawar, R.V.; Hage, N.D.; Kakad, G.J.; Fathepurkar, S.C.; Sable, A.S. Effect of growth regulators and chemicals on growth and flowering of Gladiolus. Asian J. Hortic. 2008, 3, 93–94. [Google Scholar]
- Janowska, B.; Andrzejak, R.; Kosiada, T.; Kwiatkowska, M.; Smolińska, D. The flowering and nutritional status of Gladiolus hybridus cv. Black Velvet following a cytokinin treatment. J. Elementol. 2018, 23, 1119–1128. [Google Scholar] [CrossRef]
- Janowska, B.; Andrzejak, R.; Kosiada, T.; Kwiatkowska, M.; Smolińska, D. The flowering and nutritional status of Gladiolus hybridus ‘Black Velvet’ following gibberellin treatment. Hort. Sci. 2018, 45, 205–210. [Google Scholar] [CrossRef] [Green Version]
- Hiscox, J.D.; Israelstam, G.F. A method for the extraction of chlorophyll from leaf tissue without maceration. Can. J. Bot. 1979, 57, 1332–1334. [Google Scholar] [CrossRef]
- Arnon, D. Copper enzymes in isolated chloroplasts. polyphenoloxidase in Beta vulgaris. Plant Physiol. 1949, 24, 1–15. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kamińska, W.; Kardasz, T.; Strahl, A.; Bałucka, T.; Walczak, K.; Filipek, P. The methods of analysis in chemical-agricultural station. Part II. In Analisis of Plants; IUNG: Puławy, Poland, 1972. [Google Scholar]
- Prisa, D.; Sarrocco, S.; Forti, M.; Burchi, G.; Vannacci, G. Endophytic ability of Trichoderma spp. as inoculants for ornamental plants innovative substrates. J. Plant Pathol. 2013, 86, 169–174. [Google Scholar]
- Janowska, B.; Andrzejak, R.; Kosiada, T. The influence of fungi of the Trichoderma genus on the flowering of Freesia refracta Klatt ‘Argentea’ in winter. Hort. Sci. 2020, 47, 203–210. [Google Scholar] [CrossRef]
- Andrzejak, R.; Janowska, B.; Reńska, B.; Kosiada, T. Effect of Trichoderma spp. and fertilization on the flowering of Begonia × tuberhybrida Voss. ‘Picotee Sunburst’. Agronomy 2021, 11, 1278. [Google Scholar] [CrossRef]
- Błaszczyk, L.; Witaszak, N.; Basińska-Barczak, A.; Marczak, Ł.; Sawikowska, A.; Perlikowski, D.; Kosmala, A. Reakcja roślin pszenicy zwyczajnej (Triticum aestivum L.) na kolonizację korzeni przez grzyby Trichoderma. Biul. IHAR 2019, 285, 129–230. [Google Scholar]
- Souza, R.; Ambrosini, A.; Passaglia, L.M. Plant growth-promoting bacteria as inoculants in agricultural soils. Genet. Mol. Biol. 2015, 38, 401–419. [Google Scholar] [CrossRef]
- Berg, G. Plant-microbe interactions promoting plant growth and health: Perspectives for controlled use of microorganisms in agriculture. Appl. Microbiol. Biotechnol. 2009, 84, 11–18. [Google Scholar] [CrossRef] [PubMed]
- Druzhinina, I.S.; Seidl-Seiboth, V.; Herrera-Estrella, A.; Horwitz, B.A.; Kenerley, C.M.; Monte, E.; Mukherjee, P.K.; Zeilinger, S.; Grigoriev, I.V.; Kubicek, C.P. Trichoderma: The genomics of opportunistic success. Nat. Rev. Microbiol. 2011, 9, 749–759. [Google Scholar] [CrossRef]
- Vargas, W.A.; Crutcher, F.K.; Kenerley, C.M. Functional characterisation of a plant-like sucrose transporter from the beneficial fungus Trichoderma virens. Regulation of the symbiotic association with plants by sucrose metabolism inside the fungal cells. New Phytol. 2011, 189, 777–789. [Google Scholar] [CrossRef] [PubMed]
- Sisodia, A.; Pal, A.; Singh, A.K. Varietal response and effect of Trichoderma on flowering in gladiolus. J. Pharmacogn. Phytochem. 2018, 7, 3658–3660. [Google Scholar]
- da Cruz, L.; Ludwig, F.; Steffen, G.; Joseila Maldaner, J. Development and quality of gladiolus stems with the use of vermicompost and Trichoderma sp. in substrate. Ornam. Hortic. 2018, 24, 70–77. [Google Scholar] [CrossRef] [Green Version]
- Prisa, D. Trichoderma harzianum: Biocontrol to Rhizoctonia solani and biostimulation in Pachyphytum oviferum and Crassula falcata. WJARR 2019, 3, 11–18. [Google Scholar]
- Harman, G.E.; Doni, F.; Khadka, R.B.; Uphoff, N. Endophytic strains of Trichoderma increase plants’ photosynthetic capability. J. Appl. Microbiol. 2021, 130, 529–546. [Google Scholar] [CrossRef]
- Yadav, S.; Kushwaha, H.R.; Kumar, K.; Verma, P.K. Comparative structural modeling of a monothiol GRX from chickpea: Insight in iron-sulfur cluster assembly. Int. J. Biol. Macromol. 2012, 51, 266–273. [Google Scholar] [CrossRef]
- Hasanuzzaman, M.; Bhuyan, M.H.; Parvin, K.; Bhuiyan, T.F.; Anee, T.I.; Nahar, K.; Hossen, M.; Zulfiqar, F.; Alam, M.; Fujita, M. Regulation of ROS metabolism in plants under environmental stress: A review of Recent Experimental Evidence. Int. J. Mol. Sci. 2020, 21, 8695. [Google Scholar] [CrossRef]
- Andersson, S.C. Carotenoids, Tocochromanols and Chlorophylls in Sea Buckthorn Berries (Hippophae rhamnoides) and Rose Hips (Rosa sp.). Ph.D. Thesis, Swedish University of Agricultural Sciences, Alnarp, Sweden, 2009; pp. 15–20. [Google Scholar]
- Simkin, A.J. Carotenoids and apocarotenoids in planta: Their role in plant development, contribution to the flavour and aroma of fruits and flowers, and their nutraceutical benefits. Plants 2021, 10, 2321. [Google Scholar] [CrossRef]
- Widomska, J.; Kostecka-Gugała, A.; Latowski, D.; Gruszecki, W.I.; Strzałka, K. Calorimetric studies of the effect of cis-carotenoids on the thermotropic phase behavior of phosphatidylcholine bilayers. Biophys. Chem. 2008, 140, 108–114. [Google Scholar] [CrossRef] [PubMed]
- Metwally, R.A.; Al-Amri, S. Individual and interactive role of Trichoderma viride and arbuscular mycorrhizal fungi on growth and pigment content of onion plants. Lett. Appl. Microbiol. 2020, 70, 79–86. [Google Scholar] [CrossRef] [PubMed]
- Abdel-Fattah, G.M.; Shabana, Y.M.; Adel EIsmail, A.E.; Rashad, Y.M. Trichoderma harzianum: A biocontrol agent against Bipolaris oryzae. Mycopathologia 2007, 164, 81–89. [Google Scholar] [CrossRef] [PubMed]
- da Silva, L.R.; Valadares-Inglis, M.C.; Peixoto, G.H.; de Luccas, B.E.; Muniz, P.H.; Magalhães, D.M.; Moraes, M.C.; de Mello, S.C. Volatile organic compounds emitted by Trichoderma azevedoi promote the growth of lettuce plants and delay the symptoms of white mold. Biol. Control 2021, 152, 104447. [Google Scholar] [CrossRef]
- Hosseinzeynali, A.; Abbaszadeh Dahaji, P.; Alaei, H.; Hosseinifard, J.; Akhgar, A. Effect of Trichoderma on growth and nutrition of Pistachio trees under ommon garden conditio. J. Soil Biol. 2021, 8, 115–129. [Google Scholar]
- Metwally, R.A. Arbuscular mycorrhizal fungi and Trichoderma viride cooperative effect on biochemical, mineral content, and protein pattern of onion plants. J. Basic Microbiol. 2020, 60, 712–721. [Google Scholar] [CrossRef]
- Chandra, H.; Kumari, P.; Bisht, R.; Prasad, R.; Yadav, S. Plant growth promoting Pseudomonas aeruginosa from Valeriana wallichii displays antagonistic potential against three phytopathogenic fungi. Mol. Biol. Rep. 2020, 47, 6015–6026. [Google Scholar] [CrossRef]
- Khosa, S.S.; Younis, A.; Rayit, A.; Yasmeen, S.; Riaz, A. Effect of foliar application of macro and micro nutrients on growth and flowering of Gerbera jamesonii L. AEJAES 2011, 11, 736–757. [Google Scholar]
- Lahijie, M.F. Application of micronutrients FeSO4 and ZnSO4 on the growth and development of Gladiolus variety ‘Oscar’. IJACS 2012, 4, 718–720. [Google Scholar]
- Khan, M.Y.; Haque, M.M.; Molla, A.H.; Rahman, M.M.; Alam, M.Z. Antioxidant compounds and minerals in tomatoes by Trichoderma-enriched biofertilizer and their relationship with the soil environments. JIA 2017, 16, 691–703. [Google Scholar] [CrossRef]
- de Santiago, A.; Quintero, J.M.; Avilés, M.; Delgado, A. Effect of Trichoderma asperellum strain T34 on iron, copper, manganese, and zinc uptake by wheat grown on a calcareous medium. Plant Soil. 2011, 342, 97–104. [Google Scholar] [CrossRef]
- Borrero, C.; Trillas, M.I.; Delgado, A.; Avilés, M. Effect of ammonium/nitrate ratio in nutrient solution on control of Fusarium wilt of tomato by Trichoderma asperellum T34. Plant Pathol. 2012, 61, 132–139. [Google Scholar] [CrossRef]
- Vukelić, I.D.; Prokić, L.T.; Racić, G.M.; Pešić, M.B.; Bojović, M.M.; Sierka, E.M.; Kalaji, H.M.; Panković, D.M. Effects of Trichoderma harzianum on photosynthetic characteristics and fruit quality of Tomato plants. Int. J. Mol. Sci. 2021, 22, 6961. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Year | Trichoderma spp. | Mean | |
|---|---|---|---|
| No | Yes | ||
| % in DW | |||
| N | |||
| 2018 | 2.62 a | 2.51 a | 2.56 a |
| 2019 | 2.42 a | 2.49 a | 2.45 a |
| Mean | 2.52 a | 2.50 a | |
| P | |||
| 2018 | 0.51 a | 0.79 b | 2.56 b |
| 2019 | 0.47 a | 0.81 b | 2.45 b |
| Mean | 0.49 a | 0.80 c | |
| K | |||
| 2018 | 4.92 a | 6.52 b | 5.72 b |
| 2019 | 4.76 a | 7.34 c | 6.05 b |
| Mean | 4.84 a | 6.93 c | |
| Mg | |||
| 2018 | 0.21 a | 0.18 a | 0.19 a |
| 2019 | 0.15 a | 0.16 a | 0.15 a |
| Mean | 0.18 a | 0.17 a | |
| Ca | |||
| 2018 | 1.20 a | 1.36 b | 1.28 b |
| 2019 | 1.18 a | 1.42 c | 1.30 b |
| Mean | 1.19 a | 1.39 c | |
| Year | Trichoderma spp. | Mean | |
|---|---|---|---|
| No | Yes | ||
| mgˑkg−1 in DW | |||
| Mn | |||
| 2018 | 124.4 a | 125.0 a | 124.7 a |
| 2019 | 127.2 a | 126.2 a | 126.7 a |
| Mean | 125.8 a | 125. a | |
| Cu | |||
| 2018 | 10.8 a | 11.0 a | 10.9 a |
| 2019 | 11.2 a | 11.2 a | 11.2 a |
| Mean | 11.0 a | 11.1 a | |
| Zn | |||
| 2018 | 20.2 a | 40.4 b | 30.3 b |
| 2019 | 18.6 a | 42.2 b | 30.4 b |
| Mean | 19.4 a | 41.3 c | |
| Fe | |||
| 2018 | 200.0 a | 262.0 b | 231.0 b |
| 2019 | 202.2 a | 258.4 b | 230.3 b |
| Mean | 201.1 a | 260.2 c | |
| B | |||
| 2018 | 52.2 a | 60.4 b | 56.3 b |
| 2019 | 54.6 a | 61.6 b | 58.1 b |
| Mean | 53.4 a | 61.0 b | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Andrzejak, R.; Janowska, B. Flowering, Nutritional Status, and Content of Chloroplast Pigments in Leaves of Gladiolus hybridus L. ‘Advances Red’ after Application of Trichoderma spp. Sustainability 2022, 14, 4576. https://0-doi-org.brum.beds.ac.uk/10.3390/su14084576
Andrzejak R, Janowska B. Flowering, Nutritional Status, and Content of Chloroplast Pigments in Leaves of Gladiolus hybridus L. ‘Advances Red’ after Application of Trichoderma spp. Sustainability. 2022; 14(8):4576. https://0-doi-org.brum.beds.ac.uk/10.3390/su14084576
Chicago/Turabian StyleAndrzejak, Roman, and Beata Janowska. 2022. "Flowering, Nutritional Status, and Content of Chloroplast Pigments in Leaves of Gladiolus hybridus L. ‘Advances Red’ after Application of Trichoderma spp." Sustainability 14, no. 8: 4576. https://0-doi-org.brum.beds.ac.uk/10.3390/su14084576