
Growth Performance, Gut Health, Welfare and Qualitative Behavior Characteristics of Broilers Fed Diets Supplemented with Dried Common (Olea europaea) Olive Pulp
 , , , , , and
, , , , , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Animals, Diets and Experimental Design
2.2. Production Traits and Fecal Sampling
2.3. Welfare and Behavior Indicators
2.4. Microbiota Analysis
2.4.1. Sample Collection and Processing
2.4.2. 16S rRNA Library Construction and Sequencing
2.5. Statistical Analysis
Bioinformatic Analysis
2.6. Availability of Data and Materials
3. Results
3.1. Growth Performance
3.2. Welfare and Behavior Indicators
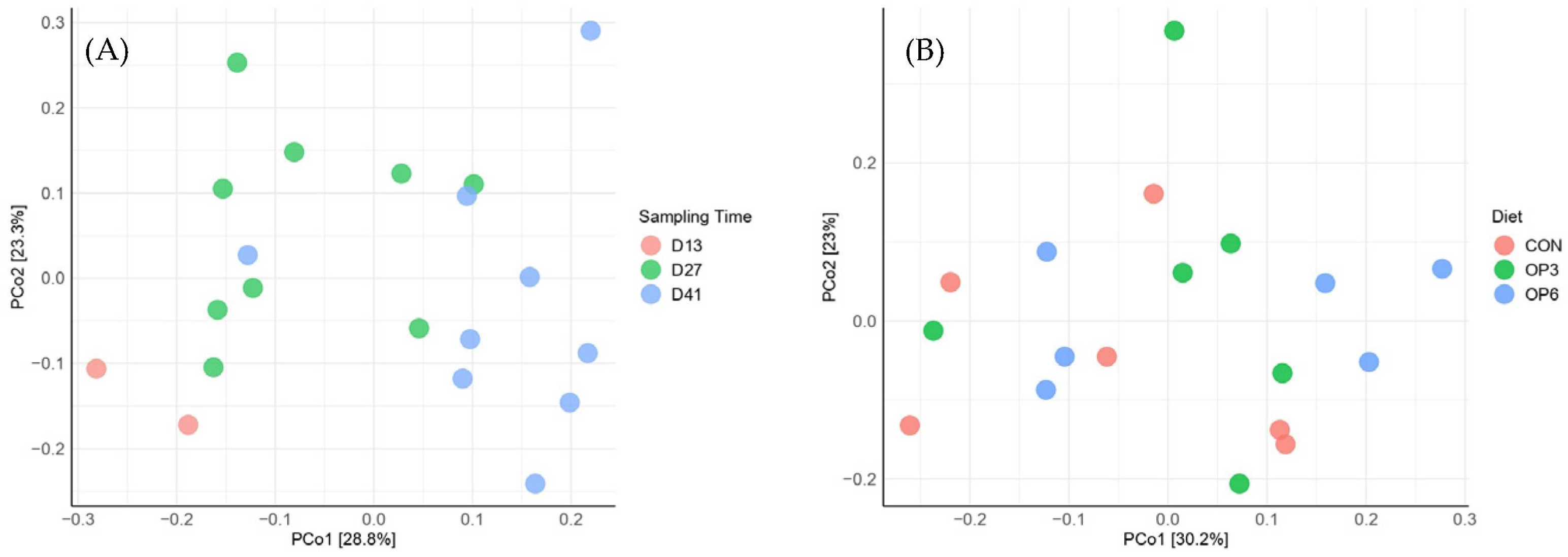
3.3. Dietary Effects of OP on Fecal Microbiome
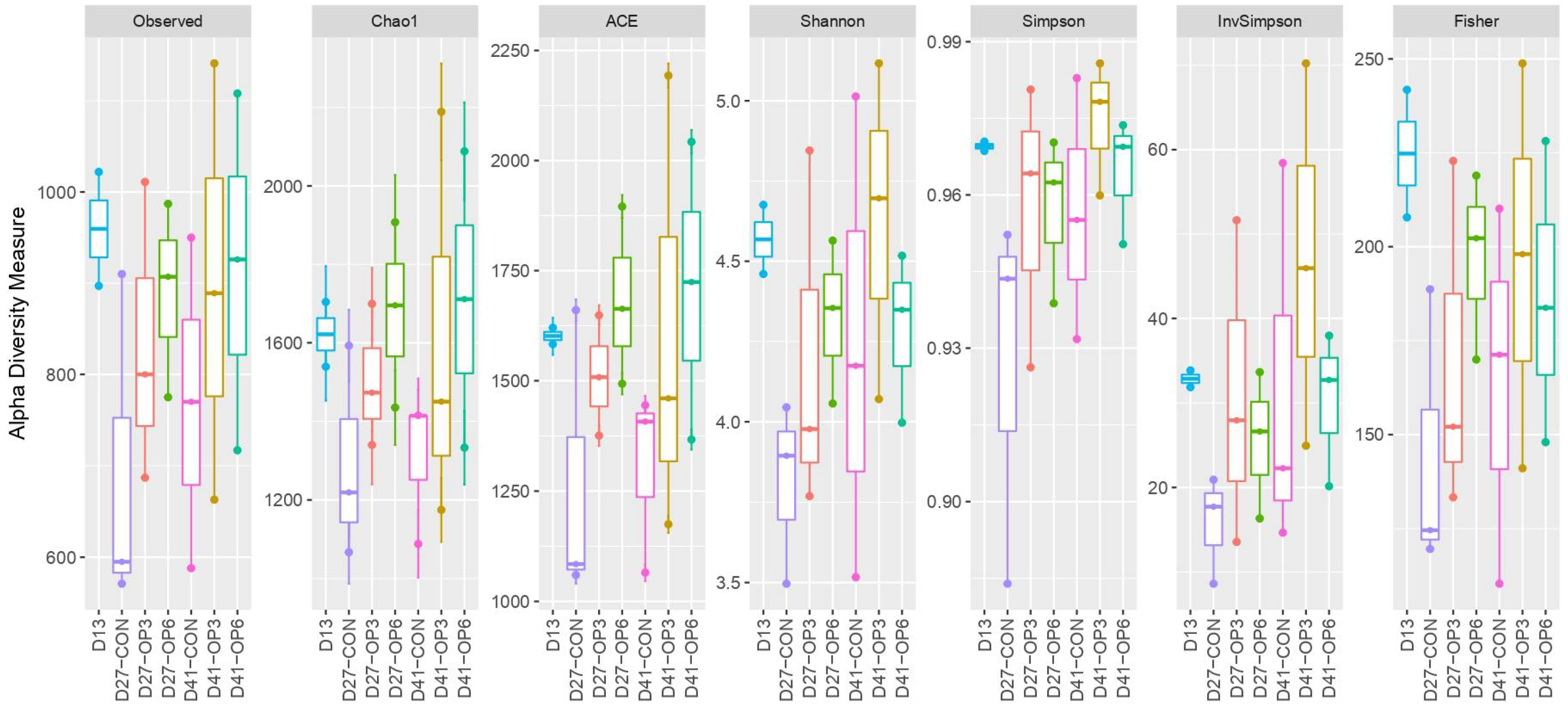
3.3.1. Microbiome Diversity Measures
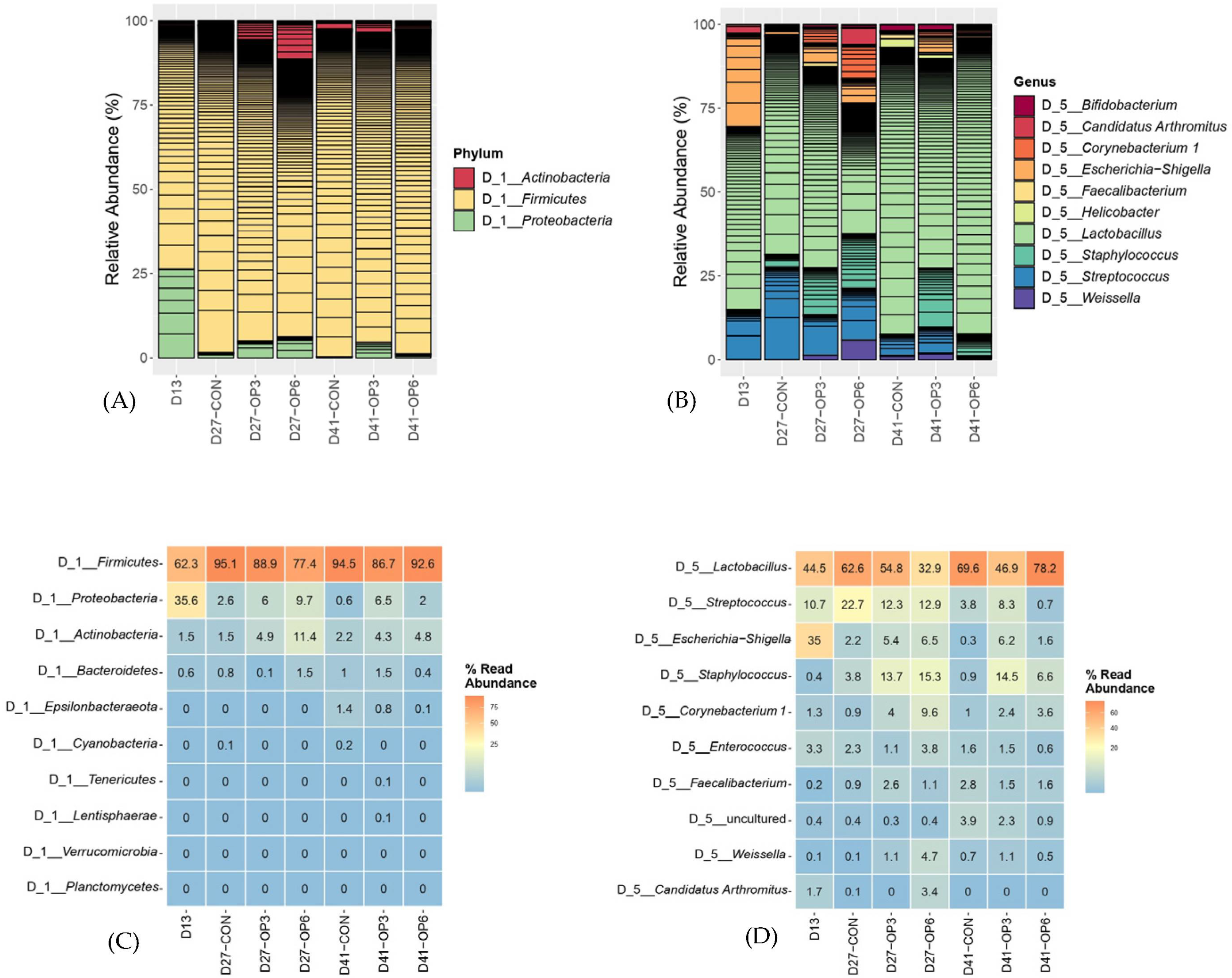
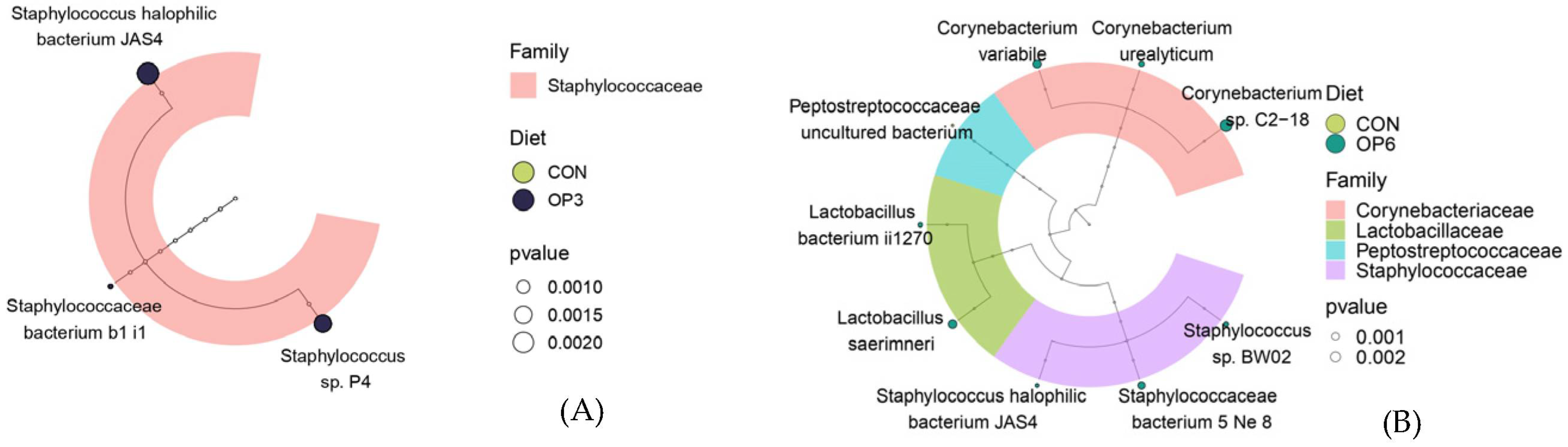
3.3.2. Microbial Community Composition
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Diaz Carrasco, J.M.; Casanova, N.A.; Fernández Miyakawa, M.E. Microbiota, Gut Health and Chicken Productivity: What Is the Connection? Microorganisms 2019, 7, 374. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ducatelle, R.; Goossens, E.; De Meyer, F.; Eeckhaut, V.; Antonissen, G.; Haesebrouck, F.; Van Immerseel, F. Biomarkers for monitoring intestinal health in poultry: Present status and future perspectives. Vet. Res. 2018, 49, 43. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- O’Mahony, S.M.; Clarke, G.; Borre, Y.E.; Dinan, T.G.; Cryan, J.F. Serotonin, tryptophan metabolism and the brain-gut-microbiome axis. Behav. Brain Res. 2015, 277, 32–48. [Google Scholar] [CrossRef] [PubMed]
- Foster, J.A.; Rinaman, L.; Cryan, J.F. Stress & the gut-brain axis: Regulation by the microbiome. Neurobiol. Stress 2017, 7, 124–136. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hoban, A.E.; Moloney, R.D.; Golubeva, A.V.; McVey Neufeld, K.A.; O’Sullivan, O.; Patterson, E.; Stanton, C.; Dinan, T.G.; Clarke, G.; Cryan, J.F. Behavioural and neurochemical consequences of chronic gut microbiota depletion during adulthood in the rat. Neuroscience 2016, 339, 463–477. [Google Scholar] [CrossRef] [PubMed]
- Ceylani, T.; Jakubowska-Doğru, E.; Gurbanov, R.; Teker, H.T.; Gozen, A.G. The effects of repeated antibiotic administration to juvenile BALB/c mice on the microbiota status and animal behavior at the adult age. Heliyon 2018, 4, e00644. [Google Scholar] [CrossRef] [PubMed]
- Cao, Y.-N.; Feng, L.-J.; Liu, Y.-Y.; Jiang, K.; Zhang, M.-J.; Gu, Y.-X.; Wang, B.-M.; Gao, J.; Wang, Z.-L.; Wang, Y.-M. Effect of Lactobacillus rhamnosus GG supernatant on serotonin transporter expression in rats with post-infectious irritable bowel syndrome. WJG 2018, 24, 338–350. [Google Scholar] [CrossRef]
- Yousefi, B.; Eslami, M.; Ghasemian, A.; Kokhaei, P.; Salek Farrokhi, A.; Darabi, N. Probiotics importance and their immunomodulatory properties. J. Cell. Physiol. 2019, 234, 8008–8018. [Google Scholar] [CrossRef]
- Cox, C.M.; Dalloul, R.A. Immunomodulatory role of probiotics in poultry and potential in ovo application. Benef. Microbes 2015, 6, 45–52. [Google Scholar] [CrossRef]
- Parois, S.; Calandreau, L.; Kraimi, N.; Gabriel, I.; Leterrier, C. The influence of a probiotic supplementation on memory in quail suggests a role of gut microbiota on cognitive abilities in birds. Behav. Brain Res. 2017, 331, 47–53. [Google Scholar] [CrossRef]
- Yan, F.F.; Wang, W.C.; Cheng, H.W. Bacillus subtilis based probiotic improved bone mass and altered brain serotoninergic and dopaminergic systems in broiler chickens. J. Funct. Foods 2018, 49, 501–509. [Google Scholar] [CrossRef]
- van der Eijk, J.A.J.; de Vries, H.; Kjaer, J.B.; Naguib, M.; Kemp, B.; Smidt, H.; Rodenburg, T.B.; Lammers, A. Differences in gut microbiota composition of laying hen lines divergently selected on feather pecking. Poult. Sci. 2019, 98, 7009–7021. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Liang, S.; Li, X.; Yang, X.; Long, F.; Yang, X. Effects of encapsulated essential oils and organic acids on laying performance, egg quality, intestinal morphology, barrier function, and microflora count of hens during the early laying period. Poult. Sci. 2019, 98, 6751–6760. [Google Scholar] [CrossRef] [PubMed]
- EUR-lex. Regulation (EC) No 1831/2003 of the European Parliament and of the Council of 22 September 2003 on Additives for Use in Animal Nutrition. 2003. Available online: https://eur-lex.europa.eu/legal-content/EN/TXT/?uri=CELEX%3A32003R1831 (accessed on 12 January 2022).
- The Community Summary Report on Trends and Sources of Zoonoses and Zoonotic Agents in the European Union in 2007. EFSA J. 2009, 7, 223r. [CrossRef]
- Fernando, U.; Biswas, D.; Allan, B.; Willson, P.; Potter, A.A. Influence of Campylobacter jejuni fliA, rpoN and flgK genes on colonization of the chicken gut. Int. J. Food Microbiol. 2007, 118, 194–200. [Google Scholar] [CrossRef] [PubMed]
- Papatsiros, V.G.; Katsoulos, P.D.; Koutoulis, K.C.; Karatzia, M.; Dedousi, A.; Christodoulopoulos, G. Alternatives to antibiotics for farm animals. CABI Rev. 2013, 8, 32. [Google Scholar] [CrossRef]
- Lillehoj, H.; Liu, Y.; Calsamiglia, S.; Fernandez-Miyakawa, M.E.; Chi, F.; Cravens, R.L.; Oh, S.; Gay, C.G. Phytochemicals as antibiotic alternatives to promote growth and enhance host health. Vet. Res. 2018, 49, 76. [Google Scholar] [CrossRef] [Green Version]
- Herrero-Encinas, J.; Blanch, M.; Pastor, J.J.; Menoyo, D. Diet Supplementation with a Bioactive Pomace Extract from Olea europaea Partially Mitigates Negative Effects on Gut Health Arising from a Short-Term Fasting Period in Broiler Chickens. Animals 2020, 10, 349. [Google Scholar] [CrossRef] [Green Version]
- Rebollada-Merino, A.; Bárcena, C.; Ugarte-Ruiz, M.; Porras, N.; Mayoral-Alegre, F.J.; Tomé-Sánchez, I.; Domínguez, L.; Rodríguez-Bertos, A. Effects on Intestinal Mucosal Morphology, Productive Parameters and Microbiota Composition after Supplementation with Fermented Defatted Alperujo (FDA) in Laying Hens. Antibiotics 2019, 8, 215. [Google Scholar] [CrossRef] [Green Version]
- Salomone, R.; Ioppolo, G. Environmental impacts of olive oil production: A Life Cycle Assessment case study in the province of Messina (Sicily). J. Clean. Prod. 2012, 28, 88–100. [Google Scholar] [CrossRef]
- Zangeneh, S.; Torki, M. Effects of B-Mannanase Supplementing of Olive Pulp-Included Diet on Performance of Laying Hens, Egg Quality Characteristics, Humoral and Cellular Immune Response and Blood Parameters. Glob. Vet. 2011, 7, 391–398. [Google Scholar]
- Upadhaya, S.D.; Kim, I.H. Efficacy of Phytogenic Feed Additive on Performance, Production and Health Status of Monogastric Animals—A Review. Ann. Anim. Sci. 2017, 17, 929–948. [Google Scholar] [CrossRef] [Green Version]
- Ibrahim, N.S.; Sabic, E.M.; Abu-Taleb, A.M. Effect of inclusion irradiated olive pulp in laying quail diets on biological performance. J. Radiat. Res. Appl. Sci. 2018, 11, 340–346. [Google Scholar] [CrossRef] [Green Version]
- Sayehban, P.; Seidavi, A.; Dadashbeiki, M.; Ghorbani, A.; Araújo, W.A.G.; Albino, L.F.T. Effects of Different Levels of Two Types of Olive Pulp with or without Exogenous Enzyme Supplementation on Broiler Performance and Economic Parameters. Rev. Bras. Cienc. Avic. 2016, 18, 489–500. [Google Scholar] [CrossRef] [Green Version]
- Abdel-Moneim, A.E.; Shehata, A.M.; Alzahrani, S.O.; Shafi, M.E.; Mesalam, N.M.; Taha, A.E.; Swelum, A.A.; Arif, M.; Fayyaz, M.; Abd El-Hack, M.E. The role of polyphenols in poultry nutrition. J. Anim. Physiol. Anim. Nutr. 2020, 104, 1851–1866. [Google Scholar] [CrossRef] [PubMed]
- Visioli, F.; Poli, A.; Gall, C. Antioxidant and other biological activities of phenols from olives and olive oil. Med. Res. Rev. 2002, 22, 65–75. [Google Scholar] [CrossRef] [PubMed]
- Omar, S.H. Oleuropein in Olive and its Pharmacological Effects. Sci. Pharm. 2010, 78, 133–154. [Google Scholar] [CrossRef] [Green Version]
- Barbaro, B.; Toietta, G.; Maggio, R.; Arciello, M.; Tarocchi, M.; Galli, A.; Balsano, C. Effects of the Olive-Derived Polyphenol Oleuropein on Human Health. IJMS 2014, 15, 18508–18524. [Google Scholar] [CrossRef] [Green Version]
- Bulotta, S.; Celano, M.; Lepore, S.M.; Montalcini, T.; Pujia, A.; Russo, D. Beneficial effects of the olive oil phenolic components oleuropein and hydroxytyrosol: Focus on protection against cardiovascular and metabolic diseases. J. Transl. Med. 2014, 12, 219. [Google Scholar] [CrossRef] [Green Version]
- Casamenti, F.; Grossi, C.; Rigacci, S.; Pantano, D.; Luccarini, I.; Stefani, M. Oleuropein Aglycone: A Possible Drug against Degenerative Conditions. Vivo Evid. Eff. Against Alzheimer’s Dis. 2015, 45, 679–688. [Google Scholar] [CrossRef]
- Duborija-Kovacevic, N.; Shavrina, K. New findings on the pharmacodynamic actions of olive oil: Our contribution to better evidence about its remedial properties. Prog. Nutr. 2018, 20, 30–38. [Google Scholar] [CrossRef]
- Abd El-Galil, K.; Morsy, A.S.; Emam, K.R.S.; Hassan, A.M. Physiological responses and productive performance of laying hens fed olive cake under South Sinai conditions. Egypt. Poult. Sci. J. 2017, 37, 293–304. [Google Scholar]
- Ranalli, A.; Pollastri, L.; Contento, S.; Di Loreto, G.; Iannucci, E.; Lucera, L.; Russi, F. Acylglycerol and Fatty Acid Components of Pulp, Seed, and Whole Olive Fruit Oils. Their Use to Characterize Fruit Variety by Chemometrics. J. Agric. Food Chem. 2002, 50, 3775–3779. [Google Scholar] [CrossRef]
- Tufarelli, V.; Introna, M.; Cazzato, E.; Mazzei, D.; Laudadio, V. Suitability of partly destoned exhausted olive cake as by-product feed ingredient for lamb production. J. Anim. Sci. 2013, 91, 872–877. [Google Scholar] [CrossRef] [Green Version]
- Rabayaa, E.; Omar, J.M.A.; Othman, R.A. Utilization of Olive Pulp in Broiler Rations. An-Najah Univ. J. Res. 2001, 15, 133–144. [Google Scholar]
- Mohamed, M.E.; Ali, A.M.; Shetaewi, M.M.; Said, K.I.; Abdel-Ghaffar, M.A. Utilization of Olive Pomace Meal and Orange By-Product Meal In Broiler Rations. Sinai J. Appl. Sci. 2013, 2, 113–128. [Google Scholar] [CrossRef] [Green Version]
- Al-Harthi, M.A. The efficacy of using olive cake as a by-product in broiler feeding with or without yeast. Ital. J. Anim. Sci. 2016, 15, 512–520. [Google Scholar] [CrossRef]
- Al-Harthi, M. The Effect of Olive Cake, with or Without Enzymes Supplementation, on Growth Performance, Carcass Characteristics, Lymphoid Organs and Lipid Metabolism of Broiler Chickens. Rev. Bras. Cienc. Avic. 2017, 19, 83–90. [Google Scholar] [CrossRef] [Green Version]
- Sateri, S.; Seidavi, A.; Bouyeh, M.; Neumann, P.; Kutzler, M.; Laudadio, V.; Loperfido, F.; Tufarelli, V. Effect of olive meal and supplemental enzymes on performance traits, blood biochemistry, humoral immunity response and caecal microbiota of broilers. SA J. An. Sci. 2017, 47, 804. [Google Scholar] [CrossRef] [Green Version]
- Papadomichelakis, G.; Pappas, A.C.; Tsiplakou, E.; Symeon, G.K.; Sotirakoglou, K.; Mpekelis, V.; Fegeros, K.; Zervas, G. Effects of dietary dried olive pulp inclusion on growth performance and meat quality of broiler chickens. Livest. Sci. 2019, 221, 115–122. [Google Scholar] [CrossRef]
- Pappas, A.C.; Tsiplakou, E.; Papadomichelakis, G.; Mitsiopoulou, C.; Sotirakoglou, K.; Mpekelis, V.; Haroutounian, S.A.; Fegeros, K.; Zervas, G. Effects of olive pulp addition to broiler diets on performance, selected biochemical parameters and antioxidant enzymes. J. Hellenic Vet. Med. Soc. 2019, 70, 1687–1696. [Google Scholar] [CrossRef] [Green Version]
- ELbaz, A.M.; Thabet, H.; Gad, G. Productive and Physiological Performance of Broilers Fed Diets Containing Different Levels of Olive Pulp. J. Anim. Poult. Prod. 2020, 11, 435–439. [Google Scholar] [CrossRef]
- Saleh, A.A.; Paray, B.A.; Dawood, M.A.O. Olive Cake Meal and Bacillus licheniformis Impacted the Growth Performance, Muscle Fatty Acid Content, and Health Status of Broiler Chickens. Animals 2020, 10, 695. [Google Scholar] [CrossRef]
- Fotou, E.; Moulasioti, V.; Angelis, I.; Tsiouris, V.; Patsias, A.; Tellis, C.; Moussis, V.; Boti, M.; Tsoukatos, D. Influence of dietary olive paste flour on the performance and oxidative stress in chickens raised in field conditions. J. Hellenic Vet. Med. Soc. 2021, 72, 3239–3248. [Google Scholar] [CrossRef]
- Saleh, A.; Alzawqari, M. Effects of Replacing Yellow Corn with Olive Cake Meal on Growth Performance, Plasma Lipid Profile, and Muscle Fatty Acid Content in Broilers. Animals 2021, 11, 2240. [Google Scholar] [CrossRef]
- Herrero-Encinas, J.; Blanch, M.; Pastor, J.J.; Mereu, A.; Ipharraguerre, I.R.; Menoyo, D. Effects of a bioactive olive pomace extract from Olea europaea on growth performance, gut function, and intestinal microbiota in broiler chickens. Poult. Sci. 2020, 99, 2–10. [Google Scholar] [CrossRef]
- Debbou-Iouknane, N.; Nerín, C.; Amrane, M.; Ghemghar, M.; Madani, K.; Ayad, A. In Vitro Anticoccidial Activity of Olive Pulp (Olea europaea L. var. Chemlal) Extract Against Eimeria Oocysts in Broiler Chickens. Acta Parasit. 2019, 64, 887–897. [Google Scholar] [CrossRef] [Green Version]
- EUR-lex. COUNCIL DIRECTIVE 2007/43/EC of 28 June 2007 Laying down Minimum Rules for the Protection of Chickens Kept for Meat Production. 2007. Available online: https://eur-lex.europa.eu/legal-content/EN/TXT/HTML/?uri=CELEX:32007L0043&from=EN (accessed on 12 April 2022).
- Aviagen, W. Ross 308: Broiler’s Management and Nutrition Specification. ROSS An Aviagen Brand 2018. Available online: https://en.aviagen.com/assets/Tech_Center/Ross_Broiler/Ross-BroilerHandbook2018-EN.pdf (accessed on 20 April 2022).
- Dedousi, A.; Kritsa, M.-Z.; Đukić Stojčić, M.; Sfetsas, T.; Sentas, A.; Sossidou, E. Production Performance, Egg Quality Characteristics, Fatty Acid Profile and Health Lipid Indices of Produced Eggs, Blood Biochemical Parameters and Welfare Indicators of Laying Hens Fed Dried Olive Pulp. Sustainability 2022, 14, 3157. [Google Scholar] [CrossRef]
- Welfare Quality®. Welfare Quality® Assessment Protocol for Poultry (Broilers, Laying Hens); Welfare Quality® Consortium: Lelystad, The Netherlands, 2009.
- Klindworth, A.; Pruesse, E.; Schweer, T.; Peplies, J.; Quast, C.; Horn, M.; Glöckner, F.O. Evaluation of general 16S ribosomal RNA gene PCR primers for classical and next-generation sequencing-based diversity studies. Nucleic Acids Res. 2013, 41, e1. [Google Scholar] [CrossRef]
- JASP Team. JASP (Version 0.14) [Computer Software]. Available online: https://jasp-stats.org/previous-versions/ (accessed on 21 December 2020).
- Bolyen, E.; Rideout, J.R.; Dillon, M.R.; Bokulich, N.A.; Abnet, C.C.; Al-Ghalith, G.A.; Alexander, H.; Alm, E.J.; Arumugam, M.; Asnicar, F.; et al. Reproducible, interactive, scalable and extensible microbiome data science using QIIME 2. Nat. Biotechnol. 2019, 37, 852–857. [Google Scholar] [CrossRef]
- Martin, M. Cutadapt removes adapter sequences from high-throughput sequencing reads. EMBnet J. 2011, 17, 10. [Google Scholar] [CrossRef]
- Rognes, T.; Flouri, T.; Nichols, B.; Quince, C.; Mahé, F. VSEARCH: A versatile open source tool for metagenomics. PeerJ 2016, 4, e2584. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Quast, C.; Pruesse, E.; Yilmaz, P.; Gerken, J.; Schweer, T.; Yarza, P.; Peplies, J.; Glöckner, F.O. The SILVA ribosomal RNA gene database project: Improved data processing and web-based tools. Nucleic Acids Res. 2012, 41, D590–D596. [Google Scholar] [CrossRef] [PubMed]
- Camacho, C.; Coulouris, G.; Avagyan, V.; Ma, N.; Papadopoulos, J.; Bealer, K.; Madden, T.L. BLAST+: Architecture and applications. BMC Bioinform. 2009, 10, 421. [Google Scholar] [CrossRef] [Green Version]
- R Core Team. 2020. Available online: http://www.r-project.org/index.html (accessed on 15 December 2020).
- McMurdie, P.J.; Holmes, S. phyloseq: An R Package for Reproducible Interactive Analysis and Graphics of Microbiome Census Data. PLoS ONE 2013, 8, e61217. [Google Scholar] [CrossRef] [Green Version]
- Andersen, K.S.; Kirkegaard, R.H.; Karst, S.M.; Albertsen, M. Ampvis2: An R Package to Analyse and Visualise 16S RRNA Amplicon Data. bioRxiv 2018. [Google Scholar] [CrossRef] [Green Version]
- Wickham, H. ggplot2: Elegant Graphics for Data Analysis; Springer: Cham, Switzerland, 2016; ISBN 978-3-319-24275-0. [Google Scholar]
- MicrobiotaProcess: An R Package for Analysis, Visualization and Biomarker Discovery of Microbiome. Available online: https://github.com/YuLab-SMU/MicrobiotaProcess (accessed on 22 April 2022).
- Oksanen, J.; Blanchet, F.G.; Friendly, M.; Kindt, R.; Legendre, P.; McGlinn, D.; Minchin, P.R.; O’Hara, R.B.; Simpson, G.L.; Solymos, P.; et al. Vegan: Community Ecology Package. Available online: https://CRAN.R-project.org/package=vegan (accessed on 23 March 2022).
- Love, M.I.; Huber, W.; Anders, S. Moderated estimation of fold change and dispersion for RNA-seq data with DESeq2. Genome Biol. 2014, 15, 550. [Google Scholar] [CrossRef] [Green Version]
- Zhu, A.; Ibrahim, J.G.; Love, M.I. Heavy-tailed prior distributions for sequence count data: Removing the noise and preserving large differences. Bioinformatics 2019, 35, 2084–2092. [Google Scholar] [CrossRef] [Green Version]
- Sklan, D.; Noy, Y. Functional development and intestinal absorption in the young poult. Br. Poult. Sci. 2003, 44, 651–658. [Google Scholar] [CrossRef]
- Thomas, D.V.; Ravindran, V.; Ravindran, G. Nutrient digestibility and energy utilisation of diets based on wheat, sorghum or maize by the newly hatched broiler chick. Br. Poult. Sci. 2008, 49, 429–435. [Google Scholar] [CrossRef]
- Jiménez-Moreno, E.; González-Alvarado, J.M.; de Coca-Sinova, A.; Lázaro, R.P.; Cámara, L.; Mateos, G.G. Insoluble fiber sources in mash or pellets diets for young broilers. 2. Effects on gastrointestinal tract development and nutrient digestibility. Poult. Sci. 2019, 98, 2531–2547. [Google Scholar] [CrossRef]
- Choct, M. Fibre—Chemistry and Functions in Poultry Nutrition. 2015. Available online: https://www.wpsa-aeca.es/aeca_imgs_docs/16478_fibra_mingan.pdf (accessed on 22 July 2022).
- Zubair, A.K.; Leeson, S. Compensatory growth in the broiler chicken: A review. World’s Poult. Sci. J. 1996, 52, 189–201. [Google Scholar] [CrossRef]
- Tejeda, O.J.; Kim, W.K. Role of Dietary Fiber in Poultry Nutrition. Animals 2021, 11, 461. [Google Scholar] [CrossRef] [PubMed]
- Ravindran, V.; Abdollahi, M.R. Nutrition and Digestive Physiology of the Broiler Chick: State of the Art and Outlook. Animals 2021, 11, 2795. [Google Scholar] [CrossRef] [PubMed]
- Harrow, S.A.; Ravindran, V.; Butler, R.C.; Marshall, J.W.; Tannock, G.W. Real-Time Quantitative PCR Measurement of Ileal Lactobacillus salivarius Populations from Broiler Chickens To Determine the Influence of Farming Practices. Appl. Environ. Microbiol. 2007, 73, 7123–7127. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McBurney, W.; Tannock, G.; Ravindran, V. A culture-independent approach to the analysis of the gut microflora of broilers. Proc. Aust. Poult. Sci. Symp. 2003, 15, 131–134. [Google Scholar]
- Rinttilä, T.; Apajalahti, J. Intestinal microbiota and metabolites—Implications for broiler chicken health and performance. J. Appl. Poult. Res. 2013, 22, 647–658. [Google Scholar] [CrossRef]
- Joven, M.; Pintos, E.; Latorre, M.A.; Suárez-Belloch, J.; Guada, J.A.; Fondevila, M. Effect of replacing barley by increasing levels of olive cake in the diet of finishing pigs: Growth performances, digestibility, carcass, meat and fat quality. Anim. Feed. Sci. Technol. 2014, 197, 185–193. [Google Scholar] [CrossRef]
- Amer, M.M. REVIEW: Footpad dermatitis (FPD) in chickens. Korean J. Food Health Converg. 2020, 6, 11–16. [Google Scholar] [CrossRef]
- Swiatkiewicz, S.; Arczewska-Wlosek, A.; Jozefiak, D. The nutrition of poultry as a factor affecting litter quality and foot pad dermatitis—An updated review. J. Anim. Physiol. Anim. Nutr. 2017, 101, e14–e20. [Google Scholar] [CrossRef]
- Freeman, N.; Tuyttens, F.A.M.; Johnson, A.; Marshall, V.; Garmyn, A.; Jacobs, L. Remedying Contact Dermatitis in Broiler Chickens with Novel Flooring Treatments. Animals 2020, 10, 1761. [Google Scholar] [CrossRef]
- Choi, J.; Kim, W.K. Dietary Application of Tannins as a Potential Mitigation Strategy for Current Challenges in Poultry Production: A Review. Animals 2020, 10, 2389. [Google Scholar] [CrossRef] [PubMed]
- Heidemann Olsen, R.; Christensen, H.; Kabell, S.; Bisgaard, M. Characterization of prevalent bacterial pathogens associated with pododermatitis in table egg layers. Avian Pathol. 2018, 47, 281–285. [Google Scholar] [CrossRef] [PubMed]
- Saraiva, S.; Saraiva, C.; Stilwell, G. Feather conditions and clinical scores as indicators of broilers welfare at the slaughterhouse. Res. Vet. Sci. 2016, 107, 75–79. [Google Scholar] [CrossRef] [PubMed]
- Arnould, C.; Butterworth, A.; Knierim, U. Standardisation of clinical scoring in poultry. In Assessment of Animal Welfare Measures for Layers and Broilers; Forkman, B., Keeling, L., Eds.; School of City and Regional Planning, Cardiff University: Cardiff, UK, 2009; pp. 7–30. ISBN 978-1-902647-79-1. [Google Scholar]
- Grimes, J.L.; Smith, J.; Williams, C.M. Some alternative litter materials used for growing broilers and turkeys. World’s Poult. Sci. J. 2002, 58, 515–526. [Google Scholar] [CrossRef]
- Kaukonen, E.; Norring, M.; Valros, A. Effect of litter quality on foot pad dermatitis, hock burns and breast blisters in broiler breeders during the production period. Avian Pathol. 2016, 45, 667–673. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mayne, R.K. A review of the aetiology and possible causative factors of foot pad dermatitis in growing turkeys and broilers. World’s Poult. Sci. J. 2005, 61, 256–267. [Google Scholar] [CrossRef]
- de Jong, I.C.; Gunnink, H.; van Harn, J. Wet litter not only induces footpad dermatitis but also reduces overall welfare, technical performance, and carcass yield in broiler chickens. J. Appl. Poult. Res. 2014, 23, 51–58. [Google Scholar] [CrossRef]
- Göransson, L.; Yngvesson, J.; Gunnarsson, S. Bird Health, Housing and Management Routines on Swedish Organic Broiler Chicken Farms. Animals 2020, 10, 2098. [Google Scholar] [CrossRef]
- Kheravii, S.K.; Morgan, N.K.; Swick, R.A.; Choct, M.; Wu, S.-B. Roles of dietary fibre and ingredient particle size in broiler nutrition. World’s Poult. Sci. J. 2018, 74, 301–316. [Google Scholar] [CrossRef]
- Farran, M.T.; Pietsch, M.; Chabrillat, T. Effect of lignocellulose on the litter quality and the ready to cook carcass yield of male broilers. In Proceedings of the Actes des 10èmes Journées de la Recherche Avicole et Palmipèdes à Foie Gras, La Rochelle, France, 26–28 March 2013; pp. 917–921. [Google Scholar]
- Bogusławska-Tryk, M.; Szymeczko, R.; Piotrowska, A.; Burlikowska, K.; Śliżewska, K. Ileal and Cecal Microbial Population and Short-Chain Fatty Acid Profile in Broiler Chickens Fed Diets Supplemented with Lignocellulose. Pak. Vet. J. 2015, 35, 212–216. [Google Scholar]
- Milošević, N.; Đukić Stojčić, M.; Perić, L.; Vukić-Vranješ, M. Effect of lignocellulose on egg production and egg quality. In Proceedings of the 20th European Symposium on Poultry Nutrition, Prague, Czech Republic, 24–27 August 2015; p. P-046. [Google Scholar]
- Kheravii, S.K.; Swick, R.A.; Choct, M.; Wu, S.-B. Coarse particle inclusion and lignocellulose-rich fiber addition in feed benefit performance and health of broiler chickens. Poult. Sci. 2017, 96, 3272–3281. [Google Scholar] [CrossRef] [PubMed]
- Musch, M.W.; Bookstein, C.; Xie, Y.; Sellin, J.H.; Chang, E.B. SCFA increase intestinal Na absorption by induction of NHE3 in rat colon and human intestinal C2/bbe cells. Am. J. Physiol. Gastrointest. Liver Physiol. 2001, 280, G687–G693. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gupta, N.; Martin, P.M.; Prasad, P.D.; Ganapathy, V. SLC5A8 (SMCT1)-mediated transport of butyrate forms the basis for the tumor suppressive function of the transporter. Life Sci. 2006, 78, 2419–2425. [Google Scholar] [CrossRef] [PubMed]
- Cherrington, C.A.; Hinton, M.; Mead, G.C.; Chopra, I. Organic Acids: Chemistry, Antibacterial Activity and Practical Applications. In Advances in Microbial Physiology; Elsevier: Amsterdam, The Netherlands, 1991; Volume 32, pp. 87–108. ISBN 978-0-12-027732-2. [Google Scholar]
- van der Wielen, P.W.J.J.; Biesterveld, S.; Notermans, S.; Hofstra, H.; Urlings, B.A.P.; van Knapen, F. Role of Volatile Fatty Acids in Development of the Cecal Microflora in Broiler Chickens during Growth. Appl. Environ. Microbiol. 2000, 66, 2536–2540. [Google Scholar] [CrossRef]
- Michalak, M.; Pierzak, M.; Kręcisz, B.; Suliga, E. Bioactive Compounds for Skin Health: A Review. Nutrients 2021, 13, 203. [Google Scholar] [CrossRef]
- Anunciato, T.P.; da Rocha Filho, P.A. Carotenoids and polyphenols in nutricosmetics, nutraceuticals, and cosmeceuticals: Nutricosmetics. J. Cosmet. Dermatol. 2012, 11, 51–54. [Google Scholar] [CrossRef]
- Nichols, J.A.; Katiyar, S.K. Skin photoprotection by natural polyphenols: Anti-inflammatory, antioxidant and DNA repair mechanisms. Arch. Dermatol. Res. 2010, 302, 71–83. [Google Scholar] [CrossRef] [Green Version]
- Zillich, O.V.; Schweiggert-Weisz, U.; Eisner, P.; Kerscher, M. Polyphenols as active ingredients for cosmetic products. Int. J. Cosmet. Sci. 2015, 37, 455–464. [Google Scholar] [CrossRef]
- Haslam, S.M.; Knowles, T.G.; Brown, S.N.; Wilkins, L.J.; Kestin, S.C.; Warriss, P.D.; Nicol, C.J. Factors affecting the prevalence of foot pad dermatitis, hock burn and breast burn in broiler chicken. Br. Poult. Sci. 2007, 48, 264–275. [Google Scholar] [CrossRef] [Green Version]
- Hocking, P.M.; Wu, K. Traditional and commercial turkeys show similar susceptibility to foot pad dermatitis and behavioural evidence of pain. Br. Poult. Sci. 2013, 54, 281–288. [Google Scholar] [CrossRef] [PubMed]
- Videnska, P.; Sedlar, K.; Lukac, M.; Faldynova, M.; Gerzova, L.; Cejkova, D.; Sisak, F.; Rychlik, I. Succession and Replacement of Bacterial Populations in the Caecum of Egg Laying Hens over Their Whole Life. PLoS ONE 2014, 9, e115142. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yan, W.; Sun, C.; Yuan, J.; Yang, N. Gut metagenomic analysis reveals prominent roles of Lactobacillus and cecal microbiota in chicken feed efficiency. Sci. Rep. 2017, 7, 45308. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sarıca, S.; Ürkmez, D. The use of grape seed-, olive leaf- and pomegranate peel-extracts as alternative natural antimicrobial feed additives in broiler diets. Eur. Poult. Sci. 2016, 80, 121. [Google Scholar]
- Amini, Z.; Parsaei, S.; Houshmand, M.; Naghiha, R. Effect of Olive leaf powder on performance and ileal bacteria count of broilers. Vet. Res. Forum. 2019, 10, 255–259. [Google Scholar] [CrossRef]
- García-Ruiz, A.; Bartolomé, B.; Martínez-Rodríguez, A.J.; Pueyo, E.; Martín-Álvarez, P.J.; Moreno-Arribas, M.V. Potential of phenolic compounds for controlling lactic acid bacteria growth in wine. Food Control 2008, 19, 835–841. [Google Scholar] [CrossRef]
- Alizadeh, M.; Shojadoost, B.; Astill, J.; Taha-Abdelaziz, K.; Karimi, S.H.; Bavananthasivam, J.; Kulkarni, R.R.; Sharif, S. Effects of in ovo Inoculation of Multi-Strain Lactobacilli on Cytokine Gene Expression and Antibody-Mediated Immune Responses in Chickens. Front. Vet. Sci. 2020, 7, 105. [Google Scholar] [CrossRef] [Green Version]
- Syed, M.A.; Ullah, H.; Tabassum, S.; Fatima, B.; Woodley, T.A.; Ramadan, H.; Jackson, C.R. Staphylococci in poultry intestines: A comparison between farmed and household chickens. Poult. Sci. 2020, 99, 4549–4557. [Google Scholar] [CrossRef]
- Videnska, P.; Rahman, M.M.; Faldynova, M.; Babak, V.; Matulova, M.E.; Prukner-Radovcic, E.; Krizek, I.; Smole-Mozina, S.; Kovac, J.; Szmolka, A.; et al. Characterization of Egg Laying Hen and Broiler Fecal Microbiota in Poultry Farms in Croatia, Czech Republic, Hungary and Slovenia. PLoS ONE 2014, 9, e110076. [Google Scholar] [CrossRef]
- Liehr, M.; Mereu, A.; Pastor, J.J.; Quintela, J.C.; Staats, S.; Rimbach, G.; Ipharraguerre, I.R. Olive oil bioactives protect pigs against experimentally-induced chronic inflammation independently of alterations in gut microbiota. PLoS ONE 2017, 12, e0174239. [Google Scholar] [CrossRef] [Green Version]
- Stanley, D.; Geier, M.S.; Chen, H.; Hughes, R.J.; Moore, R.J. Comparison of fecal and cecal microbiotas reveals qualitative similarities but quantitative differences. BMC Microbiol. 2015, 15, 51. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Grower (13–20 Days of Age) | Finisher 1 (21–32 Days of Age) | Finisher 2 (33–41 Days of Age) | |||||||
|---|---|---|---|---|---|---|---|---|---|
| Items | CON | OP3 | OP6 | CON | OP3 | OP6 | CON | OP3 | OP6 |
| Ingredients | |||||||||
| Wheat | 36.395 | 33.615 | 30.955 | 40.95 | 38.105 | 35.35 | 44.87 | 42.135 | 39.28 |
| Corn | 25 | 25 | 25 | 25 | 25 | 25 | 25 | 25 | 25 |
| Soya meal | 31.3 | 31.4 | 31.47 | 26.43 | 26.58 | 26.72 | 22.33 | 22.43 | 22.6 |
| Olive pulp | 0 | 3 | 6 | 0 | 3 | 6 | 0 | 3 | 6 |
| Sunflower oil | 4.1 | 3.9 | 3.6 | 4.9 | 4.7 | 4.45 | 5.15 | 4.9 | 4.7 |
| MCP * | 0.42 | 0.42 | 0.43 | 0.215 | 0.22 | 0.22 | 0.185 | 0.19 | 0.195 |
| Premix 1 | 0.4 | 0.4 | 0.4 | 0.4 | 0.4 | 0.4 | 0.4 | 0.4 | 0.4 |
| Limestone | 1.05 | 0.93 | 0.8 | 0.86 | 0.74 | 0.61 | 0.9 | 0.78 | 0.66 |
| NaCl | 0.28 | 0.28 | 0.28 | 0.28 | 0.28 | 0.28 | 0.28 | 0.28 | 0.28 |
| Methionine | 0.35 | 0.34 | 0.35 | 0.3 | 0.3 | 0.3 | 0.25 | 0.25 | 0.25 |
| Lysine | 0.26 | 0.27 | 0.27 | 0.24 | 0.25 | 0.25 | 0.24 | 0.24 | 0.24 |
| Threonine | 0.15 | 0.15 | 0.15 | 0.135 | 0.135 | 0.135 | 0.11 | 0.11 | 0.11 |
| RONOZYME® HiPhos | 0.02 | 0.02 | 0.02 | 0.015 | 0.015 | 0.015 | 0.01 | 0.01 | 0.01 |
| Mycotoxins Binder | 0.2 | 0.2 | 0.2 | 0.2 | 0.2 | 0.2 | 0.2 | 0.2 | 0.2 |
| Coccidiostat | 0.05 | 0.05 | 0.05 | 0.05 | 0.05 | 0.05 | 0.05 | 0.05 | 0.05 |
| Antioxidant | 0.015 | 0.015 | 0.015 | 0.015 | 0.015 | 0.015 | 0.015 | 0.015 | 0.015 |
| Xylanase | 0.01 | 0.01 | 0.01 | 0.01 | 0.01 | 0.01 | 0.01 | 0.01 | 0.01 |
| Total | 100 | 100 | 100 | 100 | 100 | 100 | 100 | 100 | 100 |
| Chemical analysis | |||||||||
| Crude protein (%) | 20.774 | 20.8 | 20.833 | 18.985 | 19.003 | 19.008 | 17.49 | 17.52 | 17.57 |
| Crude fiber (%) | 2.75 | 3.45 | 4.15 | 2.65 | 3.35 | 4.05 | 2.57 | 3.27 | 3.91 |
| Fat (%) | 5.95 | 6.15 | 6.25 | 6.75 | 6.95 | 7.1 | 7.01 | 7.15 | 7.25 |
| Ash (%) | 4.76 | 4.8 | 4.85 | 4.12 | 4.18 | 4.22 | 3.93 | 3.98 | 4.04 |
| Calculated analysis | |||||||||
| ME (kcal/kg) | 2901 | 2903 | 2903 | 2993 | 2998 | 2999 | 3040 | 3042 | 3044 |
| Lysine (%) | 1.256 | 1.259 | 1.254 | 1.117 | 1.12 | 1.118 | 1.013 | 1.009 | 1.075 |
| Methionine (%) | 0.652 | 0.649 | 0.648 | 0.579 | 0.576 | 0.577 | 0.511 | 0.509 | 0.507 |
| Ca (%) | 0.63 | 0.631 | 0.63 | 0.515 | 0.516 | 0.514 | 0.514 | 0.516 | 0.517 |
| P (%) | 0.462 | 0.461 | 0.46 | 0.398 | 0.397 | 0.397 | 0.377 | 0.376 | 0.375 |
| Na (%) | 0.12 | 0.12 | 0.12 | 0.12 | 0.12 | 0.12 | 0.12 | 0.12 | 0.12 |
| CON | OP3 | OP6 | |
|---|---|---|---|
| Body weight (g) | |||
| Day 13 | 428.61 ± 2.34 | 426.39 ± 2.87 | 429.03 ± 2.88 |
| Day 20 | 873.33 ± 8.18 a | 809.86 ± 9.70 b | 812.22 ± 12.45 b |
| Day 27 | 1346.67 ± 17.52 a | 1276.11 ± 19.00 b | 1350.71 ± 19.51 a |
| Day 34 | 1943.75 ± 33.79 a | 1791.94 ± 32.70 b | 1879.00 ± 30.56 ab |
| Day 41 | 2604.44 ± 37.06 | 2492.78 ± 38.79 | 2531.32 ± 45.67 |
| Body weight gained (g) | |||
| 13–20 d | 444.72 ±7.51 a | 383.47 ± 5.47 b | 383.19 ± 18.51 b |
| 21–27 d | 473.33 ±18.28 | 466.25 ± 18.19 | 537.17 ± 18.34 |
| 28–34 d | 597.08 ± 24.81 | 515.83 ± 35.63 | 530.34 ± 37.26 |
| 35–41 d | 660.69 ± 30.21 | 700.83 ± 19.14 | 649.41 ± 34.54 |
| Total period (13–41 d) | 543.96 ± 28.25 | 516.60 ± 36.37 | 525.03 ± 31.00 |
| Feed consumption (g) | |||
| 13–20 d | 754.58 ± 31.28 | 825.00 ± 58.21 | 845.28 ± 18.55 |
| 21–27 d | 1062.50 ± 25.09 a | 1462.36 ± 86.81 b | 1472.16 ± 102.20 b |
| 28–34 d | 1274.03 ± 11.40 | 1483.47 ± 55.25 | 1567.12 ± 122.41 |
| 35–41 d | 2155.69 ± 37.29 a | 1501.53 ± 66.77 b | 1627.97 ± 67.16 b |
| Total period (13–41 d) | 1311.70 ± 157.57 | 1318.09 ± 90.68 | 1378.13 ± 101.30 |
| FCR | |||
| 13–20 d | 1.70 ± 0.05 | 2.16 ± 0.18 | 2.22 ± 0.11 |
| 21–27 d | 2.25 ± 0.05 | 3.16 ± 0.31 | 2.75 ± 0.25 |
| 28–34 d | 2.14 ± 0.09 a | 2.91 ± 0.28 ab | 2.96 ± 0.11 b |
| 35–41 d | 3.27 ± 0.09 a | 2.14 ± 0.09 b | 2.51 ± 0.03 c |
| Total period (13–41 d) | 2.34 ± 0.18 | 2.59 ± 0.17 | 2.61 ± 0.11 |
| Day 34 | Day 41 | |||||
|---|---|---|---|---|---|---|
| Score | CON | OP3 | OP6 | CON | OP3 | OP6 |
| Feather cleanliness 1 | ||||||
| 0 | 36.11 | 61.11 | 51.43 | 13.89 a | 47.22 b | 50.00 b |
| 1 | 44.44 | 38.89 | 48.57 | 47.22 | 38.89 | 44.12 |
| 2 | 19.44 a | 0.00 b | 0.00 b | 36.11 a | 13.89 ab | 5.88 b |
| 3 | - | - | - | 2.78 | 0 | 0 |
| Foot pad dermatitis 2 | ||||||
| 0 | 47.22 a | 91.67 b | 82.86 b | 41.67 a | 86.11 b | 79.41 b |
| 1 | 27.78 a | 5.56 b | 2.86 b | 13.89 | 11.11 | 8.82 |
| 2 | 22.22 a | 2.78 b | 8.57 ab | 25.00 a | 2,78 b | 2.94 b |
| 3 | 2.78 | 0.00 | 5.71 | 16.67 a | 0 b | 2.94 ab |
| 4 | - | - | - | 2.78 | 0 | 5.88 |
| Hock burn 3 | ||||||
| 0 | 97.22 | 94.44 | 100 | 94.44 | 91.67 | 97.06 |
| 1 | 2.78 | 2.78 | 0 | 5.56 | 2.78 | 2.94 |
| 2 | - | - | - | 0 | 2.78 | 0 |
| 3 | - | - | - | - | - | - |
| 4 | 0 | 2.78 | 0 | 0 | 2.78 | 0 |
| Quality Behavior Traits | Day 34 | Day 41 | ||||
|---|---|---|---|---|---|---|
| CON | OP3 | OP6 | CON | OP3 | OP6 | |
| Active | 19.44 | 30.56 | 22.22 | 16.67 | 25 | 30.55 |
| Fearful | 0 | 0 | 0 | 0 | 0 | 0 |
| Depressed | 0 | 0 | 0 | 0 | 0 | 0 |
| Calm | 63.89 | 55.55 | 61.11 | 72.22 | 52.78 | 52.78 |
| Bored | 0 | 0 | 2.78 | 0 | 0 | 0 |
| Friendly | 0 | 2.78 | 0 | 0 | 2.78 | 0 |
| Feeding | 16.67 | 11.11 | 13.89 | 11.11 | 19.44 | 16.67 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Dedousi, A.; Kotzamanidis, C.; Kritsa, M.-Z.; Tsoureki, A.; Andreadelli, A.; Patsios, S.I.; Sossidou, E. Growth Performance, Gut Health, Welfare and Qualitative Behavior Characteristics of Broilers Fed Diets Supplemented with Dried Common (Olea europaea) Olive Pulp. Sustainability 2023, 15, 501. https://0-doi-org.brum.beds.ac.uk/10.3390/su15010501
Dedousi A, Kotzamanidis C, Kritsa M-Z, Tsoureki A, Andreadelli A, Patsios SI, Sossidou E. Growth Performance, Gut Health, Welfare and Qualitative Behavior Characteristics of Broilers Fed Diets Supplemented with Dried Common (Olea europaea) Olive Pulp. Sustainability. 2023; 15(1):501. https://0-doi-org.brum.beds.ac.uk/10.3390/su15010501
Chicago/Turabian StyleDedousi, Anna, Charalampos Kotzamanidis, Maria-Zoi Kritsa, Antiopi Tsoureki, Aggeliki Andreadelli, Sotiris I. Patsios, and Evangelia Sossidou. 2023. "Growth Performance, Gut Health, Welfare and Qualitative Behavior Characteristics of Broilers Fed Diets Supplemented with Dried Common (Olea europaea) Olive Pulp" Sustainability 15, no. 1: 501. https://0-doi-org.brum.beds.ac.uk/10.3390/su15010501