
Pollen and Molecular Biomarkers from Sedimentary Archives in the Central Po Plain (N Italy): Assessing Their Potential to Deepen Changes in Natural and Agricultural Systems



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
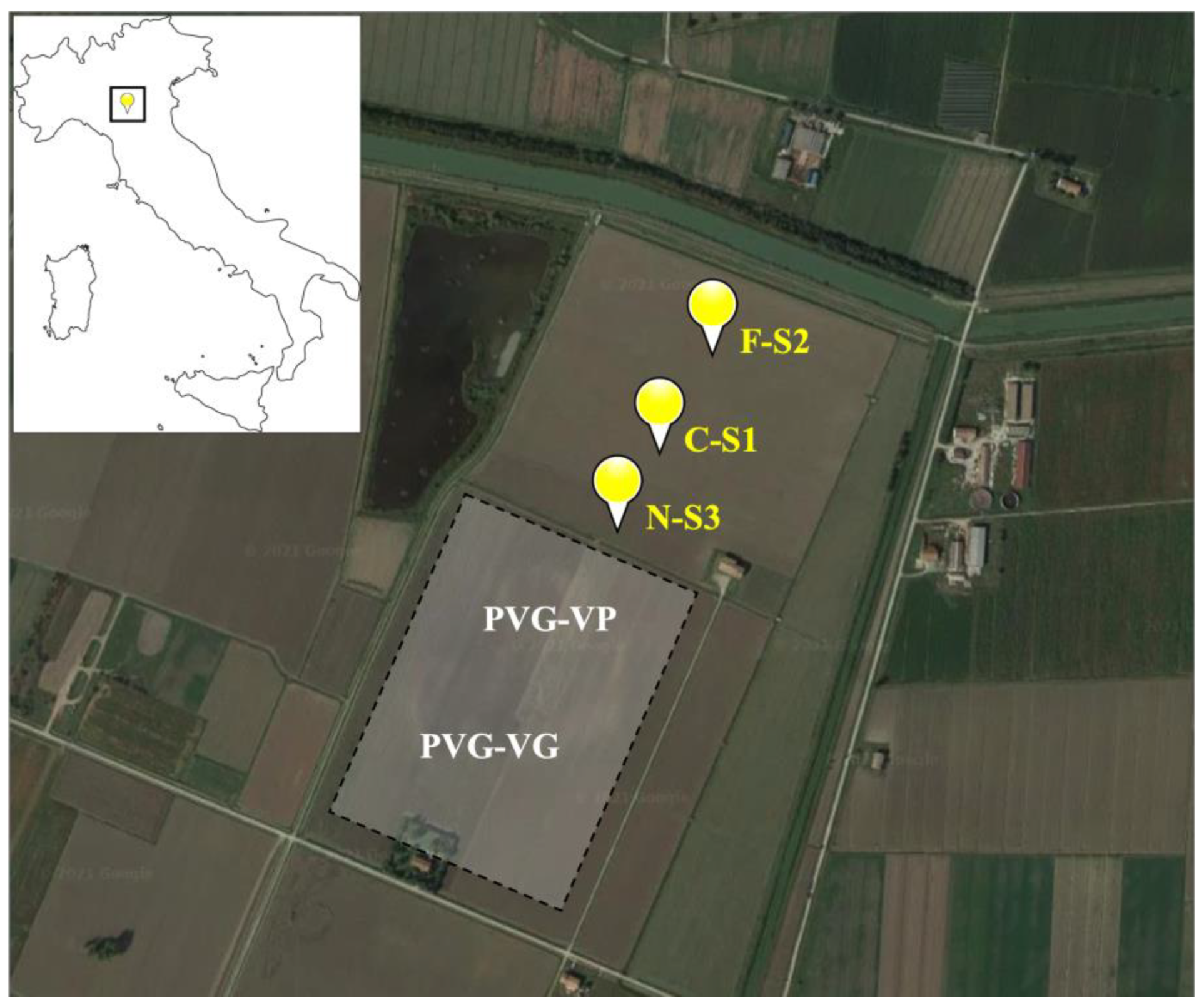
2.1. Study Area
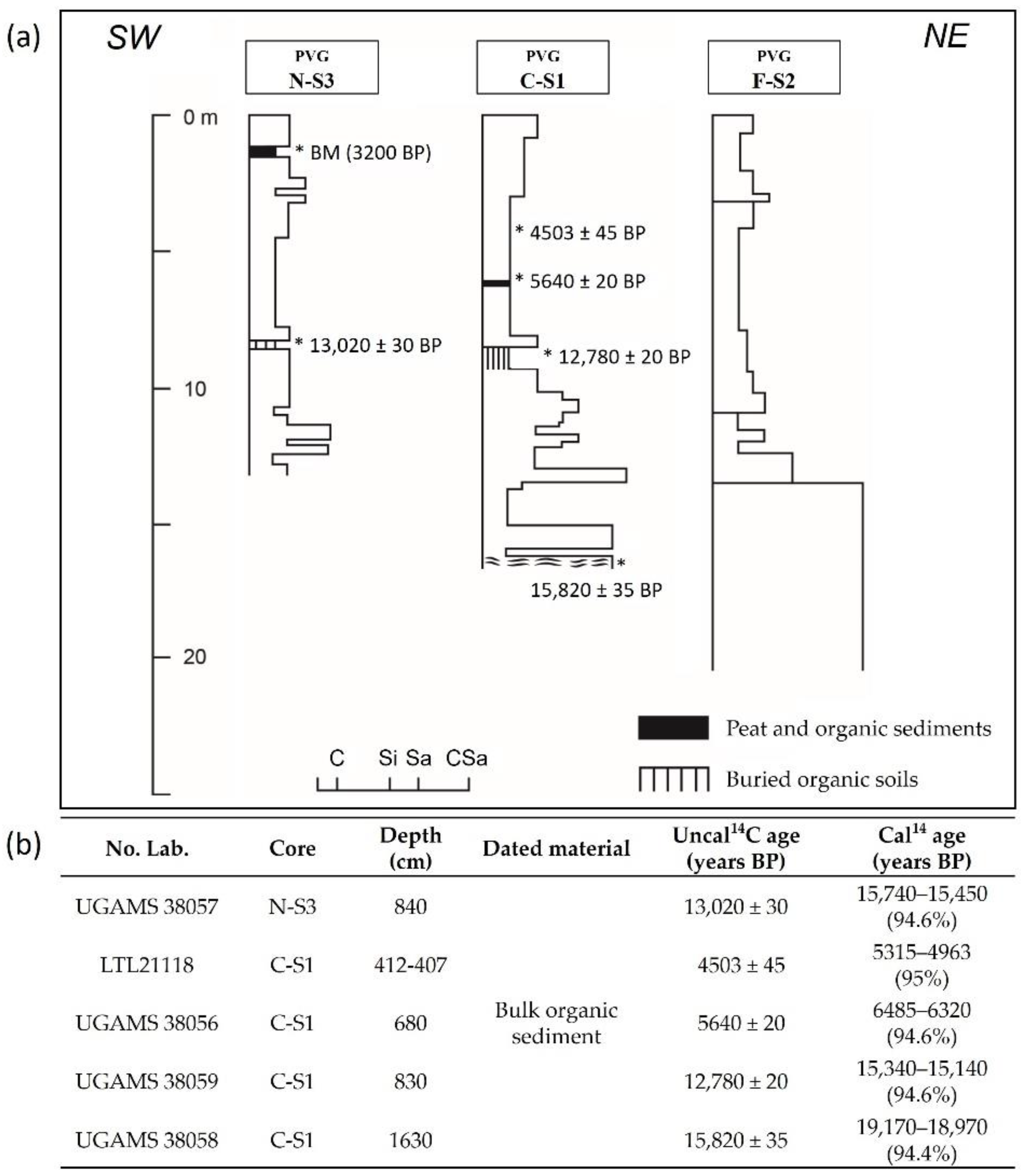
2.2. Sediment Samples
2.3. Pollen Treatment and Analysis
2.4. Biomolecular Analysis
3. Results
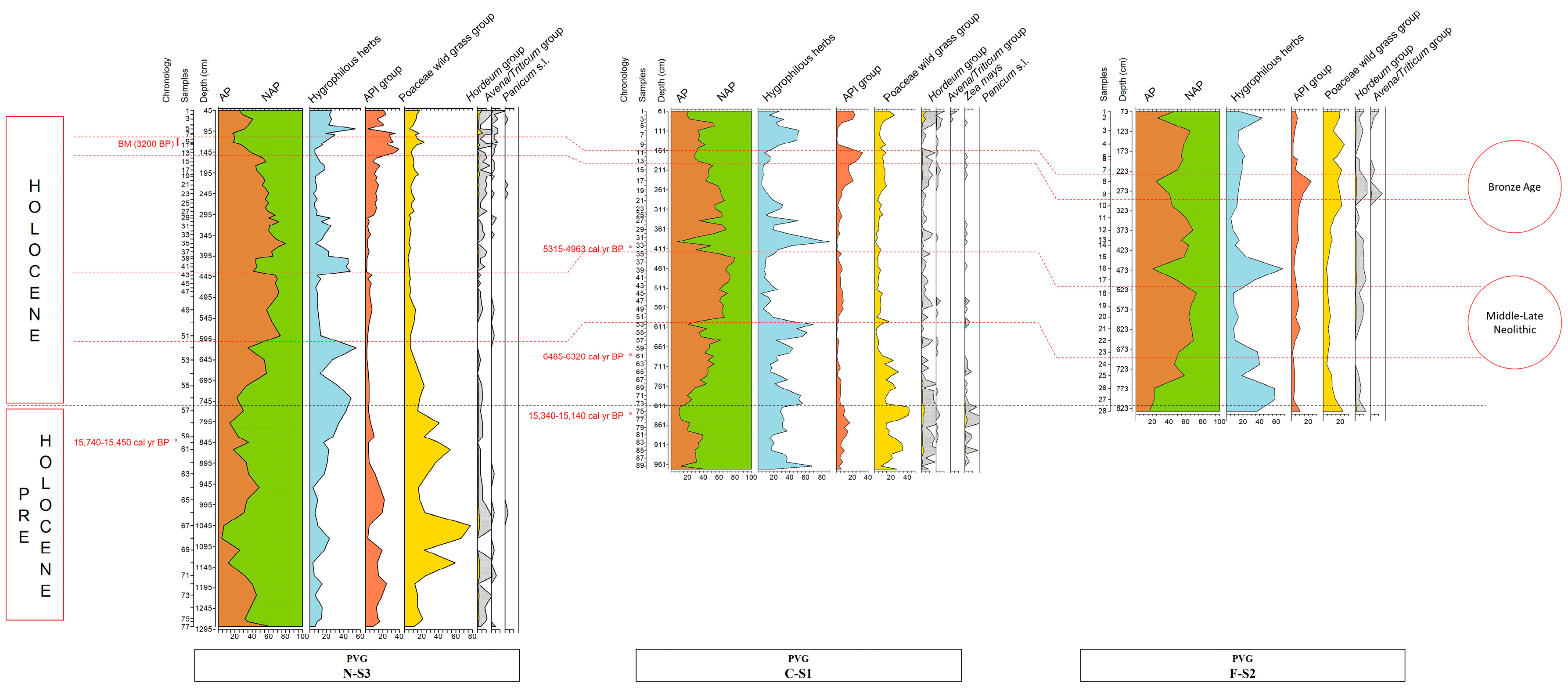
3.1. Pollen of P. Miliaceum and Other Poaceae
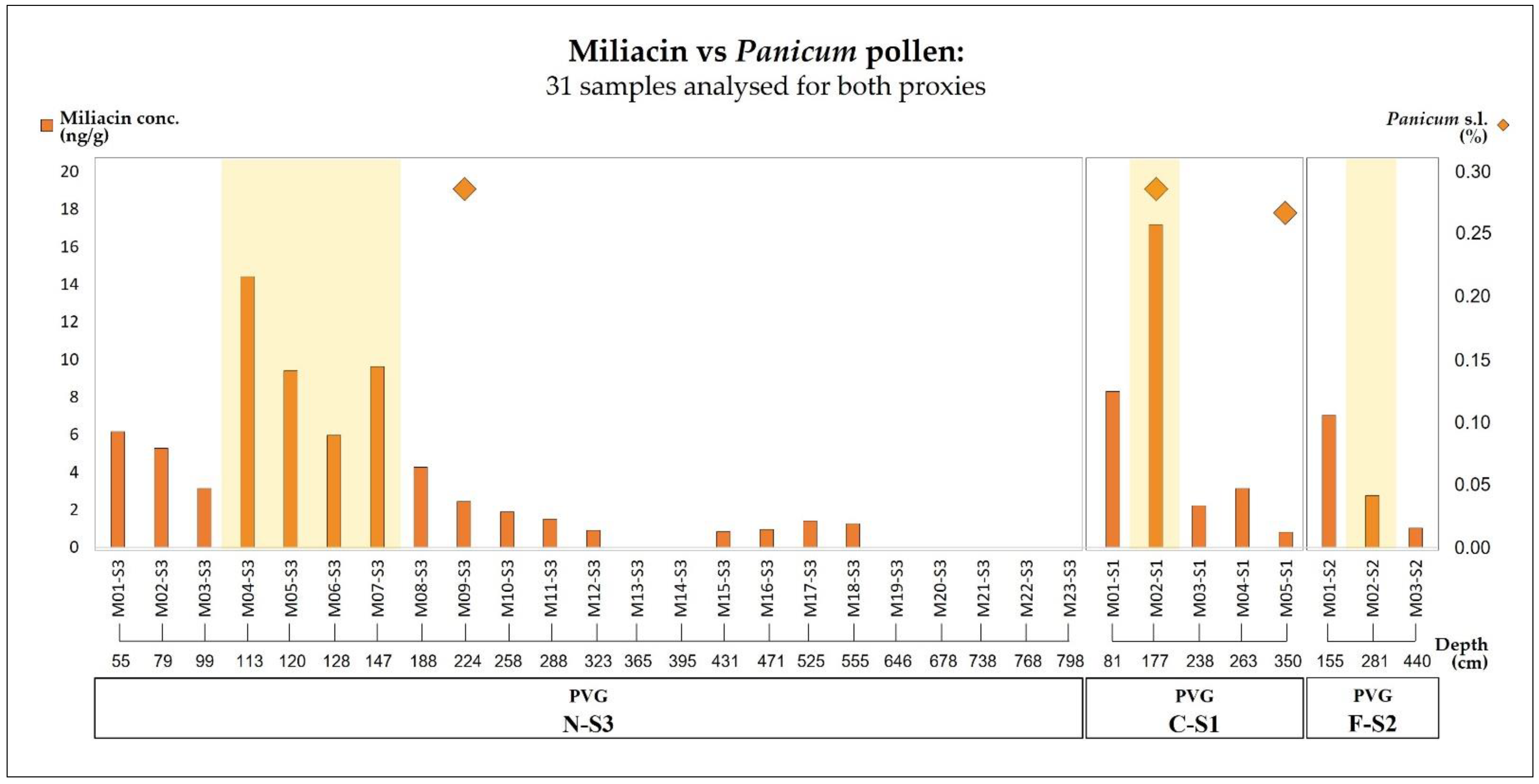
3.2. Molecular Biomarkers
4. Discussion
4.1. The Dualistic Interpretation of the Open Environment Inferred from Poaceae Pollen Curves
4.2. The Combined Evidence of Panicum Pollen and Sedimentary Miliacin during the Holocene
4.3. Wild-Cereal Hints from Large-Size Cereal Pollen (Pre-Holocene Phases)
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Castañeda, I.S.; Schouten, S. A review of molecular organic proxies for examining modern and ancient lacustrine environments. Quat. Sci. Rev. 2011, 30, 2581–2891. [Google Scholar] [CrossRef]
- Dubois, N.; Jacob, J. Molecular Biomarkers of Anthropic Impacts in Natural Archives: A Review. Front. Ecol. Evol. 2016, 4, 92. [Google Scholar] [CrossRef] [Green Version]
- Jansen, B.; Hooghiemstra, H.; de Goede, S.P.C.; van Mourik, J.M. Chapter 5—Biomarker analysis of soil archives. In Developments in Quaternary Sciences; van Mourik, J.M., Van Der Meer, J.J.M., Eds.; Elsevier: Amsterdam, The Netherlands, 2019; Volume 18, pp. 163–222. [Google Scholar]
- Zhang, Y.; Peng, T.; Yu, J.; Zhengwen, L. Natural and anthropogenically driven changes in the n-alkanols of lake sediments and implications for their use in paleoenvironmental studies of lakes. Catena 2021, 207, 105591. [Google Scholar] [CrossRef]
- Jansen, B.; van Loon, E.E.; Hooghiemstra, H.; Verstraten, J.B. Improved reconstruction of palaeo-environments through unravelling of preserved vegetation biomarker patterns. Palaeogeogr. Palaeoclimatol. Palaeoecol. 2010, 285, 119–130. [Google Scholar] [CrossRef]
- Brocks, J.; Summons, R. Sedimentary hydrocarbons, biomarkers for early life. In Treatise on Geochemistry; Schlesinger, W.H., Ed.; Elsevier: Oxford, UK, 2003; Volume 8, pp. 63–115. [Google Scholar]
- Simoneit, B.R.T.; Grimalt, J.O.; Wang, T.G.; Cox, R.E.; Hatcher, P.G.; Nissenbaum, A. Cyclic terpenoids of contemporary resinous plant detritus and of fossil woods, ambers and coals. Org. Geochem. 1985, 10, 877–889. [Google Scholar] [CrossRef]
- Abe, I.; Rohmer, M.; Prestwich, G.D. Enzymatic cyclization of squalene and oxidosqualene to sterols and triterpens. Chem. Rev. 1993, 93, 2189–2208. [Google Scholar] [CrossRef]
- Jacob, J.; Disnar, J.-R.; Boussafir, M.; Spadano Albuquerque, A.L.; Sifeddine, A.; Turcq, B. Pentacyclic triterpene methyl ethers in recent lacustrine sediments (Lagoado Caçó, Brazil). Org. Geochem. 2005, 36, 449–461. [Google Scholar] [CrossRef] [Green Version]
- Lavrieux, M.; Jacob, J.; Le Milbeau, C.; Zocatelli, R.; Masuda, K.; Bréheret, J.G.; Disnar, J.R. Occurrence of triterpenyl acetates in soil and their potential as chemotaxonomical markers of Asteraceae. Org. Geochem. 2011, 42, 1315–1323. [Google Scholar] [CrossRef] [Green Version]
- Bossard, N.; Jacob, J.; Le Milbeau, C.; Sauze, J.; Terwilliger, V.; Poissonnier, B.; Vergès, E. Distribution of miliacin (olean-18-en-3β-ol methyl ether) and related compounds in broomcorn millet (Panicum miliaceum) and other reputed sources: Implications for the use of sedimentary miliacin as a tracer of millet. Org. Geochem. 2013, 63, 48–55. [Google Scholar] [CrossRef] [Green Version]
- Jacob, J.; Disnar, J.-R.; Arnaud, F.; Chapron, E.; Debret, M.; Lallier-Vergès, E.; Desmet, M.; Revel-Rolland, M. Millet cultivation history in the French Alps as evidenced by a sedimentary molecule. J. Archaeol. Sci. 2008, 35, 814–820. [Google Scholar] [CrossRef] [Green Version]
- Jacob, J.; Disnar, J.-R.; Bardoux, G. Carbon isotope evidence for sedimentary miliacin as a tracer of Panicum miliaceum (broomcorn millet) in the sediments of Lake le Bourget (French Alps). Org. Geochem. 2008, 39, 1077–1080. [Google Scholar] [CrossRef] [Green Version]
- Jacob, J.; Simonneau, A.; Bajard, M.; Messager, E.; Billaud, Y.; Le Milbeau, C.; Poulenard, J.; Giguet-Covex, C.; Rapuc, W.; Doyen, E.; et al. Diffusion et dynamique de la culture du millet dans les Alpes ouest-européennes sur la base des concentrations en miliacine dans les sédiments lacustres. In Actes des 41èmes Rencontres Internationales d’Archéologie et d’Histoire—Biodiversités, Environnements et Sociétés Depuis la Préhistoire: Nouveaux Marqueurs et Approches Intégrées; Nicoud, E., Balasse, M., Desclaux, E., Théry-Parisot, I., Eds.; Éditions APDCA: Antibes, France, 2021; pp. 123–134. [Google Scholar]
- Rajasekaran, R.; Francis, N.; Mani, V.; Ganesan, J. Proso millet (Panicum miliaceum L.). In Neglected and Underutilized Crops: Future Smart Food; Farooq, M., Siddique, K.H.M., Eds.; Academic Press: London, UK, 2023; pp. 247–278. [Google Scholar] [CrossRef]
- Miller, N.F.; Spengler, R.N.; Frachetti, M. Millet cultivation across Eurasia: Origins, spread, and the influence of seasonal climate. Holocene 2016, 26, 1566–1575. [Google Scholar] [CrossRef] [Green Version]
- Hunt, H.V.; Vander Linden, M.; Liu, X.; Motuzaite-Matuzeviciute, G.; College, S.; Jones, M.K. Millet across Eurasia: Chronology and context of early records of the genera Panicum and Setaria from archaeological sites in the Old World. Veg. Hist. Archaeobotany 2008, 17, 5–18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Motuzaite-Matuzeviciute, G.; Staff, R.A.; Hunt, H.V.; Liu, X.; Jones, M.K. The early chronology of broomcorn millet (Panicum miliaceum) in Europe. Antiquity 2013, 338, 1073–1085. [Google Scholar] [CrossRef] [Green Version]
- Stika, H.-P.; Heiss, A.G. Plant cultivation in the bronze age. In The Oxford Handbook of the European Bronze Age; Fokkens, H., Harding, A., Eds.; Oxford University Press: Oxford, UK, 2013; pp. 369–384. [Google Scholar]
- Filipović, D.; Meadows, J.; Dal Corso, M.; Kirleis, W.; Alsleben, A.; Akeret, Ö.; Bittmann, F.; Bosi, G.; Ciută, B.; Dreslerová, D.; et al. New AMS 14C dates track the arrival and spread of broomcorn millet cultivation and agricultural change in prehistoric Europe. Sci. Rep. 2020, 10, 13698. [Google Scholar] [CrossRef]
- Ohmoto, T.; Ikuse, M.; Natori, S. Triterpenoids of the Gramineae. Phytochemistry 1970, 6, 2137–2148. [Google Scholar] [CrossRef]
- Lu, H.; Zhang, J.; Wu, N.; Liu, K.-B.; Xu, D.; Li, Q. Phytoliths Analysis for the Discrimination of Foxtail Millet (Setaria italica) and Common Millet (Panicum miliaceum). PLoS ONE 2009, 4, e4448. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Out, W.A.; Madella, M. Morphometric distinction between bilobate phytoliths from Panicum miliaceum and Setaria italica leaves. Archaeol. Anthropol. Sci. 2016, 8, 505–521. [Google Scholar] [CrossRef]
- Dal Corso, M.; Nicosia, C.; Balista, C.; Cupitò, M.; Dalla Longa, E.; Leonardi, G.; Kirleis, W. Bronze Age crop processing evidence in the phytolith assemblages from the ditch and fen around Fondo Paviani, northern Italy. Veg. Hist. Archaeobotany 2017, 26, 5–24. [Google Scholar] [CrossRef]
- Faegri, K.; Iversen, J. Textbook of Pollen Analysis; John Wiley & Sons: Chichester, UK, 1989. [Google Scholar]
- Miras, Y.; Guenet, P.; Richard, H. Holocene vegetation, landscape, and reconstruction of human activity from prehistory to the roman period based on new pollen data performed in “the plateau de Millevaches” (Limousin, Massif Central, France). Quaternaire 2011, 22, 147–164. [Google Scholar] [CrossRef] [Green Version]
- Mercuri, A.M. Genesis and evolution of the cultural landscape in central Mediterranean: The ‘where, when and how’ through the palynological approach. Landsc. Ecol. 2014, 29, 1799–1810. [Google Scholar] [CrossRef]
- Hjelle, K.; Overland, A.; Gran, M.M.; Romundset, A.; Ystgaard, I. Two thousand years of Landscape—Human interactions at a coastal peninsula in Norway revealed through pollen analysis, shoreline reconstruction, and radiocarbon dates from archaeological sites. Front. Ecol. Evol. 2022, 10, 911780. [Google Scholar] [CrossRef]
- Vignola, C.; Hättestrand, M.; Bonnier, A.; Finné, M.; Izdebski, A.; Katrantsiotis, C.; Kouli, K.; Liakopoulos, G.C.; Norström, E.; Papadaki, M.; et al. Mid-late Holocene vegetation history of the Argive Plain (Peloponnese, Greece) as inferred from a pollen record from ancient Lake Lerna. PLoS ONE 2022, 17, e0271548. [Google Scholar] [CrossRef]
- Sadori, L.; Mercuri, A.M.; Mariotti Lippi, M. Reconstructing past cultural landscape and human impact using pollen and plant macroremains. Plant Biosyst. 2010, 144, 940–951. [Google Scholar] [CrossRef]
- Mercuri, A.M.; Florenzano, A.; Burjachs, F.; Giardini, M.; Kouli, K.; Masi, A.; Picornell-Gelabert, L.; Revelles, J.; Servera-Vives, G.; Sadori, L.; et al. From influence to impact: The multifunctional land-use in Mediterranean prehistory emerging from palynology of Archaeological sites (8.2–2.8 ka BP). Holocene 2019, 29, 830–846. [Google Scholar] [CrossRef] [Green Version]
- Revelles, J. The Role of Palynology in Archaeoecological Research: Reconstructing Human-Environment Interactions during Neolithic in the Western Mediterranean. Appl. Sci. 2021, 11, 4073. [Google Scholar] [CrossRef]
- Baltensperger, D.D. Foxtail and proso millet. In Progress in New Crops; Janick, J., Ed.; ASHS Press: Alexandria, VA, USA, 1996; pp. 182–190. [Google Scholar]
- Mercuri, A.M.; Accorsi, C.A.; Bandini Mazzanti, M.; Bosi, G.; Trevisan Grandi, G.; Cardarelli, A.; Labate, D.; Marchesini, M.; Olmi, L.; Torri, P. Cereal fields from the middle-recent Bronze Age, as found in the Terramara di Montale, in the Po Plain (Emilia-Romagna, Northern Italy), based on pollen, seeds/fruits and microcharchoals. In The Archaeology of Crop Fields and Gardens; Morel, J.-P., Tresserras, J., Matalama, J.C., Eds.; Edipuglia: Bari, Italy, 2006; pp. 251–270. [Google Scholar]
- Clò, E. Climate Change and Human Impact in a Long-Term Perspective: Palynology of Central Po Plain Deposits Since the Lateglacial. Ph.D. Thesis, Università degli Studi di Modena e Reggio Emilia, Modena, Italy, 2022. [Google Scholar]
- Mercuri, A.M.; Accorsi, C.A.; Bandini Mazzanti, M.; Bosi, G.; Cardarelli, A.; Labate, D.; Marchesini, M.; Trevisan Grandi, G. Economy and environment of Bronze Age settlements e Terramaras e in the Po Plain (Northern Italy): First results of the archaeobotanical research at the Terramara di Montale. Veg. Hist. Archaeobotany 2006, 16, 43–60. [Google Scholar] [CrossRef]
- Cremaschi, M.; Mercuri, A.M.; Torri, P.; Florenzano, A.; Pizzi, C.; Marchesini, M.; Zerboni, A. Climate change versus land management in the Po Plain (Northern Italy) during the Bronze Age: New insights from the VP/VG sequence of the Terramara Santa Rosa di Poviglio. Quat. Sci. Rev. 2016, 136, 153–172. [Google Scholar] [CrossRef]
- Dal Corso, M.; Marchesini, M.; Leonardi, G.; Kirleis, W. Environmental changes and human impact during Bronze Age in northern Italy: On-site palynological investigation at Fondo Paviani (Legnago, Verona). In Collapse or Continuity? Environment and Development of Bronze Age Human Landscapes. Proceedings of the International Workshop “Socio-Environmental Dynamics over the Last 12,000 Years: The Creation of Landscapes II (14–18 March 2011)” in Kiel; Kneisel, J., Kirleis, W., Dal Corso, M., Taylor, N., Tiedtke, V., Eds.; Kiel Archaeology: Kiel, Germany, 2012; Volume 1, pp. 71–84. [Google Scholar]
- Dal Corso, M.; Zanon, M.; Heron, C.; Rottoli, M.; Cupitò, M.; Dalla Longa, E.; Kirleis, W. Tracing millet through biomarker analysis in archaeological sites in alluvial plains: The first miliacin data from the northern Italian Bronze Age. In Millet and What Else? The Wider Context of the Adoption of Millet Cultivation in Europe; Kirleis, W., Dal Corso, M., Filipović, D., Eds.; Sidestone Press: Leiden, The Netherlands, 2022; pp. 231–252. [Google Scholar]
- Brandolini, F.; Cremaschi, M. The Impact of Late Holocene Flood Management on the Central Po Plain (Northern Italy). Sustainability 2018, 10, 3968. [Google Scholar] [CrossRef] [Green Version]
- Valsecchi, V.; Tinner, W.; Finsinger, W.; Ammann, B. Human impact during the Bronze Age in the vegetation at Lago Lucone (northern Italy). Veg. Hist. Archaeobotany 2006, 15, 99–113. [Google Scholar] [CrossRef] [Green Version]
- Vescovi, E.; Kaltenrieder, P.; Tinner, W. Late-glacial and Holocene vegetation history of Pavullo nel Frignano (Northern Apennines, Italy). Rev. Palaeobot. Palynol. 2010, 160, 32–45. [Google Scholar] [CrossRef]
- Bosi, G.; Mercuri, A.M.; Bandini Mazzanti, M.; Florenzano, A.; Montecchi, M.C.; Torri, P.; Labate, D.; Rinaldi, R. The evolution of Roman urban environments through the archaeobotanical remains in Modena e Northern Italy. J. Archaeol. Sci. 2015, 53, 19–31. [Google Scholar] [CrossRef]
- Mercuri, A.M.; Bandini Mazzanti, M.; Torri, P.; Vigliotti, L.; Bosi, G.; Florenzano, A.; Olmi, L.; Massamba N’siala, I. A marine/terrestrial integration for mid-late Holocene vegetation history and the development of the cultural landscape in the Po Valley as a result of human impact and climate change. Veg. Hist. Archaeobotany 2012, 21, 353–372. [Google Scholar] [CrossRef]
- Mele, M.; Cremaschi, M.; Giudici, M.; Lozej, A.; Pizzi, C.; Bassi, A. The terramare and the surrounding hydraulic structures: A geophysical survey of the Santa Rosa site at Poviglio (Bronze Age; Northern Italy). J. Archaeol. Sci. 2013, 40, 4648–4662. [Google Scholar] [CrossRef]
- Bernabò Brea, M.; Cremaschi, M. Il Villaggio Piccolo Della Terramara di S. Rosa a Poviglio. Scavi 1987/1992; Istituto Italiano Preistoria e Protostoria: Firenze, Italy, 2004. [Google Scholar]
- Reimer, P.J.; Austin, W.E.N.; Bard, E.; Bayliss, A.; Blackwell, P.G.; Bronk Ramsey, C.; Butzin, M.; Cheng, H.; Edwards, R.L.; Friedrich, M.; et al. The IntCal20 Northern Hemisphere radiocarbon age calibration curve (0–55 kcal BP). Radiocarbon 2020, 62, 725–757. [Google Scholar] [CrossRef]
- van der Kaars, S.; Penny, D.; Tibby, J.; Fluin, J.; Dam, R.; Suparan, P. Late Quaternary palaeoecology, palynology and palaeolimnology of a tropical lowland swamp: Rawa Danau, West Java, Indonesia. Palaeogeogr. Palaeoclimatol. Palaeoecol. 2001, 171, 185–212. [Google Scholar] [CrossRef]
- Florenzano, A.; Mercuri, A.M.; Pederzoli, A.; Torri, P.; Bosi, G.; Olmi, L.; Rinaldi, R.; Bandini Mazzanti, M. The significance of intestinal parasite remains in pollen samples from Medieval pits in the Piazza Garibaldi of Parma, Emilia Romagna, Northern Italy. Geoarchaeology 2012, 27, 34–47. [Google Scholar] [CrossRef]
- Moore, P.D.; Webb, J.A.; Collins, M.E. Pollen Analysis, 2nd ed.; Blackwell Scientific Publications: Oxford, UK, 1991. [Google Scholar]
- Reille, M. Pollen et Spores d’Europe et d’Afrique du Nord; Laboratoire de Botanique Historique et Palinologie: Marseille, France, 1992. [Google Scholar]
- Beug, H.J. Leitfaden der Pollenbestimmung für Mitteleuropa und Angrezende Gebiete; Gustav Fischer: Stuttgart, Germany, 1961. [Google Scholar]
- Andersen, S.T. Identification of Wild Grasses and Cereal Pollen; Danmarks Geologiske Undersøgelse: Copenhagen, Denmark, 1979. [Google Scholar]
- Joly, C.; Barillé, L.; Barreau, M.; Mancheron, A.; Visset, L. Grain and annulus diameter as criteria for distinguishing pollen grains of cereals from wild grasses. Rev. Palaeobot. Palynol. 2007, 146, 221–233. [Google Scholar] [CrossRef]
- Beug, H.J. Leitfaden der Pollenbestimmung für Mitteleuropa und Angrenzende Gebiete; Dr. Friedrich Pfeil: Munich, Germany, 2015. [Google Scholar]
- Mercuri, A.M. Palynological analysis of the Early Holocene sequence. In The Uan Afuda Cave Hunter-gatherer Societies of Central Sahara, Arid Zone Archaeology, Monographs 1; Di Lernia, S., Ed.; All’Insegna del Giglio: Florence, Italy, 1999; pp. 149–181. [Google Scholar]
- Grimm, E.C. TILIA and TGView; Illinois State Museum: Springfield, IL, USA, 2004. [Google Scholar]
- Mercuri, A.M.; Bandini Mazzanti, M.; Florenzano, A.; Montecchi, M.C.; Rattighieri, E.; Torri, P. Anthropogenic Pollen Indicators (API) from archaeological sites as local evidence of human-induced environments in the Italian peninsula. Ann. Bot. (Rome) 2013, 3, 143–153. [Google Scholar] [CrossRef]
- Shiojima, K.; Masuda, K.; Lin, T.; Suzuki, H.; Ageta, H.; Inoue, M.; Ishida, T. Composite constituents: Three gammacer-16-ene derivatives, novel triterpenoids isolated from roots of Picris hieracioides L. subsp. japonica. Tetrahedron Lett. 1989, 30, 4977–4980. [Google Scholar] [CrossRef]
- Shiojima, K.; Masuda, K.; Ooishi, Y.; Suzuki, H.; Ageta, H. Composite constituents: New migrated gammacerane triterpenoids from roots of Picris hieracioides L. subsp. japonica. Tetrahedron Lett. 1989, 30, 6873–6874. [Google Scholar] [CrossRef]
- Shiojima, K.; Masuda, K.; Suzuki, H.; Lin, T.; Ooishi, Y.; Ageta, H. Composite constituents: Forty-two triterpenoids including eight novel compounds isolated from Picris hieracioides subsp. japonica. Chem. Pharm. Bull. 1995, 43, 1634–1639. [Google Scholar] [CrossRef] [Green Version]
- Jakupovic, J.; Tsichrintzis, F.; Zdero, C.; Hahn, A.; Lipp, W.; Smalla, B.; Berendsohn, W.G. Natural Substances in the Compositae: The Bohlmann Files. Available online: http://www.bgbm.org/BioDivInf/projects/bohlmannfiles/ (accessed on 4 December 2011).
- Lavrieux, M. Biomarqueurs Moléculaires d’Occupation des Sols, du Sol au Sédiment: Exemple du Bassin-Versant et du Lac d’Aydat (Puy-de-Dôme). Ph.D. Thesis, Université de Tours, Tours, France, 2011. [Google Scholar]
- Ravazzi, C.; Peresani, M.; Pini, R.; Vescovi, E. Il Tardoglaciale nelle alpi italiane e in pianura padana. Evoluzione stratigrafica, storia della vegetazione e del popolamento antropico. Alp. Mediterr. Quat. 2007, 20, 163–184. [Google Scholar]
- Ravazzi, C.; Deaddis, M.; De Amicis, M.; Marchetti, M.; Vezzoli, G.; Zanchi, A. The last 40 ka evolution of the Central Po Plain between the Adda and Serio rivers. Géomorphologie 2012, 18, 131–154. [Google Scholar] [CrossRef] [Green Version]
- Vittori Antisari, L.; Bianchini, G.; Cremonini, S.; Di Giuseppe, D.; Falsone, G.; Marchesini, M.; Marvelli, S.; Vianello, G. Multidisciplinary study of a Lateglacial-Holocene sedimentary sequence near Bologna (Italy): Insights on natural and anthropogenic impacts on the landscape dynamics. J. Soils Sediments 2016, 16, 645–662. [Google Scholar] [CrossRef]
- Pignatti, S. Flora d’Italia; Edagricole: Bologna, Italy, 1982. [Google Scholar]
- Ravazzi, C.; Donegana, M.; Vescovi, E.; Arpenti, E.; Caccianiga, M.; Kaltenrieder, P.; Londeix, L.; Marabini, S.; Mariani, S.; Pini, R.; et al. A new Lateglacial site with Picea abies in the Northern Apennine foothills: A population failing the model of glacial refugia trees. Veg. Hist. Archaeobotany 2006, 15, 357–371. [Google Scholar] [CrossRef]
- Clò, E.; Florenzano, A. Heterocysts of Rivularia Type for Interpreting a Palaeoenvironmental Context of the Late Quaternary in Northern Italy. Sustainability 2022, 14, 15332. [Google Scholar] [CrossRef]
- Magny, M. Holocene climatic variability as reflected by mid-European lake-level fluctuations, and its probable impact on prehistoric human settlements. Quat. Int. 2004, 113, 65–79. [Google Scholar] [CrossRef]
- Bottazzi, G.; Bronzoni, L.; Mutti, A. Carta Archeologica del Comune di Poviglio 1986–1989; IRIS: Poviglio, Italy, 1995. [Google Scholar]
- Mutti, A. Gli scavi nella terramara Santa Rosa a Fodico di Poviglio. In Guida All’esposizione; Fondazione P. Manodori: Reggio Emilia, Italy, 2007. [Google Scholar]
- Brandolini, F.; Trombino, L.; Sibilia, E.; Cremaschi, M. Micromorphology and site formation processes in the Castrum Popilii Medieval Motte (N Italy). J. Archaeol. Sci. Rep. 2018, 20, 18–32. [Google Scholar] [CrossRef]
- Cazzola, F. L’introduzione del mais in Italia e la sua utilizzazione alimentare (sec. XVI–XVIII). In La Préparation Alimentaire des Céréales; Fournier, S., Sigaut, F., Eds.; Centro Universitario Europeo per i Beni Culturali: Ravello, Italy, 1992; pp. 109–127. [Google Scholar]
- Zohary, D.; Hopf, M.; Weiss, E. Domestication of Plants in the Old World, 4th ed.; Clarendon Press: Oxford, UK, 2012. [Google Scholar]
- Rottoli, M.; Pessina, A. Neolithic agriculture in Italy: An update of archaeobotanical data with particular emphasis on northern settlements. In The Origins and Spread of Domestic Plants in Southwest Asia and Europe; Colledge, S., Conolly, J., Eds.; Routledge: London, UK, 2016; pp. 141–154. [Google Scholar]
- Kirleis, W. Atlas of Neolithic Plant Remains from Northern Central Europe; Barkhuis Publishing: Groningen, The Netherlands, 2019. [Google Scholar]
- Liu, X.Y.; Motuzaite-Matuzeviciute, G.; Hunt, H.V. From a fertile idea to a fertile arc: The origins of broomcorn millet 15 years on. In Far from the Hearth: Essays in Honour of Martin K. Jones; Lightfoot, E., Liu, X.Y., Fuller, D.Q., Eds.; McDonald Institute of Archaeology: Cambridge, UK, 2019; pp. 155–164. [Google Scholar]
- Tafuri, M.A.; Craig, O.E.; Canci, A. Stable isotope evidence for the consumption of millet and other plants in Bronze Age Italy. Assoc. Phys. Anthropol. 2009, 139, 146–153. [Google Scholar] [CrossRef]
- Chiaffarelli, G.; Indipolage Kariyawasam, T.; Proserpio, B.; Rottoli, M. Indipolage Kariyawasam, T.; Proserpio, B.; Rottoli, M. I macroresti vegetali nelle strutture al margine del Villaggio Grande della Terramara Santa Rosa di poviglio. In Terramara Santa Rosa di Poviglio—Le Strutture Idrauliche al Margine del Villaggio Grande. Scavi 1998–2011; Cremaschi, M., Pizzi, C., Eds.; Istituto Italiano di Preistoria e Protostoria: Florence, Italy, 2021; Origines, 35; pp. 389–401. [Google Scholar]
- Florenzano, A.; Clò, E.; Zappa, J.; Montecchi, M.C.; Furia, E.; Torri, P.; Mercuri, A.M. Paesaggio vegetale sulla base delle analisi del riempimento dei pozzi al margine del Villaggio Grande della Terramara Santa Rosa di Poviglio. In Terramara Santa Rosa di Poviglio—Le Strutture Idrauliche al Margine del Villaggio Grande. Scavi 1998–2011; Cremaschi, M., Pizzi, C., Eds.; Istituto Italiano di Preistoria e Protostoria: Florence, Italy, 2021; Origines, 35; pp. 373–387. [Google Scholar]
- Dal Corso, M. Environmental History and Development of the Human Landscape in a Northeastern Italian Lowland during the Bronze Age: A Multidisciplinary Case-Study. In Universitätsforschungen zur Prehistorischen Archäologie—Band 312, HDL 12; Dr. Rudolf Habelt GmbH: Bonn, Germany, 2018. [Google Scholar]
- Fundurulic, A.; Valenti, I.; Celant, A.; Barbaro, B.; Costa, M.; Manhita, A.; Severi, E.; Barrocas Dias, C.; Magri, D. Millets and Cereal Meals from the Early Iron Age Underwater Settlement of “Gran Carro” (Bolsena Lake, Central Italy). Sustainability 2022, 14, 3941. [Google Scholar] [CrossRef]
- Castiglioni, E.; Rottoli, M. Broomcorn millet, foxtail millet and sorghum in north Italian Early Medieval sites. PCA 2013, 3, 131–144. [Google Scholar]
- Murphy, C. Finding millet in the Roman world. Archaeol. Anthropol. Sci. 2016, 8, 65–78. [Google Scholar] [CrossRef] [Green Version]
- Bosi, G.; Castiglioni, E.; Rinaldi, R.; Mazzanti, M.; Marchesini, M.; Rottoli, M. Archaeobotanical evidence of food plants in Northern Italy during the Roman period. Veg. Hist. Archaeobotany 2020, 29, 681–697. [Google Scholar] [CrossRef]
- Ganzarolli, G.; Alexander, M.; Chavarria, A.; Oliver, A.; Craig, E. Direct evidence from lipid residue analysis for the routine consumption of millet in Early Medieval Italy. J. Archaeol. Sci. 2018, 96, 124–130. [Google Scholar] [CrossRef]
- Mercuri, A.M.; Fornaciari, R.; Gallinaro, M.; Vanin, S.; di Lernia, S. Plant behaviour from human imprints and the cultivation of wild cereals in Holocene Sahara. Nat. Plants 2018, 4, 71–81. [Google Scholar] [CrossRef] [Green Version]
- Andrieu-Ponel, V.; Rochette, P.; Demory, F.; Alçiçek, H.; Boulbes, N.; Bourlès, D.; Helvacı, C.; Lebatard, A.-E.; Mayda, S.; Michaud, H.; et al. Continuous presence of proto-cereals in Anatolia since 2.3 Ma, and their possible co-evolution with large herbivores and hominins. Sci. Rep. 2021, 11, 8914. [Google Scholar] [CrossRef]
- Ejsmond, M.J.; Wrońska-Pilarek, D.; Ejsmond, A.; Dragosz-Kluska, D.; Karpińska-Kołaczek, M.; Kołaczek, P.; Kozłowski, J. Does climate affect pollen morphology? Optimal size and shape of pollen grains under various desiccation intensity. Ecosphere 2011, 2, 117. [Google Scholar] [CrossRef]
- Manzaneda, A.J.; Rey, P.J.; Bastida, J.M.; Weiss-Lehman, C.; Raskin, E.; Mitchell-Olds, T. Environmental aridity is associated with cytotype segregation and polyploidy occurrence in Brachypodium distachyon (Poaceae). New Phytol. 2012, 193, 797–805. [Google Scholar] [CrossRef] [Green Version]
- Fox, D.T.; Soltis, D.E.; Soltis, P.S.; Ashman, T.-L.; Van de Peer, Y. Polyploidy: A biological force from cells to ecosystems. Trends Cell Biol. 2020, 30, 688–694. [Google Scholar] [CrossRef]
- Mercuri, A.M.; Clò, E.; Florenzano, A. Multiporate Pollen of Poaceae as Bioindicator of Environmental Stress: First Archaeobotanical Evidence from the Early–Middle Holocene Site of Takarkori in the Central Sahara. Quaternary 2022, 5, 41. [Google Scholar] [CrossRef]






Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Florenzano, A.; Clò, E.; Jacob, J. Pollen and Molecular Biomarkers from Sedimentary Archives in the Central Po Plain (N Italy): Assessing Their Potential to Deepen Changes in Natural and Agricultural Systems. Sustainability 2023, 15, 10408. https://0-doi-org.brum.beds.ac.uk/10.3390/su151310408
Florenzano A, Clò E, Jacob J. Pollen and Molecular Biomarkers from Sedimentary Archives in the Central Po Plain (N Italy): Assessing Their Potential to Deepen Changes in Natural and Agricultural Systems. Sustainability. 2023; 15(13):10408. https://0-doi-org.brum.beds.ac.uk/10.3390/su151310408
Chicago/Turabian StyleFlorenzano, Assunta, Eleonora Clò, and Jérémy Jacob. 2023. "Pollen and Molecular Biomarkers from Sedimentary Archives in the Central Po Plain (N Italy): Assessing Their Potential to Deepen Changes in Natural and Agricultural Systems" Sustainability 15, no. 13: 10408. https://0-doi-org.brum.beds.ac.uk/10.3390/su151310408