
Physiological, Morphological, and Biochemical Responses of Soybean [Glycine max (L.) Merr.] to Loquat (Eriobotrya japonica Lindl.) Leaf Extract Application on Pb-Contaminated Soil


Abstract
:1. Introduction
2. Materials and Methods
2.1. Experimental Site
2.2. Preparation of Loquat Leaf Extract
2.3. Growth Traits and Pb Uptake
2.4. Pigments
2.5. Osmolytes
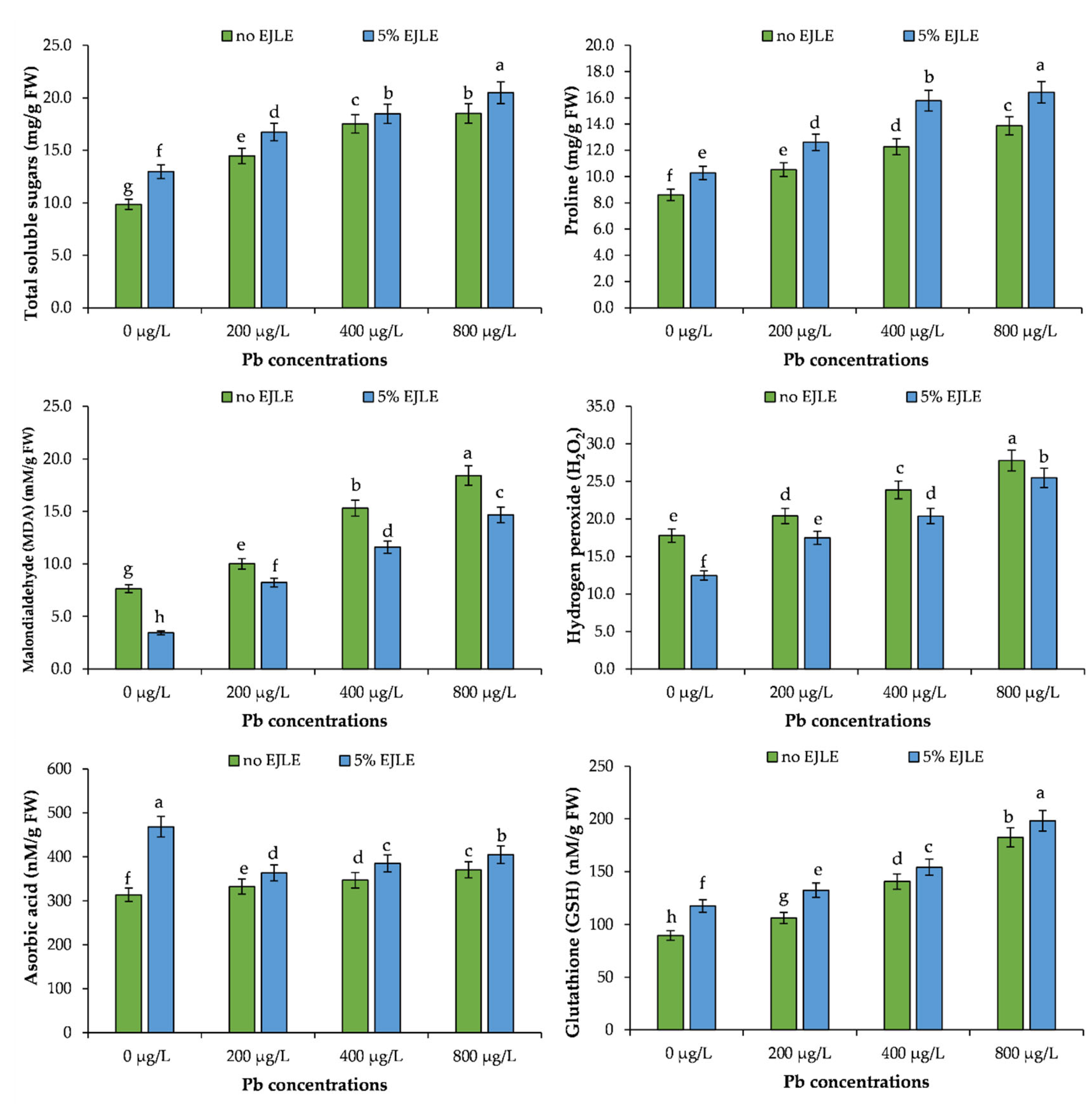
2.6. Stress Indicators
2.7. Ascorbic Acid
2.8. Glutathione, Total Phenolics, and Flavonoids
2.9. Activities of Antioxidant Enzymes
2.10. Statistical Analysis
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Masindi, V.; Muedi, K.L. Environmental Contamination by Heavy Metals. In Heavy Metals; InTech: Jakarta Selatan, Indonesia, 2018. [Google Scholar]
- Vareda, J.P.; Valente, A.J.M.; Durães, L. Assessment of Heavy Metal Pollution from Anthropogenic Activities and Remediation Strategies: A Review. J. Environ. Manag. 2019, 246, 101–118. [Google Scholar] [CrossRef] [PubMed]
- Nagajyoti, P.C.; Lee, K.D.; Sreekanth, T.V.M. Heavy Metals, Occurrence and Toxicity for Plants: A Review. Environ. Chem. Lett. 2010, 8, 199–216. [Google Scholar] [CrossRef]
- Kaya, C.; Ugurlar, F.; Ashraf, M.; Alyemeni, M.N.; Bajguz, A.; Ahmad, P. The Involvement of Hydrogen Sulphide in Melatonin-Induced Tolerance to Arsenic Toxicity in Pepper (Capsicum annuum L.) Plants by Regulating Sequestration and Subcellular Distribution of Arsenic, and Antioxidant Defense System. Chemosphere 2022, 309, 136678. [Google Scholar] [CrossRef] [PubMed]
- Kaya, C.; Ugurlar, F.; Ashraf, M.; El-Sheikh, M.A.; Bajguz, A.; Ahmad, P. The Participation of Nitric Oxide in Hydrogen Sulphide-Mediated Chromium Tolerance in Pepper (Capsicum annuum L.) Plants by Modulating Subcellular Distribution of Chromium and the Ascorbate-Glutathione Cycle. Environ. Pollut. 2022, 313, 120229. [Google Scholar] [CrossRef] [PubMed]
- Ben Massoud, M.; Sakouhi, L.; Karmous, I.; Zhu, Y.; El Ferjani, E.; Sheehan, D.; Chaoui, A. Protective Role of Exogenous Phytohormones on Redox Status in Pea Seedlings under Copper Stress. J. Plant Physiol. 2018, 221, 51–61. [Google Scholar] [CrossRef]
- Kaya, C.; Ugurlar, F.; Farooq, S.; Ashraf, M.; Alyemeni, M.N.; Ahmad, P. Combined Application of Asparagine and Thiourea Improves Tolerance to Lead Stress in Wheat by Modulating AsA-GSH Cycle, Lead Detoxification and Nitrogen Metabolism. Plant Physiol. Biochem. 2022, 190, 119–132. [Google Scholar] [CrossRef]
- Bahmani, R.; Kim, D.; Modareszadeh, M.; Hwang, S. Cadmium Enhances Root Hair Elongation through Reactive Oxygen Species in Arabidopsis. Environ. Exp. Bot. 2022, 196, 104813. [Google Scholar] [CrossRef]
- Yang, L.; Yang, H.; Bian, Z.; Lu, H.; Zhang, L.; Chen, J. The Defensive Role of Endogenous H2S in Brassica rapa against Mercury-Selenium Combined Stress. Int. J. Mol. Sci. 2022, 23, 2854. [Google Scholar] [CrossRef]
- Akhoundnejad, Y.; Karakas, O.; Demirci, O. Response of Lettuce to Silver Nanoparticles Under Drought Conditions. Iran. J. Sci. Technol. Trans. A Sci. 2022, 46, 111–120. [Google Scholar] [CrossRef]
- Kabeer, R.; Sylas, V.P.; Praveen Kumar, C.S.; Thomas, A.P.; Shanthiprabha, V.; Radhakrishnan, E.K.; Baiju, K.R. Role of Heavy Metal Tolerant Rhizosphere Bacteria in the Phytoremediation of Cu and Pb Using Eichhornia crassipes (Mart.) Solms. Int. J. Phytoremediation 2022, 24, 1120–1132. [Google Scholar] [CrossRef]
- Ahmed, S.F.; Kumar, P.S.; Rozbu, M.R.; Chowdhury, A.T.; Nuzhat, S.; Rafa, N.; Mahlia, T.M.I.; Ong, H.C.; Mofijur, M. Heavy Metal Toxicity, Sources, and Remediation Techniques for Contaminated Water and Soil. Environ. Technol. Innov. 2022, 25, 102114. [Google Scholar] [CrossRef]
- Gottesfeld, P.; Were, F.H.; Adogame, L.; Gharbi, S.; San, D.; Nota, M.M.; Kuepouo, G. Soil Contamination from Lead Battery Manufacturing and Recycling in Seven African Countries. Environ. Res. 2018, 161, 609–614. [Google Scholar] [CrossRef]
- Zulfiqar, U.; Farooq, M.; Hussain, S.; Maqsood, M.; Hussain, M.; Ishfaq, M.; Ahmad, M.; Anjum, M.Z. Lead Toxicity in Plants: Impacts and Remediation. J. Environ. Manag. 2019, 250, 109557. [Google Scholar] [CrossRef]
- Chen, F.; Aqeel, M.; Maqsood, M.F.; Khalid, N.; Irshad, M.K.; Ibrahim, M.; Akhter, N.; Afzaal, M.; Ma, J.; Hashem, M.; et al. Mitigation of Lead Toxicity in Vigna Radiata Genotypes by Silver Nanoparticles. Environ. Pollut. 2022, 308, 119606. [Google Scholar] [CrossRef]
- Amari, T.; Ghnaya, T.; Abdelly, C. Nickel, Cadmium and Lead Phytotoxicity and Potential of Halophytic Plants in Heavy Metal Extraction. S. Afr. J. Bot. 2017, 111, 99–110. [Google Scholar] [CrossRef]
- Alamri, S.A.; Siddiqui, M.H.; Al-Khaishany, M.Y.Y.; Nasir Khan, M.; Ali, H.M.; Alaraidh, I.A.; Alsahli, A.A.; Al-Rabiah, H.; Mateen, M. Ascorbic Acid Improves the Tolerance of Wheat Plants to Lead Toxicity. J. Plant Interact. 2018, 13, 409–419. [Google Scholar] [CrossRef] [Green Version]
- Pratap, A.; Gupta, S.K.; Kumar, J.; Solanki, R.K. Soybean. In Technological Innovations in Major World Oil Crops; Springer New York: New York, NY, USA, 2012; Volume 1, pp. 293–321. [Google Scholar]
- Dixit, A.; Antony, J.I.; Sharma, N.K.; Tiwari, R.K. Soybean Constituents and Their Functional Benefits. In Opportunity, Challenge and Scope of Natural Products in Medicinal Chemistry; Research Signpost: Kerala, India, 2011; pp. 367–383. ISBN 9788130804484. [Google Scholar]
- Medic, J.; Atkinson, C.; Hurburgh, C.R. Current Knowledge in Soybean Composition. J. Am. Oil Chem. Soc. 2014, 91, 363–384. [Google Scholar] [CrossRef]
- Nikolić, Z.; Vasiljević, I.; Zdjelar, G.; Đorđević, V.; Ignjatov, M.; Jovičić, D.; Milošević, D. Detection of Genetically Modified Soybean in Crude Soybean Oil. Food Chem. 2014, 145, 1072–1075. [Google Scholar] [CrossRef] [PubMed]
- Bakhoum, G.S.; Badr, E.A.E.; Sadak, M.S.; Kabesh, M.O.; Amin, G.A. Improving Growth, Some Biochemical Aspects and Yield of Three Cultivars of Soybean Plant by Methionine Treatment Under Sandy Soil Condition. Int. J. Environ. Res. 2019, 13, 35–43. [Google Scholar] [CrossRef]
- Bilal, S.; Khan, A.L.; Shahzad, R.; Kim, Y.-H.; Imran, M.; Khan, M.J.; Al-Harrasi, A.; Kim, T.H.; Lee, I.-J. Mechanisms of Cr(VI) Resistance by Endophytic Sphingomonas Sp. LK11 and Its Cr(VI) Phytotoxic Mitigating Effects in Soybean (Glycine max L.). Ecotoxicol. Environ. Saf. 2018, 164, 648–658. [Google Scholar] [CrossRef] [PubMed]
- da Rosa Couto, R.; Faversani, J.; Ceretta, C.A.; Ferreira, P.A.A.; Marchezan, C.; Basso Facco, D.; Garlet, L.P.; Silva, J.S.; Comin, J.J.; Bizzi, C.A.; et al. Health Risk Assessment and Soil and Plant Heavy Metal and Bromine Contents in Field Plots after Ten Years of Organic and Mineral Fertilization. Ecotoxicol. Environ. Saf. 2018, 153, 142–150. [Google Scholar] [CrossRef] [PubMed]
- Armendariz, A.L.; Talano, M.A.; Olmos Nicotra, M.F.; Escudero, L.; Breser, M.L.; Porporatto, C.; Agostini, E. Impact of Double Inoculation with Bradyrhizobium japonicum E109 and Azospirillum brasilense Az39 on Soybean Plants Grown under Arsenic Stress. Plant Physiol. Biochem. 2019, 138, 26–35. [Google Scholar] [CrossRef] [PubMed]
- Adeyemi, N.O.; Atayese, M.O.; Sakariyawo, O.S.; Azeez, J.O.; Abayomi Sobowale, S.P.; Olubode, A.; Mudathir, R.; Adebayo, R.; Adeoye, S. Alleviation of Heavy Metal Stress by Arbuscular Mycorrhizal Symbiosis in Glycine max (L.) Grown in Copper, Lead and Zinc Contaminated Soils. Rhizosphere 2021, 18, 100325. [Google Scholar] [CrossRef]
- Fu, Y.; Yuan, Q.; Lin, S.; Liu, W.; Du, G.; Zhao, L.; Zhang, Q.; Lin, D.-R.; Liu, Y.-T.; Qin, W.; et al. Physicochemical Characteristics and Biological Activities of Polysaccharides from the Leaves of Different Loquat (Eriobotrya japonica) Cultivars. Int. J. Biol. Macromol. 2019, 135, 274–281. [Google Scholar] [CrossRef] [PubMed]
- Dhiman, A.; Suhag, R.; Thakur, D.; Gupta, V.; Prabhakar, P.K. Current Status of Loquat (Eriobotrya japonica Lindl.): Bioactive Functions, Preservation Approaches, and Processed Products. Food Rev. Int. 2022, 38, 286–316. [Google Scholar] [CrossRef]
- Kim, H.H.; Heo, M.R.; Lee, S.; Yim, S.H. Validation of Analytical Method and Antioxidant Properties of Eriobotrya japonica Lindl. Leaf Extract According to Extraction Solvent. Korean J. Food Sci. Technol. 2019, 51, 301–308. [Google Scholar] [CrossRef]
- Hoagland, D.R.; Arnon, D.I. The Water-Culture Method for Growing Plants without Soil; California Agricultural Experiment Station: Berkeley, CA, USA, 1950; Volume 347. [Google Scholar]
- Rao, B.; Tang, R.-C. Green Synthesis of Silver Nanoparticles with Antibacterial Activities Using Aqueous Eriobotrya japonica Leaf Extract. Adv. Nat. Sci. Nanosci. Nanotechnol. 2017, 8, 015014. [Google Scholar] [CrossRef] [Green Version]
- Malik, R.N.; Husain, S.Z.; Nazir, I. Heavy Metal Contamination and Accumulation in Soil and Wild Plant Species from Industrial Area of Islamabad, Pakistan. Pak. J. Bot. 2010, 42, 291–301. [Google Scholar]
- Arnon, D. Copper Enzyme in Isolated Chloroplast and Chlorophyll Expressed in Terms of Mg per Gram. Plant Physiol. 1949, 24, 21–24. [Google Scholar]
- Ravelo-Pérez, L.M.; Hernández-Borges, J.; Rodríguez-Delgado, M.A.; Borges-Miquel, T. Spectrophotometric Analysis of Lycopene in Tomatoes and Watermelons: A Practical Class. Chem. Educ. 2008, 13, 11–13. [Google Scholar]
- Bates, L.S.; Waldren, R.P.; Teare, I.D. Rapid Determination of Free Proline for Water-Stress Studies. Plant Soil 1973, 39, 205–207. [Google Scholar] [CrossRef]
- Irigoyen, J.J.; Einerich, D.W.; Sánchez-Díaz, M. Water Stress Induced Changes in Concentrations of Proline and Total Soluble Sugars in Nodulated Alfalfa (Medicago sativa) Plants. Physiol. Plant. 1992, 84, 55–60. [Google Scholar] [CrossRef]
- Zhang, J.; Kirkham, M.B. Drought-Stress-Induced Changes in Activities of Superoxide Dismutase, Catalase, and Peroxidase in Wheat Species. Plant Cell Physiol. 1994, 35, 785–791. [Google Scholar] [CrossRef]
- Jana, S.; Choudhuri, M.A. Glycolate Metabolism of Three Submersed Aquatic Angiosperms: Effect of Heavy Metals. Aquat. Bot. 1981, 11, 67–77. [Google Scholar] [CrossRef]
- Kampfenkel, K.; Vanmontagu, M.; Inze, D. Extraction and Determination of Ascorbate and Dehydroascorbate from Plant Tissue. Anal. Biochem. 1995, 225, 165–167. [Google Scholar] [CrossRef]
- Anderson, M.E. Determination of Glutathione and Glutathione Disulfide in Biological Samples. In Methods in Enzymology; Academic Press: Cambridge, MA, USA, 1985; Volume 113, pp. 548–555. [Google Scholar]
- Velioglu, Y.S.; Mazza, G.; Gao, L.; Oomah, B.D. Antioxidant Activity and Total Phenolics in Selected Fruits, Vegetables, and Grain Products. J. Agric. Food Chem. 1998, 46, 4113–4117. [Google Scholar] [CrossRef]
- Zhishen, J.; Mengcheng, T.; Jianming, W. The Determination of Flavonoid Contents in Mulberry and Their Scavenging Effects on Superoxide Radicals. Food Chem. 1999, 64, 555–559. [Google Scholar] [CrossRef]
- Zhang, X.Z. The Measurement and Mechanism of Lipid Peroxidation and SOD, POD and CAT Activities in Biological System. In Research Methodology in Crop Physiology; Agriculture Press: Beijing, China, 1992; pp. 208–211. [Google Scholar]
- Aebi, H. Catalase in Vitro. In Methods in Enzymology; Academic Press: Cambridge, MA, USA, 1984; pp. 121–126. [Google Scholar]
- Zhou, W.; Leul, M. Uniconazole-Induced Tolerance of Rape Plants to Heat Stress in Relation to Changes in Hormonal Levels, Enzyme Activities and Lipid Peroxidation. Plant Growth Regul. 1999, 27, 99–104. [Google Scholar] [CrossRef]
- Nakano, Y.; Asada, K. Hydrogen Peroxide Is Scavenged by Ascorbate-Specific Peroxidase in Spinach Chloroplasts. Plant Cell Physiol. 1981, 22, 867–880. [Google Scholar] [CrossRef]
- Rao, M.V.; Paliyath, G.; Ormrod, D.P. Ultraviolet-B- and Ozone-Induced Biochemical Changes in Antioxidant Enzymes of Arabidopsis thaliana. Plant Physiol. 1996, 110, 125–136. [Google Scholar] [CrossRef] [Green Version]
- Shapiro, S.S.; Wilk, M.B. An Analysis of Variance Test for Normality (Complete Samples). Biometrika 1965, 52, 591–611. [Google Scholar] [CrossRef]
- Steel, R.G.D.; Torrie, J.H.; Dickey, D. Principles and Procedure of Statistics: A Biometrical Approach, 3rd ed.; McGraw HillBookCo. Inc.: New York, NY, USA, 1997; pp. 352–358. [Google Scholar]
- IBM SPSS Statistics for Windows; IBM Corp: Armonk, NY, USA, 2015.
- Sharma, P.; Dubey, R.S. Lead Toxicity in Plants. Braz. J. Plant Physiol. 2005, 17, 35–52. [Google Scholar] [CrossRef] [Green Version]
- Parrotta, L. Target or Barrier? The Cell Wall of Early- and Later-Diverging Plants vs Cadmium Toxicity: Differences in the Response Mechanisms. Front. Plant Sci. 2015, 6, 133. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mishra, S.; Srivastava, S.; Tripathi, R.D.; Kumar, R.; Seth, C.S.; Gupta, D.K. Lead Detoxification by Coontail (Ceratophyllum demersum L.) Involves Induction of Phytochelatins and Antioxidant System in Response to Its Accumulation. Chemosphere 2006, 65, 1027–1039. [Google Scholar] [CrossRef] [PubMed]
- Bücker-Neto, L.; Paiva, A.L.S.; Machado, R.D.; Arenhart, R.A.; Margis-Pinheiro, M. Interactions between Plant Hormones and Heavy Metals Responses. Genet. Mol. Biol. 2017, 40, 373–386. [Google Scholar] [CrossRef]
- Vega, A.; Delgado, N.; Handford, M. Increasing Heavy Metal Tolerance by the Exogenous Application of Organic Acids. Int. J. Mol. Sci. 2022, 23, 5438. [Google Scholar] [CrossRef]
- Kim, S.-M.; Kim, T.-K.; Kang, M.-C.; Cha, J.Y.; Yong, H.I.; Choi, Y.-S. Effects of Loquat (Eriobotrya japonica Lindl.) Leaf Extract with or without Ascorbic Acid on the Quality Characteristics of Semi-Dried Restructured Jerky during Storage. Food Sci. Anim. Resour. 2022, 42, 566–579. [Google Scholar] [CrossRef] [PubMed]
- Dogan, M.; Karatas, M.; Aasim, M. Cadmium and Lead Bioaccumulation Potentials of an Aquatic Macrophyte Ceratophyllum demersum L.: A Laboratory Study. Ecotoxicol. Environ. Saf. 2018, 148, 431–440. [Google Scholar] [CrossRef] [PubMed]
- Shu, X.; Yin, L.; Zhang, Q.; Wang, W. Effect of Pb Toxicity on Leaf Growth, Antioxidant Enzyme Activities, and Photosynthesis in Cuttings and Seedlings of Jatropha curcas L. Environ. Sci. Pollut. Res. 2012, 19, 893–902. [Google Scholar] [CrossRef]
- Rucińska-Sobkowiak, R. Water Relations in Plants Subjected to Heavy Metal Stresses. Acta Physiol. Plant. 2016, 38, 257. [Google Scholar] [CrossRef] [Green Version]
- Chandra, R.; Kang, H. Mixed Heavy Metal Stress on Photosynthesis, Transpiration Rate, and Chlorophyll Content in Poplar Hybrids. For. Sci. Technol. 2016, 12, 55–61. [Google Scholar] [CrossRef] [Green Version]
- Yadav, S.K. Heavy Metals Toxicity in Plants: An Overview on the Role of Glutathione and Phytochelatins in Heavy Metal Stress Tolerance of Plants. S. Afr. J. Bot. 2010, 76, 167–179. [Google Scholar] [CrossRef] [Green Version]
- Vidal, C.; Ruiz, A.; Ortiz, J.; Larama, G.; Perez, R.; Santander, C.; Ferreira, P.A.A.; Cornejo, P. Antioxidant Responses of Phenolic Compounds and Immobilization of Copper in Imperata Cylindrica, a Plant with Potential Use for Bioremediation of Cu Contaminated Environments. Plants 2020, 9, 1397. [Google Scholar] [CrossRef]
- Ren, S.-C.; Sun, J.-T. Changes in Phenolic Content, Phenylalanine Ammonia-Lyase (PAL) Activity, and Antioxidant Capacity of Two Buckwheat Sprouts in Relation to Germination. J. Funct. Foods 2014, 7, 298–304. [Google Scholar] [CrossRef]
- Adhikari, A.; Adhikari, S.; Ghosh, S.; Azahar, I.; Shaw, A.K.; Roy, D.; Roy, S.; Saha, S.; Hossain, Z. Imbalance of Redox Homeostasis and Antioxidant Defense Status in Maize under Chromium (VI) Stress. Environ. Exp. Bot. 2020, 169, 103873. [Google Scholar] [CrossRef]
- Ashraf, U.; Kanu, A.S.; Mo, Z.; Hussain, S.; Anjum, S.A.; Khan, I.; Abbas, R.N.; Tang, X. Lead Toxicity in Rice: Effects, Mechanisms, and Mitigation Strategies—A Mini Review. Environ. Sci. Pollut. Res. 2015, 22, 18318–18332. [Google Scholar] [CrossRef] [PubMed]
- Sofy, M.R.; Seleiman, M.F.; Alhammad, B.A.; Alharbi, B.M.; Mohamed, H.I. Minimizing Adverse Effects of Pb on Maize Plants by Combined Treatment with Jasmonic, Salicylic Acids and Proline. Agronomy 2020, 10, 699. [Google Scholar] [CrossRef]
- Dezhban, A.; Shirvany, A.; Attarod, P.; Delshad, M.; Matinizadeh, M.; Khoshnevis, M. Cadmium and Lead Effects on Chlorophyll Fluorescence, Chlorophyll Pigments and Proline of Robinia pseudoacacia. J. For. Res. 2015, 26, 323–329. [Google Scholar] [CrossRef]
- Ben Rejeb, K.; Abdelly, C.; Savouré, A. How Reactive Oxygen Species and Proline Face Stress Together. Plant Physiol. Biochem. 2014, 80, 278–284. [Google Scholar] [CrossRef] [PubMed]
- Xu, H.; Chen, J. Commercial Quality, Major Bioactive Compound Content and Antioxidant Capacity of 12 Cultivars of Loquat (Eriobotrya japonica Lindl.) Fruits. J. Sci. Food Agric. 2011, 91, 1057–1063. [Google Scholar] [CrossRef]
- Kısa, D.; Elmastaş, M.; Öztürk, L.; Kayır, Ö. Responses of the Phenolic Compounds of Zea mays under Heavy Metal Stress. Appl. Biol. Chem. 2016, 59, 813–820. [Google Scholar] [CrossRef]
- Márquez-García, B.; Fernández-Recamales, M.Á.; Córdoba, F. Effects of Cadmium on Phenolic Composition and Antioxidant Activities of Erica Andevalensis. J. Bot. 2012, 2012, 936950. [Google Scholar] [CrossRef] [Green Version]
- Giannakoula, A.; Therios, I.; Chatzissavvidis, C. Effect of Lead and Copper on Photosynthetic Apparatus in Citrus (Citrus aurantium L.) Plants. The Role of Antioxidants in Oxidative Damage as a Response to Heavy Metal Stress. Plants 2021, 10, 155. [Google Scholar] [CrossRef] [PubMed]
- Thakur, S.; Singh, L.; Zularisam, A.W.; Sakinah, M.; Din, M.F.M. Lead Induced Oxidative Stress and Alteration in the Activities of Antioxidative Enzymes in Rice Shoots. Biol. Plant. 2017, 61, 595–598. [Google Scholar] [CrossRef]
- Turrens, J.F. Mitochondrial Formation of Reactive Oxygen Species. J. Physiol. 2003, 552, 335–344. [Google Scholar] [CrossRef]
- Rajput, V.D.; Harish; Singh, R.K.; Verma, K.K.; Sharma, L.; Quiroz-Figueroa, F.R.; Meena, M.; Gour, V.S.; Minkina, T.; Sushkova, S.; et al. Recent Developments in Enzymatic Antioxidant Defence Mechanism in Plants with Special Reference to Abiotic Stress. Biology 2021, 10, 267. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Source | DF | Sum of Squares | Mean Squares | F Value | p Value |
|---|---|---|---|---|---|
| Shoot dry biomass | |||||
| Pb | 3 | 1.992 | 0.664 | 451.407 | <0.0001 |
| EJLE | 1 | 0.112 | 0.112 | 76.193 | <0.0001 |
| Pb × EJLE | 3 | 0.010 | 0.003 | 2.289 | 0.007 |
| Root dry biomass | |||||
| Pb | 3 | 0.155 | 0.052 | 343.444 | <0.0001 |
| EJLE | 1 | 0.031 | 0.031 | 205.444 | <0.0001 |
| Pb × EJLE | 3 | 0.001 | 0.000 | 2.259 | 0.001 |
| Chlorophyll contents | |||||
| Pb | 3 | 133.933 | 44.644 | 327.304 | <0.0001 |
| EJLE | 1 | 21.755 | 21.755 | 159.495 | <0.0001 |
| Pb × EJLE | 3 | 1.817 | 0.606 | 4.441 | 0.019 |
| Carotenoid contents | |||||
| Pb | 3 | 90.760 | 30.253 | 277.957 | <0.0001 |
| EJLE | 1 | 19.260 | 19.260 | 176.958 | <0.0001 |
| Pb × EJLE | 3 | 4.657 | 1.552 | 14.262 | <0.0001 |
| Anthocyanin contents | |||||
| Pb | 3 | 66.499 | 22.166 | 236.471 | <0.0001 |
| EJLE | 1 | 14.369 | 14.369 | 153.285 | <0.0001 |
| Pb × EJLE | 3 | 3.275 | 1.092 | 11.644 | 0.000 |
| Lycopene contents | |||||
| Pb | 3 | 134.656 | 44.885 | 427.819 | <0.0001 |
| EJLE | 1 | 12.327 | 12.327 | 117.490 | <0.0001 |
| Pb × EJLE | 3 | 8.453 | 2.818 | 26.857 | <0.0001 |
| Pb in roots | |||||
| Pb | 3 | 605,785.83 | 201,928.611 | 6319.708 | <0.0001 |
| EJLE | 1 | 34,898.101 | 34,898.101 | 1092.197 | <0.0001 |
| Pb × EJLE | 3 | 12,993.148 | 4331.049 | 135.548 | <0.0001 |
| Pb in leaves | |||||
| Pb | 3 | 187,399.403 | 62,466.468 | 1565.556 | <0.0001 |
| EJLE | 1 | 24,400.315 | 24,400.315 | 611.529 | <0.0001 |
| Pb × EJLE | 3 | 14,045.921 | 4681.974 | 117.341 | <0.0001 |
| Pb translocation factor | |||||
| Pb | 3 | 1.101 | 0.367 | 1577.826 | <0.0001 |
| EJLE | 1 | 0.038 | 0.038 | 161.552 | <0.0001 |
| Pb × EJLE | 3 | 0.018 | 0.006 | 26.438 | <0.0001 |
| Source | DF | Sum of Squares | Mean Squares | F Value | p Value |
|---|---|---|---|---|---|
| Proline contents | |||||
| Pb | 3 | 117.536 | 39.179 | 701.078 | <0.0001 |
| EJLE | 1 | 36.064 | 36.064 | 645.345 | <0.0001 |
| Pb × EJLE | 3 | 2.781 | 0.927 | 16.586 | <0.0001 |
| Total soluble sugars | |||||
| Pb | 3 | 224.825 | 74.942 | 658.298 | <0.0001 |
| EJLE | 1 | 25.855 | 25.855 | 227.109 | <0.0001 |
| Pb × EJLE | 3 | 3.613 | 1.204 | 10.578 | 0.000 |
| Malondialdehyde | |||||
| Pb | 3 | 419.985 | 139.995 | 1524.793 | <0.0001 |
| EJLE | 1 | 67.670 | 67.670 | 737.050 | <0.0001 |
| Pb × EJLE | 3 | 5.256 | 1.752 | 19.081 | <0.0001 |
| Hydrogen peroxide | |||||
| Pb | 3 | 428.681 | 142.894 | 2330.737 | <0.0001 |
| EJLE | 1 | 74.026 | 74.026 | 1207.437 | <0.0001 |
| Pb × EJLE | 3 | 7.769 | 2.590 | 42.240 | <0.0001 |
| Ascorbic acid | |||||
| Pb | 3 | 9924.944 | 3308.315 | 216.585 | <0.0001 |
| EJLE | 1 | 14,653.030 | 14,653.030 | 959.289 | <0.0001 |
| Pb × EJLE | 3 | 22,017.881 | 7339.294 | 480.481 | <0.0001 |
| Glutathione | |||||
| Pb | 3 | 26,018.168 | 8672.723 | 1044.265 | <0.0001 |
| EJLE | 1 | 2623.787 | 2623.787 | 315.925 | <0.0001 |
| Pb × EJLE | 3 | 244.641 | 81.547 | 9.819 | 0.001 |
| Total phenolic contents | |||||
| Pb | 3 | 154.907 | 51.636 | 608.553 | <0.0001 |
| EJLE | 1 | 97.486 | 97.486 | 1148.918 | <0.0001 |
| Pb × EJLE | 3 | 17.907 | 5.969 | 70.347 | <0.0001 |
| Total flavonoid contents | |||||
| Pb | 3 | 3.713 | 1.238 | 120.001 | <0.0001 |
| EJLE | 1 | 2.975 | 2.975 | 288.495 | <0.0001 |
| Pb × EJLE | 3 | 0.743 | 0.248 | 24.011 | <0.0001 |
| Source | DF | Sum of Squares | Mean Squares | F Value | p Value |
|---|---|---|---|---|---|
| SOD | |||||
| Pb | 3 | 1505.74 | 501.914 | 68.177 | <0.0001 |
| EJLE | 1 | 682.02 | 682.027 | 92.642 | <0.0001 |
| Pb × EJLE | 3 | 21.10 | 7.034 | 0.955 | 0.0043 |
| CAT | |||||
| Pb | 3 | 1335.03 | 445.011 | 66.623 | <0.0001 |
| EJLE | 1 | 1549.95 | 1549.952 | 232.046 | <0.0001 |
| Pb × EJLE | 3 | 5.37 | 1.790 | 0.268 | 0.008 |
| POD | |||||
| Pb | 3 | 887.04 | 295.681 | 81.848 | <0.0001 |
| EJLE | 1 | 1121.07 | 1121.077 | 310.327 | <0.0001 |
| Pb × EJLE | 3 | 11.78 | 3.929 | 1.088 | 0.003 |
| APX | |||||
| Pb | 3 | 909.81 | 303.271 | 43.815 | <0.0001 |
| EJLE | 1 | 573.30 | 573.304 | 82.829 | <0.0001 |
| Pb × EJLE | 3 | 2.55 | 0.850 | 0.123 | 0.005 |
| GR | |||||
| Pb | 3 | 921.65 | 307.219 | 26.546 | <0.0001 |
| EJLE | 1 | 513.37 | 513.375 | 44.360 | <0.0001 |
| Pb × EJLE | 3 | 6.88 | 2.295 | 0.198 | 0.008 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Khalofah, A.; Farooq, S. Physiological, Morphological, and Biochemical Responses of Soybean [Glycine max (L.) Merr.] to Loquat (Eriobotrya japonica Lindl.) Leaf Extract Application on Pb-Contaminated Soil. Sustainability 2023, 15, 4352. https://0-doi-org.brum.beds.ac.uk/10.3390/su15054352
Khalofah A, Farooq S. Physiological, Morphological, and Biochemical Responses of Soybean [Glycine max (L.) Merr.] to Loquat (Eriobotrya japonica Lindl.) Leaf Extract Application on Pb-Contaminated Soil. Sustainability. 2023; 15(5):4352. https://0-doi-org.brum.beds.ac.uk/10.3390/su15054352
Chicago/Turabian StyleKhalofah, Ahlam, and Shahid Farooq. 2023. "Physiological, Morphological, and Biochemical Responses of Soybean [Glycine max (L.) Merr.] to Loquat (Eriobotrya japonica Lindl.) Leaf Extract Application on Pb-Contaminated Soil" Sustainability 15, no. 5: 4352. https://0-doi-org.brum.beds.ac.uk/10.3390/su15054352