
Terrestrial and Aquatic Plastisphere: Formation, Characteristics, and Influencing Factors


Department of Environmental Science, The University of Arizona, Tucson, AZ 85721, USA
Sustainability 2024, 16(5), 2163; https://0-doi-org.brum.beds.ac.uk/10.3390/su16052163
Submission received: 14 February 2024
/
Revised: 28 February 2024
/
Accepted: 2 March 2024
/
Published: 5 March 2024
(This article belongs to the Special Issue Fates, Transports, Interactions and Monitoring of Emerging Pollutants)
Abstract
:The increasing number of plastic particles in the environment gives rise to a unique ecological niche called the plastisphere, which is characterized by a dynamic assemblage of microorganisms comprising autotrophs, heterotrophs, predators, and pathogens. This paper reviews the formation, characteristics, and factors influencing the terrestrial and aquatic plastisphere. The terrestrial and aquatic plastisphere forms when hitchhiking microorganisms, often bacteria, in the environment adhere to the surfaces of plastic particles and alter the surface properties of the particles for subsequent colonization of increasingly tightly clinging microorganisms. The terrestrial plastisphere is not as mobile as the aquatic plastisphere and is defined by the characteristics of the soil anchoring it. The microorganisms on the plastisphere are often defined by the dominant microorganisms in the surroundings, particularly those of the phyla Proteobacteria, Actinobacteria, Bacteroidota, Firmicutes, Chloroflexi, Acidobacteria, Cyanobacteria, Ascomycota, Basidiomycota, Chytridiomycota, Ciliophora, Ochrophyta, and Chlorophyta. However, the compositions and dominant microbial species on different plastic surfaces vary widely as they change with time, plastic properties, and biotic and abiotic environmental factors. Temporal changes in the plastisphere are due to succession. Plastic properties, particularly the type, surface properties, size, color, degree of aging, and chemical leaching, also affect the composition of the plastisphere. Biotic environmental factors comprising the dominant ambient microorganisms and interspecies interactions, together with the abiotic ones, including pH, temperature, nutrient availability, soil properties, salinity, and light, significantly shape the plastisphere. This review provides insight into the biodiversity of the plastisphere, and its roles in spreading pathogens and degrading plastics.
1. Introduction
Global plastic pollution has led to a tremendous increase in research related to microplastics and nanoplastics. According to the Global Plastic Outlook of the Organization for Economic Co-operation and Development, approximately 82 million tons of plastic waste produced worldwide is mismanaged, of which 13 million tons enter the terrestrial environments, and 6 million tons enter the rivers or coastlines [1]. Subsequently, 1.7 million tons of mismanaged plastic entering the environment is transported to the ocean, indicating that oceans receive about 0.5% of the global plastic waste [1]. This plastic waste is a direct source of microplastics. Microplastics can be released in their primary form from plastic particles smaller than 5 mm such as microbeads, plastic pellets or nurdles, and plastic fibers. Microplastics can also come from the degradation of larger plastic items in the environment, and these are called secondary microplastics [2].
In the environment, microplastics interact with the media they are in, as well as the organisms and chemicals in the media [3]. With microplastics garnering global attention and a surge in microplastics-related research, the term ‘plastisphere’ has increasingly appeared in the scientific literature. The plastisphere refers to the ecosystem that exists on the surface of plastic debris, especially in marine environments. This ecosystem consists of various microorganisms, including bacteria, algae, and other single-celled organisms that have evolved to live in human-made plastic environments [4]. Though research on the plastisphere was initially focused on marine environments, there have been increasing studies on the plastisphere in terrestrial environments lately [5,6,7,8]. The plastisphere is unique because it differs from other floating materials that naturally occur due to plastic’s unique chemical properties and slow biodegradation rate. Over 1000 different species of microbes are able to inhabit just one microplastic of 5 mm. The plastisphere can also act as a transport medium for these organisms over long distances, potentially introducing invasive species to different ecosystems [9].
The term ‘plastisphere’ was first coined by Zettler et al. in 2013 [8]. Since its introduction, the term has expanded beyond microbiology to include interactions of any organisms with plastics [10]. As studies of the plastisphere in soil grow, the definition of the plastisphere has moved beyond the microbial communities inhabiting plastic particles in the aquatic environment. While some scholars still adopt the original definition of plastisphere intended for the aquatic environment, others promulgate the concept of habitat or niche on plastic surfaces, leading to the emergence of terms such as the ‘plastisphere microbiome’ or the ‘plastisphere microbial community’ [11]. These terms suggest that the plastisphere is not limited to microorganisms on plastic surfaces but also includes those in soil affected by the presence of plastics. The leaching of chemicals from plastics changes their immediate surroundings and these changes are more prevalent in the solid matrix of soil than in water. Therefore, the plastisphere is not analogous to biofilm, particularly in the terrestrial context [4]. While research related to the plastisphere mounts, there are currently few reviews related to the plastisphere. Wang et al. reviewed the methods for examining the soil plastisphere and the microbial interactions in the plastisphere to enable a better understanding of its ecological risks [12]. Du et al. reviewed the microorganisms comprising bacteria, fungi, and autotrophs in the marine plastisphere, as well as how ocean depth and seawater nutrients affect the plastisphere’s microbial structures [10]. Another review illustrates the techniques employed to image plastispheres [13]. Additionally, the microbial communities in the aquatic plastisphere and their interactions with the plastics have been reviewed [14]. However, currently no review has delved into the formation and characteristics of the aquatic and terrestrial plastisphere, or discussed the factors shaping the plastisphere. Furthermore, there is a lack of consensus on the specific characteristics of the plastisphere, and the variability of the plastisphere is not well understood. This review, therefore, aims to present the formation and characteristics of the plastisphere in the aquatic and terrestrial environments and synthesize the plastisphere-influencing factors.
The plastisphere represents a unique ecological habitat. This review is foreseen to contribute to a better understanding of how microorganisms colonize and adapt to this new anthropogenic habitat and the many factors shaping the plastisphere. It compares the findings of studies on the characteristics of the plastisphere to identify their similarities and differences. It may shed light on developing strategies to mitigate the impacts of plastic pollution. To achieve this aim, this review includes peer-reviewed articles related to the plastisphere that have been published in the past 10 years. Papers published in the 5 years were prioritized in the review. The literature search was conducted using online journal databases comprising Web of Science, Scopus, and ProQuest, with keywords including aquatic, terrestrial, plastisphere, formation, features, implications, factors, influence, and characteristics. Combinations of the keywords such as influencing factors of terrestrial plastisphere, characteristics of aquatic plastisphere, and formation of aquatic and terrestrial plastisphere were entered to refine the search. The inclusion criteria are (1) the articles are related to aquatic or terrestrial plastisphere; (2) the articles describe the formation, characteristics, or influencing factors of plastisphere; (3) the articles were written in English; and (4) the articles were published in the past 10 years.
2. Formation of the Plastisphere
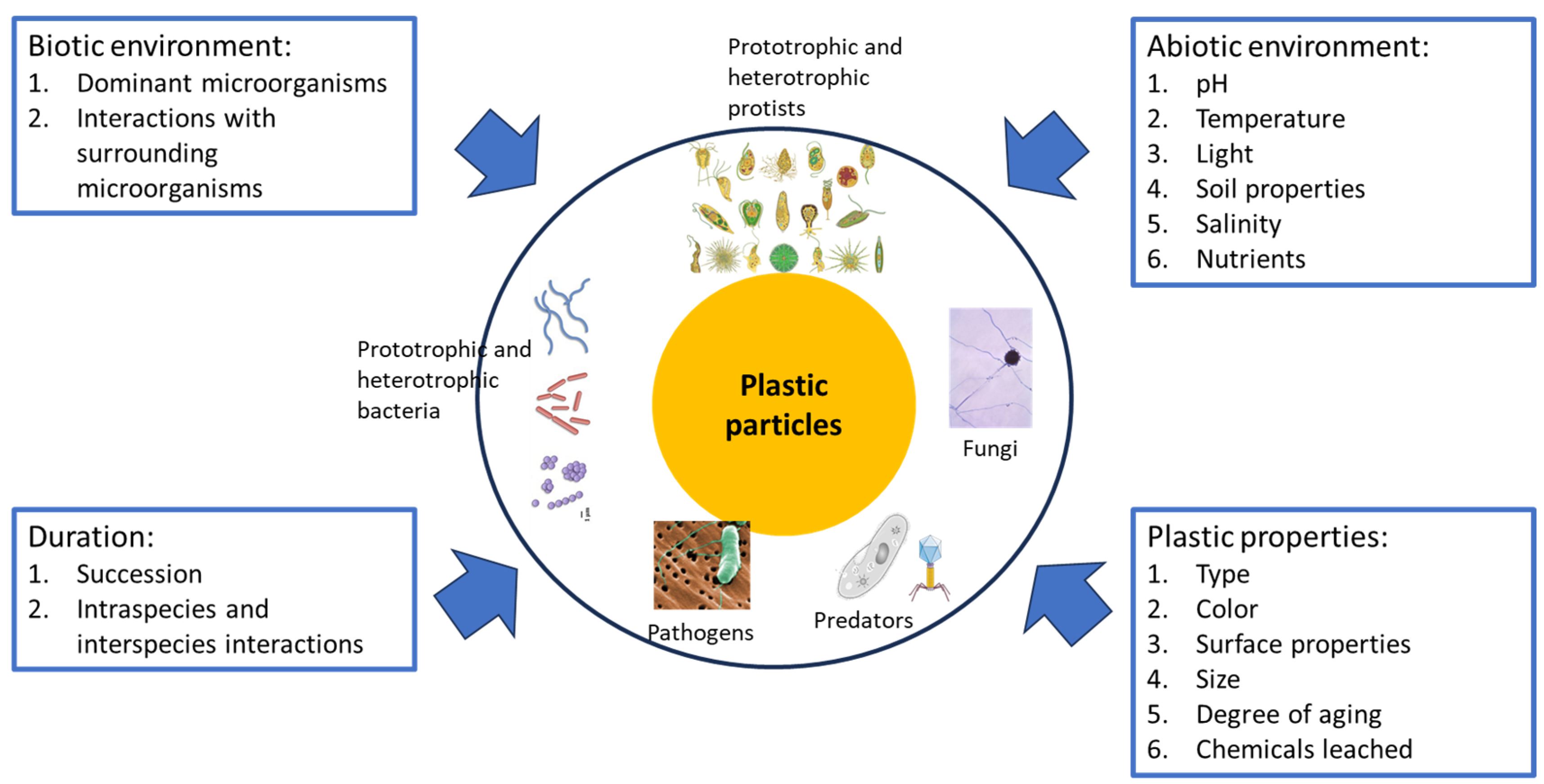
The plastisphere in aquatic environments forms when plastic waste is transformed into microplastics through physical and chemical disruption processes within aquatic environments [15]. Physical forces such as waves can grind plastics against rocks, sand, and other floatables in the ocean, causing the plastics to break into smaller pieces [16]. Plastics can also degrade chemically in the ocean through photo-oxidation, which involves the absorption of light by the plastics, resulting in the formation of free radicals [17]. The free radicals can react with oxygen in the air to form hydroxy, carbonyl, and carboxyl groups. The presence of these groups increases the susceptibility of plastics to further degradation through cracking, and fragmentation, which subsequently causes the formation of microplastics [17]. Microplastics colonized by biofilm-forming microbial communities are known as the plastisphere. This unique substrate can aid microbial dispersal, and it has synergistic effects on the development, transportation, persistence, and ecology of microorganisms [18]. The microbial community assemblages in the plastisphere include autotrophs, heterotrophs, predators, and pathogens (Figure 1). The plastic materials, either biodegradable or non-biodegradable, are used by opportunistic aquatic microorganisms as adhesion surfaces rather than carbon sources [14]. Additionally, the plastisphere in freshwater and seawater could be nuanced in formation and characteristics, due to the difference in plastic and microplastic movements. In freshwater, microplastics are likely to follow a spiraling movement, affecting the composition and activity of microorganisms in the plastisphere [7,19].
The plastisphere in the soil forms when microplastic debris accumulates, creating a unique microenvironment that provides new ecological niches for microorganisms [4]. Microplastic characteristics and soil environmental factors can drive the formation and succession of soil plastisphere communities. Notably, the dynamics and behaviors of the soil plastisphere diverge from its marine counterpart, where it was initially defined, thereby likely exhibiting uncharacterized features in terms of ecological effects and biogeochemistry [20]. The soil plastisphere does not exhibit the mobility of the oceanic plastisphere, which moves with currents. It is significantly influenced by the characteristics of the soil anchoring it [4]. As soils are commonly heterogeneous, plastic particles can be incorporated into the soil aggregates and distributed to different microenvironments within the same soil mass. The redox reactions are different in soil environments, leading to distinct microbial communities [21]. Studies have pointed to the differences in the microbial communities between the soil plastisphere and its surroundings, with Proteobacteria being more prevalent in the former [11,22]. This aligns with the predominance of Proteobacteria in the marine plastisphere [8]. The reason for this is that numerous species of Proteobacteria can produce extracellular polymeric substances and have a clinging structure that enables them to stick to the surfaces of plastic particles. Furthermore, the soil plastisphere is affected by two major factors, namely the plastic material and the microbial communities on and around plastics. Plastics provide the surface for anchoring microorganisms. Concurrently, their presence changes the physicochemical soil environment by modifying soil structure and leaching chemicals [12]. In soil, plastics are less mobile and have a significantly longer interaction time with their surroundings with more pronounced spatial differences [23]. For instance, plastic particles in hotspots such as the rhizosphere are likely to anchor microorganisms typical of the rhizosphere, instead of those in the bulk soil. The plastisphere can be perceived to be incorporated into the soil compartments in soil environment [23].
Biogeochemistry modulates microbial community composition and mineral formation in water, soil, and sediment over spatial and temporal scales. In the coastal environment, comprising the water column and sediment, the microbes and minerals associated with microplastics vary in different parts of the environment due to the redox conditions and the biogeochemistry [24]. A study revealed the decreasing coverage of the plastisphere and minerals on polyethylene (PE) and polystyrene (PS) films as the depth of the sediment increased, with those in the bottom sediment having the lowest density of microorganisms [24]. Diatoms were observed in the plastisphere together with bacteria of diverse morphologies, as well as crystalline and amorphous metal oxide minerals. PS films adsorbed more metals than PE films. The metal concentrations decreased in the bottom sediment. Exposed plastics in water and surface sediment usually undergo more oxidation than those buried in deeper sediment [24]. This indicates that the plastisphere formed in different compartments of an environment is likely to have varying characteristics.
There is an increasing interest in the eco-corona formed around microplastics and nanoplastics, which potentially affects the plastisphere. An eco-corona refers to a layer of ecological macromolecules, such as natural organic matter, extracellular polymeric substances, proteins, and metabolites, bound to the plastic particles through adsorption [25]. It usually forms in natural aquatic environments but can also form in the soil. When plastic particles enter the soil, they can interact with soil metabolomes produced by plant root exudation, microbial and animal metabolism, and the decomposition of soil organic matter. The adsorption of these metabolomes directly onto the plastic particles leads to the formation of an eco-corona [26]. Alternatively, the macromolecules attached to the particles can facilitate bridging interactions with other materials in the surrounding environment, forming an eco-corona. The main components of an eco-corona are lipids or lipid-like molecules, phenylpropanoids and polyketides, nucleosides, nucleotides, and their analogs. The eco-corona alters the physicochemical properties of plastic particles and influences the types of microorganisms that can colonize the plastisphere [27]. The composition of the eco-corona can also affect microbial attachment in a substrate-dependent manner. Moreover, the eco-corona may cause the aggregation of nanoplastics, which facilitates microbial colonization. It plays a crucial role in shaping the plastisphere [28]. Besemer (2016) suggested the presence of core bacteria in the freshwater plastisphere, which is composed of those from the Burkholderiaceae, Illumatobacteraceae, Sphingomonadaceae, and Rhodobacteraceae families [29]. This indicates that the formation of the plastisphere is more closely linked to certain bacteria, particularly those which are more common and abundant in the environment.
3. Migration and Distribution of the Plastisphere
The migration of the plastisphere frequently follows that of the plastic particles. Microplastics enter terrestrial environments through sources such as tire wear, sewage sludge, compost, and biosolids [30]. They tend to migrate from the soil surface to deeper parts of the soil together with rain infiltration through preferential pathways in soil matrices, agronomic practices such as tilling and irrigation, plant root elongations, and soil-burrowing animals [2]. Terrestrial microplastics can be transported by rainfall and air to nearby waterways or soils. Some microplastics are sufficiently light to be transported over long distances through the air before settling due to gravity or being washed down by rain. These microplastics may end up on land or in waterbodies [31,32]. Microplastics in rivers may sink to the bottom and become trapped in the sediments depending on their densities and the flow of the rivers [33]. Those in the water column eventually enter the oceans, where they may remain floating or sink and be incorporated into the sediments [24]. Transfer of microplastics between the terrestrial and aquatic environments is possible, for instance, through flooding, which causes the migration of microplastics from riverbanks into the rivers, as well as tides and storm surges, which move microplastics from the coast into the seawater. These migration modes suggest that the plastisphere is not static and could alter when moving from one environment to another [34].
However, unlike uncolonized microplastics, the formation of the plastisphere alters the physical properties of microplastics, thus changing their movements. The plastisphere may enhance the vertical movement of microplastics through water columns by altering the buoyancy of microplastics and promoting the aggregation of microplastics to form larger particles. This could affect their locations in the water column and, in some instances, their tendency to settle [35]. However, the effects of the plastisphere on microplastics remain poorly understood and are often overpowered by hydrometeorological influences comprising water flow, currents, wind, and rain. Additionally, the sizes and types of microplastics play a significant role in their transport. Polyethylene terephthalate (PET) particles were found to sink in freshwater at a velocity influenced solely by their density, while the plastisphere has a greater influence on the sinking of PS along the river. The presence of buoyant microplastics, particularly PE and polypropylene (PP), below the ocean surface and in sediments suggests that their densities might have been altered through biofouling, causing them to sink [36]. However, measurements of the density of bio-fouled plastics have not been extensively conducted. Studies have indicated that the densities, sizes, and shapes of microplastics determine their sinking rates, but it is their densities that determine if they float, suspend, or sink in the water column [37]. Kooi et al. proposed an oscillatory movement of biofouled microplastics in the ocean demonstrated through repeated cycles of settling and moving upward. This movement results from the density dynamics between the plastic particles and the seawater. The densities of the plastic particles are, in turn, determined by changes in the abundance of microorganisms in the plastisphere due to factors such as light intensity, salinity and nutrient availability. The authors predicted that, if size is the main determinant of microplastic vertical movement in seawater, most microplastics would remain at intermediate depths [38].
In view of the complexity of predicting the distribution of microplastics due to the numerous factors at play, it is deemed that the plastisphere generally permeates both terrestrial and aquatic environments. The plastisphere associated with dense microplastics such as PET, high-density polyethylene (HDPE), and polyvinyl chloride (PVC) is more likely to be found in the sediments, while that associated with more buoyant microplastics is distributed in different parts of the water column. In the terrestrial environment, the plastisphere is less mobile and its distribution is related to how microplastics migrate in soils, for instance, through infiltration, root movements, agronomic practices, and burrowing animals, causing the plastisphere to permeate the deeper soil layers.
4. Characteristics of the Plastisphere
4.1. Terrestrial Environment
The terrestrial plastisphere hosts a multitude of microbial communities. Over 1000 different species of microbes can inhabit a 5 mm piece of plastic. The microbial species within the plastisphere differ from other naturally occurring floating materials due to the unique chemical nature and slow biodegradation of plastics. Studies on the terrestrial plastisphere are frequently conducted on biodegradable and/or hard-to-degrade microplastics added to soil [39,40] or aged microplastics already present in soil due to various human activities [22,41,42]. Most of the studies reviewed compared the richness and abundance of microbial communities in the plastisphere against those in the surrounding soil using alpha diversities, which are a measure of the diversity within a particular area or ecosystem. In terms of microbial ecology, analyzing the alpha diversities of amplicon sequencing data is crucial to assessing differences between microbial environments [43]. Some of the common alpha diversity indices are the Chao index, Simpson index, Shannon index, and Good’s coverage index (Table 1). The Chao index estimates species richness (number of different species) based on abundance data. The Simpson index measures the degree of concentration when individuals are classified into types [44]. The Shannon index assesses species diversity based on both richness and evenness (relative abundance of each species), while the Good’s coverage index estimates how well a sample represents the entire community. They are complementary and are used to indicate the alpha diversity, which quantifies the richness and evenness of a community. A more diverse and balanced microbial community in the plastisphere will have a higher alpha diversity [44].
While most studies have revealed a decline in the alpha diversities of the microbial communities in the plastisphere [5,39,40], some studies found the alpha diversities to be higher, for instance, on plastic-mulched croplands [45] and a maize field with dried maize straw and living clover applied [46]. Li et al. (2023) made a distinction between rare and abundant microorganisms, with the former referring to those less commonly found which may have adapted to thrive on plastic surfaces, and the latter being the commonly found generalists colonizing a variety of surfaces including the plastic surfaces [41]. Plastic-specific microorganisms are tightly attached to the polymeric surface and might be represented by rare but active species [47]. In their study, the alpha diversities of rare microorganisms were higher in the plastisphere. Interestingly, the study also revealed a higher Simpson index for abundant bacteria, indicating that a few dominant species of abundant bacteria are significantly more abundant than others in the plastisphere, and a higher Shannon index for abundant fungi, indicating that the plastisphere has a higher diversity and more balanced distribution of abundant fungi than the surroundings [41]. A study found alpha diversities in the plastisphere to be affected by temperature, where they were not significantly different from the surroundings at 15 °C but were lower at 25 °C [11].
Ascomycota is the most common fungal phylum in the plastisphere of various biodegradable and hard-to-degrade plastics or the plastisphere generally [39,40,42]. Common genera of Ascomycota which exist in the plastisphere are Fusarium, Phoma, Paecilomyces, Tetracladium, Cladosporium, and Didymella, to name a few, and the dominant genera may vary between sites or types of forests [40,42]. For instance, Dothideomycetes is the largest class of Ascomycota and it was found to be the dominant fungi at sites polluted by plastics in Kenya [42]. Basidiomycota, Chytridiomycota, and Mortierellomycota fungi are also common in the plastisphere and Mortierellomycota was observed to be enriched in the PE plastisphere [39,42]. Protists were observed to be enriched in the plastisphere of plastic-mulched croplands in China. They were categorized into phototroph, consumer, and parasite (Table 1).
In terms of bacteria, those of the phylum Actinobacteria and Proteobacteria were the most commonly reported in studies on the terrestrial plastisphere [11,22,41,48]. The common genera of Actinobacteria comprise Blastococcus, Iamia, Streptomyces, Nocardia, Mycobacterium, Arthrobacter, Rhodococcus, and Nocardioides (Table 1). The abundance of Actinobacteria was observed to be higher at 25 °C than at 15 °C, and they were enriched in the plastisphere with the application of clover to soil [11,46]. Nocardia colonizes biodegradable and hard-to-degrade plastics in the plastisphere of different types of soils and soils from different regions [5,11]. Rhodococcus seems to be a more common Actinonacteria in the plastisphere of soils from the Swiss Alps while Streptomyces is more common in those from Northern Greenland [5]. The common genera of Proteobacteria in the plastisphere are Sphingomonas, Rhizobacter, Pseudomonas, Rhizobium, Massilia, Lysobacter, Aminobacter, Collimonas, Variovorax, and Caulobacter (Table 1). The dominant genera of Proteobacteria tend to vary on different types of plastics from different soils [5,11]. Biodegradable plastics tend to enhance the plastisphere because they are more accessible sources of carbon than hard-to-degrade plastics. Microbial preferences for certain plastic types may also contribute to the genera variation and this remains poorly understood. For instance, bacteria of the order Saccharimonadales were observed to be associated with polylactic acid (PLA) and a starch-based bioplastic [49]. The dominant bacterial genera in different soils are likely to be different due to factors such as pH, moisture content, organic matter, and vegetation types. Bacteroidota is another bacterial phylum in the plastisphere reported in numerous studies, characterized by the genera Pontibacter, Pedobacter, and Dyadobacter, among others [5,25,40,46]. Additionally, some studies detected Acidobacteriota [11,48] and Firmicutes in the plastisphere [11,22,46,48]. MacLean et al. also found Cyanobacteria of the order Nostocales in the plastisphere of soils from a landfill and a plastic recycling factory [22].
The bacteria in the terrestrial plastisphere may have different functions. Cyanobacteria, for instance, are photosynthetic bacteria. Together with other phototrophs such as Bumilleriopsis, Bracteacoccus, Tetracystis, Chlorochytrium, and Deasonia, they can engage in symbiosis with other heterotrophic bacteria populating the plastisphere through fixing nitrogen and engaging in primary production [22,45]. Their presence could facilitate the establishment of other bacteria in the plastisphere by making carbon more readily available since carbon in hard-to-degrade plastics is not easy to access. Bacteria of the genera Streptomyces and Pseudomonas can be pathogenic. Pseudomonas sp. and Rhodococcus sp. are also known to have the ability to degrade plastics [50]. Parasitic protists like Spongospora and Anurofeca are also present. In certain instances, the plastisphere could be enriched with pathogenic fungi, making plastic debris in the soil a vector for plant pathogens [41].

{kind=link}
Table 1.
Microbial communities associated with terrestrial plastisphere.
| Environment | Diversity | Microbial Phylum/Class | Microbial Genus |
|---|---|---|---|
| PE, polybutylene adipate terephthalate (PBAT), and (PLA microplastics in farmland, China [39] | Alpha diversities of fungal communities were lower in the PBAT/PLA plastisphere than in the PE plastisphere; the soil has higher alpha diversities than the plastisphere. | Ascomycota | Family: Nectriaceae, e.g., Fusarium sp., Cladosporiaceae, Pleosporaceae (enriched on PBAT/PLA), Didymellaceae (enriched on PBAT/PLA), Clavicipitaceae (enriched on PE) |
| Mortierellomycota | Family: Mortierellaceae (enriched on PE) | ||
| Basidiomycota | Not available (NA) | ||
| Firm residues from greenhouses, China [41] | Alpha diversities (Shannon and Simpson) were higher for rare bacteria and fungi; Simpson index was higher for abundant bacteria; Shannon index was higher for abundant fungi. * | Actinobacteriota (abundant bacteria) | Nonomuraea, Blastococcus, Iamia, Kocuria, Streptomyces |
| Crenarchaeota (abundant bacteria) | Nitrososphaeraceae | ||
| Armatimonadota (rare bacteria) | NA | ||
| Bacteroidota (rare bacteria) | Pontibacter | ||
| Myxococcota (rare bacteria) | Bacteriap25 | ||
| Proteobacteria (rare bacteria) | Arenimonas, Skermanella, Steroidobacter, Massilia, Sphingomonas | ||
| Zygomycota (abundant and rare fungi) | Mortierella | ||
| Ascomycota (rare fungi) | Fusarium, Phoma, Paecilomyces | ||
| Poly(butylene succinate-co-butylene adipate) (PBSA) incubated in cropland soils, Germany [40] | Bacterial and fungal richness were lower in the PBSA plastisphere in coniferous forests. * | Ascomycota | In broadleaved forests (200 days): Tetracladium, Mollisina, Cladosporium |
| Ascomycota, Basidiomycota | In coniferous forests (200 days): Venturia, Phoma, Cladosporium, Vishniacozyma (Basidiomycota) | ||
| Ascomycota | In both forests (400 days): Tetracladium, Cyphellophora, Exophiala, Plenodomus | ||
| Gammaproteobacteria | Rhizobacter, Variovorax, Massilia, Pseudomonas | ||
| Alphaproteobacteria | Sphingomonas, Bradyrhizobium, Brevundimonas, Methylobacterium-Methylorubrum, Allorhizobium-Neorhizobium-Pararhizobium-Rhizobium | ||
| Actinobacteria | Pseudonocardia, Nakamurella, Pseudonocardia, Mycobacterium | ||
| Bacteroidota | Pedobacter, Ferruginibacter, Hymenobacter | ||
| PE film in a maize field amended with dried maize straw and living clover [46] | Alpha diversities and abundance of dominant bacteria improved with maize straw and living clover applications. | Bacteroidota | Orders enriched in plastisphere: Sphingomonadales, Flavobacteriales, Chitinophagales |
| Proteobacteria | Orders enriched in plastisphere: Oceanospirillales, Rhodobacterales, Rhizobiales, Caulobacterales | ||
| Firmicutes | Order enriched in plastisphere: Bacillales | ||
| Actinobacteria (enriched by clover in plastisphere) | NA | ||
| Plastic film residues from plastic-mulched croplands, China [45] | Alpha diversities (Chao1 and Simpson) were higher. * | Protists (phyla not reported) | Phototroph: Bumilleriopsis, Bracteacoccus, Tetracystis, Chlorochytrium, Deasonia, Consumer: Pseudoplatyophrya, Filosa–Sarcomonadea_XXX, Capsellina Parasite: Spongospora, Anurofeca |
| PE microplastics incubated in soil [48] | Alpha diversities were not significantly affected. * | Actinobacteria | Nocardia, Arthrobacter, Nocardioides, Aeromicrobium, Rhodococcus, Amycolatopsis |
| Proteobacteria | NA | ||
| Chloroflexi | NA | ||
| Acidobacteriota | NA | ||
| Bacteroidota | Pontibacter | ||
| Firmicutes | NA | ||
| PLA and PE incubated in two different soils at 15 °C and 25 °C [11] | Alpha diversities were not significantly different at 15 °C *; Alpha diversities were lower at 25 °C *; Differences in the relative abundances of the main phyla on PE and PLA were minimal. | Actinobacteriota (at 25 °C, higher abundance in plastisphere and in comparison to 15 °C) | Nocardia (blank clay soil and yellow loam sand soil), Streptomyces (yellow loam sand soil), |
| Proteobacteria (abundance increased at 15 °C) | Lysobacter (blank clay soil and yellow loam sand soil) | ||
| Acidobacteriota | NA | ||
| Chloroflexi | NA | ||
| Firmicutes | NA | ||
| Sites with high human activity and high plastic pollution, Kenya [42] | NA | Ascomycota (Dothideomycetes, 62%; Sordariomycetes, 18%) | Acremonium, Aspergillus, Bipolaris, Cladosporium, Clonostachys, Curvularia, Didymella, Fusarium, Myrothecium, Podospora |
| Basidiomycota | Calvatia, Chlorophyllum, Chroogomphus, Cyathus, Cystobasidium, Hannaella, Minimedusa, Papiliotrema, Rhodotorula | ||
| Chytridiomycota | Gaertneriomyces, Rhizophlyctis, Spizellomyces. Triparticalcar | ||
| An abandoned landfill and a plastic recycling factory, Germany [22] | Simpson index, Shannon index, and Pielou index were lower. * | Proteobacteria | Aminobacter, Pseudox anthozoans, Paracoccus, Rickettsiella, Devosia |
| Actinobacteria | Actinomycetospora, Arthrobacter, Rhodococcus, Rubrobacter, Cellulosimicobium | ||
| Firmicutes | Aeribacillus, Brevibacillus, Planifilum, Paenibacillu | ||
| Cyanobacteria | Nostocales (Order) | ||
| PLA, PBAT, and PE buried/incubated in soils from the Swiss Alps [5] | Alpha diversities were lower. * | Proteobacteria (PLA) | Collimonas, Rhizobiaceae, Variovorax, Pseudomonas |
| Actinobacteria (PLA) | Nocardia, Streptacidiphilus, Catenulispora | ||
| Verrucomicrobia (PLA) | Luteolibacter | ||
| Proteobacteria (PBAT) | Collimonas, Pseudomonas, Variovorax, Mesorhizobium | ||
| Actinobacteria (PBAT) | Nocardia, Rhodococcus, Umezawaea | ||
| Bacteroidetes (PBAT) | Sediminibacterium | ||
| Actinobacteria (PE) | Nocardia, Nocardioides, Rhodococcus | ||
| PLA, PBAT, and PE buried/incubated in soils from Northern Greenland [5] | Alpha diversities were lower. * | Proteobacteria (PLA) | Caulobacter, Brevundimonas, Sphingorhabdus |
| Actinobacteria (PLA) | Nocardia, Nocardioides, Streptomyces | ||
| Bacteroidetes (PLA) | Dyadobacter, Ohtaekwangia | ||
| Proteobacteria (PBAT) | Aquabacterium, Acidovorax, Caulobacter, Brevundimonas | ||
| Actinobacteria (PBAT) | Nocardia, Actinocorallia, Streptomyces | ||
| Actinobacteria (PE) | Nocardia, Nocardioides, Streptomyces, Aeromicrobium | ||
| Proteobacteria (PE) | Aquabacterium, Brevundimonas, Sphingorhabdus, Solimonas |
* Relative to the surrounding soil.
4.2. Aquatic Environment
The plastisphere in the aquatic environment is also occupied by diverse microorganisms. Most studies reviewed reported a higher diversity and richness of microbial communities in the aquatic plastisphere [51,52,53], except for a study by Xu et al. conducted on plastic items incubated in an urban river in China [54]. Delacuvellerie et al. observed the riverine plastisphere to have a higher bacterial richness than the marine environments, indicating the differences in microbial enrichment across the plastisphere in different aquatic environments [55]. It is possible for different microbial communities to demonstrate different diversities. Li et al. reported that the fungal communities in the marine plastisphere have a lower richness than those in the surrounding environment, in contrast to the enriched bacterial communities in the plastisphere [56].
As with the plastisphere in the terrestrial environment, the common bacterial phyla in the aquatic environment are Proteobacteria, Actinobacteria, Bacteroidetes, Firmicutes, and Cyanobacteria [51,54,57]. Additionally, bacteria of the phyla Planctomycetes, Deinococcos-Thermus, Chloroflexi, Acidobacteria, and Verrucomirobia were observed [51,55,56,58]. Certain bacteria reported for the terrestrial plastisphere, particularly the rare bacteria comprising Armatimonadota and Myxococcota, have not been reported in the aquatic plastisphere (Table 1 and Table 2). In fact, cultured Myxococcota is usually derived from aerobic soil environments, though an order of the bacteria called Bradymonadales is known to inhabit saline environments [59]. Deinococcus-Thermus bacteria seem to be more predominant in the aquatic plastisphere (Table 2). The genera of bacteria in the aquatic plastisphere are diverse and they differ on biodegradable and hard-to-degrade plastics. The proteobacteria in the plastisphere of biodegradable plastics in freshwater ecosystems in China consisted of Desulfobacca, Phreatobacter, Phaselicystis, Candidatus, Nitrosomonas and Roseomonas, among others, but those in the plastisphere of hard-to-degrade plastics include Reyranella, Candidatus, Novosphingobium, Syntrophobacter, Bdellovibrio, and Phreotobacter [51]. While certain bacterial genera, such as Phreotobacter, Roseomonas, Arenimonas, and Legionella, were found on both plastics, the abundance could be different, which can probably be attributed to the surface properties of both plastics. Hard-to-degrade plastics generally have more hydrophobic surfaces and this can lead to a strong interface in aquatic environments, which deters microbial attachment. Their surface hydrophobicity decreases with aging and the sorption of organic and inorganic materials, favoring the formation of plastisphere. Additionally, due to differences in functional groups, the plastic particles might adsorb varying types and amounts of organic materials, thus, affecting the types and abundance of bacteria colonizing their surfaces [60]. Similarly, the main Actinobacteria in the plastisphere of biodegradable plastics were Fodinicola, and Lysinimonas but those in the plastisphere of hard-to-degrade plastics were hgcI_clade, Iamia, Tessaracoccus, and Gaiella. Acidobacteria were observed on hard-to-degrade plastics but not biodegradable plastics (Table 2). Nuances in the type and abundance of microorganisms on biodegradable and hard-to-degrade microplastics in an Arctic freshwater lake were also observed, where Proteobacteria like Erythromicrobium and Rhodoferax were more abundant on hard-to-degrade microplastics, but Paranamyces fungi under Chytridiomycota were only found on hard-to-degrade microplastics [52]. The same study only found Arrhenia fungi under Basidiomycota on biodegradable microplastics (Table 2).
Furthermore, Delacuvallerie et al. revealed that dominant plastisphere bacteria, particularly Cyanobacteria, Bacteroidetes, Betaproteobacteria, and Deinococcus-Thermus, were more prevalent in riverine environments, whereas Bacteroidetes, Gammaproteobacteria and Alphaproteobacteria were more prevalent in marine environments [55]. Li et al. confirmed the differences in the dominant microorganisms on the microplastics in freshwater and seawater ecosystems in China, particularly bacteria of the phyla Proteobacteria, Actinobacteria, and Acidobacteria, and fungi of the phylum Chytridiomycota. This conferred the functional differences between the microorganisms in both ecosystems [56]. Moreover, the dominant bacteria on microplastics of different colors incubated in an aquaculture pond were observed to be different, with unclassified bacteria under the family of Burkholderiaceae being predominant on blue and yellow microplastics while those under the family of Rhodobacteraceae were predominant on blue and transparent plastics [53]. Bacteria of Chloroflexi phylum were more prevalent on yellow and transparent microplastics and a type of bacteria from the family Caldilineaceae under the phylum was found exclusively on yellow microplastics [53]. The compositions of bacterial communities on hard-to-degrade plastic films were reported to be different. Hirschia and Erythrobacter bacteria under the class Alphaproteobacteria were more abundant on polyurethane (PU) prepolymer than HDPE, low-density polyethylene (LDPE), and PP, where Phyllobacteriaceae family bacteria were dominant [47]. Flexithrix bacteria were more abundant on PVC plastic films whereas those under the Saprospiraceae family were more abundant on PS (Table 2).
Fungi were observed in the aquatic plastisphere. Like the terrestrial plastisphere, the common fungal phyla are Chytridiomycota, Ascomycota, and Basidiomycota [52,56]. The compositions of fungal communities vary across different habitats and types of plastics (Table 1). Li et al. reported that Ascomycota fungi of the order Saccharomycetales and Chytridiomycota fungi of the order Tremellales dominated the microplastics from freshwater and seawater ecosystems in their study [56]. Eukaryotes comprising those of the phyla Ciliophora, Ochrophyta, Chlorophyta, Fungi, Cryptophyta, and Dinoflagellata were associated with the microplastics in an Arctic freshwater lake [52] (Gonzalez-Pleiter et al., 2021). Ciliophora, and Dinoflagellata, together with other eukaryotes under Archaeplastida, Opisthokonta, and SAR group were also detected in the lacustrine plastisphere in Italy [57] (Table 2).
In comparison to the terrestrial plastisphere, cyanobacteria are more commonly reported in studies on the aquatic plastisphere [51,55,57]. A reason for this is that the microbial communities in the aquatic plastisphere are more reliant on Cyanobacteria as a source of nutrients since carbon is less concentrated and accessible in aquatic environments than in soil environments. Cyanobacteria are a phylum of autotrophic Gram-negative bacteria capable of carrying out photosynthesis. They serve as producers in the plastisphere, hence they are a significant source of carbon for the microorganisms therein [51]. Certain bacteria in the aquatic plastisphere, such as those of the Tenacibaculum genus, are fish pathogens. The known pathogenic species are T. discolor, T. gallaicum, T. maritimum, and T. soleae [61]. Bacteria of the family Vibrionaceae were also reported. Vibrio is the largest genera under this family and certain Vibrio sp. such as Vibro cholera, Vibrio parahaemolyticus, and Vibrio vulnificus are pathogenic [58]. While Vibrio sp. was not enriched in the marine plastisphere in the study of Koh et al., a seagrass pathogen of the genus Labyrinthula was enriched. This implies that the plastisphere can harbor pathogens that are not only harmful to animals but also plants [62].
Table 2.
Microbial communities associated with aquatic plastisphere.
| Environment | Diversity | Microbial Phylum/Class | Microbial Genus |
|---|---|---|---|
| Biodegradable and hard-to-degrade plastics sited in freshwater ecosystems, China [51] | Richness and Shannon indices increased. * | Proteobacteria (biodegradable) | Desulfobacca, Phreatobacter, Phaselicystis, Candidatus, Nitrosomonas, Roseomonas, Chthonobacter, Arenimonas, Sphingopyxis. Legionella, Rhizobium, Porphyrobacter |
| Actinobacteria (biodegradable) | Fodinicola, Lysinimonas, | ||
| Bacteroidetes (biodegradable) | Dinghuibacter, Algoriella, Terrimonas, | ||
| Firmicutes (biodegradable) | Exiguobacterium, Brochothrix, Enterococcus, Carnobacterium, Vagococcus, | ||
| Cyanobacteria (biodegradable) | Cyanobium, Planktothrix | ||
| Planctomycetes (biodegradable) | Pirellula | ||
| Chloroflexi (biodegradable) | uncultured | ||
| Deinococcus-Thermus (biodegradable) | Deinococcus | ||
| Proteobacteria (hard-to-degrade) | Reyranella, Candidatus, Novosphingobium, Syntrophobacter, Bdellovibrio, Phreotobacter, Roseomonas, Arenimonas, Legionella, Nitrosomonas, Rhizobium | ||
| Bacteroidetes (hard-to-degrade) | Flavobacterium, Paludibacter, Bacteriodes, | ||
| Firmicutes (hard-to-degrade) | Carnobacterium, Clostridium, Exiguobacterium, Lactococcus, Proteiniclasticum, Catenibacterium | ||
| Planctomycetes (hard-to-degrade) | Singulisphaera, Fimbriiglobus, Planctopirus, Pirellula, | ||
| Chloroflexi (hard-to-degrade) | Anaerolinea | ||
| Cyanobacteria (hard-to-degrade) | Cyanobium, Planktothrix, | ||
| Actinobacteria (hard-to-degrade) | hgcI_clade, Iamia, Tessaracoccus, Gaiella | ||
| Acidobacteria (hard-to-degrade) | Geothrix, Stenotrophobacter | ||
| Deinococcus-Thermus (hard-to-degrade) | Deinococcus | ||
| Garbage bags, shopping bags, and plastic bottles incubated in an urban river, China [54] | The diversity indices of bacterial community on microplastics were lower than those of free-living bacteria. | Gammaproteobacteria | NA |
| Alphaproteobacteria | NA | ||
| Bacteroidia | NA | ||
| Plastic debris from river to sea, France [55] | The river had higher richness indices than marine environments. | Cyanobacteria (10.7%) (river) | Chamaesiphon (8.3%) |
| Bacteroidetes (7.5%) (river) | Hymenobacter (1.3%) | ||
| Betaproteobacteria (7.5%) (river) | |||
| Deinococcus-Thermus (5.2%) (river) | Deinococcus (5.2%) | ||
| Bacteroidetes (26%) (marine) | Cytophaga, Saprospira, Tenacibaculum | ||
| Gammaproteobacteria (10%) (marine) | Unclassified | ||
| Alphaproteobacteria (22%) (marine) | Unclassified | ||
| Microplastics from lakes in Italy [57] | NA | Archaeplastida (kingdom) | Chloroplastida (Phylum) → Ulvellaceae, Scenedesmaceae, Desmidiaceae |
| Opisthokonta (clade) | Holozoa (clade) → Metazoa | ||
| SAR group | Alveolata (Infrakingdom) → Dinoflagellates, Ciliophora; Stramenopiles (Phylum) → Pennales, Peronosporomycetes, Bacillariophyceae | ||
| Cyanobacteria | Family: Nostocaceae (family) | ||
| Proteobacteria | Family: Burkolderaceae, Rhodobacteraceae, Sphingomonadaceae, Rhizobiaceae | ||
| Bacteroidota | Family: Flavobacteraceae, Bacteroidaceae, Rikenellaceae | ||
| Firmicutes | Family: Clostridiaceae | ||
| Deinococcus-Thermus | Family: Deinococcaceae | ||
| Polyhydroxybutyrate (PHB), HDPE, and LDPE microplastics in an Arctic freshwater lake [52] | Shannon index was higher. * | Proteobacteria (76.1%) | Mycoplana (24.8%) (dominate biodegradable and hard-to-degrade microplastics), Erythromicrobium (6.3%) (more abundant on hard-to-degrade microplastics), Comamonadaceae_unclassified (4.6%), Rhodobacter (4.1%) (more abundant on hard-to degrade microplastics), Rhodoferax (2.9%) (more abundant on biodegradable microplastics), Moraxellaceae_unclassified (2.7%) (more abundant on biodegradable microplastics), Sphingomonadaceae_unclassified (2.6%), Zymomonas (2.5%) |
| Cyanobacteria (7.9%) | NA | ||
| Bacteroidetes (6.3%) | NA | ||
| Actinobacteria (3.1%) | NA | ||
| Verrucomicrobia (2.3%) | NA | ||
| Chytridiomycota (52.0%) | Betamyces (22.2%) (predominant on hard-to-degrade and biodegradable microplastics), Chytridiomycota_unclassified (19.9%), Paranamyces (1.0%) (only on hard-to-degrade microplastics), Polychytriales_unclassified (1.0%) | ||
| Ascomycota (7.3%) | NA | ||
| Basidiomycota (4.3%) | Cryptococcus (1.4%) (only on hard-to-degrade microplastics), Arrhenia (1.2%) (only on biodegradable microplastics) | ||
| Ciliophora (62.8%) | Stentor (45.9%) (dominant on both microplastics), Vorticella (7.5%), Uroleptus (3.5%), Sessilida_unclassified (2.6%), Chilodonellidae_unclassified (1.5%) | ||
| Ochrophyta (15.9%) | Chrysophyceae_Clade-C_unclassified (3.9%), Epipyxis (2.7%), Chrysophyceae_X_unclassified (1.9%) | ||
| Chlorophyta (8.2%) | Tetraselmis (2.9%), Chlamydomonas (1.4%) | ||
| Fungi (4.7%) | NA | ||
| Cryptophyta (4.1%) | Cryptomonas (3.8%) | ||
| Dinoflagellata (1.7%) | NA | ||
| Microplastics from three freshwater and three seawater ecosystems, China [56] | The richness of bacterial communities was higher; the richness of fungal communities was lower. * | Proteobacteria (functional difference) | Order: Kiloniellales, Methylophilales, Legionellales, Tepidisphaerales, Methylococcales |
| Bacteroidetes | Order: Sphingomonadales | ||
| Actinobacteria (functional difference) | Order: Acidimicrobiales, Actinomycetales, Nitriliruptorales, | ||
| Firmicutes | Order: Bacillales | ||
| Verrucomicrobia | Order: Opitutales | ||
| Planctomycetes | NA | ||
| Acidobacteria (functional difference) | NA | ||
| Ascomycota | Order: Saccharomycetales | ||
| Basidiomycota | NA | ||
| Chytridiomycota (functional difference) | Order: Tremellales | ||
| Rozellomycota | NA | ||
| Mortierellomycota | NA | ||
| Cercozoa | NA | ||
| Yellow, blue, and transparent microplastics incubated in an aquaculture pond, China [53] | Alpha diversities of the plastisphere were higher. * | Proteobacteria | Allorhizobium-Neorhizobium-Pararhizobium-Rhizobium, Aquabacterium, Herbaspirillum, Bradyrhizobium, unclassified_f__Burkholderiaceae (blue and yellow), unclassified_f__Rhodobacteraceae (blue and transparent), Pseudomonas (yellow), Aquabacterium (blue) |
| Chloroflexi (yellow and transparent) | norank_f__Caldilineaceae (yellow) | ||
| Actinobacteria (transparent) | NA | ||
| Dependentiae (transparent) | NA | ||
| Firmicutes (blue) | NA | ||
| Microplastics in lentic Italian ecosystems [63] | NA | Alphaproteobacteria | Sphingorhabdus, Altererythrobacter, Sphingomonas, Porphyrobacter, Novosphingobium, Pseudorhodobacter, Rhodobacter |
| Betaproteobacteria | Aquabacterium, Massilia, Acidovorax, Hydrogenophaga, Ideonella, Leptothrix | ||
| Gammaproteobacteria | NA | ||
| Bacteroidetes | Muricauda, Mesoflavibacter | ||
| Cyanobacteria | Aphanothece, Chroococcus, Synechocystis Phormidium, Leptolyngbya, Pseudoanabaena, Calothrix | ||
| Plastic debris in the Urumqi River, China [58] | Alpha diversities of bacteria on plastic debris were mostly higher. * | Proteobacteria | Pseudomonas, Acinetobacter, Alteromonadaceae (Family), Vibrionaceae (Family) |
| Bacteroidetes | NA | ||
| Cyanobacteria | NA | ||
| Firmicutes | NA | ||
| Actinobacteria | Nocardiaceae (Family) | ||
| Chloroflexi | NA | ||
| Nitrospirae | NA | ||
| Plastic films incubated in a simulated seawater environment [47] | NA. The study aimed to identify plastic-specific bacteria | Gammaproteobacteria | NA |
| Alphaproteobacteria | Hirschia, Erythrobacter (PU prepolymer); uncultured Phyllobacteriaceae family (HDPE, LDPE, PP) | ||
| Sphingobacteria (on PLA) | NA | ||
| Bacteroidota | Flexithrix (PVC); Saprospiraceae family (PS) | ||
| Plastic debris from off-coast seawater, Singapore [53] | Alpha diversities of the prokaryotes were higher but those of the eukaryotes were lower * | Cyanobacteria | Pleurocapsa PCC-7319, Schizothrix LEGE 07164, Myxosarcina GI1, Chroococcidiopsis PCC-6712 |
| Bacteroidota | Rubrivirga, Aquimarina, Muricauda, Rubricoccus, Winogradskyella, Maribacter, Lewinella | ||
| Proteobacteria | Erythrobacter, Sphingomicrobium, Nioella, Sphingomonas, Hyphomonas | ||
| Deinococcota | Truepera | ||
| Acidobacteriota | Blastocatella | ||
| Ochrophyta | Navicula, Amphora, Nitzchia, Psammodictyon, Cymbella | ||
| Ciliophora | Acineta_1, Holosticha, Ephelota, Trochilia, Chlamydonellopsis | ||
| Chlorophyta | Ulva, Acrochaete, Ruthnielsenia, | ||
| Cercozoa | Thalassomyxa, Massisteria, Nanofila | ||
| Dinoflagellata | Amphidinium, Symbiodinium, Durusdinium | ||
| Sagenista | Labyrinthula, Labyrinthulaceae_ANT10_3, Aplanochytrium |
* Relative to the surrounding environment or the free-living bacteria.
5. Factors Influencing the Composition of the Plastisphere
Based on the studies reviewed in Section 4, the factors affecting the composition of the plastisphere can generally be categorized into three overarching categories, namely duration, environmental factors, and the properties of plastics (Figure 1).
5.1. Duration
Studies have shown that the terrestrial and aquatic plastisphere is not static. The microbial community changes over time. Wright et al. followed the microbial succession in the PET plastisphere over 6 weeks of incubation with microbial communities from marine plastic debris and found that the microbial communities in the plastisphere were significantly distinct from the inoculum [64]. Gammaproteobacteria, particularly Alteromonadaceae, and Thalassospiraceae, were particularly abundant in the early incubation stage, while the abundance of Vibrionaceae was relatively constant over time. The abundance of Alcanivoraceae increased towards the end of the experiment [64]. Similarly, another study revealed Gammaproteobacteria as the dominant bacterial phylum, particularly the Oleibacter sp., in the coastal plastisphere during the plastisphere-forming stage but was later succeeded by Alphaproteobacteria and Flavobacteria. Succession constantly occurred in the plastisphere, favoring inter-specific cooperation instead of competition [65]. The pioneer microbial community is thought to be able to attach to the hydrophobic surfaces of microplastics through secreting exopolysaccharides, which facilitate the attachment of other microorganisms. Zhang et al. reported the presence of Alteromonas, Neptuniibacter, Roseobacter, and Thalassobius in the early marine plastisphere, together with diatom and cyanobacteria. Bacteroidetes, particularly Flavobacteriaceae, subsequently increased in abundance, which is probably because they can feed on the organic materials produced by the pioneer colonizers [66]. However, a study reported Flavobacteriaceae as a pioneering species in the marine plastisphere of PE, in addition to Rhodobacteraceae and Microtrichaceae. Bacillaceae and Moraxellaceae became dominant from day 75 onwards before Flavobacteriaceae and Rhodobacteraceae made a comeback on day 135 [67]. Proteobacteria are consistent pioneer colonizers in multiple studies. Adhesion to the plastic surfaces is deemed to increase with microbial succession where subsequent colonizers have pili, adhesion proteins, and mechanisms that enable them to strongly and, sometimes, irreversibly attach to the plastics [68].
Studies on the marine plastisphere have also found the presence of phototrophic microorganisms on microplastics exposed to sunlight. A common phototroph is cyanobacteria. Cyanobacteria were observed to be more abundant in the riverine plastisphere in one study, indicating that their primary production could serve as a source of nutrients to other microorganisms therein [55]. Their presence might alter the availability of nutrients on the surface of microplastics, which subsequently affects the composition of microorganisms [69]. Variation in the microbial composition of the terrestrial plastisphere occurs with time. Having adapted to hydrophobic plastic surfaces or overcome electrostatic repulsion, bacteria produced more adhesion proteins and extracellular materials to better adhere to the surfaces. The dominant bacteria on microplastics incubated at a dump site changed over 14 months. Proteobacteria were consistently present on the microplastics but their abundance fluctuated in response to that of Chloroflexi. On PE and PP, the abundance of Actinobacteria was high but declined sharply after nine months of incubation [70]. In addition, Ju et al. observed changes in the microbial composition on the surface of biodegradable plastic mulches with time. Similar to the extant studies, Proteobacteria, Actinobacteria, Acidobacteria, Chloroflexi, and Bacteriodetes were the most common bacteria phyla on the plastic mulches [71]. The orders of bacteria demonstrated a clearer variation with time. The abundances of Burkholderiales and Pseudonocardiales fluctuated for 5 months though they were consistently present on the mulches. In PLA, for instance, the Rhizobiales population increased steadily before a decline after 4 months [71]. The succession of microbiota in the terrestrial and aquatic plastisphere is common as the pioneer colonies make the microenvironment more suitable for colonization by other microorganisms. Succession is linked to the environmental conditions, especially the predominant microorganisms present in the surroundings, and the microplastic properties since studies have shown that microbial succession on different types of microplastics diverges [64,70]. Degraders and pathogens were found to be enriched more on the surfaces of biodegradable plastic mulches than the hard-to-degrade ones [71].
5.2. Plastic Properties
Be it in terrestrial or aquatic environments, plastic properties have a significant influence on the plastisphere formed. According to the previous section, the microorganisms in the plastisphere are selectively enriched on different plastic types. For instance, bacteria of the families Nectriaceae, Cladosporiaceae, and Pleosporaceae were more abundant on biodegradable microplastics like PBAT and PLA, while those of Clavicipitaceae were more abundant on PE in a terrestrial environment (Table 1). Ruthi et al. observed different abundances of Proteobacteria, Actinobacteria, and Bacteoidetes on PLA, PBAT, and PE buried in the soils of two distinct habitats [5]. Similarly, Miao et al. also found that bacteria of common phyla, particularly Proteobacteria, Actinobacteria, Bacteroidetes, Firmicutes, Cyanobacteria, Planctomycetes, Chloroflexi, and Deinococcus-Thermus, could be differentially enriched in the plastispheres of biodegradable and hard-to-degrade plastics in freshwater ecosystems [51]. Wen et al. revealed that the colors of microplastics have an effect on the microbial composition of the associated plastisphere, where Chloroflexi bacteria were enriched on yellow and transparent microplastics, and Firmicutes bacteria were enriched on blue microplastics. The differences in microbiota could be attributed to the different wavelengths of light reflected by the microplastics [53].
Additionally, studies have pointed to the noticeable differences between the microbial compositions among hard-to-degrade microplastics, where the microbiota on PS microplastics was widely different from that on PE and PP [72]. Even the sizes of plastic debris have an implication for the microbial composition of the plastisphere. Debroas et al. observed that Alphaproteobacteria and Gammaproteobacteria were more dominant on the surfaces of PET and PE mesoplastics, while Burkholderiales were more abundant on the surfaces of PE microplastics in the plastic debris retrieved from the North Atlantic Ocean [73]. Sulfate-reducing bacteria were found to be enriched on biodegradable polyhydroxyalkanoates (PHA) [6]. The surface properties of plastic particles affect the microorganisms colonizing the plastisphere. Different plastics have different surface charges, which can lead to varying adsorption interactions of microorganisms [14]. Bacteria equipped with certain structures can more easily overcome the repulsive forces between the negatively charged cells and the plastic surface and, thus, are better able to colonize the surfaces of certain plastics. The additives leached from the plastic particles also influence the types of microorganisms that can colonize their surfaces [74]. Over time, microplastics in the environment undergo physical and chemical changes due to exposure to sunlight, oxygen, and other environmental factors. This aging process alters the surface properties of microplastics, and hence the structure of the plastisphere [30]. Aging may increase the surface area, polarity, and roughness of microplastics, making the microplastics more favorable to some microorganisms and less favorable to others [75]. A study not targeting the plastisphere showed organic plastic additives could adversely affect autotrophic and heterotrophic bacteria in artificial seawater [76].
5.3. Environmental Factors
Environmental factors encompass a large group of factors that affect the composition of the plastisphere. It can broadly be divided into biotic and abiotic. Biotic environmental factors center on the microorganisms and organisms in the environment (Figure 1). The dominant bacteria in the environment often determine the bacteria in the plastisphere. This is called bacterial hitchhiking, causing the bacterial compositions on different substrates to resemble their surroundings [73]. This is demonstrated through the predominance of bacteria of the class Gammaproteobacteria in the young plastisphere as they are abundant in the environment. These hitchhikers are the pioneer colonizers that alter the plastic surfaces for the colonization of other microorganisms [14]. Cyanobacteria and other phototrophs can engage in symbiotic relationships with the heterotrophs by converting carbon dioxide into organic carbon through photosynthesis, which the heterotrophs can use for growth and maintenance. In return, the heterotrophs can contribute to the degradation of the plastics and their death may release substances which the phototrophs can use. Symbiosis is important for the establishment of plastisphere in nutrient-depleted and extreme environments [14]. The presence of predators, such as protozoa and bacteriophages, contributes to shaping the composition of the plastisphere. Ingestion of microplastics by aquatic organisms may also result in the development of the plastisphere that is compositionally similar to the microbiome in the guts of the organisms [77].
The abiotic environmental factors range from nutrient availability to the physicochemical parameters of the environmental media, such as light, pH, temperature, and salinity (Figure 1). A study revealed that the level of nutrient enrichment in lakes affected the heterotrophic activities in the plastisphere. The functional diversity profiles of the lacustrine plastisphere were different from the surrounding water. The plastisphere of the oligo-mesotrophic lake had a higher functional richness and biomass than the eutrophic lake and the ambient water [78]. Salinity seems to significantly affect the estuarine and marine plastisphere, and it was observed to correlate positively with the abundance of Vibrio sp. [77]. Vibrio sp. are potential pathogens and their enrichment on microplastics indicates that microplastics could be vectors for pathogens [79]. Limited carbon resources in the aquatic environment may drive microbial succession in the plastisphere towards communities that tightly adhere to the microplastics [47]. Ruthi et al. reported different abundances and compositions of bacteria under the phyla of Proteobacteria, Actinobacteria, and Bacteroidetes in biodegradable and hard-to-degrade microplastics buried in soils from the Swiss Alps and Northern Greenland, implying that plastisphere characteristics can be site-specific [5]. Different temperatures resulted in the selective enrichment of certain bacteria. Sun et al. reported lower alpha diversities of the plastisphere than the surroundings at 25 °C but the difference was not apparent at 15 °C [11]. The plastisphere had a higher abundance of Actinobacteriota at 25 °C and a higher abundance of proteobacteria at 15 °C. At 25 °C, bacteria of different genera of Actinobacteriota were also differentially enriched on microplastics in different soils (Table 1), showing that the effects of the abiotic environment are highly diverse and complex. Even different amendments applied on the same site could result in the selective enrichment of the microbial communities [46]. The availability of light is crucial for the colonization of the plastisphere by phototrophs. As different microorganisms have varying pH preferences, the pH of the surrounding environment can determine the types of microorganisms in the plastisphere [8]. The multitude of widely varying abiotic environmental factors confers uniqueness to the plastisphere. While the microbial phyla in the plastisphere are largely similar, the abundance and diversity of microbial species are often distinct, even for plastic particles on the same site.
6. Conclusions
The plastisphere has gained attention due to the unique networks of bacteria that dwell around plastic particles, forming new ecological niches. The terrestrial and aquatic plastisphere formation follows a similar track, with plastic particles occupied by opportunistic microorganisms or hitchhikers. These pioneer microorganisms are often represented by the dominant microorganisms in the surrounding environment. Colonization of the plastic particles by the pioneer colonies modifies the surface properties of the particles for colonization by other microorganisms. The most common microorganisms in the plastisphere are bacteria of the phyla Proteobacteria, Actinobacteria, Bacteroidota, Firmicutes, Chloroflexi, Acidobacteria, and Cyanobacteria, as well as fungi of the phyla Ascomycota, Basidiomycota, and Chytridiomycota. Protists, for instance, those in the phyla Ciliophora, Ochrophyta, and Chlorophyta, also exist in the plastisphere. While the formation and succession processes of the terrestrial and aquatic plastispheres are the same, the dynamics and behaviors of the plastisphere in soil and aquatic environments differ. The soil plastisphere does not exhibit the mobility of the oceanic plastisphere and is significantly influenced by the characteristics of the soil anchoring it. The factors affecting the plastisphere can generally be categorized into duration, plastic properties, and environmental factors. Duration signifies the succession and changes in the microbial communities in the plastisphere over time towards the establishment of the communities that tightly adhere to the plastic particles. The abundance and dominant species of microorganisms on different plastic particles may be nuanced due to the type, surface properties, size, color, and aging of the particles. Chemicals leached from the particles can also affect the microbiome of the particles. Environmental factors are highly diverse, ranging from biotic factors characterized by the dominant microorganisms in the ambient environment and interspecies interactions to abiotic factors encompassing pH, temperature, nutrient availability, soil properties, salinity, and light.
Understanding the characteristics of the plastisphere enables a better understanding of the functions of microplastics as vectors of pathogens. Certain pathogens have been found to exist and, sometimes, increase in the plastisphere. Additionally, the plastisphere may contribute to the breakdown of plastic debris. Certain microorganisms in the plastisphere can utilize plastics as a source of carbon. Studying the plastisphere also contributes to a better insight into the diversity of microorganisms in the soil and aquatic environments since some microorganisms that survive on the plastic debris are not typically encountered in the surrounding environment. However, there is much to be explored concerning the plastisphere, particularly microbe-microbe interactions, microbe-substrate interactions, the abiotic factors shaping the plastisphere, and the effects of plastic chemical leaching on the plastisphere.
Funding
This research received no external funding.
Acknowledgments
The author wishes to thank the University of Arizona for the administrative support given.
Conflicts of Interest
The author declares no conflicts of interest.
References
- OECD Global Plastic Outlook: Policy Scenarios to 2060. Available online: https://0-www-oecd--ilibrary-org.brum.beds.ac.uk/sites/aa1edf33-en/index.html?itemId=/content/publication/aa1edf33-en (accessed on 1 February 2024).
- Tang, K.H.D. Effects of Microplastics on Agriculture: A Mini-review. Asian J. Environ. Ecol. 2020, 13, 1–9. [Google Scholar] [CrossRef]
- Tang, K.H.D. Abundance of Microplastics in Wastewater Treatment Sludge. J. Hum. Earth Futur. 2022, 3, 138–146. [Google Scholar] [CrossRef]
- Rillig, M.C.; Kim, S.W.; Zhu, Y.-G. The soil plastisphere. Nat. Rev. Microbiol. 2024, 22, 64–74. [Google Scholar] [CrossRef] [PubMed]
- Rüthi, J.; Bölsterli, D.; Pardi-Comensoli, L.; Brunner, I.; Frey, B. The “Plastisphere” of Biodegradable Plastics Is Characterized by Specific Microbial Taxa of Alpine and Arctic Soils. Front. Environ. Sci. 2020, 8, 562263. [Google Scholar] [CrossRef]
- Pinnell, L.J.; Turner, J.W. Shotgun Metagenomics Reveals the Benthic Microbial Community Response to Plastic and Bioplastic in a Coastal Marine Environment. Front. Microbiol. 2019, 10, 1252. [Google Scholar] [CrossRef] [PubMed]
- Oberbeckmann, S.; Loeder, M.G.J.; Gerdts, G.; Osborn, A.M. Spatial and seasonal variation in diversity and structure of microbial biofilms on marine plastics in Northern European waters. FEMS Microbiol. Ecol. 2014, 90, 478–492. [Google Scholar] [CrossRef] [PubMed]
- Zettler, E.R.; Mincer, T.J.; Amaral-Zettler, L.A. Life in the “Plastisphere”: Microbial Communities on Plastic Marine Debris. Environ. Sci. Technol. 2013, 47, 7137–7146. [Google Scholar] [CrossRef] [PubMed]
- Stabnikova, O.; Stabnikov, V.; Marinin, A.; Klavins, M.; Klavins, L.; Vaseashta, A. Microbial Life on the Surface of Microplastics in Natural Waters. Appl. Sci. 2021, 11, 11692. [Google Scholar] [CrossRef]
- Du, Y.; Liu, X.; Dong, X.; Yin, Z. A review on marine plastisphere: Biodiversity, formation, and role in degradation. Comput. Struct. Biotechnol. J. 2022, 20, 975–988. [Google Scholar] [CrossRef]
- Sun, Y.; Shi, J.; Wang, X.; Ding, C.; Wang, J. Deciphering the Mechanisms Shaping the Plastisphere Microbiota in Soil. mSystems 2022, 7, e00352-22. [Google Scholar] [CrossRef]
- Wang, C.; Wang, L.; Ok, Y.S.; Tsang, D.C.W.; Hou, D. Soil plastisphere: Exploration methods, influencing factors, and ecological insights. J. Hazard. Mater. 2022, 430, 128503. [Google Scholar] [CrossRef]
- Arias-Andres, M. Who is where in the Plastisphere, and why does it matter? Mol. Ecol. Resour. 2020, 20, 617–619. [Google Scholar] [CrossRef]
- Dey, S.; Rout, A.K.; Behera, B.K.; Ghosh, K. Plastisphere community assemblage of aquatic environment: Plastic-microbe interaction, role in degradation and characterization technologies. Environ. Microbiome 2022, 17, 32. [Google Scholar] [CrossRef] [PubMed]
- Amaral-Zettler, L.A.; Zettler, E.R.; Slikas, B.; Boyd, G.D.; Melvin, D.W.; Morrall, C.E.; Proskurowski, G.; Mincer, T.J. The biogeography of the Plastisphere: Implications for policy. Front. Ecol. Environ. 2015, 13, 541–546. [Google Scholar] [CrossRef] [PubMed]
- Li, C.; Tang, K.H.D. Effects of pH and Temperature on the Leaching of Di (2-Ethylhexyl) Phthalate and Di-n-butyl Phthalate from Microplastics in Simulated Marine Environment. Biointerface Res. Appl. Chem. 2023, 13, 269. [Google Scholar]
- Conradie, W.; Dorfling, C.; Chimphango, A.; Booth, A.M.; Sørensen, L.; Akdogan, G. Investigating the Physicochemical Property Changes of Plastic Packaging Exposed to UV Irradiation and Different Aqueous Environments. Microplastics 2022, 1, 456–476. [Google Scholar] [CrossRef]
- Barros, J.; Seena, S. Plastisphere in freshwaters: An emerging concern. Environ. Pollut. 2021, 290, 118123. [Google Scholar] [CrossRef]
- Harrison, J.P.; Hoellein, T.J.; Sapp, M.; Tagg, A.S.; Ju-Nam, Y.; Ojeda, J.J. Microplastic-Associated Biofilms: A Comparison of Freshwater and Marine Environments. In Freshwater Microplastics: Emerging Environmental Contaminants? Wagner, M., Lambert, S., Eds.; Springer International Publishing: Cham, Switzerland, 2018; pp. 181–201. ISBN 978-3-319-61615-5. [Google Scholar]
- Su, X.; Yang, K. Exploring the blindspot: The soil plastisphere. Soil Ecol. Lett. 2023, 6, 230209. [Google Scholar] [CrossRef]
- Zhang, G.S.; Liu, Y.F. The distribution of microplastics in soil aggregate fractions in southwestern China. Sci. Total Environ. 2018, 642, 12–20. [Google Scholar] [CrossRef]
- MacLean, J.; Mayanna, S.; Benning, L.G.; Horn, F.; Bartholomäus, A.; Wiesner, Y.; Wagner, D.; Liebner, S. The Terrestrial Plastisphere: Diversity and Polymer-Colonizing Potential of Plastic-Associated Microbial Communities in Soil. Microorganisms 2021, 9, 1876. [Google Scholar] [CrossRef]
- Kuzyakov, Y.; Blagodatskaya, E. Microbial hotspots and hot moments in soil: Concept & review. Soil Biol. Biochem. 2015, 83, 184–199. [Google Scholar] [CrossRef]
- Dodhia, M.S.; Rogers, K.L.; Fernández-Juárez, V.; Carreres-Calabuig, J.A.; Löscher, C.R.; Tisserand, A.A.; Keulen, N.; Riemann, L.; Shashoua, Y.; Posth, N.R. Microbe-mineral interactions in the Plastisphere: Coastal biogeochemistry and consequences for degradation of plastics. Front. Mar. Sci. 2023, 10, 1134815. [Google Scholar] [CrossRef]
- Liu, S.; Zhang, X.; Zeng, K.; He, C.; Huang, Y.; Xin, G.; Huang, X. Insights into eco-corona formation and its role in the biological effects of nanomaterials from a molecular mechanisms perspective. Sci. Total Environ. 2023, 858, 159867. [Google Scholar] [CrossRef] [PubMed]
- Shi, X.; Chen, Z.; Wei, W.; Chen, J.; Ni, B.-J. Toxicity of micro/nanoplastics in the environment: Roles of plastisphere and eco-corona. Soil Environ. Health 2023, 1, 100002. [Google Scholar] [CrossRef]
- Liu, S.; Junaid, M.; Liao, H.; Liu, X.; Wu, Y.; Wang, J. Eco-corona formation and associated ecotoxicological impacts of nanoplastics in the environment. Sci. Total Environ. 2022, 836, 155703. [Google Scholar] [CrossRef] [PubMed]
- ter Halle, A.; Ghiglione, J.F. Nanoplastics: A Complex, Polluting Terra Incognita. Environ. Sci. Technol. 2021, 55, 14466–14469. [Google Scholar] [CrossRef] [PubMed]
- Besemer, K. Microbial Biodiversity in Natural Biofilms. In Aquatic Biofilms: Ecology, Water Quality and Wastewater Treatment; Caister Academic Press: Poole, UK, 2016; pp. 63–88. [Google Scholar]
- Tang, K.H.D. Microplastics in agricultural soils in China: Sources, impacts and solutions. Environ. Pollut. 2023, 322, 121235. [Google Scholar] [CrossRef]
- Campanale, C.; Galafassi, S.; Savino, I.; Massarelli, C.; Ancona, V.; Volta, P.; Uricchio, V.F. Microplastics pollution in the terrestrial environments: Poorly known diffuse sources and implications for plants. Sci. Total Environ. 2022, 805, 150431. [Google Scholar] [CrossRef]
- Tang, K.H.D. Microplastics in and Near Landlocked Countries of Central and East Asia: A Review of Occurrence and Characteristics. Trop. Aquat. Soil Pollut. 2023, 3, 120–130. [Google Scholar] [CrossRef]
- Weber, C.J.; Opp, C. Spatial patterns of mesoplastics and coarse microplastics in floodplain soils as resulting from land use and fluvial processes. Environ. Pollut. 2020, 267, 115390. [Google Scholar] [CrossRef]
- McCormick, A.R.; Hoellein, T.J. Anthropogenic litter is abundant, diverse, and mobile in urban rivers: Insights from cross-ecosystem analyses using ecosystem and community ecology tools. Limnol. Oceanogr. 2016, 61, 1718–1734. [Google Scholar] [CrossRef]
- Vercauteren, M.; Lambert, S.; Hoogerwerf, E.; Janssen, C.R.; Asselman, J. Microplastic-specific biofilm growth determines the vertical transport of plastics in freshwater. Sci. Total Environ. 2024, 910, 168399. [Google Scholar] [CrossRef]
- Amaral-Zettler, L.A.; Zettler, E.R.; Mincer, T.J.; Klaassen, M.A.; Gallager, S.M. Biofouling impacts on polyethylene density and sinking in coastal waters: A macro/micro tipping point? Water Res. 2021, 201, 117289. [Google Scholar] [CrossRef]
- Kaiser, D.; Kowalski, N.; Waniek, J.J. Effects of biofouling on the sinking behavior of microplastics. Environ. Res. Lett. 2017, 12, 124003. [Google Scholar] [CrossRef]
- Kooi, M.; van Nes, E.H.; Scheffer, M.; Koelmans, A.A. Ups and Downs in the Ocean: Effects of Biofouling on Vertical Transport of Microplastics. Environ. Sci. Technol. 2017, 51, 7963–7971. [Google Scholar] [CrossRef]
- Li, K.; Xu, L.; Bai, X.; Zhang, G.; Zhang, M.; Huang, Y. Differential fungal assemblages and functions between the plastisphere of biodegradable and conventional microplastics in farmland. Sci. Total Environ. 2024, 906, 167478. [Google Scholar] [CrossRef]
- Tanunchai, B.; Ji, L.; Schröder, O.; Gawol, S.J.; Geissler, A.; Wahdan, S.F.M.; Buscot, F.; Kalkhof, S.; Schulze, E.-D.; Noll, M.; et al. Fate of a biodegradable plastic in forest soil: Dominant tree species and forest types drive changes in microbial community assembly, influence the composition of plastisphere, and affect poly(butylene succinate-co-adipate) degradation. Sci. Total Environ. 2023, 873, 162230. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Gao, W.; Wang, C.; Gao, M. Distinct distribution patterns and functional potentials of rare and abundant microorganisms between plastisphere and soils. Sci. Total Environ. 2023, 873, 162413. [Google Scholar] [CrossRef]
- Gkoutselis, G.; Rohrbach, S.; Harjes, J.; Obst, M.; Brachmann, A.; Horn, M.A.; Rambold, G. Microplastics accumulate fungal pathogens in terrestrial ecosystems. Sci. Rep. 2021, 11, 13214. [Google Scholar] [CrossRef] [PubMed]
- Willis, A.D. Rarefaction, Alpha Diversity, and Statistics. Front. Microbiol. 2019, 10, 2407. [Google Scholar] [CrossRef]
- Jost, L. Partitioning diversity into independent alpha and beta components. Ecology 2007, 88, 2427–2439. [Google Scholar] [CrossRef]
- Li, Y.; Yang, R.; Guo, L.; Gao, W.; Su, P.; Xu, Z.; Xiao, H.; Ma, Z.; Liu, X.; Gao, P.; et al. The composition, biotic network, and assembly of plastisphere protistan taxonomic and functional communities in plastic-mulching croplands. J. Hazard. Mater. 2022, 430, 128390. [Google Scholar] [CrossRef]
- Zhao, Z.-Y.; Wang, P.-Y.; Xiong, X.-B.; Zhou, R.; Li, F.-M.; Cheng, Z.-G.; Wang, W.; Mo, F.; Cheruiyot, K.W.; Wang, W.-Y.; et al. Plant biomass mediates the decomposition of polythene film-sourced pollutants in soil through plastisphere bacteria island effect. Environ. Int. 2023, 178, 108114. [Google Scholar] [CrossRef] [PubMed]
- Kirstein, I.V.; Wichels, A.; Gullans, E.; Krohne, G.; Gerdts, G. The Plastisphere—Uncovering tightly attached plastic “specific” microorganisms. PLoS ONE 2019, 14, e0215859. [Google Scholar] [CrossRef] [PubMed]
- Ya, H.; Xing, Y.; Zhang, T.; Lv, M.; Jiang, B. LDPE microplastics affect soil microbial community and form a unique plastisphere on microplastics. Appl. Soil Ecol. 2022, 180, 104623. [Google Scholar] [CrossRef]
- Wicaksono, J.A.; Purwadaria, T.; Yulandi, A.; Tan, W.A. Bacterial dynamics during the burial of starch-based bioplastic and oxo-low-density-polyethylene in compost soil. BMC Microbiol. 2022, 22, 309. [Google Scholar] [CrossRef] [PubMed]
- Yoshida, S.; Hiraga, K.; Takehana, T.; Taniguchi, I.; Yamaji, H.; Maeda, Y.; Toyohara, K.; Miyamoto, K.; Kimura, Y.; Oda, K. A bacterium that degrades and assimilates poly(ethylene terephthalate). Science 2016, 351, 1196–1199. [Google Scholar] [CrossRef] [PubMed]
- Miao, L.; Li, W.; Adyel, T.M.; Yao, Y.; Deng, Y.; Wu, J.; Zhou, Y.; Yu, Y.; Hou, J. Spatio-temporal succession of microbial communities in plastisphere and their potentials for plastic degradation in freshwater ecosystems. Water Res. 2023, 229, 119406. [Google Scholar] [CrossRef] [PubMed]
- González-Pleiter, M.; Velázquez, D.; Casero, M.C.; Tytgat, B.; Verleyen, E.; Leganés, F.; Rosal, R.; Quesada, A.; Fernández-Piñas, F. Microbial colonizers of microplastics in an Arctic freshwater lake. Sci. Total Environ. 2021, 795, 148640. [Google Scholar] [CrossRef] [PubMed]
- Wen, B.; Liu, J.-H.; Zhang, Y.; Zhang, H.-R.; Gao, J.-Z.; Chen, Z.-Z. Community structure and functional diversity of the plastisphere in aquaculture waters: Does plastic color matter? Sci. Total Environ. 2020, 740, 140082. [Google Scholar] [CrossRef]
- Xu, L.; Li, K.; Zhang, M.; Guo, J.; Jia, W.; Bai, X.; Tian, X.; Huang, Y. Plastic substrate and residual time of microplastics in the urban river shape the composition and structure of bacterial communities in plastisphere. J. Environ. Manag. 2023, 345, 118710. [Google Scholar] [CrossRef] [PubMed]
- Delacuvellerie, A.; Ballerini, T.; Frère, L.; Matallana-Surget, S.; Dumontet, B.; Wattiez, R. From rivers to marine environments: A constantly evolving microbial community within the plastisphere. Mar. Pollut. Bull. 2022, 179, 113660. [Google Scholar] [CrossRef]
- Li, C.; Wang, L.; Ji, S.; Chang, M.; Wang, L.; Gan, Y.; Liu, J. The ecology of the plastisphere: Microbial composition, function, assembly, and network in the freshwater and seawater ecosystems. Water Res. 2021, 202, 117428. [Google Scholar] [CrossRef] [PubMed]
- Di Pippo, F.; Crognale, S.; Levantesi, C.; Vitanza, L.; Sighicelli, M.; Pietrelli, L.; Di Vito, S.; Amalfitano, S.; Rossetti, S. Plastisphere in lake waters: Microbial diversity, biofilm structure, and potential implications for freshwater ecosystems. Environ. Pollut. 2022, 310, 119876. [Google Scholar] [CrossRef] [PubMed]
- Xue, N.; Wang, L.; Li, W.; Wang, S.; Pan, X.; Zhang, D. Increased inheritance of structure and function of bacterial communities and pathogen propagation in plastisphere along a river with increasing antibiotics pollution gradient. Environ. Pollut. 2020, 265, 114641. [Google Scholar] [CrossRef] [PubMed]
- Li, L.; Huang, D.; Hu, Y.; Rudling, N.M.; Canniffe, D.P.; Wang, F.; Wang, Y. Globally distributed Myxococcota with photosynthesis gene clusters illuminate the origin and evolution of a potentially chimeric lifestyle. Nat. Commun. 2023, 14, 6450. [Google Scholar] [CrossRef] [PubMed]
- Wright, R.J.; Erni-Cassola, G.; Zadjelovic, V.; Latva, M.; Christie-Oleza, J.A. Marine Plastic Debris: A New Surface for Microbial Colonization. Environ. Sci. Technol. 2020, 54, 11657–11672. [Google Scholar] [CrossRef]
- Fernández-Álvarez, C.; Santos, Y. Identification and typing of fish pathogenic species of the genus Tenacibaculum. Appl. Microbiol. Biotechnol. 2018, 102, 9973–9989. [Google Scholar] [CrossRef]
- Koh, J.; Bairoliya, S.; Salta, M.; Cho, Z.T.; Fong, J.; Neo, M.L.; Cragg, S.; Cao, B. Sediment-driven plastisphere community assembly on plastic debris in tropical coastal and marine environments. Environ. Int. 2023, 179, 108153. [Google Scholar] [CrossRef]
- Di Pippo, F.; Venezia, C.; Sighicelli, M.; Pietrelli, L.; Di Vito, S.; Nuglio, S.; Rossetti, S. Microplastic-associated biofilms in lentic Italian ecosystems. Water Res. 2020, 187, 116429. [Google Scholar] [CrossRef]
- Wright, R.J.; Bosch, R.; Langille, M.G.I.; Gibson, M.I.; Christie-Oleza, J.A. A multi-OMIC characterisation of biodegradation and microbial community succession within the PET plastisphere. Microbiome 2021, 9, 141. [Google Scholar] [CrossRef]
- Pollet, T.; Berdjeb, L.; Garnier, C.; Durrieu, G.; Le Poupon, C.; Misson, B.; Briand, J.-F. Prokaryotic community successions and interactions in marine biofilms: The key role of Flavobacteriia. FEMS Microbiol. Ecol. 2018, 94, fiy083. [Google Scholar] [CrossRef]
- Zhang, S.-J.; Zeng, Y.-H.; Zhu, J.-M.; Cai, Z.-H.; Zhou, J. The structure and assembly mechanisms of plastisphere microbial community in natural marine environment. J. Hazard. Mater. 2022, 421, 126780. [Google Scholar] [CrossRef]
- Tu, C.; Chen, T.; Zhou, Q.; Liu, Y.; Wei, J.; Waniek, J.J.; Luo, Y. Biofilm formation and its influences on the properties of microplastics as affected by exposure time and depth in the seawater. Sci. Total Environ. 2020, 734, 139237. [Google Scholar] [CrossRef] [PubMed]
- Dussud, C.; Hudec, C.; George, M.; Fabre, P.; Higgs, P.; Bruzaud, S.; Delort, A.-M.; Eyheraguibel, B.; Meistertzheim, A.-L.; Jacquin, J.; et al. Colonization of Non-biodegradable and Biodegradable Plastics by Marine Microorganisms. Front. Microbiol. 2018, 9, 1571. [Google Scholar] [CrossRef] [PubMed]
- Amaral-Zettler, L.A.; Zettler, E.R.; Mincer, T.J. Ecology of the plastisphere. Nat. Rev. Microbiol. 2020, 18, 139–151. [Google Scholar] [CrossRef] [PubMed]
- Rohrbach, S.; Gkoutselis, G.; Hink, L.; Weig, A.R.; Obst, M.; Diekmann, A.; Ho, A.; Rambold, G.; Horn, M.A. Microplastic polymer properties as deterministic factors driving terrestrial plastisphere microbiome assembly and succession in the field. Environ. Microbiol. 2023, 25, 2681–2697. [Google Scholar] [CrossRef] [PubMed]
- Ju, Z.; Du, X.; Feng, K.; Li, S.; Gu, S.; Jin, D.; Deng, Y. The Succession of Bacterial Community Attached on Biodegradable Plastic Mulches During the Degradation in Soil. Front. Microbiol. 2021, 12, 785737. [Google Scholar] [CrossRef] [PubMed]
- Parrish, K.; Fahrenfeld, N.L. Microplastic biofilm in fresh- and wastewater as a function of microparticle type and size class. Environ. Sci. Water Res. Technol. 2019, 5, 495–505. [Google Scholar] [CrossRef]
- Debroas, D.; Mone, A.; Ter Halle, A. Plastics in the North Atlantic garbage patch: A boat-microbe for hitchhikers and plastic degraders. Sci. Total Environ. 2017, 599–600, 1222–1232. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Liu, C.; Yang, H.; He, W.; Li, B.; Zhu, X.; Liu, S.; Jia, S.; Li, R.; Tang, K.H.D. Leaching of chemicals from microplastics: A review of chemical types, leaching mechanisms and influencing factors. Sci. Total Environ. 2024, 906, 167666. [Google Scholar] [CrossRef]
- Jemec Kokalj, A.; Kuehnel, D.; Puntar, B.; Žgajnar Gotvajn, A.; Kalčikova, G. An exploratory ecotoxicity study of primary microplastics versus aged in natural waters and wastewaters. Environ. Pollut. 2019, 254, 112980. [Google Scholar] [CrossRef]
- Fernández-Juárez, V.; López-Alforja, X.; Frank-Comas, A.; Echeveste, P.; Bennasar-Figueras, A.; Ramis-Munar, G.; Gomila, R.M.; Agawin, N.S.R. “The Good, the Bad and the Double-Sword” Effects of Microplastics and Their Organic Additives in Marine Bacteria. Front. Microbiol. 2021, 11, 581118. [Google Scholar] [CrossRef] [PubMed]
- Kesy, K.; Oberbeckmann, S.; Kreikemeyer, B.; Labrenz, M. Spatial Environmental Heterogeneity Determines Young Biofilm Assemblages on Microplastics in Baltic Sea Mesocosms. Front. Microbiol. 2019, 10, 1665. [Google Scholar] [CrossRef] [PubMed]
- Arias-Andres, M.; Kettner, M.T.; Miki, T.; Grossart, H.-P. Microplastics: New substrates for heterotrophic activity contribute to altering organic matter cycles in aquatic ecosystems. Sci. Total Environ. 2018, 635, 1152–1159. [Google Scholar] [CrossRef] [PubMed]
- Tang, K.H.D. Environmental Co-existence of Microplastics and Perfluorochemicals: A Review of Their Interactions. Biointerface Res. Appl. Chem. 2023, 13, 587. [Google Scholar]
Figure 1.
The composition of the plastisphere and the factors affecting it.

Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Tang, K.H.D. Terrestrial and Aquatic Plastisphere: Formation, Characteristics, and Influencing Factors. Sustainability 2024, 16, 2163. https://0-doi-org.brum.beds.ac.uk/10.3390/su16052163
AMA Style
Tang KHD. Terrestrial and Aquatic Plastisphere: Formation, Characteristics, and Influencing Factors. Sustainability. 2024; 16(5):2163. https://0-doi-org.brum.beds.ac.uk/10.3390/su16052163
Chicago/Turabian StyleTang, Kuok Ho Daniel. 2024. "Terrestrial and Aquatic Plastisphere: Formation, Characteristics, and Influencing Factors" Sustainability 16, no. 5: 2163. https://0-doi-org.brum.beds.ac.uk/10.3390/su16052163
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.