1. Introduction
Among all of the strategies seeking for the relationships between exposure to chemicals and the effects on human health, the predictive toxicology approach has the potential to better identify biological effects from exposure to environmental mixtures and predict the final health outcome, by using in vitro methods supported by high throughput approaches and linking them to known key steps in disease progression. Alternative methods to animal testing are considered as suitable tools to support hazard identification and are also of growing interest for predicting the toxicological risks.
The simultaneous presence of a huge number of different chemicals at low concentrations could often result in misleading characterization of the hazard associated with complex mixtures, leading to underestimation of the risk, since the possible additive or more than additive interactions among chemicals could not be properly identified [
1]. Airborne particulate matter (PM) could be regarded as the prototypical example, as it is nearly impossible to identify and measure all components in the PM extracts. Moreover, individual pollutants in airborne PM samples are often under the acceptable concentration level established by the current legislations and near or even under the method detection limits. In this context, the establishment of
in vitro methods able to characterize the toxic effects and the carcinogenic potential of mixtures could be relevant for hazard and risk assessment.
Numerous scientific studies have linked exposure to environmental pollution to a variety of acute and chronic health adverse outcomes, including mortality and cardiovascular and respiratory diseases [
2]. In October, 2013, the International Agency for Research on Cancer (IARC) classified outdoor air pollution as “carcinogenic to humans” (Group 1) [
3].
The prediction of carcinogenicity is still a major issue, since cancer is a multifactorial process consisting of a sequence of stages. The
in vivo rodent bioassay is currently considered to be the reference test for the evaluation of the carcinogenicity end-point, but, for economic and technical reasons, it is rarely suitable for mixtures of environmental origin. Among
in vitro tests reproducing several stages of the multiphasic process of carcinogenesis, the cell transformation assays (CTAs) appear to be the most suitable tools to predict the carcinogenic properties of chemicals [
4,
5] and to evaluate the carcinogenic risk associated with environmental samples. The suitability of CTAs, as a model to forecast the carcinogenic potential of chemicals, was recently confirmed by the results of the prevalidation study coordinated by European center for validation of alternative methods (ECVAM) aiming at assessing the predictivity and reproducibility of
in vitro CTAs in the carcinogenicity testing of chemicals [
6,
7]. The exposure of immortalized embryonic mouse fibroblasts to chemicals having a transforming ability resulted in the loss of the contact-inhibition and the onset of morphologically-transformed foci [
8].
In this study, we evaluated the combined effect of multiple chemicals in real environmental mixtures, using alternative testing strategies, including a model to highlight the toxic and transforming properties of urban airborne samples and microarray-based transcriptomics to identify genes modulated as a consequence of the exposure to air samples collected at different sites.
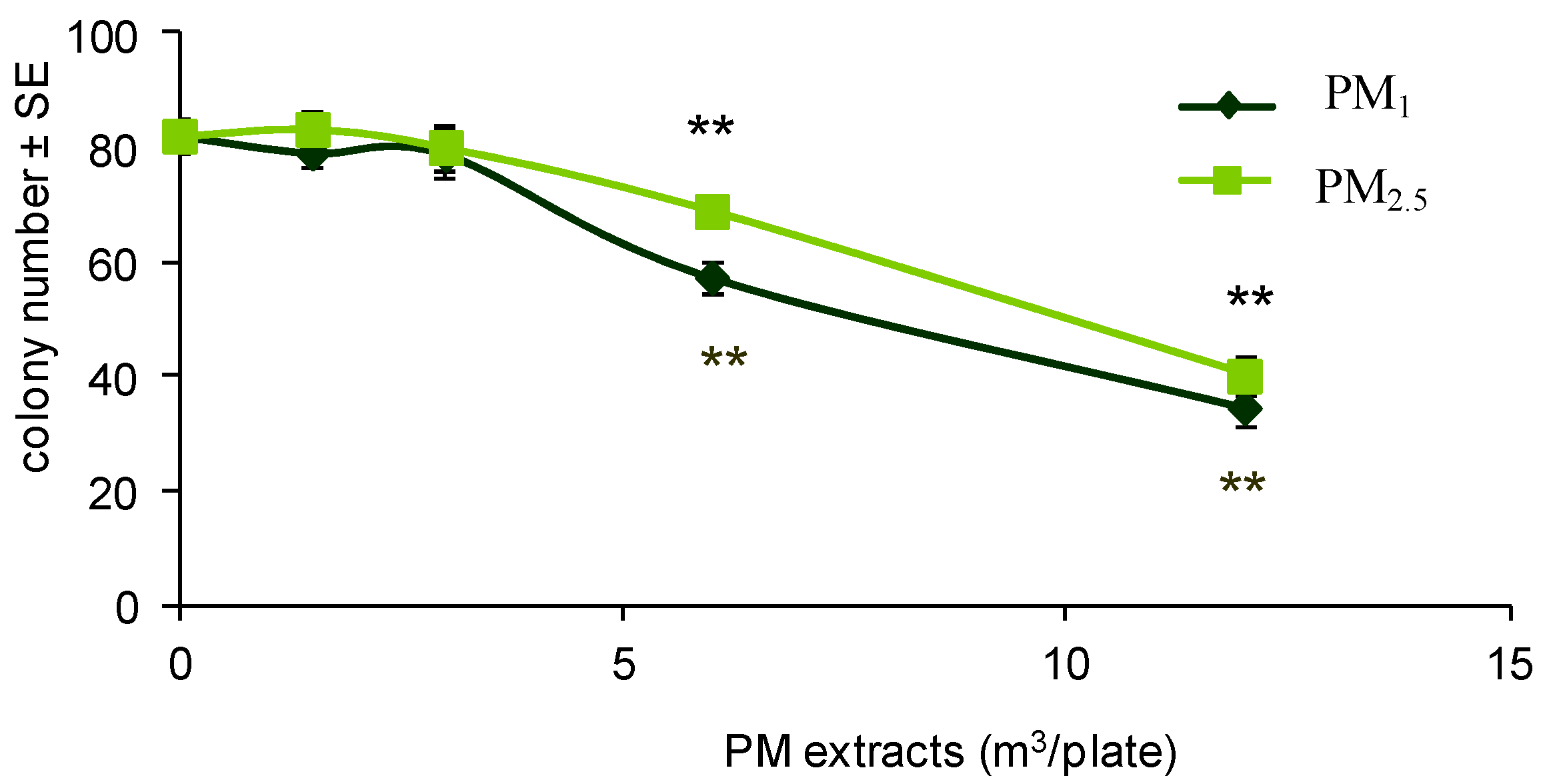
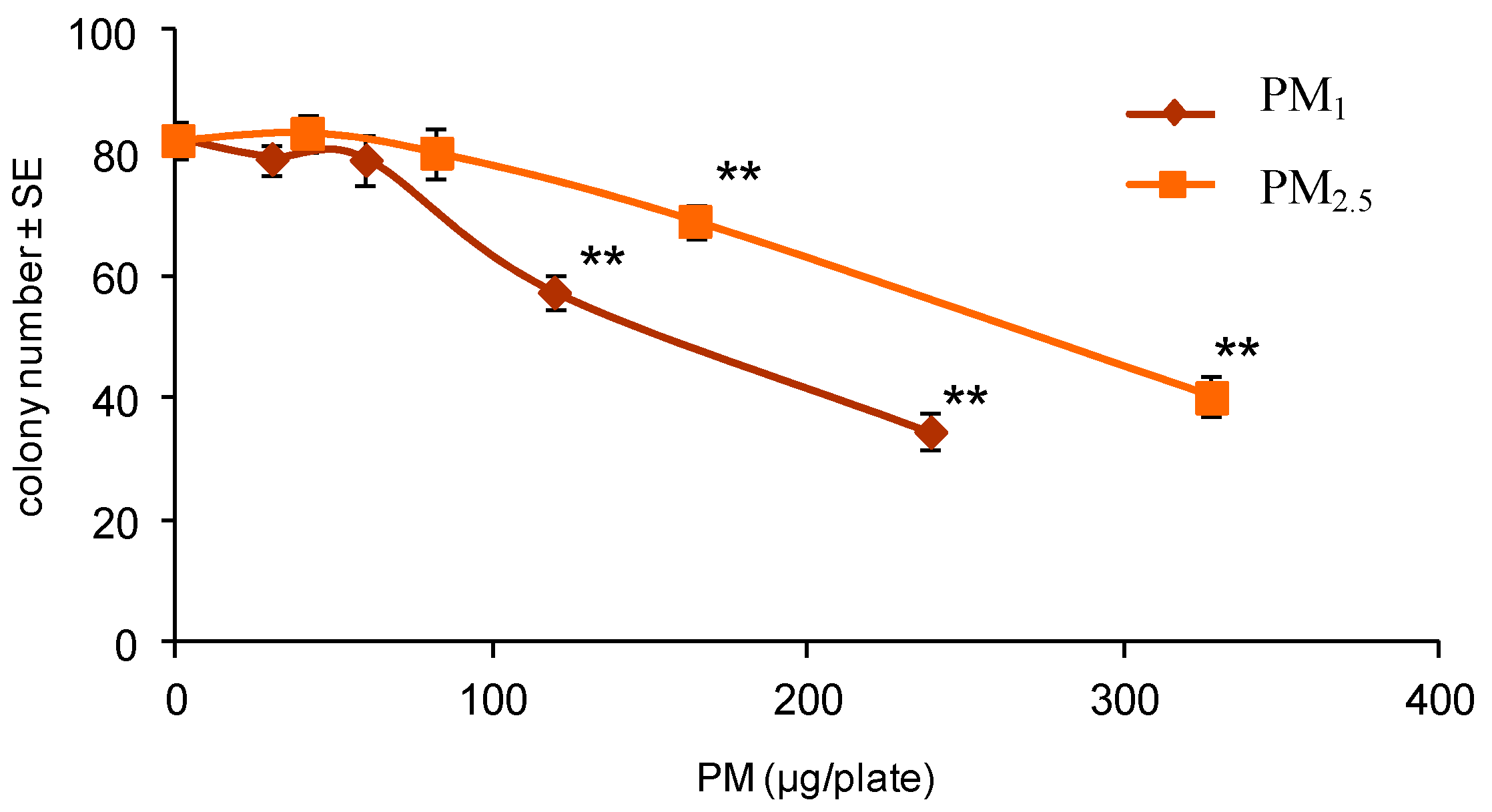
A 3T3 CTA was performed by treating cells with PM extracts. The air samples of PM2.5 and PM1 were collected during different seasons at a site that is located in the northern area of the city of Bologna. This site is considered the urban background. The organic extracts from the collected samples were evaluated for cytotoxicity and transforming activity in order to identify dose-related effects possibly associated with different aerodynamic diameters or airborne chemical components.
The integration of omics techniques with in vitro methodologies helps in clarifying the mode(s) and/or mechanism(s) of action of cell transformation following environmental exposure. The functional genomics, based on the microarray technology, could assess the impact of exposure to multiple cellular processes within a single experiment, providing information about the metabolic, signaling and regulatory networks of cells affected by mixtures. The generated signatures can be used also to derive possible biomarkers of toxicity and exposure.
It has been reported that exposures to environmental complex mixtures often affect the cell growth and several signaling pathways involved in initiating repair, allowing adaptation and promoting survival [
9], suggesting that the exposure of cells to organic PM extracts could induce the modulation of genes involved in similar pathways and processes.
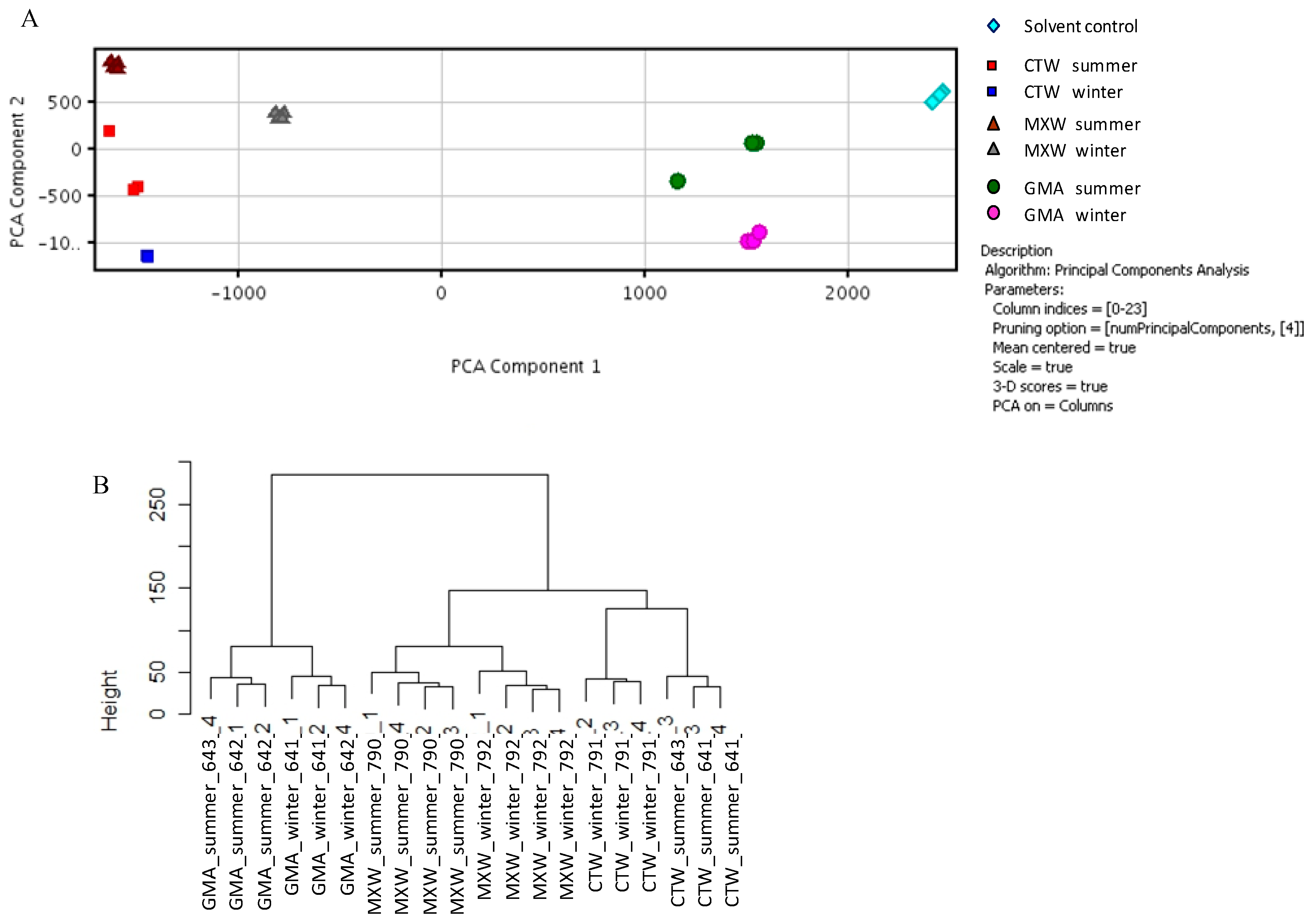
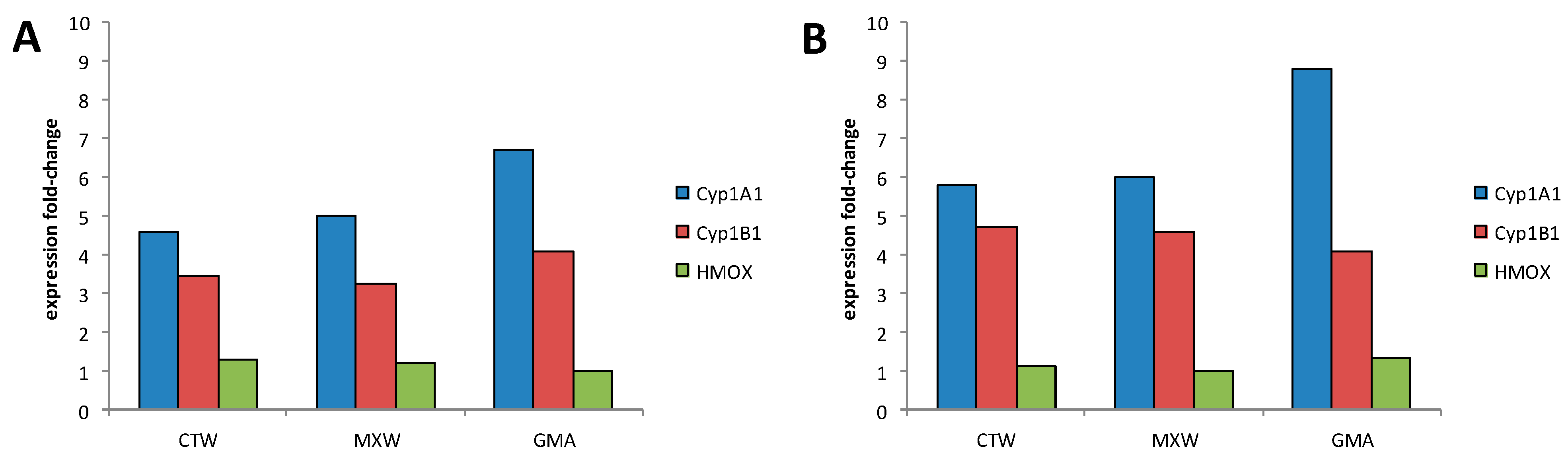
To confirm previous findings, gene expression profiles of A549 cells treated with organic extracts derived from PM2.5 collected at three sites in the surroundings of Bologna were obtained. The sampling sites were differently impacted by pollutant sources present in the sampling area.
The A549 cell line, derived from a human lung adenocarcinoma, was chosen as the cellular target for transcriptomic studies, since it has been previously confirmed to be a useful
in vitro model to study the PM effects on lung cells [
10,
11]. Indeed, A549 exhibits characteristics resembling human alveolar type II cells [
12]. The identification lists of genes affected by the treatment was used to identify the biological pathways and processes involved in the cellular response and eventually relate them to adverse health outcomes associated with PM
2.5 exposure.
2. Materials and Methods
2.1. Cells
Cell cultures were routinely maintained in a humidified incubator with an atmosphere of 5% CO2 in air at 37 °C. The BALB/c 3T3 A31-1-1 cell line was purchased from the Health Science Research Resource Bank (Osaka, Japan). The cells were grown in minimum essential medium (MEM) with 10% fetal bovine serum (FBS), cryoconserved in MEM 10% FBS solution containing 5% dimethyl sulfoxide (DMSO) and used for the CTAs at Passages 3–5 from arrival. For the transformation assays, only sub-confluent cells (about 70% confluence) were used. The target cells were not maintained in culture beyond the third passage after thawing. The A549 cells (ATCC CCL185) were used for the microarray experiments. They were obtained from LGC standards Co. and were grown in F12K medium with 10% FBS.
2.2. Air Samples
The PM
2.5 and PM
1 used for the CTAs were sampled in an urban background site during autumn, 2012 (October 23–November 12, 2012). The PM
2.5 fraction was collected by the high volume air flow sampler, Air Flow PM
2.5-HVS (UNI-EN 14907 compliant, suction flow 500 L/min, Air Monitoring Systems-Analitica, Pesaro-Italy). Air Flow polyurethane foam (PUF) (suction flow 200 L/min, Air Monitoring Systems-Analitica, Pesaro-Italy), which could collect simultaneously gas (using PUF-polyurethane foam) and PM
1, was used for PM
1 sampling. For the microarray experiments, samples of PM
2.5 were collected daily by low volume air flow samplers (Skypost TCR TECORA) during summer, 2008, (June 13–July 24, 2008) and winter, 2009 (January 15–March 12, 2009). A brief description of the sampling site is reported in
Table 1.
Table 1.
Sampling sites for toxicological analysis. PM, particulate matter.
Table 1.
Sampling sites for toxicological analysis. PM, particulate matter.
| Name | Description | Sampling Period | PM2.5 (µg/m3) | PM1 (µg/m3) | Toxicological Assay |
|---|
| Supersite | urban background site located in the northern area of the City of Bologna | Autumn | 27.27 | 19.86 | Cytotoxicity assay (CTA) |
| MXW | site located in the surroundings of Bologna and impacted by the waste-to-energy plant | Summer | 23.33 | ND | Microarray experiments |
| Winter | 35.42 | ND |
| CTW | site located in the surroundings of Bologna and impacted by all of the same sources as MXW, except the waste-to-energy plant | Summer | 21.11 | ND |
| Winter | 34.47 | ND |
| GMA | Urban background site located in the southern area of Bologna (public park) | Summer | 18.74 | ND |
| Winter | 29.56 | ND |
The gravimetric data were obtained daily by weighing the filters before and after collection. The filters were pooled to obtain seasonal samples and extracted with acetone, using a Soxhlet apparatus, then reduced to dryness and dissolved in DMSO at 800 m3 equivalents/mL. The treatment solutions were prepared by diluting the stock solutions in the culture media immediately before using. The final concentration of the vehicle DMSO was 0.5%.
2.4. Microarray Experiments
A549 cells at a density of 2 × 105 were seeded in 60-mm plates and incubated at 37 °C in a 5% CO2 humidified atmosphere. After 48 h, the cells were treated with PM2.5 extracts (8 m3 equivalents) for 4 h. At the end of the incubation time, total RNA was isolated from cells by using TRIzol Reagent (Life Technologies, Carlsbad, CA, USA) and purified on an RneasyR affinity column (Qiagen, Valencia, CA, USA). RNA quantification and quality were assessed respectively by a Nanodrop 1000 (Thermo Scientific, Wilmington, DE, USA) and an Agilent bioanalyzer 2100 using the RNA Nano kit (Agilent Technologies, Santa Clara, CA, USA).
Fluorescently-labeled cRNA was generated starting from 1 µg of total RNA by using the Low RNA Input Fluorescent Linear Amplification Kit (Agilent Technologies) according to the instructions from the supplier (Agilent Technologies). Cyanine 3-CTP (Cy3) (Perkin-Elmer, NEN Life Science, Boston, MA, USA) -labeled cRNAs were combined and purified with QIAquick spin columns (Qiagen) and then applied to the oligonucleotide slides (Whole Human Genome Microarray kit, 4 × 44 K), according to the Agilent 60-mer oligomicroarray processing protocol (G4140-90040_One-Color_GE_5.7, available online at Agilent Technologies) [
18].
Slides were scanned in the Cy-3 channel with an Agilent’s High-Resolution C Scanner (Agilent, G2565AA). Scanned images were analyzed by the Agilent Feature Extraction software 9.1 to derive the raw intensity data used in the next steps of analysis.
4. Conclusions
The evaluation of the toxicity and carcinogenicity of complex mixtures by alternative methods to animal bioassays could help in the prediction of risk linked to carcinogenic compounds present in the environmental sample.
The association between results derived from
in vitro toxicology assays, expression profiles and health outcomes needs further characterization before gene expression profiling data could be completely incorporated into the risk-assessment process. However, the global gene expression profiling performed
in vitro demonstrated that, despite the difference in the cell types, microarray platforms, incubation time and PM sources and doses, common sets of genes and pathways were modified by PM exposure, and the results were consistent with those observed in animal studies [
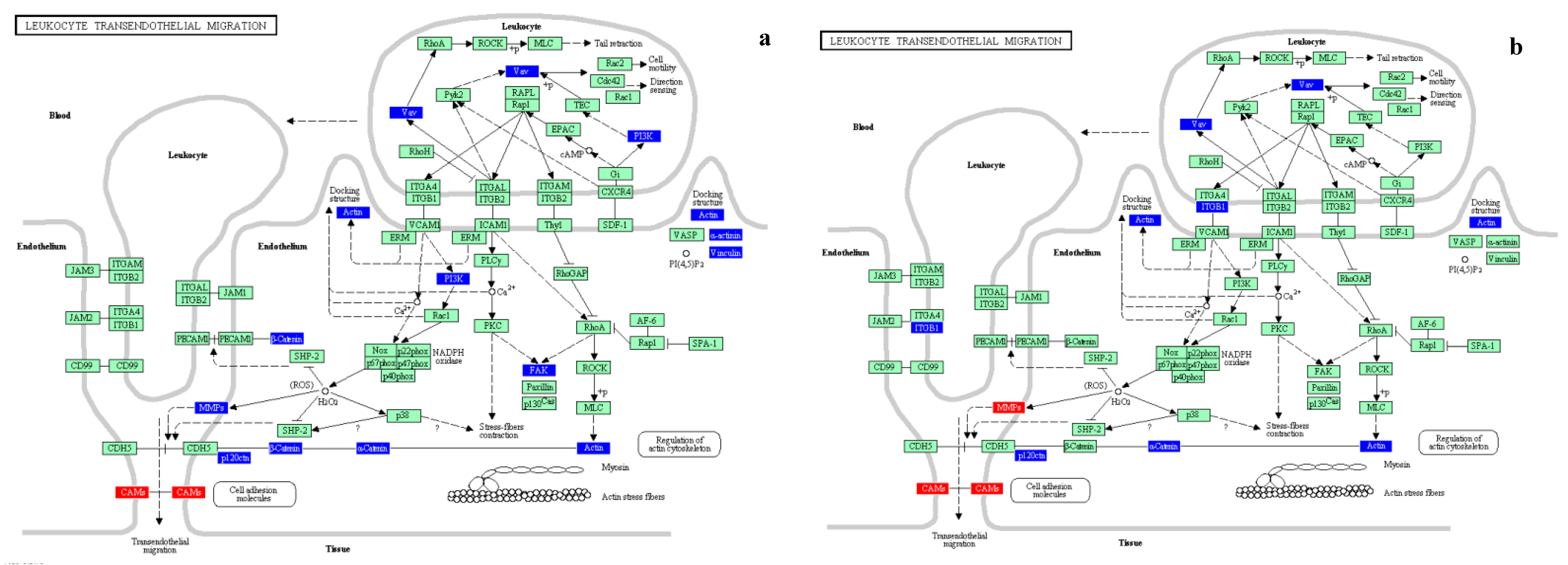
46]. Our data on biomarkers of exposure confirmed the induction of common genes related to xenobiotic metabolism and, to a lesser extent, oxidative stress. Our results demonstrated also that it is possible to differentiate environmental samples on the bases of transcriptomic profiles, stressing differences that could be possibly related to local sources of pollution. Moreover, the identification of the involvement of the adhesion process in pathways underlying the adverse health effects induced by PM could be used as supporting evidence for human exposure studies.
These findings strongly support our approach, suggesting that gene expression data coupled with in vitro assays could drive the identification of indicators of adverse health effects due to a complex mixture exposure and could be valuable in supporting the prediction of risk from environmental exposure.


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}




